



Введение
Из всех костных морфологических структур позвоночных наиболее консервативной в своем строении и в то же время информационно емкой является череп. Этот орган эволюционно формировался для выполнения сразу нескольких функций: во-первых, для защиты головного мозга, во-вторых, защиты и фиксации расположения органов чувств, в-третьих, захвата, удержания и поглощения добычи. Ввиду того, что череп сосредотачивает в себе жизненно важные функции, его строение для каждого вида позвоночных, и млекопитающих в частности, строго детерминировано образом жизни и наследственностью. В строении черепа вида (тип и строение зубной системы, размер и форма черепа) заключена информация о родственных отношениях с другими видами, специализации в пищевой сети экосистемы, условиях среды обитания. Эти закономерности давно были подмечены исследователями и в настоящее время используются для классификации диких видов (Лебедев, Лисовский, 2008; Аверьянов, Лопатин, 2011) и пород домашних животных (Боронецкая и др., 2017), а также в исследовании внутривидовых межпопуляционных различий (Васильева и др., 2008; Барышников, Пузаченко, 2009).
Основой краниологических исследований по-прежнему остается классический метод стандартного краниометрического анализа, построенный на системе линейных промеров черепа с последующим статистическим анализом полученных данных (Амшокова, 2010; Барышников, Пузаченко, 2012; Кораблёв и др., 2012, 2014). Однако в настоящее время в краниологических исследованиях все чаще прибегают к методам получения цифровых изображений черепов (Cox et al., 2013; Quintela et al., 2016) и, в частности, цифровых 3D-моделей черепов с последующим анализом их формы методами геометрической морфометрии (Maga et al., 2015). У этого подхода есть свои преимущества – он дает полную информацию о форме черепа как единой интегрированной морфоструктуры, а следовательно, увеличивает зону поиска как различий, так и инвариантных характеристик – основного предмета в исследовании форм морфологических структур биообъектов (Пузаченко, 2013, 2016).
Изменение морфологии вида, и черепа в частности, может происходить при смене условий обитания вида. Внедрение в экосистемы видов-вселенцев является наглядным примером таких процессов. Проблеме внедрения в экосистемы чужеродных видов-вселенцев в последние десятилетия уделяется достаточное внимание со стороны исследователей (Хляп и др., 2008; Олейников, 2013; Бобров, 2015; Дгебуадзе, 2014). Кардинальное внедрение нового вида в экосистему ведет к изменению ее естественного баланса и взаимосвязи между видами, нарушению устоявшейся иерархии пищевых цепочек. Таким примером может служить интродукция на территории Евразии североамериканского грызуна ондатры (Оndatra zibethicus L.), достигшая беспрецедентных масштабов в бывшем СССР, где с момента начала интродукции вида в 1928 г. по 1970 г. в пределах его территории было расселено около 300 тыс. зверьков (Ондатра…, 1993). В настоящее время ареал ондатры простирается от Европы до Дальнего Востока, захватывая территории Казахстана, Монголии и Китая (Чащухин, 2007).
Межпопуляционные различия интродуцированной ондатры изучены в Западной Сибири с применением традиционного краниометрического анализа на основе линейных промеров черепа (Васильев и др., 1999), а также методами геометрической морфометрии на материале двумерных изображений нижней челюсти (Васильев и др., 2014, 2016). Применение анализа формы нижней челюсти методами геометрической морфометрии позволило выявить более подробную картину морфогенетических изменений в популяциях интродуцированной ондатры. Однако изучение формы черепа (за исключением нижней челюсти), с учетом анализа симметричной и асимметричной компонент формы, методами геометрической морфометрии в популяциях ондатры до настоящего момента не проводилось.
В задачу настоящего исследования входило на примере выборок из контрастных географически удаленных друг от друга популяций интродуцированной ондатры (Нижегородское Поволжье и Южный Казахстан) использовать комплексный подход в исследовании краниологических показателей посредством классических методов, а также методов геометрической морфометрии с помощью анализа цифровых 3D-моделей черепов, включая анализ симметричной и асимметричной компонент формы.
Материалы
Материалом для исследований послужили выборки черепов ондатры Зоологического музея ННГУ им. Н. И. Лобачевского двух географических популяций, удаленных друг от друга более чем на 2000 км: окрестности д. Аристово Семеновского района Нижегородская области (собраны в 1949 г.) и долины р. Или Балхашского района Казахстана (собраны в 1951 г.).
Для анализа были использованы только черепа взрослых особей, возраст которых составлял 9 и более месяцев. Возраст ондатр определялся по двум признакам: особенностям формы черепа и степени развитости ламбдоидального и сагиттального гребней (Карасева и др., 2008). Признак степени развитости корней и стертости коронки для определения возраста ондатр применен не был ввиду невозможности препарирования музейных образцов.
Объем выборок, использованных в анализе, составил для нижегородской популяции 15 особей (7 самцов и 8 самок), для казахстанской – 21 особь (12 самцов, 4 самки и 5 особей без определения половой принадлежности).
Методы
Для краниометрического анализа черепов были выполнены 8 стандартных краниометрических промеров (Васильев и др., 1999), приведенных на рис. 1.
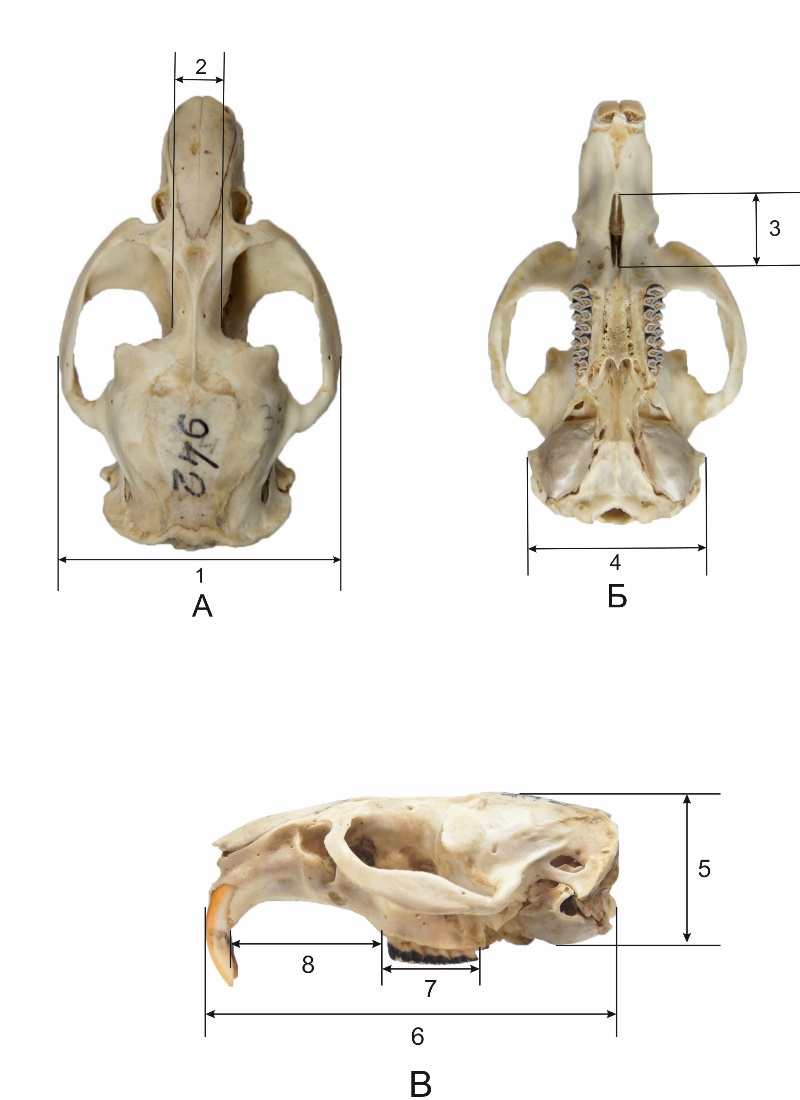
Рис. 1. Схема стандартных краниометрических промеров ондатры (Ondatra zibethicus L.). А – вид сверху: 1 – скуловая ширина (СШ), 2 – межглазничная ширина (МГШ). Б – вид снизу: 3 – длина резцового отверстия (ДРО), 4 – наибольшая ширина черепа (НШ). В – вид сбоку: 5 – высота черепа в области мозговой капсулы (ВЧ), 6 – кондилобазальная длина (КБД), 7 – альвеолярная длина верхнего зубного ряда коренных зубов (ДВЗР), 8 – длина диастемы верхней (ДДВ)
Fig. 1. Scheme of standard craniometric measurements of muskrats (Ondatra zibethicus L.). A – top view: 1 – zygomatic width, 2 – interorbital width. B – bottom view: 3 – stensen's foramen, 4 – maximum width of the skull. B – side view: 5 – the height of the skull in the area of the cerebral capsule, 6 – condylobasal length, 7 – alveolar length of the upper dentition of the molars; 8– length of the upper diastema
Трехмерные цифровые изображения черепов (3D-модели), полученные с помощью 3D-сканера Range Vision ScanCenter, использовались для анализа различий формы и оценки флуктуирующей асимметрии (ФА) методом геометрической морфометрии (Klingenberg, McIntyre, 1998). Оцифровку образцов черепов производили с помощью специализированного программного обеспечения, поставляемого в комплекте с 3D-сканером. Череп фиксировался на подложке поворотного стола, входящего в комплект сканера, таким образом, чтобы избежать изменения его положения при повороте диска стола. Череп сканировался вертикально в двух положениях с фиксацией на затылочной части и ростральной части. Для полной оцифровки поверхности черепа двумя цифровыми камерами 3D-сканера диск стола делал полный оборот в 360º. При этом съемка производилась секторами (проекциями) с шагом поворота стола на 30º. Полный поворот диска стола на 360º соответствовал 12 проекциям, составляющим 1 съемку (по 2 итерации на каждое положение). Таким образом, после совершения полного цикла оцифровки череп был представлен 12 цифровыми проекциями, которые затем объединялись в единую модель в специализированной программе-редакторе RangeVision ScanMerge. Объединение проекций в полноценную модель осуществлялось с точностью до 0.02 мм.
Для решения задач по исследованию формы черепа методами геометрической морфометрии в программе Landmark (версия 3.0.0.6) на поверхности каждой из полученных 3D-моделей черепов в характерных точках (Lalis et al., 2009; Cox et al., 2013; Ge et al., 2015; Maga et al., 2015; Quintela et al., 2016) были расставлены 65 меток (рис. 2), каждая из которых характеризовалась 3 координатами (x, y, z). Исследуемые черепа характеризовались билатеральной симметрией, при этом часть меток располагались на плоскости симметрии, а часть являлись парными. Для учета эффекта ошибки измерения каждый череп был отсканирован и оцифрован дважды.
Статистический анализ данных проводился в программной среде R (версия 3.4.2). Для анализа гендерных и межпопуляционных отличий краниометрических промеров использован двухфакторный дисперсионный анализ с факторами «популяция» и «пол». Для анализа межпопуляционных отличий по всему комплексу краниометрических промеров использован многомерный дисперсионный анализ, для визуализации результатов анализа этого типа использован метод главных компонент. Значимость действия фактора определялась непараметрически с использованием перестановочной процедуры (Good, 2005), реализованной в функциях пакета lmPerm.
Анализ формы черепа методами геометрической морфометрии проводился средствами пакета geomorph (Adams, Otárola-Castillo, 2013; Adams et al., 2017). При анализе формы использовался алгоритм с процедурой отображения парных меток и вычислением симметричной и асимметричной компонент формы (Klingenberg et al., 2002). Анализ гендерных и межпопуляционных отличий формы проводился с помощью двухфакторного прокрустова дисперсионного анализа (procrustes ANOVA) симметричной компоненты с факторами «популяция» и «пол».
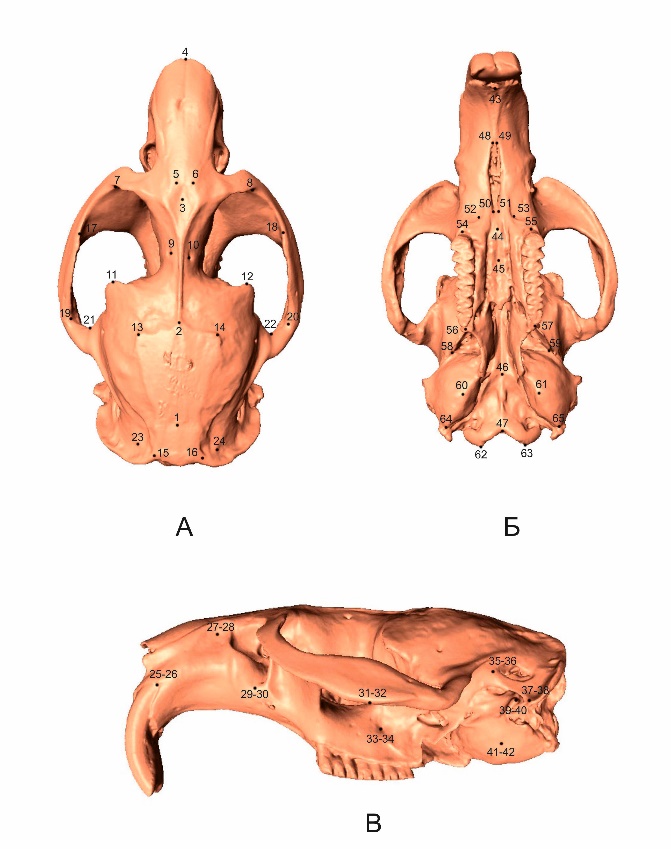
Рис. 2. Схема расстановки меток на 3D-модели черепа ондатры. А – вид сверху (1–24 метки); Б – вид снизу (43–65 метки); В – вид сбоку (25–42 метки)
Fig. 2. Scheme of arrangement of labels on 3D-models of the muskrat skull. А – top view (1–24 marks); B – bottom view (43–65 marks); B – side view (25–42 marks)
Анализ асимметрии формы черепа проводился путем двухфакторного прокрустова дисперсионного анализа, где факторами выступали «организм», «сторона» и взаимодействие «организм × сторона». Этот анализ позволяет делать количественные выводы о присутствии статистически значимой направленной и флуктуирующей асимметрии (Klingenberg, McIntyre, 1998). В этом виде анализа значимость эффекта фактора «организм» означает наличие внутрипопуляционной изменчивости признака (выявляется практически всегда), значимость эффекта фактора «сторона» означает наличие направленной асимметрии, а значимость эффекта взаимодействия «организм × сторона» указывает на присутствие флуктуирующей асимметрии.
Результаты
- Традиционный краниометрический анализ различий нижегородской и казахстанской популяций ондатр
В табл. 1 представлены средние и стандартные отклонения 8 краниометрических промеров для нижегородской и казахстанской популяций ондатры. Двухфакторный дисперсионный анализ не выявил статистически значимого влияния пола ни по одному из признаков (см. столбец pgender). Межпопуляционные отличия выявлены только по длине резцового отверстия (см. столбец psite), которая оказалась выше в казахстанской популяции ондатры.
Таблица 1. Традиционные краниометрические показатели и результаты анализа межполовых и межпопуляционных отличий в выборках ондатры нижегородской и казахстанской популяций
| Признаки | Описательная статистика | Результаты двухфакторного дисперсионного анализа | ||||
| нижегородская популяция (n = 15) |
казахстанская популяция (n = 21) | |||||
| m | s | m | s | pgender | psite | |
| Кондилобазальная длина | 63.2180 | 2.7368 | 64.3162 | 1.8305 | 1.0000 | 0.4508 |
| Скуловая ширина | 38.8907 | 1.9691 | 38.8462 | 1.4714 | 0.2508 | 0.7255 |
| Межглазничная ширина | 6.2247 | 0.5537 | 6.0810 | 0.3660 | 0.7451 | 0.6333 |
| Высота черепа в области мозговой капсулы | 21.4780 | 0.8581 | 21.5886 | 0.8387 | 0.2714 | 0.7843 |
| Длина резцового отверстия | 12.7560 | 1.0407 | 13.5686 | 1.0161 | 0.7647 | 0.0471 |
| Диастема верхняя, длина | 23.2560 | 1.4065 | 23.6129 | 1.0763 | 0.8039 | 0.5341 |
| Альвеолярная длина | 15.8907 | 0.7782 | 15.9943 | 0.7235 | 0.6667 | 0.9804 |
| Наибольшая ширина черепа | 27.1340 | 1.3004 | 27.1086 | 0.9149 | 0.3875 | 0.5402 |
Примечание. n – объем выборки; m – среднее; s – стандартное отклонение; pgender – значимость межполовых различий; psite – значимость межпопуляционных различий.
Отсутствие внутрипопуляционных гендерных различий по краниометрическим промерам позволило на этапе многомерного анализа межпопуляционных различий объединить самцов и самок в единую выборку. Многомерный дисперсионный анализ (MANOVA) значимых межпопуляционных отличий между выборками ондатры по совокупности 8 краниометрических показателей не выявил (λ Уилкса = 0.77; F = 1; p = 0.43). В целях визуализации результатов многомерного анализа проведен анализ главных компонент. Диаграмма рассеяния в пространстве двух первых главных компонент представлена на рис. 3. Хорошо видно отсутствие систематического смещения одной группы точек относительной другой, эллипсы рассеяния практически полностью пересекаются.
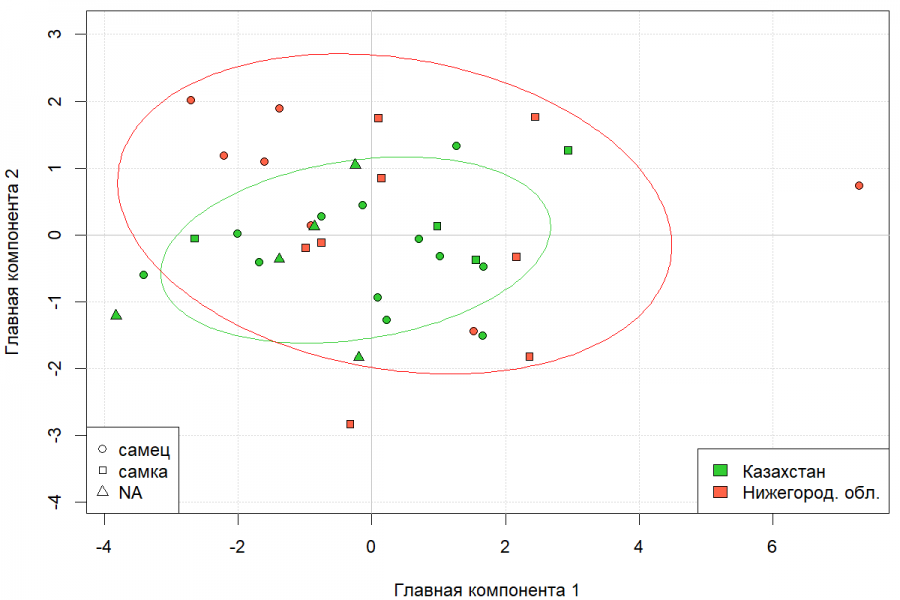
Рис. 3. Диаграмма рассеяния особей казахстанской и нижегородской популяций ондатры в пространстве первых двух главных компонент, вычисленных по 8 краниометрическим признакам (1-я главная компонента – 54.53 % общей дисперсии, 2-я главная компонента – 16.62 % общей дисперсии)
Fig. 3. Diagram of dispersion of individuals from Kazakhstan and Nizhny Novgorod muskrat populations in the space of the first two main components, calculated according to 8 craniometric features (the 1st main component – 54.53 % of the total dispersion, the 2nd main component – 16.62 % of the total dispersion)
Таким образом, традиционный краниометрический анализ с использованием совокупности 8 стандартных промеров черепа ондатры не выявил статистически значимых различий между выборками географически удаленных популяций как на внутрипопуляционном, так и на межпопуляционном уровне. Статистически значимые межпопуляционные отличия выявлены только по длине резцового отверстия.
2. Анализ различий формы черепов нижегородской и казахстанской популяций ондатры
Сравнительный анализ формы черепов популяций ондатры проводился с помощью методов геометрической морфометрии. Вначале была проведена процедура отображения координат парных меток относительно плоскости симметрии. Затем был проведен обобщенный прокрустов анализ, по результатам которого были вычислены переменные формы (координаты упорядоченных конфигураций меток без учета размерной составляющей) для исходных и отображенных конфигураций меток. На третьем этапе были рассчитаны симметричная и асимметричная компоненты формы (Klingenberg et al., 2002).
Симметричные компоненты формы использованы в двухфакторном прокрустовом дисперсионном анализе межполовых и межпопуляционных отличий. Результаты представлены в табл. 2. Межполовых отличий по форме черепа не выявлено. Отличия же между популяциями статистически значимы. Фактор межпопуляционных отличий объясняет 7.63 % общей изменчивости формы.
Таблица 2. Результаты прокрустова дисперсионного анализа межполовых и межпопуляционных различий симметричной компоненты формы черепа выборок ондатры нижегородской и казахстанской популяций
Фактор |
Статистические показатели |
|||||
| df | SS | MS | F | η2 | р | |
| Популяция | 1 | 0.00350 | 0.00350 | 2.3863 | 0.07631 | < 0.001 |
| Пол | 1 | 0.00158 | 0.00158 | 1.0765 | 0.03443 | 0.2340 |
| Популяция × пол | 1 | 0.00118 | 0.00118 | 0.8065 | 0.02578 | 0.6101 |
| Остаточная изменчивость | 27 | 0.03965 | 0.00147 | |||
Примечание. df – число степеней свободы; SS – сумма квадратов; MS – средний квадрат; F – отношение средних квадратов; η2 – доля объясненной дисперсии.
Для наглядной демонстрации обнаруженных различий по форме черепа на рис. 4 представлена диаграмма рассеяния значений первых двух главных компонент, вычисленных по переменным формы (координаты x, y, z 65 меток черепа в упорядоченных и масштабированных конфигурациях). Хорошо видно, что нижегородская и казахстанская выборки расходятся по 1 главной компоненте.
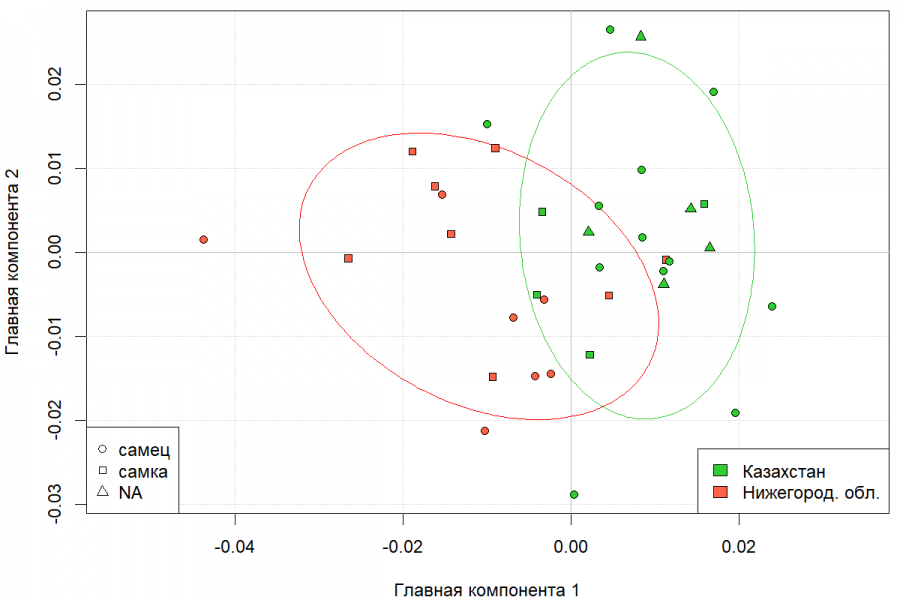
Рис. 4. Диаграмма рассеяния особей казахстанской и нижегородской популяций ондатры в пространстве первых двух главных компонент, вычисленных по симметричной компоненте формы черепа
Fig. 4 Diagram of dispersion of individuals from the Kazakhstan and Nizhny Novgorod muskrat populations in the space of the first two main components, calculated from the symmetric component of the skull shape
3. Оценка межпопуляционных различий уровня асимметрии формы черепов нижегородской и казахстанской популяций ондатры методами геометрической морфометрии
Двухфакторный дисперсионный прокрустов анализ формы черепа обнаружил статистически значимые различия как по фактору «сторона», выявив наличие направленной асимметрии, так и по фактору «организм × сторона», подтвердив наличие ФА (табл. 3).
Таблица 3. Результаты прокрустова дисперсионного анализа асимметрии формы черепа ондатры нижегородской
и казахстанской популяций
| Фактор | Статистические показатели |
|||||
| df | SS | MS | F | η2 | р | |
| Организм | 35 | 0.20863 | 0.00596 | 6.8903 | 0.67909 | < 0.001 |
| Сторона | 1 | 0.00288 | 0.00288 | 3.3283 | 0.00937 | < 0.001 |
| Организм × сторона | 35 | 0.03028 | 0.00087 | 0.9520 | 0.09856 | < 0.001 |
| Остаточная изменчивость | 72 | 0.06543 | 0.00091 | |||
Примечание. Обозначения соответствуют табл. 2.
Анализ показал, что 67.9 % изменчивости формы приходится на отличия между организмами (см. показатель η2). Отличия между сторонами (направленная асимметрия) объясняет 0.9 % изменчивости, тогда как на долю флуктуирующей асимметрии приходится 9.8 %. Оставшиеся 21.3 % составляет случайная необъясненная изменчивость. Таким образом, флуктуирующая асимметрия по величине примерно в 10 раз выше направленной.
Представленный анализ не учитывает возможность различного проявления направленной асимметрии в нижегородской и казахстанской популяциях ондатры. Для выявления этого аспекта был проведен отдельный прокрустов дисперсионный анализ на основе асимметричной компоненты формы черепа ондатры. Результаты выявили статистически значимые межпопуляционные отличия по асимметричной компоненте формы (F = 1.48, p = 0.029, η2 = 0.042). Это означает, что две популяции характеризуются разной направленной асимметрией формы черепа.
Индивидуальный показатель флуктуирующей асимметрии рассчитывался для каждой особи как квадратный корень из суммы квадратов разностей координат между исходной и отображенной конфигурациями меток за вычетом среднего популяционного уровня асимметрии по каждой координате. Межпопуляционные отличия в уровне флуктуирующей асимметрии анализировались путем перестановочной процедуры на основе однофакторного дисперсионного анализа. Отличия не выявлены (p = 0.53).
Обсуждение
В настоящем исследовании реализован комплексный подход к краниометрическому анализу двух популяций ондатры на ранних этапах ее интродукции на территории Нижегородской области и Казахстана (1949 и 1951 г. соответственно). Анализ традиционных краниометрических промеров дополнен результатами анализа симметричной и асимметричной компонент формы черепа методами геометрической морфометрии на основе 3D-моделей.
Межполовые отличия не были выявлены как по промерам, так и методами геометрической морфометрии. Ранее межполовые отличия по краниометрическим промерам были проанализированы для популяций ондатры, обитающих на полуострове Ямал и на территории Курганской области (Васильев и др., 1999). Для курганских популяций отличия были выявлены, для ямальских – нет. Соответственно отсутствие полового диморфизма в изученных нами популяциях не противоречит ранее полученным данным.
Межпопуляционные отличия между нижегородской и казахстанской популяциями ондатры выявлены только для одного краниометрического промера – длины резцового отверстия. По всей совокупности 8 промеров межпопуляционные отличия не выявлены. Напротив, геометрическая морфометрия выявила статистически значимые отличия по симметричной компоненте формы черепа между популяциями. Полученный результат свидетельствует об адаптационных процессах приспособления к различным условиям обитания даже на ранних этапах акклиматизации. Полученный результат соответствует ранее проведенному анализу отличий между курганской и ямальской популяциями ондатры на ранних этапах акклиматизации на основе краниометрических промеров (Васильев и др., 1999) и на основе геометрической морфометрии нижней челюсти (Васильев и др., 2014, 2016). Оба метода выявили межпопуляционные отличия между географически удаленными популяциями ондатры. Наше исследование дополняет указанные исследования в части демонстрации межпопуляционной изменчивости формы черепа.
Нами также впервые была проанализирована межпопуляционная изменчивость асимметричной компоненты черепа ондатры. Показано, что и нижегородская, и казахстанская популяции ондатры характеризуются наличием направленной асимметрией формы черепа. При этом выявлены статистически значимые отличия между популяциями по направленной асимметрии. Это означает, что систематическое искривление черепа, так же как и форма черепа, обладает межпопуляционной спецификой. На данном этапе исследований невозможно выяснить, имеет ли эта специфика адаптивное значение. С учетом того, что межпопуляционные отличия объясняют только 4.2 % межиндивидуальной изменчивости асимметричной компоненты формы, мы склоняемся к тому, что эти отличия являются результатом изоляции и генетико-автоматических процессов.
Уровень флуктуирующей асимметрии часто используется в популяционных исследованиях в качестве меры стабильности развития, что, в свою очередь, может отражать степень «благополучия» популяций (Захаров, 2001; Захаров и др., 2001). Так как отличий между популяциями по уровню флуктуирующей асимметрии в нашем анализе не выявлено, можно предположить отсутствие существенных изменений окружающей среды в момент развития организмов. Также следует учитывать, что череп является функционально важной структурой, развитие которой высоко канализовано и устойчиво к ненаправленным случайным отклонениям в ходе онтогенеза.
Заключение
Несмотря на широкое внедрение молекулярно-генетических методов, методов биологии развития и других исследовательских технологий, основной объем информации о разнообразии организмов по-прежнему добывается в процессе количественных морфологических исследований (Klingenberg et al., 2002). В последние годы исследования варьирования размеров и формы организмов проводятся с широким использованием аппарата статистического многомерного анализа в рамках направления, получившего название «морфометрия» (Павлинов, 2008; Поздняков, 2010; Klingenberg, 1996, 2008; Badyaev et al., 2000; Stevens, Willing, 2000; Zelditch et al., 2004). В общем случае многомерный морфометрический анализ направлен на обнаружение порядка (правил, закономерностей) в варьировании множества морфологических измерений на выборках организмов и отображение положения признаков («промеров») или особей в моделируемом многомерном пространстве, обозначаемом как «морфологическое пространство». В целом методы морфометрии показали свою эффективность в решении задач таксономии, индивидуального развития, эволюции и др.
Можно согласиться с мнением Пузаченко (2013, 2016), что варьирование микро- и макропараметров морфологического разнообразия черепа взрослых животных ограничено таким образом, что большая часть значений попадает в относительно узкий интервал или «норму», верхняя граница которой отражает организованность морфологического разнообразия, инвариантную для черепа млекопитающих. Классический метод стандартного краниометрического анализа, основанный на системе линейных промеров черепа, характеризует именно эту консервативную (инвариантную) составляющую часть видоспецифичности. Подчеркнем, что, по Пузаченко (2013), одним из показателей, позволяющим отнести макропараметр к группе инвариантных показателей, является отсутствие полового диморфизма, что и наблюдалось в краниометрических показателях сравниваемых популяций ондатры.
Мы полагаем, что использование 3D-моделей черепов открывает перспективу расширения и углубления морфометрических исследований, в том числе и с использованием аппарата теории групп, а предпосылкой к развитию этого направления является проведенный нами теоретико-групповой анализ псевдосимметрии двумерных изображений гидробионтов классов Conjugatophyceae, Bacillariophyceae, Acantharia, Asteroidea, а также преобразований симметрии в онтогенезе иглокожих (Гелашвили и др., 2016, 2018). Следующим логическим шагом в изучении морфометрии биообъектов, в том числе черепов млекопитающих, включая их псевдосимметрию, будет теоретико-групповой анализ их трехмерных изображений.
Библиография
Аверьянов А. О., Лопатин А. В. Филогения трикодонтов и симметродонтов и происхождение современных млекопитающих // Доклады Академии наук. 2011. Т. 436. № 2. С. 276–279.
Амшокова А. Х. Изменчивость краниометрических признаков малой лесной мыши (Sylvaemus uralensis Pall.) на разных уровнях в условиях Центрального Кавказа // Вестник Нижегородского университета им. Н. И. Лобачевского. 2010. № 3-1. С. 126–133.
Барышников Г. Ф., Пузаченко А. Ю. Краниометрическое разнообразие островных популяций бурого медведя (Ursus arctos, Carnivora) с Хоккайдо, Сахалина и Южных Курил // Труды Зоологического института РАН. 2009. Т. 313. № 2. С. 119–142.
Барышников Г. Ф., Пузаченко А. Ю. Краниометрическая изменчивость речной выдры (Lutra lutra: Carnivora: Mustelidae) в Северной Евразии // Труды Зоологического института РАН. 2012. Т. 316. № 3. С. 203–222.
Бобров В. В. Виды-вселенцы млекопитающих в России (обзор исследований) // Социально-экологические технологии. 2015. № 1-2. С. 21–30.
Боронецкая О. И., Никифоров А. И., Чикурова Е. А. Государственный музей животноводства имени Е. Ф. Лискуна – центр краниологических исследований // Известия Тимирязевской сельскохозяйственной академии. 2017. № 3. С. 70–83.
Васильев А. Г., Большаков В. Н., Малафеев Ю. М., Валяева Е. А. Эволюционно-экологические процессы в популяциях ондатры при акклиматизации в условиях севера // Экология. 1999. № 6. С. 433–441.
Васильев А. Г., Большаков В. Н., Синева Н. В. Отдаленные морфогенетические последствия акклиматизации ондатры в Западной Сибири // Доклады Академии наук. 2014. Т. 455. № 4. С. 478–480.
Васильев А. Г., Большаков В. Н., Васильева И. А., Синева Н. В. Последствия интродукции ондатры в Западной Сибири: морфофункциональный аспект // Российский журнал биологических инвазий. 2016. № 4. С. 2–13.
Васильева И. А., Васильев А. Г., Большаков В. Н. Морфологическая дивергенция скальных полевок подрода Aschizomys (Rodentia, Cricetidae) // Зоологические исследования / Под ред. И. Я. Павлинова, М. В. Калякина. М.: Изд-во Моск. ун-та, 2008. С. 210–255.
Гелашвили Д. Б., Чупрунов Е. В., Сомов Н. В., Марычев М. О., Нижегородцев А. А., Маркелов И. Н., Якимов В. Н. Псевдосимметрия в живой природе: Монография / Под общ. ред. проф. Д. Б. Гелашвили и проф. Е. В. Чупрунова. Н. Новгород: Изд-во ННГУ, 2016. 363 с.
Гелашвили Д. Б., Чупрунов Е. В., Сомов Н. В., Марычев М. О., Нижегородцев А. А., Маркелов И. Н., Якимов В. Н. Теоретико-групповой анализ преобразований симметрии на примере некоторых гидробионтов // Журнал общей биологии. 2018. Т. 79. № 3. С. 169–182.
Дгебуадзе Ю. Ю. Чужеродные виды в голарктике: некоторые результаты и перспективы исследований // Российский журнал биологических инвазий. 2014. № 1. С. 2–8.
Захаров В. М. Онтогенез и популяция (стабильность развития и популяционная изменчивость) // Экология. 2001. № 3. С. 177–191.
Захаров В. М., Жданова Н. П., Кирик Е. Ф., Шкиль Ф. Н. Онтогенез и популяция: оценка стабильности развития в природных популяциях // Онтогенез. 2001. Т. 32. № 6. С. 404–421.
Карасева Е. В., Телицына А. Ю., Жигальский О. А. Методы изучения грызунов в полевых условиях . М.: Изд-во ЛКИ, 2008. 416 с.
Кораблёв М. П., Кораблёв Н. П., Кораблёв П. Н. Морфо-фенетический анализ популяций американской норки (Neovison vison) Каспийско-Балтийского водораздела // Российский журнал биологических инвазий. 2012. № 4. С. 36–56.
Кораблёв Н. П., Кораблёв М. П., Кораблёв П. Н., Туманов И. Л. Факторы морфологического разнообразия краниометрических признаков американской норки (Neovison vison) // Российский журнал биологических инвазий. 2014. № 4. С. 30–54.
Лебедев В. С., Лисовский А. А. Географическая изменчивость метрических признаков черепа и таксономическая структура хомячков Cricetulus группы Barabensis (Rodentia, Cricetidae) // Зоологический журнал. 2008. Т. 87. № 3. С. 361–374.
Олейников А. Ю. Размещение аборигенных и интродуцированных полуводных млекопитающих на Сихотэ-Алине // Российский журнал биологических инвазий. 2013. № 2. С. 35–50.
Ондатра: морфология, систематика, экология . М.: Наука, 1993. 542 с.
Павлинов И. Я., Нанова О. Г., Спасская Н. Н. К изучению морфологического разнообразия размерных признаков черепа млекопитающих. 1. Соотношение разных форм групповой изменчивости // Журнал общей биологии. 2008. Т. 69. № 5. С. 344–354.
Поздняков А. А. Морфологическая изменчивость животных (методы, результаты изучения на примере разных таксонов) // Сообщества и популяции животных: экологический и морфологический анализ. Новосибирск; М.: Товарищество научных изданий КМК, 2010. С. 133–157.
Пузаченко А. Ю. Инварианты и динамика морфологического разнообразия (на примере черепа млекопитающих) : Автореф. дис. … д-ра биол. наук. М., 2013. 48 с.
Пузаченко А. Ю. Количественные закономерности морфологического разнообразия черепа млекопитающих // Сборник трудов Зоологического музея МГУ им. М. В. Ломоносова. 2016. Т. 54. С. 229–268.
Хляп Л. А., Бобров В. В., Варшавский А. А. Биологические инвазии на территории России: млекопитающие // Российский журнал биологических инвазий. 2008. № 2. С. 78–96.
Чащухин В. А. Ондатра: причины и следствия биологической инвазии . М.: Т-во научных изданий КМК, 2007. 133 с.
Adams D. C., Otárola-Castillo E. Geomorph: an R package for the collection and analysis of geometric morphometric shape data // Methods in Ecology and Evolution. 2013. Vol. 4. Issue 4. P. 393–399. DOI: 10.1111/2041-210X.12035
Adams D. C., Collyer M. L., Kaliontzopoulou A., Sherratt E. Geomorph: Software for geometric morphometric analyses. R package version 0.5. 2017. P. 1–139. URL: https://cran.r-project.org/package=geomorph.
Badyaev A. V., Foresman K. R. Extreme environmental change and evolution: stress-induced morphological variation is strongly concordant with patterns of evolutionary divergence in shrew mandibles // Proc. R. Soc. Lond. 2000. Issue 267. P. 371-377.
Cox P. G., Kirkham J., Herre A. Masticatory biomechanics of the Laotian rock rat, Laonastes aenigmamus, and the function of the zygomaticomandibularis muscle // PeerJ. 1: e160. 2013. DOI: 10.7717/peerj.160
Ge D., Yao L., Xia L., Zhang Z., Yang Q. Geometric morphometric analysis of skull morphology reveals loss of phylogenetic signal at the generic level in extant lagomorphs (Mammalia: Lagomorpha) // Contributions to Zoology. 2015. Vol. 84 (4). P. 267–284.
Good P. Permutation, parametric and bootstrap tests of hypotheses. N.Y.: Springer, 2005. 315 p.
Klingenberg C. P. A combined morphometric and phylogenetic analysis of an ecomorphological trend: pelagization in Antarctic fishes (Perciformes: Nototheniidae) // Biological Journal of the Linnean Society. 1996. Issue 59. P. 143–177.
Klingenberg C. P. Morphological Integration and developmental modularity // Annu. Rev. Ecol. Evol. Syst. 2008. Issue 39. P. 115–132.
Klingenberg C. P., McIntyre G. S. Geometric morphometrics of developmental instability: Analyzing patterns of fluctuating asymmetry with procrustes methods // Evolution. 1998. Vol. 52. Issue 5. P. 1363–1375.
Klingenberg C. P., Barluenga M., Meyer A. Shape analysis of symmetric structures: Quantifying variation among individuals and asymmetry // Evolution. 2002. Vol. 56. Issue 10. P. 1909–1920.
Lalis A., Evin A., Denys C. Morphological identification of sibling species: the case of West African Mastomys (Rodentia: Muridae) in sympatry // C. R. Biologies. 2009. Vol. 332. P. 480–488.
Maga A. M., Navarro N., Cunningham M. L., Cox T. C. Quantitative trait loci affecting the 3D skull shape and size in mouse and prioritization of candidate genes in-silico // Frontiers in Physiology. 2015. Vol. 6. Article 92. DOI: 10.3389/fphys.2015.00092
Stevens R. D., Willing M. R. Community structure, abundance, and morphology // OIKOS. 2000. Vol. 88. P. 48–56.
Quintela F. M., Fornel R., Freitas T. R. O. Geographic variation in skull shape of the water rat Scapteromys tumidus (Cricetidae, Sigmodontinae): isolation-by-distance plus environmental and geographic barrier effects? // Anais da Academia Brasileira de Ciê. 2016. Vol. 88 (1 Suppl.). P. 451–466. DOI: 10.1590/0001-3765201620140631
Zelditch M. L., Swiderski D. L., Sheets H. D., & Fink W. L. Introduction // Geometric Morphometrics for Biologists. 2004. P. 1–20.

 © 2011 - 2026
© 2011 - 2026