



Введение
Фрагментация – расчленение обширных местообитаний на изолированные участки, которые слишком ограничены для того, чтобы поддерживать характерный видовой состав долгое время, представляет собой одну из главных причин снижения общего биоразнообразия (Meffe, 1997; Лосев, 2006 и др.). Лишайники, являясь чувствительными организмами к антропогенному нарушению, обладают большим потенциалом в изучении влияния фрагментации на экосистемы (Esseen, Renhorn, 1998). Исследования по этой теме начались сравнительно недавно – с конца прошлого века. За это время опубликовано несколько десятков работ, выполненных в бореальных, реже в тропических и субтропических лесах в Европе, Северной и Южной Америке. Часть исследований посвящена изучению влияния фрагментации местообитаний на общее разнообразие лишайников (Svoboda et al., 2010 и др.); выделяются потенциально уязвимые к фрагментации виды лишайников (Dettki et al., 1998). Другое направление в этой области образуют исследования, касающиеся изучения так называемого краевого эффекта (edge effect), складывающегося на окраинах лесных растительных сообществ, граничащих с открытыми пространствами, в том числе вырубками (Rheault et al., 2003; Gignac, Dale, 2005; Boudreault et al., 2008; Hedenеs, Ericson, 2008 и др.). В ряде работ оцениваются возможности распространения отдельных редких видов лишайников в ландшафтах, подверженных высокой степени фрагментации лесной растительности (Esseen, Renhorn, 1998; Keon, Muir, 2002; Esseen, 2006; Coxson, Stevenson, 2007, и др.), в том числе и с применением методов молекулярной биологии (Otólora et al., 2011). Однако выявленных закономерностей все еще не достаточно для детальной оценки влияния на лишайники такого сложного явления, как фрагментация местообитаний. И это связано, в первую очередь, с разнообразием специфических экологических условий, складывающихся в разных сообществах, различиями в биологии и экологии видов, разными жизненными стратегиями отдельных лишайников, а также с технической трудностью изучения и сложностью интерпретации полученных результатов. Вопрос о размере фрагмента, необходимого для поддержания качественных и количественных характеристик лишайникового покрова, пока остается открытым. Имеющаяся в литературе информация, в основном касается лесных естественных территорий, в которых влияние загрязнения не значительно и не учитывается. Вместе с тем в городских условиях следует ожидать более выраженного эффекта воздействия фрагментации местообитаний на такие организмы как лишайники, являющиеся высокочувствительными к загрязнению воздушной среды (Бязров, 2002). Цель исследования - выявление закономерностей формирования лишайникового покрова стволов сосны под влиянием фрагментации местообитаний в сосновых сообществах на территории г. Костомукши.Материалы
Город Костомукша построен на северо-западе Республики Карелия в 1983 г., площадь 4046 км2, население около 30 тыс. человек (по данным 2010 г.). Возникновение города связано со строительством Костомукшского горно-обогатительного комбината (КГОКа). Растительность района относится к подзоне северной тайги с преобладанием сосновых лесов (Волков, 2008). Город относительно комбината расположен в противоположной стороне действия преобладающих ветров; жилищное строительство и проведение коммуникаций – с учетом максимального сохранения участков леса.
На территории города повсеместно встречаются фрагменты естественных насаждений – сосняков зеленомошных с разным уровнем антропогенного нарушения (рис. 1). Основными видами нарушений являются рекреация (вытаптывание, кострища, механическое повреждение деревьев, выгул домашних животных) и загрязнение. Согласно имеющимся данным, в 2009 г. Костомукшским городским округом (в том числе и комбинатом) было выброшено в атмосферу 45.6 тыс. т загрязняющих веществ, что составляет 41 % всех выбросов в Карелии. В состав основных выбросов комбината входят твердые вещества – 11 %, диоксид серы – 82 %, оксид углерода – 3.1 % и оксиды азота – 3.3 %. При этом вклад автотранспорта в суммарные выбросы города составляет 9.6 %, из них оксида углерода – 69.2 %, оксидов азота – 41.5 % (Государственный доклад…, 2010).
Рис. 1. Местоположение исследованных фрагментов сосновых сообществ в г. Костомукша.
Fig. 1. Location of the investigated fragments of pine forest in Kostomuksha town.
Методы
Для каждого изученного фрагмента была определена его площадь (га) при помощи компьютерной программы MapInfo. В пределах каждого фрагмента случайным образом отбирались по 8 деревьев сосны обыкновенной (Pinus sylvestris L.), соответствующих следующим параметрам: прямостоячие, внешне не поврежденные, возрастом более 100 лет, высотой 19-22 м, диаметром ствола 22-28 см, радиусом кроны 1.3–2.1 м. Описания эпифитного лишайникового покрова производились при помощи рамки 10×20 см у основания ствола и на высоте130 см от земли с двух сторон света (север и юг) (Горшков, 1990). В описаниях оценивали следующие характеристики: видовое разнообразие, общее проективное покрытие и покрытия отдельных видов лишайников. Всего было выполнено 1792 описания на 448 деревьях. Для анализа общего видового разнообразия и встречаемости лишайников были учтены все виды, обитающие на стволах деревьев на высоте от 0 до 2 м.
Для каждого фрагмента были рассчитаны следующие показатели эпифитного лишайникового покрова: общее число видов, среднее число видов в описании, среднее общее проективное покрытие, встречаемость и покрытие отдельных видов лишайников. Обработка данных выполнена на основе регрессионного анализа в среде StatGraphics (Ивантер, Коросов, 2003).
Результаты
Видовой состав эпифитных лишайников на стволах сосны в условиях г. Костомукши насчитывает 25 видов. Согласно литературным данным, на территориях, непосредственно граничащих с городом – в заповеднике «Костомукшский», а также в промышленной зоне г. Костомукши, на стволах сосны встречается 42 вида лишайников (Фадеева, Дубровина, 1995). Таким образом, на территории города выявлено около 60 % потенциально возможного биоразнообразия лишайников. Общее проективное покрытие лишайников в отдельных описаниях варьирует от 0 до 85 %, составляя в среднем 10 %. Число описаний без лишайников – 548 (31 %). Число видов в описаниях изменяется от 0 до 9. Доминантными видами являются Hypogymnia physodes, Parmeliopsis ambigua, P. hyperopta, Imshaugia aleurites, виды рода Cladonia, что типично для Фенноскандии и Северо-Запада России (Горшков, 1990; Halonen et al., 1991; Bruteig, 1993; Гимельбрант, Ростова, 1998; Катенина, 1999; Горшков, Тарасова, 2000; Лебедева, 2004 и др.). Единично встретились такие виды, как Alectoria sarmentosa, Cladonia macilenta, Pseudevernia fufruraceae. В 6 фрагментах сосновых сообществ был обнаружен вид Bryoria fremontii, занесенный в Красную книгу Российской Федерации и Республики Карелия (1988, 2007).
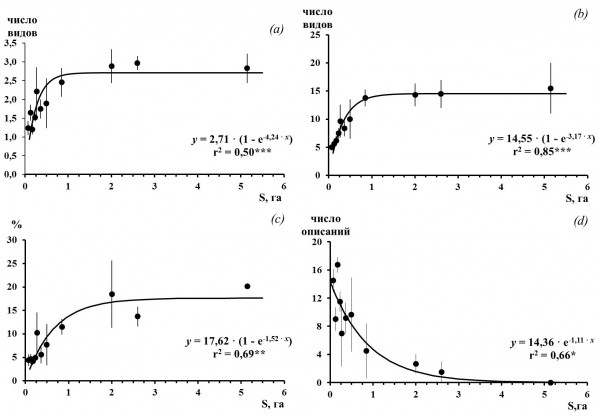
Рис. 2. Среднее число видов в описании (a), общее число видов во фрагменте (b), среднее общее покрытие лишайников (c) и число описаний без лишайников (d) во фрагментах сосновых сообществ разной площади на территории г. Костомукши. 1– среднее арифметическое и его ошибка; 2 – коэффициент аппроксимации; 3 – уровень значимости, соответственно 0.05 (*), 0.01(**) и 0.001 (***)
Fig. 2. The average number of lichen species in the sample plots (a), the total number of lichen species in fragments (b), the average total lichen cover (c) and the number sample plots without lichens (d) in fragments of pine forests in Kostomuksha. 1 – the mean and its error; 2 – coefficient of regression; 3 – the level of significance, respectively, 0.05 (*), 0.01 (**) and 0.001 (***)
Площадь 56 изученных фрагментов естественных сосновых насаждений варьирует от 0.04 до 6.13 га. Средняя площадь фрагмента составляет 0.62 га, 49 фрагментов (88 %) имеют площадь менее 1 га. Более мелкие фрагменты находятся, как правило, в центральной части города, а более крупные – на окраине. Выявлена статистически значимая зависимость между площадью сосновых насаждений и основными характеристиками эпифитного лишайникового покрова (рис. 2). Так, общее проективное покрытие лишайников, общее число видов, среднее число видов в описании с увеличением площади сосновых насаждений увеличиваются, а число описаний без лишайников, напротив, снижается. Стабилизация значений характеристик видового разнообразия (общего и среднего числа видов) регистрируется в насаждениях площадью 1 га и выше. Характеристики обилия – общее проективное покрытие лишайников и процент описаний с лишайниками достигают максимума и в дальнейшем не изменяются в сообществах площадью 2 га и более.
Обсуждение
Одной из основных причин снижения разнообразия и обилия лишайников при фрагментации местообитаний является невозможность сообщества поддерживать стабильные условия микроклимата во фрагментах малой площади (Gignac, Dale, 2005; Coxson, Stevenson, 2007; Esseen, 2006; Hedenеs, Ericson, 2008; Svoboda et al., 2010). Так, показано, что значения микроклиматических параметров (освещенность, температура, влажность) в зависимости от расстояния от края фрагмента стабилизируются лишь на расстоянии 12-30 м (Gignac, Dale, 2005). Кроме этого, предполагается, что данный фактор влияет на общий гидрологический режим территорий, и в ландшафтах с высокой степенью фрагментации уровень влажности в целом ниже, чем в менее фрагментированных (Svoboda et al., 2010). Поэтому ожидается, что ответ видов, чувствительных к уровню влажности, на действие фрагментации, будет более выраженным. Например, в исследовании, касающемся семейства Collemetaceae (изобилующего гигрофильными видами) показано, что виды рода Collema оказались более чувствительными к действию краевого эффекта, чем род Leptogium (Hedenеs, Ericson, 2008).
Другая причина «эффекта фрагментации» связана со способностью размножения некоторых видов преимущественно фрагментами талломов (Dettki, 1998; Dettki et al., 1998; Keon, Muir, 2002; Hilmo et al., 2005 и др.). И иногда наличие источника новых талломов для успешного расселения вида является более важным, чем наличие подходящих субстратов и микроклиматических условий. Этот факт был показан в исследовании роста трансплантантов талломов редкого вида Usnea longissima, размножающегося фрагментами слоевищ (Keon, Muir, 2002). Однако часто оба фактора влияют одновременно. Так, для хвойных лесовпоказано, что биомасса и длина талломов Alectoria sarmentosa (с данным типом размножения) увеличиваются от края вглубь леса и максимальные значения этих показателей отмечены в Швеции на расстоянии 58 м от опушки, в Канаде – 25-50 м (Еsseen, Renhorn, 1998; Essen, 2006). Авторы предположили, что для расселения данного вида важны также наличие подходящего субстрата и стабильные показатели микроклимата. На опушке леса талломы Alectoria sarmentosa получают физические повреждения сильным ветром.
В лихенофлоре г. Костомукши лишь у одного вида - Alectoria sarmentosa - преобладает такой тип размножения, но этот вид не может оказать заметного влияния на показатели лишайникового покрова. Остальные виды размножаются спорами полового размножения и специализированными органами вегетативного размножения – соредиями и изидиями, и, вероятно, этот процесс не зависит от фрагментации.
Вопрос о необходимом минимальном размере фрагмента, способном поддерживать высокие показатели биоразнообразия и обилия лишайников, пока остается открытым. В исследовании, выполненном в 27 фрагментах площадью от 0.03 до 4 га в хвойных лесах Канады, низкая биомасса Usnea spp. была зарегистрирована в семи массивах, наименьших по площади (Bryoria spp., Evernia mesomorpha); зависимости параметров лишайников от размера фрагментов лесных насаждений не выявлено (Rheault at al., 2003). В другом канадском исследовании, биомасса видов родов Bryoria, Evernia, Usnea возрастала от края фрагмента вглубь сообщества, стабилизация значений регистрировалась на расстоянии 30 м (Boudreault et al., 2008). Для осиновых лесов Канады, например, минимальный размер фрагмента, в котором возможно поддержание микроклимата, сохранение биоразнообразия и обилия мхов и лишайников, составляет 900 м2 (Gignac, Dale, 2005). В нашем исследовании выявлена необходимость сохранения более крупных фрагментов лесных сообществ (1‒2 га). Вероятно, на лишайники из небольших лесных массивов, расположенных в городе, оказывает влияние не только нарушение микроклимата, но и фоновое городское загрязнения, приводящее к усилению «эффекта фрагментации».
Заключение
В условиях урбанизированных территорий размер фрагментов растительного сообщества, обеспечивающий поддержание видового разнообразия лихенобиоты, увеличивается более чем в 10 раз. Проведенные исследования показали, что размеры фрагментов естественного леса на территории города таежной зоны определяются не только зональным типом растительности, но и фактором загрязнения, что приводит к необходимости увеличения их минимальной площади до 1-2 га.
Библиография
Бязров Л. Г. Лишайники в экологическом мониторинге. М.: Научный мир, 2002. 336 с.
Волков А. Д. Типы леса Карелии. Петрозаводск: Изд-во КарНЦ РАН, 2008. 180 с.
Гимельбрант Д. Е., Ростова Н. С. Вертикальная изменчивость структуры лишайникового покрова и биотические группировки эпифитных лишайников сосны (Pinus sylvestris L.) на Валаамском архипелаге // Комплексные природоведческие исследования на Северо-Западе России: Валаамская и Кургальская экспедиции СПбОЕ: Тр. С.-Петербургского общества естествоиспытателей. СПб, 1998. Т. 92. Сер. 1. С. 29-42.
Горшков В. В. Изменение во внеярусной растительности при атмосферном загрязнении // Влияние промышленного атмосферного загрязнения на сосновые леса Кольского полуострова. Л, 1990. С. 145-167.
Горшков В. В., Тарасова В. Н. Влияние лесных пожаров на эпифитный лишайниковый покров сосновых лесов южной Карелии // Раст. ресурсы. 2000. Т. 36. № 1. С. 18-29.
Государственный доклад о состоянии окружающей среды Республики Карелия в 2009 г. / Мин-во сел., рыб. хоз-ва и экологии Республики Карелия. Петрозаводск, 2010. 296 с.
Ивантер Э. В., Коросов А. В. Введение в количественную биологию: Учеб. пособие. Изд-во ПетрГУ, Петрозаводск, 2003. 304 с.
Катенина О. А. Эпифитные лишайники в составе лихенофлоры Новгородской области // Новости систематики низших растений. СПб.: Наука, 1999. Т. 33. С. 130-138.
Красная книга Республики Карелия. Петрозаводск: Карелия, 2007. 364 с.
Красная книга РСФСР: Растения. М.: Росагропромиздат, 1988. 591 с.
Лебедева М. Ю. Видовой состав эпифитных лишайников сосны севера русской равнины (на примере Карелии, Ленинградской, Московской и Новгородской областей) // Новости систематики низших растений. СПб.: Наука, 2004. Т. 37. С. 271-284.
Лосев К. С. Преобразование естественных ландшафтов // Современные глобальные изменения природной среды. В 2 т. Т. 2. М.: Научный мир, 2006. С. 203-212.
Фадеева М. А., Дубровина Н. Н. Материалы к флоре лишайников заповедника "Костомукшский" и промышленной зоны г. Костомукши // Флористические исследования в Карелии. Вып. 2. Петрозаводск, 1995. С. 68-84.
Boudreault C., Bergeron Y., Drapeau P., Lo´pez L. M. Edge effects on epiphytic lichens in remnant stands of managed landscapes in the eastern boreal forest of Canada // Forest Ecology and Management. № 255. 2008. P. 1461–1471.
Bruteig I. E. Large-scale survey of the distribution and ecology of common epiphytic lichens on Pinus sylvestris in Norway // Ann. Bot. Fenn. 1993. Vol. 30. № 3. P. 161-179.
Coxson D. S., Stevenson S. K. Influence of high-contrast and low-contrast forest edges on growth rates of Lobaria pulmonaria in the inland rainforest, British Columbia // Forest Ecology and Management. 2007. № 253. P. 103-111.
Dettki H. Verbreitung von Fragmenten zweier Bartflechtenarten // Sauteria. 1998. № 9. P. 123-131.
Dettki H., Edman M., Esseen P-A., Hedenеs H., Jonsson BG., Kruys N., Moen J., Renhorn K-E. Screening for species potentially sensitive to habitat fragmentation // Ecography. 1998. Vol. 21. № 6. P. 649-652.
Esseen P-A. Edge influence on the old-growth forest indicator lichen Alectoria sarmentosa in natural ecotones // Journal of Vegetation Science. 2006. № 17. P. 185-194.
Esseen P-A., Renhorn K. E. Edge effects on an epiphytic lichen in fragmented forests // Conservation Biology. 1998. Vol. 12. № 6. P. 1307-1317.
Gignac L. D., Dale M. R. T. Effects of fragment size and habitat heterogeneity on cryptogam diversity in the low-boreal forest of western Canada // The Bryologist. 2005. Vol. 108. № 1. P. 50-66.
Halonen P., Hyvärinen M., Kauppi M. The epiphytic lichen flora on conifers in relation to climate in the Finnish middle boreal subzone // Lichenologist. 1991. Vol. 23. № 1. P. 61-72.
Hedenеs H., Ericson L. Species occurrences at stand level cannot be understood without considering the landscape context: cyanolichens on aspen in boreal Sweden // Biological Conservation. 2008. Vol. 141. № 3. P. 710-718.
Hilmo O., Hytteborn H., Holien H. Do different logging strategies influence the abundance of epiphytic chlorolichens? // Lichenologist. 2005. Vol. 37. № 6. P. 543-553.
Keon D. B., Muir P. S. Growth of Usnea longissima across a variety of habitats in the Oregon Coast Range // The Bryologist. 2002. Vol. 105. № 2. P. 233-242.
Meffe G. K., Carroll C. R., and contributors. Principles of conservation biology / 2nd ed. Sunderland. Massachusetts, USA: Sinauer Association Publish., 1997. 729 p.
Otólora M. G., Martнnez I., Belinchуn R., Widmer I., Aragуn G., Escudero A., Scheidegger C. Remnants fragments preserve genetic diversity of the old forest lichen Lobaria pulmonaria in a fragmented Mediterranean mountain forest // Biodiversity and Conservation. 2011. № 20. P. 1239-1254.
Rheault H., Drapeau P., Bergeron Y., Esseen P-A. Edge effects on epiphytic lichens in managed black spruce forests of eastern North America // Canadian Journal of Forest Research. 2003. Vol. 33. № 1. P. 23-32.
Svoboda D., Peksa O., Veselá J. Epiphytic lichen diversity in central European oak forests: assessment of the effects of natural environmental factors and human influences // Environmental Pollution. 2010. Vol. 158. № 3. P. 812-819.

 © 2011 - 2026
© 2011 - 2026