



Введение
Процесс поэтапного выполнения флоро-фаунистического районирования Северной Евразии по отдельным биотическим компонентам обнажил ряд проблем, подробному обсуждению которых посвящена серия наших прежних публикаций (Блинова, Равкин, 2008; Равкин и др., 2014; Равкин и др., 2015а, б; Ravkin et al., 2015). Высокая степень индивидуальности каждой из рассматриваемых биотических групп приводит к существенным различиям в иерархии и местоположении проводимых границ, а также в объеме таксонов классификации. В основном это обусловлено групповой спецификой экологической толерантности животных и древесных растений, проявляющейся на общем фоне пространственной изменчивости современного гидротермического режима. При этом наиболее отчетливо видна связь изменчивости биоты с зональностью и провинциальностью, тогда как их интегральное влияние отражается в виде диагонального смещения границ (диагональности).
Полного совпадения зонально-подзональных границ и выделенных в процессе районирования, как правило, нет, так же как и стопроцентного сходства в списках участков, входящих в те или иные таксоны в различных схемах частного районирования. Существенные различия подобного рода нередки и в представлениях наших предшественников – в зависимости от использованных ими принципов, методов и состава анализируемых объектов. При этом авторы обычно считают или, по крайней мере, подразумевают, что каждое выполненное ими деление – не только единственно верное, но и может быть распространено на все остальные группы организмов или хотя бы их часть.
На наш взгляд, все результаты частного районирования в той или иной мере адекватности отражают реально существующую пространственную изменчивость объектов определенного слоя в общей совокупности, как в статистическом ансамбле со слабыми внутренними связями и значительным влиянием внешних средовых ограничений. Это связано с высокой степенью индивидуальности реакции различных видов и их групп на среду, а также с особенностями исследовательского аппарата. При этом разные группы объектов, подходов и методов анализа, включая индивидуальность аналитика, не антагонистичны, а дополняют в представлениях друг друга, то есть комплементарны по отношению между собой и со всей неоднородностью объектов, особенно в том случае, когда использованы формализованные методы агрегации.
В связи с этим при выполнении комплексного флоро-фаунистического районирования Северной Евразии возникает проблема обобщения полученных частных, зачастую весьма противоречивых, представлений о пространственной дифференциации отдельных биотических компонентов (Равкин и др., 2017). Определенные трудности связаны еще и с различным объемом понятия вида и других таксонов в систематике разных организмов, их видового богатства и степени изученности распространения. Поэтому простое объединение списков видов (или других таксонов) неизбежно приводит к доминированию закономерностей, свойственных преобладающим по разнообразию группам организмов. Например, беспозвоночным по отношению к растениям, растениям – по сравнению с позвоночными. Этим обусловлены полные или значимые потери информации об особенностях распространения менее представительных групп организмов, в то время как при биотическом районировании желательно максимально сохранить не только знания о специфике всех видов при их объединении, но и представления по всем анализируемым группам организмов. Для этого предложен принцип равнозначности закономерностей по выделенным группам объектов (Равкин и др., 2011). Однако это по-прежнему не отменяет актуальности проблемы оценки устойчивости проводимых при районировании границ.
Обычно подобные проблемы решаются за счет увеличения анализируемой выборки, дополнительного привлечения информации о все большем числе видов и/или прочих биотических групп. Мы использовали для этого усреднение матриц коэффициентов сходства, как адекватное, в какой-то мере, объединение закономерностей географической изменчивости флор и фаун все большего числа эталонных групп организмов, по которым имеется сравнимая информация по их распространению. Для выравнивания представительности групп организмов перед усреднением проводили нормирование на минимальную сумму значений сходства по каждой матрице в целом, без диагональных показателей. Для этого значения коэффициентов уменьшали во столько раз, во сколько сумма их больше по данной группе видов, чем матрицы с минимальной суммой. Кластерный анализ усредненных таким образом матриц сходства дает общее представление о неоднородности всех эталонных групп, вместе взятых, с учетом допущения их равнозначности.
Однако на этом проблемы не исчерпываются. Даже если стандартизировать методы, подходы и исходные ограничения, остается компонент изменчивости, связанный с репрезентативностью выборки групп организмов. Ее можно условно считать достаточной, если прибавление других эталонных групп уже не меняет полученных представлений либо колебания в принадлежности к таксонам свойственны лишь пограничным территориям. Для оценки устойчивости границ можно использовать так называемую «бутстрэп поддержку» (Efron, 1979), хотя правомерность применения этого метода к матрицам сходства еще не доказана и возникают программные трудности с допустимым объемом анализируемого числа проб.
Опыту подобной проверки стабильности границ и объемов таксонов классификации посвящена данная статья. Такая попытка осуществлена на примере дополнения трех матриц сходства: по фауне наземных беспозвоночных (в качестве эталонной группы взяты жуки – 529 видов), водных и наземных позвоночных (рыб и круглоротых – 335 видов, земноводных – 40 видов, пресмыкающихся – 170 видов, птиц – 734 вида, млекопитающих – 341 вид) и древесных растений (536 видов), всего 2156 видов. К этим трем матрицам коэффициентов сходства добавлена четвертая матрица сходства флоры всех сосудистых растений, включая древесные, по 1643 родам. В итоге объединенная выборка включает информацию по 3799 признакам (частично по видам и по родам). Предварительное тестирование различий результатов классификации по 536 видам, относящимся к 73 родам древесных растений, показало, что на видовом уровне выявлена бóльшая связь с провинциальной изменчивостью среды, а по родам – с зонально-подзональной. Причем влияние этих факторов прослеживается в обеих классификациях, но степень совпадения с зональностью возрастает по мере роста числа и ранга систематических таксонов в анализируемой выборке.
Статья, предлагаемая вниманию читателей, представляет собой продолжение нашей публикации в журнале «Принципы экологии» (2017. № 1), поэтому она написана как дополнение к ней. Текст ее сокращен по объему таким образом, чтобы максимально избежать повторений.
Материалы
Описание публикаций, где подробно охарактеризованы материалы и даны ссылки на прежние статьи, приведено во Введении. Исходные сведения о распределении сосудистых растений (на уровне рода) заимствованы из базы данных портала «Биодат» (Биодат..., 2017). Обобщение их выполнено Ю. И. Макеевой. В частности, при этом использована публикация Ю. Д. Нухимовской с соавторами (2003). Дополнительные сведения по флоре архипелага Земля Франца-Иосифа взяты нами из статьи Д. С. Мосеева и Л. А. Сергиенко (2017).
Методы
Описание методов содержат статьи, упомянутые во Введении, а также публикации Ю. С. Равкина, В. Л. Куперштоха и В. А. Трофимова (Равкин и др., 1978) и Ю. С. Равкина, С. Г. Ливанова (2008). Вкратце их особенности сводятся к следующему. Информация о классифицируемых объектах представляется в виде матрицы коэффициентов сходства фаун и/или флор выделенных участков. В качестве характеристики «разброса» значений коэффициентов принят средний квадрат отклонения от среднего по всей матрице (дисперсия). Сопоставляемые варианты сначала классифицируются таким образом, чтобы доля дисперсии исходной матрицы коэффициентов, учитываемой классификацией, была наибольшей. Для этого из коэффициентов сходства вычитается среднее по их матрице значение. В результате все коэффициенты меньше среднего становятся отрицательными. Затем ищется пара вариантов, объединение которых в один класс уменьшает начальную дисперсию на максимально возможную величину, т. е. пробы с наибольшим сходством. После этого столбцы и строки матрицы коэффициентов, соответствующие найденной паре вариантов, поэлементно суммируются. На агрегированной так матрице процедура объединения повторяется. Такой поиск и агрегацию продолжают до тех пор, пока доля учитываемой дисперсии увеличивается. Это происходит, если объединяются варианты с положительными коэффициентами. В результате получается некоторая классификация – объединение вариантов населения по их максимальному сходству в незаданное число классов. При этом коэффициенты проб внутри классов в основном положительны, а между классами – отрицательны.
Далее проводят оценку дисперсии коэффициентов сходства, учтенной составленной классификацией. Для этого все коэффициенты внутри найденных классов уменьшают на их среднее, а коэффициенты сходства между классами увеличивают на абсолютную величину их среднего. В результате получается остаточная матрица коэффициентов, дисперсия которой меньше, чем у исходной. Разница этих дисперсий, отнесенная к начальной, составляет долю уменьшения. При классификации порог значимости по представительности для выделения подобластей и провинций равен четырем участкам, то есть эти таксоны могут быть выделены, если в них входит не менее четырех участков первичного разделения территории.
Результаты
Флористическая и флоро-фаунистическая классификации
Иерархическая классификация флоры сосудистых растений по 1643 родам 597 участков Северной Евразии представлена ниже и на рис. 1. Номинация таксонов классификации выполнена формальным сочетанием названий входящих в состав таксона крайних (пограничных) участков в направлении с запада на восток и с севера на юг.
Северо-Восточный островной регион
1. Северо-Восточная (островная) полярно-пустынная подобласть (7 полярно-пустынных участков и 1 субарктический тундровый). Далее слово «участков» опущено.
Северный (Печенго-Анадырский) редколесно-тундровый регион (тундровых арктических и субарктических 26 и 43, редколесных 20, северотаежных 3).
Подобласти
2. Западная (Печенго-Байдарацкая) редколесно-тундровая (тундровых арктических и субарктических 4 и 8, редколесных 4, северотаежных 3);
3. Северная срединная (Байдарацко-Колымская) редколесно-тундровая (тундровых арктических и субарктических 21 и 16, редколесных 7);
4. Южная срединная анклавная (Пясинско-Колымская) редколесная (субарктический тундровый 1, редколесных 9);
5. Северо-Восточная (Колымско-Чукотская) тундровая (тундровых арктических и субарктических 1 и 6);
6. Юго-Восточная (Корякская) тундрово-субарктическая (тундровых субарктических 12).
Срединный (Балтийско-Камчатский) редколесно-степной регион (тундровых субарктических 4, редколесных 58, северо- и среднетаежных 29 и 76, южно- и подтаежных 45 и 25, широколиственно-лесных 16, горно-таежных 29, лесостепных и степных 20 и 73, полупустынных 8).
7. Западная (Балтийско-Байкальская) северотаежно-степная подобласть (редколесных 2, северо- и среднетаежных 24 и 32, южно- и подтаежных 20 и 16, широколиственно-лесных 14, горно-таежных 9, лесостепных и степных 20 и 45, полупустынных 2).
Провинции
7.1. Северо-Западная (Кольско-Енисейская) северо-среднетаежная (редколесных 2, северо- и среднетаежных 24 и 21, южнотаежный 1);
7.2. Западная (Балтийско-Уральская) среднетаежно-степная (средне- и южнотаежных 5 и 8, подтаежных и широколиственно-лесных 15 и 14, лесостепных и степных 12 и 17, полупустынных 2);
7.3. Срединная южная (Урало-Байкальская) среднетаежно-степная (среднетаежных 6, южно- и подтаежных 11 и 1, горно-таежных 9, лесостепных и степных 8 и 28).
8. Срединная восточная (Енисейско-Охотская) редколесно-степная подобласть (редколесных 51, северо- и среднетаежных 5 и 34, южно- и горно-таежных 7 и 20, степных 6).
Провинции
8.1. Северо-Восточная (Восточно-Сибирская) редколесно-таежная (редколесных 48, северо- и среднетаежных 3 и 17, горно-таежных 12);
8.2. Срединная северная (Эвенкийская) редколесно-среднетаежная (редколесных 3, северо- и среднетаежных 2 и 13, южнотаежный 1);
8.3. Юго-Восточная (Забайкальско-Охотская) среднетаежно-степная (средне- и южнотаежных 4 и 6, горно-таежных 8, степных 6).
Подобласти
9. Срединная притихоокеанская (Камчатско-Курильско-Сахалинская) тундрово-южнотаежная (тундровых субарктических 4, редколесных 5, средне- и южнотаежных 4 и 5).
10. Юго-Западная (Карпатско-Уральская) полупустынно-степная (подтаежных 2, степных 22, полупустынных 6);
11. Юго-Восточная притихоокеанская (Амурско-Уссурийская) среднетаежно-подтаежная (средне- и южнотаежных 6 и 13, подтаежных 7, широколиственно-лесных 2);
Юго-Западный (Кавказско-Среднеазиатский) лесо-пустынно-степной регион (широколиственно- и горно-лесных 2 и 11, равнинно- и горно-степных 8 и 13, полупустынных 29, пустынных 51).
Подобласти
12. Западная (Крымско-Кавказская) горно-лесная с проникновением в степь и полупустыни (широколиственно- и горно-лесных 2 и 11, степных 7, полупустынных 3);
13. Северо-Западная (Волжско-Эмбинская) полупустынно-пустынная (полупустынных 1, пустынных 5);
14. Северо-Восточная (Казахстанская) полупустынно-пустынная (степных 1, полупустынных 16, пустынных 28);
15. Юго-Западная (Узбекско-Туркменская) полупустынно-пустынная (горно-степных 3, полупустынных 2, пустынных 13);
16. Юго-Восточная анклавная (Таджикско-Киргизская) горно-степная, с проникновением в полупустыни и пустыни (горно-степных 10, полупустынных 6, пустынных 5).
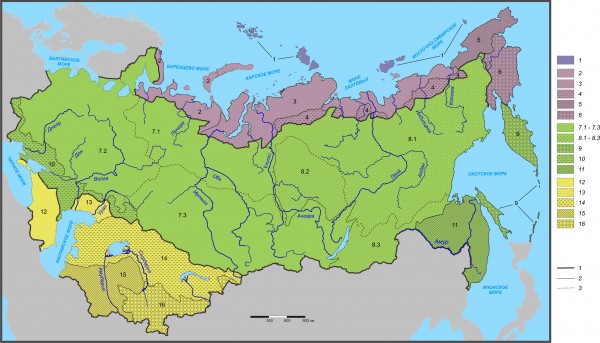
Рис. 1. Районирование Северной Евразии по флоре сосудистых растений на уровне рода. При составлении всех карт Северной Евразии использована нормальная равнопромежуточная коническая проекция В. В. Каврайского. Характеристики таксонов см. выше в тексте классификации; границы: 1 – регионов, 2 – подобластей, 3 – провинций
Fig. 1. Zoning of Northern Eurasia made on the flora of vascular plants at the genus level. In the preparation of all maps of Northern Eurasia the Kavrayskiy normal equidistant conical projection was used. See characteristics of the taxa above in text of classification; borders of: 1 – regions; 2 – subregions; 3 – provinces
Результаты флоро-фаунистического районирования по четырем матрицам – позвоночные, беспозвоночные, древесные растения по видам и все сосудистые растения по родам – приведены ниже и на рис. 2.
Полярно-пустынный островной регион
1. Полярно-пустынно-тундровая островная подобласть (участков – полярно-пустынных 7, тундровых арктических 2, субарктический 1).
Срединный тундрово-степной регион (тундровых арктических 24, субарктических 47, редколесных 78, северо- и среднетаежных 32 и 76, южно- и подтаежных 45 и 25, горно-таежных и широколиственно-лесных 29 и 16, лесостепных и степных 20 и 61).
2. Тундровая материковая подобласть (тундровых арктических 24, субарктических 30, редколесных 1).
Провинции
2.1. Западная (тундровых арктических 2, субарктических 13, редколесный 1);
2.2. Срединная арктическая (арктических тундровых 21, субарктических 11);
2.3. Восточная (арктический тундровый 1, субарктических тундровых 6).
3. Тундрово-таежная притихоокеанская (Корякско-Камчатская) подобласть (тундровых субарктических 14, редколесных 5, среднетаежных 3).
Провинции
3.1. Тундровая (тундровых субарктических 4);
3.2. Редколесно-тундровая (тундровых субарктических 10, редколесных 5, среднетаежных 3).
4. Предтундрово-редколесная подобласть (тундровых субарктических 3, редколесных 67, северотаежных 3).
Провинции
4.1. Западная (редколесных 7, северотаежный 1);
4.2. Срединная (субарктический тундровый 1, редколесных 19, северотаежный 1);
4.3. Юго-Восточная (редколесных 35, северотаежных 1);
4.4. Восточная (тундровых субарктических 2, редколесных 6).
5. Таежно-степная подобласть (редколесных 5, северо- и среднетаежных 29 и 72, южно- и горно-таежных 35 и 29, подтаежных 23 и широколиственно-лесных 14, лесостепных и степных 20 и 61).
Провинции
5.1. Северная таежная (редколесных 5, северо- и среднетаежных 29 и 62, южно- и горно-таежных 19 и 25, подтаежных 5, лесостепных и степных 4 и 5);
5.2. Юго-Западная южнотаежно-степная (южнотаежных и подтаежных 2 и 13, широколиственно-лесных 14, лесостепных и степных 16 и 33);
5.3. Южная степная (степных 17);
5.4. Юго-Восточная среднетаежно-степная (средне- и южнотаежных 10 и 14, подтаежных и горно-таежных 5 и 4, степных 6).
6. Таежно-лесная притихоокеанская подобласть (средне- и южнотаежных 1 и 10, подтаежных и широколиственно-лесных по 2).
Юго-Западный (Кавказско-Среднеазиатский) регион (широколиственных и горных лесов 2 и 11, степных равнинных и горных 37 и 20, полупустынных и пустынных 37 и 51).
Подобласти
7. Крымско-Кавказская (широколиственно-лесных 2, горно-лесных 11, степных 9, полупустынных 3).
8. Пустынно-степная Казахстанско-Среднеазиатская (горно-степных 13, степных 11, полупустынных 34, пустынных 51).
Провинции
8.1. Северо-Западная пустынно-полупустынная (степных 1, полупустынных 10, пустынных 5);
8.2. Северо-Восточная степная (степных 9);
8.3. Срединная полупустынно-пустынная (степных 1, полупустынных 18, пустынных 32);
8.4. Юго-Западная пустынно-горностепная (горно-степных 7, полупустынных 5, пустынных 14);
8.5. Юго-Восточная горно-степная (горно-степных 6, полупустынных 1).
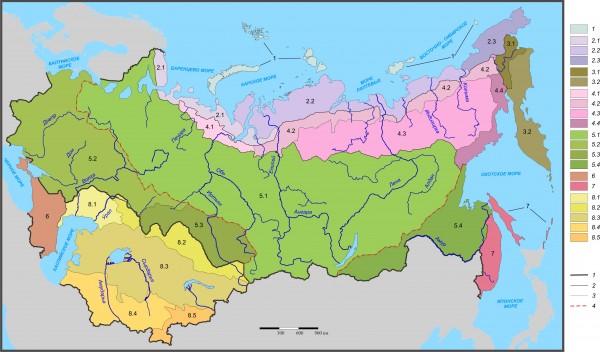
Рис. 2. Флоро-фаунистическое районирование Северной Евразии по среднему сходству четырех матриц (на уровне вида по древесным растениям, наземным и водным позвоночным; на уровне рода – по всем сосудистым растениям). Характеристики таксонов см. выше в тексте классификации; границы: 1 – регионов, 2 – подобластей, 3 – провинций, 4 – максимальной диагональности
Fig. 2. Flora-fauna zoning of Northern Eurasia by the average similarity of the four matrices (at the species level for woody plants, terrestrial and aquatic vertebrates, at the genus level – for all vascular plants). See characteristics of the taxa above in text of classification; borders of: 1 – regions, 2 – subregions, 3 – provinces, 4 – maximum diagonality
Флоро-фаунистическая структура
Структурный граф, отображающий сходство флор сосудистых растений выделенных подобластей, иллюстрирует три тренда (рис. 3). Первый из них, направленный с севера на юг, совпадает с увеличением теплообеспеченности при одновременном уменьшении влагообеспеченности. В результате этих изменений гидротермического режима, и прежде всего соотношения тепла и влаги, формируются зональные изменения. На графе они отображены вертикальным рядом изменений в направлении от полярных пустынь к тундрам, редколесьям, лесам, лесостепи, степи и далее к полупустыням и пустыням. Изменения, иллюстрируемые горизонтальным рядом, связаны с провинциальностью (континентальностью) и увеличением абсолютных высот местности. Влияние высот проявляется в основном в Крымско-Кавказской и Казахстанско-Среднеазиатской подобластях, то есть на Кавказе и Памиро-Тянь-Шанской части территории. Аналогичный граф, выстроенный по изменению сходства флор регионов, вследствие явной переагрегации иллюстрирует в основном широтные (зональные) отличия. Тогда как провинциальную изменчивость можно отразить лишь на полярных островах, поскольку западная часть этих островов из-за отепляющего влияния северо-восточного ответвления системы Гольфстрим по флоре сосудистых растений оказывается ближе к материковой флоре, а восточные острова в процессе классификации выделяются в самостоятельный регион.
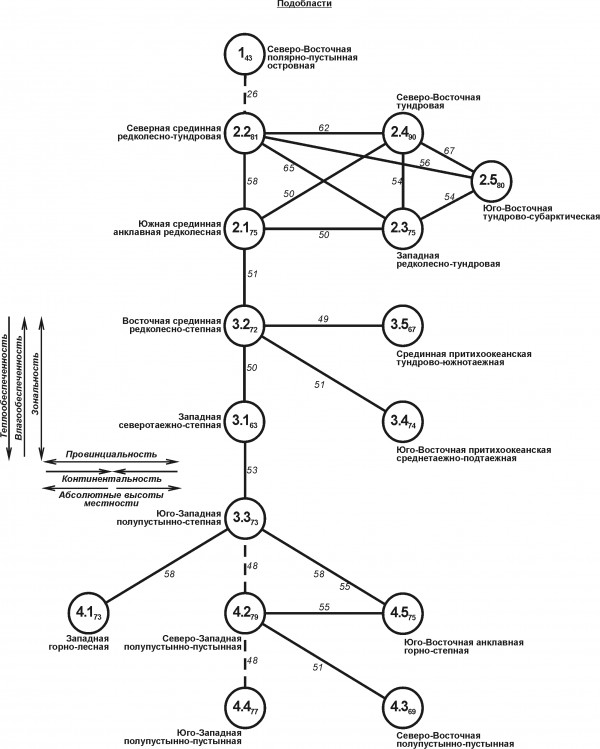
Рис. 3. Пространственно-типологическая структура флоры сосудистых растений Северной Евразии на уровне рода. Внутри значков приведены номера таксонов классификации (см. в тексте), в виде подстрочного индекса показана величина внутригруппового сходства. Между таксонами обозначена величина межгруппового сходства. Стрелки у перечня основных структурообразующих факторов среды указывают направление увеличения их влияния и флоро-фаунистические тренды
Fig. 3. Spatial-typological structure of flora of vascular plants of Northern Eurasia at the genus level. The numbers of taxa are inside the icons of the classification (see in the text), a subscript shows the value of intra-group similarity. Between the taxa the magnitude of intergroup similarities is indicated. The arrows at the list of basic structure-forming environmental factors indicate the direction of increasing their influence and floro-faunistic trends
Структурный граф флоро-фаунистического сходства по четырем усредненным матрицам полностью совпадает с таковым по трем усредненным матрицам (Равкин и др., 2017), поэтому здесь не приведен.
Экологическая организация флоро-фаунистической неоднородности
При анализе матрицы сходства по родам сосудистых растений наиболее велика связь неоднородности флоры Северной Евразии с теплообеспеченностью (62 % дисперсии матрицы коэффициентов сходства), а с зональностью и региональностью аналогичные оценки связи соответственно на 6 и 7 % меньше. Намного слабее связь с провинциальностью (17 %), а с инсулярностью и поясностью она и вовсе невелика (8 и 0.9 %). Всеми перечисленными факторами, вместе взятыми, учитывается 79 % дисперсии. При этом приращение информативности по отношению к теплообеспеченности прослежено только по региональности и инсулярности (16 и 1 %), остальные факторы полностью скоррелированы с предыдущими наборами. Природные режимы (неразделимые сочетания факторов) по иерархической и структурной классификациям увеличивают объясненную дисперсию матрицы на 7 %. Множественный коэффициент корреляции всех указанных связей сравнительно высок и равен 0.93.
Результаты аналогичных расчетов только по видам древесных растений, как правило, выше, за исключением провинциальности, связь с которой вдвое меньше. По родам всех сосудистых растений в целом по выявленным факторам оценки выше на 24 %, а с природными режимами связь на 6 % ниже. По всем сосудистым растениям факторы и режимы связаны всего на 6 % больше (множественный коэффициент корреляции всего на 0.045 больше). Результаты районирования по использованным признакам наиболее сходны с таковыми по биотическому и климатическому районированию (различия не превышают ± 2 %) и значительно отличаются от результатов флористического и физико-географического районирования (соответственно на 12 и 8 % меньше, чем по всем сосудистым растениям). Описания районирования взяты для сравнения из публикаций (Флора СССР, 1934; Рихтер, 1964; Курнаев, 1973; Udvardy, 1975; Воронов, Кучерук, 1977; Атлас СССР, 1983).
Оценки силы и общности связи тех же факторов и режимов по трем и четырем усредненным матрицам (1–3 – древесные растения, беспозвоночные и позвоночные животные по видам; 1–4 – то же самое, плюс все сосудистые растения по родам) сходны как с ранее рассчитанными значениями, так и между собой (табл. 1). В целом по четырем матрицам они на два процента ниже, чем по трем. Эти отличия явно недостоверны и свидетельствуют о достаточности представлений, полученных по первому варианту счета при усреднении трех матриц. То же свойственно и результатам географического районирования, оценки которых полностью совпадают по климатическому, биотическому и флористическому районированию (42; 39 и 33 %) и чуть меньше – по четырем матрицам для лесорастительного разделения территории (22 и 21 %).
Таблица 1. Оценка силы и общности связи неоднородности фауны и флоры сосудистых растений и факторов среды Северной Евразии
| Учтенная дисперсия | ||||
| Фактор, режим | индивидуально | нарастающим итогом | ||
| по трем матрицам | по четырем матрицам | по трем матрицам | по четырем матрицам | |
| Теплообеспеченность | 61 | 63 | 61 | 63 |
| Зональность | 51 | 52 | 61 | 63 |
| Региональность | 23 | 22 | 67 | 67 |
| Провинциальность | 20 | 20 | 67 | 67 |
| Инсулярность | 4 | 5 | 68 | 68 |
| Рельеф (равнина – горы) | 0.6 | 0.6 | 68 | 68 |
| Все факторы | 68 | 68 | 68 | 68 |
| Режимы иерархической классификации | 64 | 61 | 82 | 81 |
| Структурные | 57 | |||
| Все режимы | 70 | 70 | 83 | 81 |
| Все факторы и режимы | 83 | 81 | 83 | 81 |
| Множественный коэффициент корреляции | 0.91 | 0.90 | 0.91 | 0.90 |
Результаты взаимной (перекрестной) аппроксимации средних матриц сходства по трем и четырем исходным показывают в общем невысокое приращение информативности. Так, классификация, составленная по матрице средней по трем исходным, учитывает 64 % ее дисперсии, а выполненная по четырем исходным матрицам – 62 % (вместе 70 %). То есть приращение за счет объединения классификаций равно всего 6 %. Те же объединения по матрице средней, полученной из четырех исходных, дают соответственно чуть большее приращение: 8 % (в среднем по трем и четырем матрицам 7 %; табл. 2).
Таблица 2. Приращение информативности по двум классификациям, % учтенной дисперсии
| В среднем по матрицам | ||||||
| трем | четырем | В среднем | ||||
| трем | четырем | трем и четырем | четырем | трем | четырем и трем | |
| 64 | 62 | 70 | 61 | 63 | 69 | – |
| Приращение, % | 6 | Приращение, % | 8 | 7 | ||
Обсуждение
Сопоставляя результаты анализа неоднородности флоры сосудистых растений, выполненного на уровне рода и только древесных растений на уровне вида (Равкин и др., 2015б), можно отметить, что после включения данных по архипелагу Земля Франца-Иосифа по всем сосудистым растениям выделено четыре региона, а отдельно по видам древесных растений – три. То есть отличия сводятся к отсутствию по древесным растениям Полярно-пустынного островного региона, который входит в качестве подобласти в состав Северного региона. Это отличие в общем не принципиально и касается в основном иерархии подразделений. Гораздо больше отличия в границах Юго-Западного и Северо-Восточного регионов по видам древесных растений, а также Срединного и Юго-Западного (Кавказско-Среднеазиатского) регионов по родам всех сосудистых растений. Разница в том, что северная (европейская лесопустынно-степная) подобласть по древесным растениям входит вместе с более южными кавказскими и среднеазиатскими участками в Юго-Западный регион, а по родам сосудистых растений объединяется в Срединный регион. Это подчеркивает преобладающее влияние провинциальности в первом случае и зональности – во втором. Остальные отличия не принципиальны и так же, как ранее рассмотренные, относятся к разнице в иерархии выделенных таксонов.
По родам древесных растений отдельно и всех сосудистых растений вместе результаты районирования сходны. Значимые отличия сводятся к включению Северной редколесно-тундровой провинции, выделенной по родам древесных растений, в состав Срединного региона по родам всех сосудистых растений. Кроме того, в последний входит и кавказская часть территории, которая по родам древесных растений вместе со Среднеазиатской подобластью образует самостоятельный регион. При взаимной (перекрестной) аппроксимации этих матриц различия составляют 16 и 27 %, в среднем 22 %. Обеими классификациями эти матрицы аппроксимированы на 70 и 69 % (приращение составляет 27 и 33 % учтенной дисперсии, в среднем 30 %). Это свидетельствует об отсутствии принципиальных отличий между составленными классификациями при значительных различиях в их информативности.
При обобщенном сопоставлении конфигурации выделов карт районирования по родам всех сосудистых растений и выполненного ранее по видам древесных растений и фауне позвоночных в целом можно отметить их значительное сходство на уровне подобластей. Число регионов в них одинаково и равно четырем, но бросается в глаза вдвое меньшая дробность разделения на подобласти первого набора объектов (8 и 16).
Таким образом, можно утверждать, что в обоих случаях по сосудистым растениям (по родам) и усредненным матрицам сходства фауны и флоры только древесных растений (по видам) имеются в основном отличия в степени обобщения и в иерархии разделения территорий, отнесенных к разным таксонам классификаций. После усреднения трех и четырех матриц (по беспозвоночным и позвоночным животным, древесным растениям по видам и всем сосудистым по родам), главные тренды и их причины сохраняются прежними, так же как число выделенных подобластей. Число выделенных регионов по трем усредненным матрицам на один больше, чем по четырем. При этом выделение трех регионов свидетельствует о явной переагрегации, то есть нецелесообразности регионального уровня обобщения.
Итак, сопоставление флоро-фаунистических классификаций по трем и четырем матрицам (по видам отдельно и по видам и родам вместе) показало, что с прибавлением четвертой матрицы сходства (по сосудистым растениям в целом) принадлежность к регионам изменилась у 299 участков (50 % от 597), к подобластям – в 16 случаях (3 %) и к провинциям – у 82 (14 %). В целом число отклоняющихся участков равно 397 (66 %). Таким образом, по числу участков отклонения в классификациях очень велики, но связаны в основном с различием в составе регионов (81 сложный процент). Кроме того, следует учитывать, что из-за реализованной возможности изменения порога представительности (с 10 до 4 участков) по 28 из них выполнено дополнительное разделение двух подобластей, что тоже входит в число отклонений. Если не считать эти отличия и соотнести степень отклонений по числу участков к их сумме по трем уровням иерархии, то доля отклонений составляет 21 % (рис. 4).
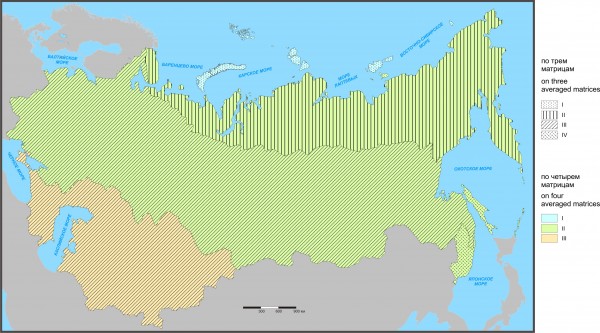
Рис. 4. Флоро-фаунистическое районирование по трем и четырем средним матрицам на уровне региона
Fig. 4. Floro-faunistic division of Northern Eurasia performed on three and four averaged matrices at the region level
Конфигурация выделов по подобластям существенно не изменилась, отмечены только смещения границ на один, реже два участка. Видимо, районирование после усреднения по трем матрицам достаточно полно отражает флоро-фаунистическую неоднородность Северной Евразии, а отличия при добавлении данных по всем сосудистым растениям (в ранге рода) принципиальных изменений на этом уровне не вносят (рис. 5). Степень несовпадения провинциальных границ показана на рис. 6.
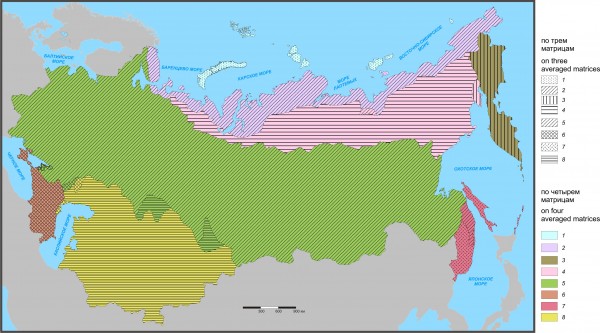
Рис. 5. Флоро-фаунистическое районирование по трем и четырем средним матрицам на уровне подобласти
Fig. 5. Floro-faunistic division of Northern Eurasia performed on three and four averaged matrices at the subregion level
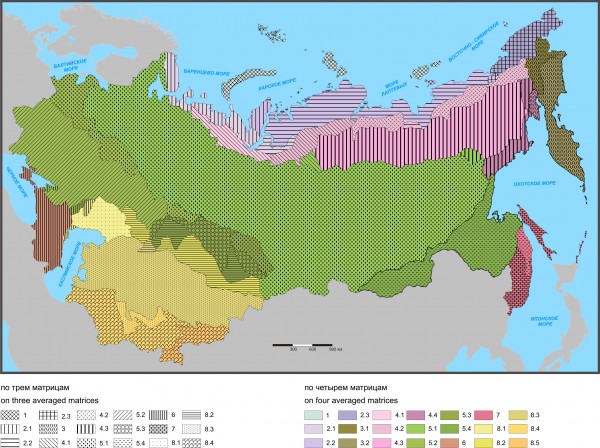
Рис. 6. Флоро-фаунистическое районирование по трем и четырем средним матрицам на уровне провинции
Fig. 6. Floro-faunistic division of Northern Eurasia performed on three and four averaged matrices at the province level
Структурные представления по трем и четырем матрицам после нормирования и усреднения абсолютно одинаковы, так же как и отдельно по всем сосудистым растениям. Оценки связи флоро-фаунистической изменчивости с факторами среды по трем и четырем матрицам (после усреднения частных матриц сходства) очень близки. Их различия колеблются в пределах от нуля до 3 %, при этом по четырем матрицам они, как правило, на эту часть доли меньше, чем по трем. Информативность результатов климатического, биотического и физико-географического районирования абсолютно одинакова и лишь лесорастительного на 1 % меньше.
Заключение
Аппроксимация средних матриц сходства классификациями, составленными по трем и четырем исходным, колеблется по модулю в пределах от 61 до 64 % учтенной дисперсии и составляет в среднем 62 %. Отклонение по этой доле колеблется от 0.8 до 3 %. В среднем оно равно 2 %. Аппроксимация матрицы сходства, полученной по четырем матрицам, классификацией, составленной по трем первым матрицам, даже чуть лучше, чем составленной по четырем матрицам (62 и 63 %, отклонение – 0.8 %).
Таким образом, по доле учтенной дисперсии различия очень невелики. Противоречие в оценках по числу участков и учтенной дисперсии связано с тем, что различия прослежены, как правило, в пограничных участках или периферических таксонах. Отличающиеся участки и их группы почти одинаково сходны с соседними таксонами, и отнесение их к тому или иному таксону равновероятно, а результаты незначительно отличаются по учтенной дисперсии, и, в общем, безразлично, какой принять вариант расчленения территории. Бóльшая часть отклонений связана с выделением регионов, деление на которые получено при повторной агрегации и в принципе мало информативно. Так, расчеты множественной оценки информативности показывают, что наиболее значимо деление территории на подобласти (после усреднения по трем и четырем матрицам – 49 и 48 % дисперсии). Деление на провинции при расчете множественной регрессии нарастающим итогом увеличивает учтенную дисперсию на 11 и 12 %, а на регионы – всего на 2 и 1 % (в целом в среднем на 13 %). Именно поэтому, несмотря на значительные отличия по числу участков, эти классификации почти не отличаются по информативности.
В целом складывается впечатление, что две составленные флоро-фаунистические классификации, существенно (на 66 %) различаются по числу участков первоначального разделения территории, но почти не отличаются по информативности как между собой, так и при перекрестной аппроксимации. На этом основании можно говорить о возможности составления двух (и, вероятно, более) классификаций с одинаковой информативностью, т. е. равнозначных по доле учитываемой ими дисперсии одной и той же матрицы сходства, но существенно различающихся в конфигурации границ. Причины этого, возможно, обусловлены наличием в одной сборной совокупности двух и более блоков с равнозначными по силе проявления, но разнонаправленными трендами. В основном же эти отличия связаны с несовпадением в иерархии выделенных таксонов и оценок информативности по иерархическому ряду: регион – подобласть – провинция. Первый из них (региональный) наименее значим по учтенной дисперсии, хотя с этим рангом деления связано наибольшее число отклонений по количеству участков. Для второго (подобластного) оценка максимальна, а для последнего (провинциального) уровня оценка занимает в ряду второе место. В итоге можно считать, что объединенное районирование по флоре древесных растений и фауне, судя по незначительным потерям учтенной дисперсии средней матрицы эталонных групп организмов, адекватно отражает общую флоро-фаунистическую неоднородность этой территории, но, естественно, в рамках сделанных допущений.
Библиография
Атлас СССР . М.: Главное управление геодезии и картографии, 1983. 260 с.
Биодат. Живая природа и биоразнообразие. Ареалы животных и растений . URL: http://biodat.ru/db/areal/index.htm (дата обращения: 15.12.2017).
Блинова Т. К., Равкин Ю. С. Орнитофаунистическое районирование Северной Евразии // Сибирский экологический журнал. 2008. Т. 15. № 1. С. 101–121.
Воронов А. Г., Кучерук В. В. Биотическое разнообразие Палеарктики: проблемы изучения и охраны // Биосферные заповедники: Труды I советско-американского симпозиума СССР, 5–17 мая 1976 г. Л.: Гидрометеоиздат, 1977. С. 7–20.
Курнаев С. Ф. Лесорастительное районирование СССР . М.: Наука, 1973. 203 с.
Мосеев Д. С., Сергиенко Л. А. К флоре островов архипелага Земля Франца-Иосифа и северной части архипелага Новая Земля (аннотированный список видов) // Ученые записки Петрозаводского государственного университета. 2017. № 4 (165). С. 48–64.
Нухимовская Ю. Д., Губанов И. А., Исаева-Петрова Л. П., Пронькина Г. А. Современное состояние биологического разнообразия на заповедных территориях России. Вып. 2. Сосудистые растения: В 2 ч. . М.: МСОП, 2003. 783 с.
Равкин Ю. С., Богомолова И. Н., Цыбулин С. М. Фаунистическое районирование Северной Евразии // Известия Российской академии наук. Серия географическая. 2015а. № 3. С. 29–40.
Равкин Ю. С., Богомолова И. Н., Цыбулин С. М. Сравнительное районирование Северной Евразии по древесным растениям и наземным позвоночным // Сибирский лесной журнал. 2015б. № 5. С. 42–53.
Равкин Ю. С., Богомолова И. Н., Цыбулин С. М. Экологическая организация флоро-фаунистической неоднородности Северной Евразии // Принципы экологии. 2017. № 1. С. 56–70.
Равкин Ю. С., Богомолова И. Н., Цыбулин С. М., Легалов А. А. Районирование Северной Евразии по фауне короедов (Scolytidae) // Евразийский энтомологический журнал. 2014. № 13(3). С. 271–279.
Равкин Ю. С., Куперштох В. Л., Трофимов В. А. Пространственная организация населения птиц // Птицы лесной зоны Приобья. Новосибирск: Наука. Сиб. отд-ние, 1978. С. 253–269.
Равкин Ю. С., Ливанов С. Г. Факторная зоогеография . Новосибирск: Наука, 2008. 205 с.
Равкин Ю. С., Седельников В. П., Сергеев М. Г., Титлянова А. А., Хмелёв В. А., Богомолова И. Н., Цыбулин С. М. Пространственно-типологическая дифференциация экосистем Западно-Сибирской равнины. Сообщение V. Экосистемы суши // Сибирский экологический журнал. 2011. Т. 18. № 6. С. 773–788.
Рихтер Г. Д. Физико-географическое районирование СССР // Физико-географический атлас мира. М.: Изд-во АН СССР и Главного управления геодезии и картографии СССР, 1964. С. 248–249.
Флора СССР . М.; Л., 1934. Т. 1. 302 с.
Efron B. Bootstrap Methods: Another Look at the Jackknife // Annals of Statistics. 1979. Vol. 7. № 1. P. 1–26.
Ravkin Yu. S., Bogomolova I. N., Tsybulin S. M., Legalov A. A. Zoning of Northern Eurasia based on the fauna of pine flower, fungus and leaf-rolling weevils (Coleoptera: Nemonychidae, Anthribidae, Rhynchitidae, Attelabidae) // Евразийский энтомологический журнал. 2015. № 14 (4). С. 367–373.
Udvardy M. D. F. Classification of the Biogeographic Provinces of the World Occasional № 18 Int. Union for Conservation of Nature. Morges, Swittzerland, 1975. P. 1–48.
Благодарности
Исследования, послужившие основой для настоящей статьи, выполнены по программе ФНИ государственных академий наук на 2013–2020 гг., проект № ААААА-А16-116121410122-4, и частично в рамках «Программы повышения конкурентоспособности Томского государственного университета». Авторы искренне признательны Т. Н. Добрецовой, В. С. Жукову, В. Г. Ивлиеву и Е. С. Равкину за ценные советы при обсуждении статьи до ее публикации.

 © 2011 - 2026
© 2011 - 2026