



Введение
Анализ демографической и пространственной структуры популяций чужеродных видов характеризует состояние колоний в новых условиях и позволяет оценить инвазивный потенциал вселенцев. На данный момент на территории юга Среднерусской возвышенности известно существование колоний ксерофильных видов наземных моллюсков: Xeropicta derbentina (Krynicki, 1836) и Brephulopsis cylindrica (Menke, 1828), расположенных в пределах г. Белгорода (Снегин и др., 2014, 2017). Естественным ареалом указанных видов являются территории Крыма, Кавказа и Малой Азии (Шилейко, 1978, 1984). На территории Крыма и Северного Причерноморья популяции данных видов в значительной степени изучены (Крамаренко, 2014), что позволяет сопоставить состояние колоний в различных частях ареала.
Целью работы являлся анализ демографической и пространственной структуры адвентивных колоний наземных моллюсков X. derbentina и B. cylindrica на территории г. Белгорода.
Материалы
Колонии моллюсков-вселенцев B. cylindrica и X. derbentina исследовали в течение весенне-осеннего сезона 2017 г. Полевые наблюдения проводили на участке, расположенном в окрестностях мелового карьера в черте г. Белгорода (50°62'64″ с. ш.; 36°51'77″ в. д.). Участок представляет собой открытую местность с преобладанием рудеральной растительности. Пробы отбирались каждый месяц с мая по сентябрь.
Методы
Для определения демографической структуры и пространственного распределения особей адвентивных колоний использовали метод пробных площадок по схеме, предложенной Крамаренко с соавторами (2014). В пределах обследуемой территории выбирали относительно однородный участок, на котором пробные площадки располагались в виде регулярной сетки: 8 трансект по 20 пробных площадок. Расстояние между площадками составляло 1.5 м. Сбор моллюсков осуществлялся в пределах каждой пробной площадки площадью 0.25 м2 (рис. 1).
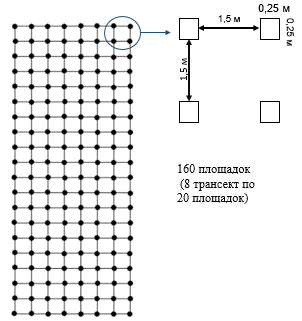
Рис. 1. Схема расположения пробных площадок
Fig. 1. Scheme of the location of sample plots
Абсолютную плотность (D) особей на участке определяли как среднее арифметическое зарегистрированных в границах каждой пробной площадки (xi) организмов данного вида (Крамаренко, 1995):
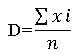
Параллельно с определением плотности популяций изучали их возрастную структуру. Особи B. cylindrica и X. derbentina были разделены на два возрастных класса: ювенильные и адуальные. Адуальными считались половозрелые особи со сформированным отворотом устья.
Для оценки пространственной структуры популяций изучаемых видов использовали глобальный индекс Морана, который рассчитывался по формуле (Anselin, 1995; Крамаренко и др., 2014):
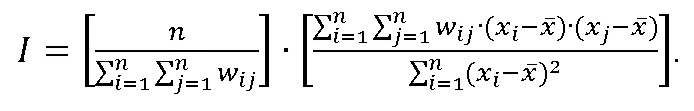
где n – число использованных площадок; xi – число особей в пределах i-й площадки; xj – число особей в пределах j-й пробной площадки; ![]() – среднее значение количества особей по всей выборке; wij – дистанция («вес») между i и j пробными площадками в пространстве.
– среднее значение количества особей по всей выборке; wij – дистанция («вес») между i и j пробными площадками в пространстве.
Для статистической обработки демографических данных использовали стандартный пакет программы R [R CoreTeam, 2016]. Расчет индекса Морана произведен в программе ArcGIS 10.2.
Результаты
Результаты полевых наблюдений представлены на рис. 2.
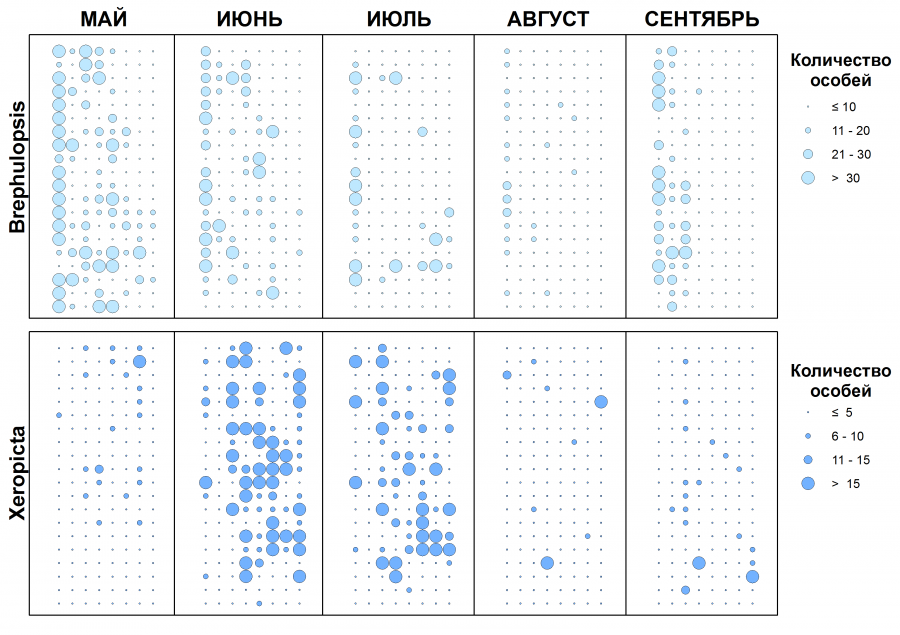
Рис. 2. Ряды наблюдений численности особей на пробных площадках в колониях B. cylindrica и X. derbentina в течение периода с мая по сентябрь
Fig. 2. Observation series of the number of individuals on sample plots in the colonies of B. cylindrica and X. derbentina from May to September
За исследуемый период отмечено изменение численности особей и демографического состава колоний B. cylindrica и X. derbentina в разные месяцы (табл. 1). Для B. cylindrica во все месяцы характерно преобладание особей новой генерации. Наиболее высокие показатели плотности с преобладанием ювенильных особей отмечены в мае и июне. Колония другого адвентивного моллюска, X. derbentina, имеет умеренную численность. В августе – сентябре отмечено снижение численности и увеличение доли половозрелых особей.
Таблица 1. Показатели плотности популяции и частота размерно-возрастных классов B. cylindrica и X. derbentina
| Месяц | B. cylindrica | X. derbentina | ||||
| D, особей/м2 | ювенильные | адуальные | D, особей/м2 | ювенильные | адуальные | |
| Май | 250.3 | 0.925 | 0.074 | 41.2 | 0.958 | 0.041 |
| Июнь | 149.4 | 0.877 | 0.123 | 136 | 0.979 | 0.021 |
| Июль | 85.5 | 0.637 | 0.353 | 110.9 | 0.950 | 0.05 |
| Август | 78.6 | 0.517 | 0.483 | 23.6 | 0.703 | 0.297 |
| Сентябрь | 145 | 0.761 | 0.239 | 27.9 | 0.502 | 0.498 |
Исследование пространственной структуры показало, что белгородские колонии B. cylindrica и X. derbentina имеют преимущественно агрегированный тип распределения (табл. 2). В начале и середине лета оба вида демонстрируют агрегированное распределение. Далее наблюдаются некоторые отличия. B. cylindrica в течение всего периода активности формирует скопления. У второго вида агрегированные тип распределения наблюдается до начала осени, а в конце сезона особи распределены случайно.
Таблица 2. Оценки локального индекса Морана (IM) в популяциях изучаемых видов
| Месяц | Показатели | B. cylindrica | X. derbentina |
| Май | IM | 0.27 | 0.11 |
| ожидаемое значение критерия, E [IM] | -0.006 | -0.006 | |
| критерий значимости z | 6.42 | 2.67 | |
| р-значение | 0 | 0.01 | |
| тип пространственного распределения | агрегированный | агрегированный | |
| Июнь | IM | 0.11 | 0.16 |
| E [IM] | -0.006 | -0.006 | |
| критерий значимости z | 2.65 | 3.87 | |
| р-значение | 0.01 | 0.0001 | |
| тип пространственного распределения | агрегированный | агрегированный | |
| Июль | IM | 0.13 | 0.15 |
| E [IM] | -0.006 | -0.006 | |
| критерий значимости z | 3.21 | 3.67 | |
| р-значение | 0.001 | 0.0002 | |
| тип пространственного распределения | агрегированный | агрегированный | |
| Август | IM | 0.28 | -0.002 |
| E [IM] | -0.006 | -0.006 | |
| критерий значимости z | 6.77 | 0.12 | |
| р-значение | 0 | 0.91 | |
| тип пространственного распределения | агрегированный | случайный | |
| Сентябрь | IM | 0.4 | 0.04 |
| E [IM] | -0.006 | -0.006 | |
| критерий значимости z | 10.37 | 1.05 | |
| р-значение | 0 | 0.29 | |
| тип пространственного распределения | агрегированный | случайный |
Обсуждение
Известно, что в начале лета для B. cylindrica и родственного ему вида B. bidens характерно появление новой генерации (Крамаренко, Попов, 1993). В середине лета мы наблюдали значительное снижение численности колонии B. cylindrica, что объясняется изменением условий среды, вызванных наступлением сухого и жаркого сезона в летние месяцы, когда большое количество особей, как правило, гибнет (Крамаренко, 1995). Кроме того, в летний период моллюски данного вида зарываются в грунт, что затрудняет их учет (Крамаренко, 1997). Причем такая форма адаптивного поведения более свойственна неполовозрелым особям (Крамаренко, 1997), что обуславливает некоторое преобладание ювенильных моллюсков в колонии B. cylindrica после выхода из летнего периода покоя. Снижение численности популяций в конце сезона активности отмечается и в пределах естественного ареала. Так, в Крыму зарегистрированы минимальные значения численности популяции B. cylindrica в сентябре – 40 особей/м2 (Крамаренко, 1997).
Для X. derbentina в пределах нативного ареала характерен однолетний жизненный цикл, при котором новая генерация появляется весной, а к началу осени моллюски достигают репродуктивной зрелости (Popov, Dragomaschenko, 1997; Крамаренко, 2002). Известно, что в некоторых инвазивных популяциях X. derbentina моллюски перестраиваются на двулетний жизненный цикл (Aubry et al., 2005). Однако для белгородской колонии данного вида, как и для крымских популяций, характерен однолетний жизненный цикл. Также на территории, заселенной адвентивной колонией X. derbentina, обнаружено большое количество пустых раковин половозрелых особей. По всей видимости, до следующего сезона доживает небольшое количество адуальных особей, которые гибнут в конце весны – начале лета.
Исследование пространственной структуры белгородских колоний во многом подтверждает известные для изучаемых видов закономерности. Для B. cylindrica аналогичные результаты оценки пространственной структуры были получены в крымских популяциях (Крамаренко и др., 2014). Считается, что формированию агрегаций ксерофильных моллюсков способствуют условия среды. Температура внутри конгломерата улиток на несколько градусов ниже температуры внешней среды (Шилейко, 2012). Кроме того, агрегированное распределение чаще характерно для популяций моллюсков с высокой численностью и плотностью (Крамаренко и др., 2014). Случайное распределение особей в колонии X. derbentina в конце сезона активности, вероятно, можно объяснить завершением летнего периода с высокими температурами, а также снижением численности (см. табл. 1).
Важно также отметить, что на исследуемом участке, занятым B. cylindrica и X. derbentina, практически отсутствует другой чужеродный моллюск − Stenomphalia ravergiensis, колонии которого довольно многочисленны в других районах г. Белгорода. Кроме того, этот вид присутствует и на некотором удалении от исследуемого участка (Снегин, Адамова, 2016). Ранее в пределах пробных площадок нами также был проведен анализ пространственной структуры колонии S. ravergiensis, который показал, что особи распределены случайно (IM = [-0.02; 0.06] при р > 0.05) и на многих пробных площадках вид отсутствует. Возможно, кавказская улитка не выдерживает конкуренции с двумя другими видами-вселенцами. В то же время на данный момент не отмечено распространение B. cylindrica и X. derbentina на территорию, занятую S. ravergiensis.
Можно предположить, что B. cylindrica и X. derbentina в настоящее время доминируют на локальном участке и характеризуются высоким репродуктивным потенциалом, что соответствует одной из стадий инвазионного процесса (Williamson, Fitter, 1996; Colautti, MacIsaac, 2004).
Заключение
Таким образом, на момент исследования численность адвентивных колоний B. cylindrica и X. derbentina находилась на достаточно высоком уровне, что характерно для многих адвентивных видов на стадии колонизации. Новые генерации изучаемых видов появляются в конце весны, максимальное количество ювенильных особей отмечено в начале лета. Пространственная структура белгородских колоний B. cylindrica и X. derbentina представляет собой агрегации, что отражает адаптации к аридным условиям естественного ареала.
Библиография
Крамаренко С. С. Некоторые методы популяционной биологии наземных моллюсков . Николаев: Волтрон, 1995. 40 с.
Крамаренко С. С. Некоторые аспекты экологии наземных моллюсков Brephulopsis cylindrica (Gastropoda; Buliminidae) // Вестник зоологии. 1997. Т. 31. № 4. С. 51–54.
Крамаренко С. С. Особенности скрещивания и репродукции наземных моллюсков Xeropicta derbentina (Pulmonata; Hygromiidae) на северной границе ареала // Вестник зоологии. 2002. Т. 36. № 5. С. 55–60.
Крамаренко С. С. Формирование паттернов пространственно-временной изменчивости наземных моллюсков: мультимасштабный подход: Дис. … д-ра биол. наук . Киев, 2014. 446 с.
Крамаренко С. С., Кунах О. Н., Жуков А. В., Андрусевич Е. В. Анализ паттернов пространственной организации популяций наземных моллюсков: подход с использованием методов геостатистики // Бюллетень Дальневосточного малакологического общества. 2014. № 18. С. 5–40.
Крамаренко С. С., Попов В. Н. Изменчивость морфологических признаков наземных моллюсков рода Brephulopsis Lindhom, 1925 (Gastropoda, Pulmonata, Buliminidae) в зоне интрогрессивной гибридизации // Журнал общей биологии. 1993. Т. 54. № 6. С. 682–690.
Снегин Э. А., Адамова В. В. Анализ демографических и морфологических показателей популяций адвентивного моллюска Stenomphalia ravergiensis (Mollusca, Gastropoda, Pulmonata) в условиях юга Среднерусской возвышенности // Вестник Томского государственного университета. Биология. 2016. № 3 (35). С. 149–160.
Снегин Э. А., Адамова В. В., Сычев А. А. Морфо-генетическая изменчивость нативных и адвентивных популяций моллюска Brephulopsis cylindrica (Gastropoda, Pulmonata, Enidae) // Ruthenica. 2017. Т. 27. № 3. С. 119–132.
Снегин Э. А., Артемчук О. Ю., Сычев А. А., Адамова В. В. Адвентивные виды наземных моллюсков на юге Среднерусской возвышенности // Биоразнообразие и устойчивость живых систем: Материалы XIII Междунар. экологической конференции / Под ред. А. В. Присного. Белгород: ИД «Белгород» НИУ БелГУ, 2014. С. 156–157.
Шилейко А. А. Наземные моллюски надсемейства Helicoidea. Фауна СССР. Моллюски. Нов. сер. Т. 3. Вып. 6 . Л.: Наука, 1978. 384 с.
Шилейко А. А. Наземные моллюски подотряда Pupillina фауны СССР (Gastropoda, Pulmonata,Geophila). Фауна СССР, Моллюски. Т. 3. Вып. 3 . Л.: Наука, 1984. 339 с.
Шилейко А. А. Разнообразие наземных моллюсков, их история и адаптация к жизни на суше // Научный журнал Павлодарского государственного педагогического института. Биологические науки Казахстана. 2012. № 4. С. 44–66.
Anselin L. Local indicators of spatial association – LISA // Geographical analysis. 1995. Vol. 27. № 2. P. 93–115.
Aubry S., Labaune C., Magnin F., Kiss L. Habitat and integration within indigenous communities of Xeropicta derbentina (Gastropoda: Hygromiidae) a recently introduced land snail in south‐eastern France // Diversity and Distributions. 2005. Vol. 11. № 6. P. 539–547.
Colautti R. I., MacIsaac H. J. A neutral terminology to define ‘invasive’species // Diversity and distributions. 2004. Vol. 10. № 2. P. 135–141.
Popov V. N., Dragomaschenko L. A. Aspect of the life cycle of land snails of the genus Xeropicta Monterosato, 1892 in Crimea (Pulmonata: Hygromiinae) // Heldia. 1997. Vol. 4. P. 263–266.
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL: https:// www.R-project.org/ (дата обращения: 06.12.2017).
Williamson M. H., Fitter A. The characters of successful invaders // Biological conservation. 1996. Vol. 78. № 1–2. P. 163–170.
Благодарности
Исследование выполнено при финансовой поддержке РФФИ в рамках научного проекта № 18-34-00010.
Выражаем благодарность С. С. Крамаренко за консультацию и ценные советы, а также П. А. Украинскому за помощь в проведении теоретических расчетов.

 © 2011 - 2026
© 2011 - 2026