



Введение
В настоящее время на земной поверхности практически не осталось территорий и акваторий, не затронутых деятельностью человека. В существующих классификациях современных ландшафтов выделяются антропогенные ландшафты, структура и возникновение которых связаны с деятельностью человека (Мильков, 1973). Водные объекты искусственного происхождения (пруды, обводненные карьеры, каналы, водохранилища и др.) характеризуются особыми гидролого-гидрохимическими условиями и структурой сообществ (Гидротехнические сооружения, 1978; Константинов, 1986).
Наиболее крупные искусственные водоемы (каналы и водохранилища) были созданы для целей судоходства и энергетики. При этом большинство водохранилищ образовались в результате зарегулирования русел существующих рек, что во многом определяет динамику их сообществ (Пидгайко, 1984). Каналы же являются водопроводящими сооружениями и имеют искусственные русла. Специфика формирования биоценозов каналов обусловлена их происхождением и особенностями сообществ водных объектов, которые они соединяют. По своим характеристикам сообщества каналов занимают промежуточное положение между типичными озерными и речными ценозами (Zhao et al., 2018).
На территории Вологодской области наиболее крупными каналами являются Белозерский обводной канал и каналы Северо-Двинской водной системы. Гидробионты этих водных объектов до настоящего времени остаются практически неизученными. Проводимые ранее исследования зоопланктона водных объектов Северо-Двинской шлюзованной системы были фрагментарными и ограничивались лишь озерами (Ривьер, 1978, 1982, 1988, 1992, 2012 и др.; Родионова, 1988).
Цель данной работы – выявление особенностей зоопланктона каналов Северо-Двинской шлюзованной системы как типичных антропогенных местообитаний.
Материалы
Исследования проводились на 6 каналах Северо-Двинской шлюзованной системы в июле 2010 г. Северо-Двинская водная система располагается в центральной части Вологодской области на территории Национального парка «Русский Север» (рис. 1). Построенная в 1825–1828 гг., она соединяет реки Шексна и Сухона, тем самым связывая бассейны рек Волги и Северной Двины. По своему экономическому значению Северо-Двинская система значительно уступает Волго-Балтийской. В последнее время увеличивается ее значение как туристического маршрута.
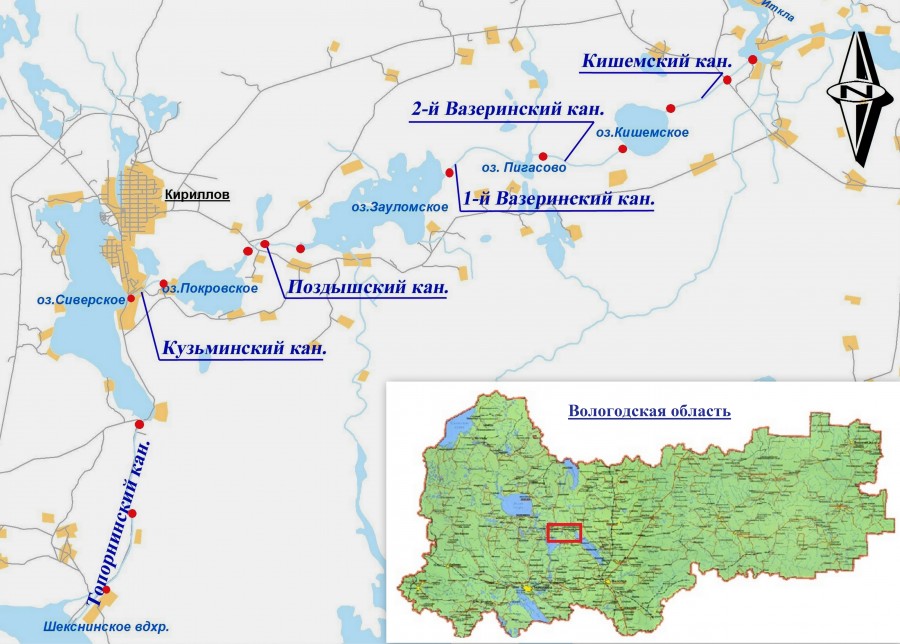
Рис. 1. Картосхема Северо-Двинской шлюзованной системы (красным пуансоном обозначены места отбора гидробиологических проб)
Fig. 1. Schematic map of Severo-Dvinsky water system (hydrological sampling sites are denoted by red punch)
В настоящее время Северо-Двинская водная система состоит из цепочки малых озер, соединенных каналами. Озера водной системы являются типичными ледниковыми водоемами. За исключением озера Сиверское все водоемы мелководны, характеризуются выраженной зарослевой зоной, занимающей до 50 % акватории (Антипов и др., 1981; Краснова, 1999). Озеро Сиверское является самым глубоким из озер водной системы (максимальная глубина 25 м), отличается выраженной температурной стратификацией и небольшим развитием макрофитов, которыми занято 5–10 % площади озера. В отличие от озер, каналы характеризуются сходными морфометрическими параметрами, что обусловлено использованием их ранее как транспортных путей (табл. 1). Для всех каналов свойственны особый волновой режим, повышенная мутность воды и отсутствие выраженных зарослей макрофитов.
Таблица 1. Основные морфометрические характеристики каналов Северо-Двинской шлюзованной системы
| № | Наименование канала | Объекты, которые соединяет канал | Длина, км | Средняя ширина, м | Глубина, м |
| 1 | Топорнинский | Шекснинское водохранилище и озеро Сиверское | 6.6 | 45.0 | 3.00 |
| 2 | Кузьминский | Озера Сиверское и Покровское | 1.3 | 40.7 | 3.00 |
| 3 | Поздышский (р. Поздышка) | Озера Покровское и Зауломское | 2.7 | 65.0 | 3.00 |
| 4 | 1-й Вазеринский | Озера Зауломское и Пигасово | 2.3 | 39.5 | 2.94 |
| 5 | 2-й Вазеринский | Озера Пигасово и Кишемское | 2.8 | 42.5 | 2.94 |
| 6 | Кишемский | Озеро Кишемское и реку Иткла | 2.7 | 38.5 | 3.00 |
Методы
Отбор проб зоопланктона проводили в июле 2010 г. Сбор осуществлялся малой сетью Джеди (ячея 74 мкм). Всего было отобрано 45 проб. Станции отбора проб были распределены на всем протяжении каналов (см. рис. 1). Для уточнения видового состава зоопланктона проводился отбор проб на участках, занятых макрофитами. Пробы фиксировали 4%-ным формалином. Камеральная обработка проб проводилась в камере Богорова в соответствии с общепринятыми в гидробиологии методами с использованием современных определителей (Методика изучения…, 1975; Определитель зоопланктона…, 2010). В составе зоопланктона учитывали и определяли видовую принадлежность коловраток (Rotifera) и ракообразных (Cladocera, Copepoda). Для каждого организма определяли общую длину тела. Биомасса зоопланктеров рассчитывалась согласно уравнениям связи индивидуальной массы организмов с длиной тела (Балушкина, Винберг, 1979).
В рамках анализа оценивали среднюю численность и биомассу видов и групп, выделяли доминирующий комплекс видов (относительная численность более 5 %), рассчитывали индексы видового разнообразия Шеннона – Уивера (по численности и биомассе зоопланктеров) и фаунистического сходства Чекановского – Серенсена (Песенко, 1982), среднюю индивидуальную массу зоопланктеров (Крючкова, 1987). Для оценки степени варьирования показателей в расчетах использовали соответствующие результаты обработки нескольких гидробиологических проб по каждому изученному водному объекту. При сравнительном анализе использованы опубликованные данные (Гордеева и др., 1978; Ривьер, 1978) и собственные неопубликованные материалы о зоопланктоне малых озер Северо-Двинской водной системы.
Результаты
В составе зоопланктона каналов Северо-Двинской системы обнаружено 33 вида, из них коловраток – 11, кладоцер – 15, копепод – 7. Видовое богатство зоопланктона отдельных каналов колебалось от 17 (Кишемский) до 28 (Топорнинский) видов (табл. 2). Особенностью зоопланктона каналов является присутствие в его составе холодолюбивого Cyclops scutifer, характерного и для сообществ малых озер водной системы (Ривьер, 1982). Различия видового богатства связаны преимущественно с разной представленностью в составе зоопланктона кладоцер, в частности видов из семейств Sididae и Chydoridae.
Таблица 2. Таксономический состав зоопланктона водных объектов Северо-Двинской шлюзованной системы
| Таксон | Каналы* | Озера водной системы | |||||
| 1 | 2 | 3 | 4 | 5 | 6 | ||
| Тип Rotifera Cuvier, 1817 | |||||||
| Сем. Philodinidae Bryce, 1910 | |||||||
| Dissotrocha sp. | – | – | – | – | + | – | – |
| Rotaria sp. | + | – | – | – | – | – | + |
| Сем. Euchlanidae Ehrenberg, 1838 | |||||||
| Euchlanis sp. | + | – | – | + | – | – | + |
| Сем. Brachionidae Ehrenberg, 1838 | |||||||
| Brachionus diversicornis (Daday, 1883) | – | + | + | – | – | – | – |
| Keratella quadrata (Müller, 1786) | + | + | + | – | – | – | +д |
| K. cochlearis (Gosse, 1851) | – | – | – | – | – | – | +д |
| Kellicottia longispina (Kellicott, 1879) | – | – | – | – | – | – | + |
| Сем. Mytilinidae Harring, 1913 | |||||||
| Mytillina sp. | + | – | – | – | – | – | – |
| Сем. Asplanchidae Eckstein, 1883 | |||||||
| Asplanchna priodonta Gosse, 1850 | +д | + | +д | + | +д | +д | + |
| Сем. Trichocercidae Harring, 1913 | |||||||
| Trichocerca sp. | – | + | + | + | + | + | + |
| Сем. Synchaetidae Hudson and Gosse, 1886 | |||||||
| Polyarthra euryptera Wierzejski, 1891 | – | – | – | – | – | – | + |
| Polyarthra sp. | + | + | + | + | + | + | + |
| Сем. Conochilidae Harring, 1913 | |||||||
| Conochilus sp. | – | – | + | + | – | + | + |
| Сем. Filiniidae Harring and Myers | |||||||
| Filinia longiseta (Ehrenberg, 1834) | + | + | +д | + | +д | + | + |
| Сем. Lecanidae Lemane, 1933 | |||||||
| Lecane sp. | – | – | – | – | – | – | + |
| Тип Arthropoda Latreille, 1829 | |||||||
| Надотряд Cladocera Latreille, 1829 | |||||||
| Сем. Sididae Baird, 1850 | |||||||
| Sida crystallina crystallina (O. F. Müller, 1776) | + | – | – | + | – | + | + |
| Diaphanosoma brachyurum Lievin, 1848 | + | + | – | + | – | + | + |
| Сем. Daphniidae Straus, 1820 | |||||||
| Daphnia сristata Sars, 1862 | + | + | + | + | + | + | + |
| D. сucullata Sars, 1862 | + | + | + | + | + | + | + |
| D. longispina O. F. Müller, 1785 | – | – | – | – | – | – | + |
| D. longiremis Sars, 1862 | – | – | – | – | – | – | + |
| Ceriodaphnia quadrangula (O. F. Müller, 1785) | + | + | + | – | + | + | + |
| Scapholeberis mucronata (O. F. Müller, 1776) | – | – | – | – | – | – | + |
| Сем. Bosminidae Sars, 1865 | |||||||
| Bosmina (Eubosmina) cf. coregoni Baird, 1857 | + | + | – | – | – | – | + |
| Bosmina (E.) cf. longispina Leydig, 1860 | – | – | – | – | – | – | + |
| B. (Bosmina) longirostris (O. F. Müller, 1785) | +д | + | + | + | + | + | +д |
| Сем. Chydoridae Dybowski et Grochowski, 1894 | |||||||
| Acroperus harpae (Baird, 1834) | + | – | – | + | + | – | + |
| Alona affinis (Leydig, 1860) | – | – | – | – | – | – | + |
| A. quadranqularis (O. F. Müller, 1785) | – | – | – | – | – | – | + |
| A. rectangula Sars, 1862 | – | – | – | – | – | – | + |
| Camptocercus lilljeborgi Schoedler, 1862 | – | – | – | + | + | – | + |
| Chydorus sphaericus (O. F. Müller, 1785) | + | + | + | +д | + | + | +д |
| Disparalona rostrata (Koch, 1841) | + | + | – | + | + | – | + |
| Graptoleberis testudinaria (Fischer, 1851) | + | – | – | – | + | – | + |
| Pleuroxus trigonellus (O. F. Müller, 1785) | + | – | – | – | – | – | – |
| Pleuroxus sp. | + | – | – | – | – | – | – |
| Сем. Eurycercidae Kurz, 1875 | |||||||
| Eurycercus (Eurycercus) lamellatus (O. F. Müller, 1776) | – | – | – | – | – | – | + |
| Сем. Polyphemidae Baird, 1845 | |||||||
| Polyphemus pediculus (Linnaeus, 1761) | – | – | – | – | – | – | + |
| Cем. Leptodoridae Lilljeborg, 1861 | |||||||
| Leptodora kindtii (Focke, 1844) | + | + | + | + | + | + | + |
| Подкласс Copepoda Milne-Edwards, 1840 | |||||||
| Cем. Diaptomidae G.O. Sars, 1903 | |||||||
| Eudiaptomus gracilis (Sars, 1863) | + | + | + | + | + | + | + |
| E. graciloides (Lilljeborg, 1888) | – | – | – | – | – | – | + |
| Сем. Temoridae Sars, 1902 | |||||||
| Heterocope appendiculata (Sars, 1863) | – | + | – | + | – | – | + |
| Ceм. Cyclopidae Dana, 1846 | |||||||
| Diacyclops bicuspidatus (Claus, 1857) | + | – | – | – | + | ||
| Thermocyclops oithonoides (Sars, 1863) | – | – | – | – | – | – | + |
| T. crassus (Fischer, 1853) | – | – | – | – | – | – | + |
| Mesocyclops leuckarti (Claus, 1857) | +д | + | + | + | +д | ||
| Paracyclops affinis (Sars, 1863) | +д | +д | +д | – | – | +д | + |
| Cyclops scutifer Sars, 1863 | +д | +д | +д | +д | +д | +д | +д |
| Cyclops strenuus Fischer, 1851 | – | – | – | – | – | – | + |
| Megacyclops viridis (Jurine, 1820) | – | – | – | – | – | – | + |
| Eucyclops serrulatus (Fischer, 1851) | – | – | – | – | – | – | + |
| Ectocyclops phaleratus (Koch, 1838) | – | – | – | – | – | – | + |
| Cyclops sp. | + | + | – | + | – | – | – |
| Harpactiformes | + | – | – | – | – | – | + |
| Общее число видов, в том числе: | 28 | 21 | 17 | 22 | 19 | 17 | 48 |
| Rotifera | 7 | 6 | 7 | 6 | 5 | 5 | 12 |
| Cladocera | 14 | 9 | 6 | 10 | 10 | 8 | 22 |
| Copepoda | 7 | 6 | 4 | 6 | 4 | 4 | 14 |
Примечание. «–» – вид не обнаружен, «+» – вид обнаружен, «+д» – доминирующий вид; * – обозначения каналов аналогичны табл. 1.
Изученные планктонные сообщества каналов характеризуются значительной выравненностью (табл. 3). Значения индекса видового разнообразия Шеннона – Уивера, рассчитанные по численности зоопланктона в каналах, колебались от 1.8 ± 0.19 (2-й Вазеринский) до 3.2 ± 0.18 (Топорнинский), по биомассе – от 2.3 ± 0.18 (Кузьминский) до 3.1 ± 0.29 (1-й Вазеринский), что соответствует показателям в олиго-, мезотрофных водоемах (Андроникова, 1996).
Средняя численность зоопланктона каналов Северо-Двинской системы составила 110.2 ± 11.82 тыс. экз./м3, биомасса – 0.6 ± 0.09 г/м3. Наибольшие величины этих характеристик отмечены в небольшом Поздышском канале (рис. 2). Доминантами по величинам численности и биомассы во всех изученных каналах являлись Copepoda (см. рис. 2). Они составляли от 68 (Кишемский) до 85 % (2-й Вазеринский) общей численности и от 67 (1-й Вазеринский и Кишемский) до 89 % (Кузьминский) общей биомассы зоопланктона.
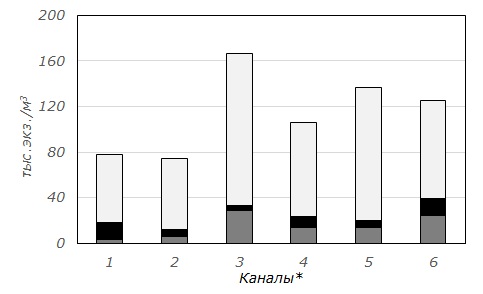
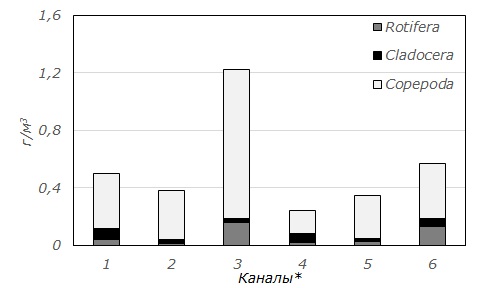
Рис. 2. Средние численность (А) и биомасса (Б) зоопланктона каналов Северо-Двинской шлюзованной системы (обозначения каналов аналогично табл. 1)
Fig. 2. The average abundance (A) and biomass (Б) of zooplankton of canals of Severo-Dvinsky water system ( canals are indicated as in Table 1)
Доминирующий комплекс зоопланктона во всех каналах сходен и включает 2–3 вида. Исключением является лишь Топорнинский канал, где состав доминантов более разнообразен. В число доминантов входят преимущественно копеподы – Cyclops scutifer, Paracyclops affinis. В отдельных каналах (Топорнинский, Поздышский, 2-й Вазеринский, Кишемский) доминантами являются и коловратки – Asplanchna priodonta и Filinia longiseta. Хищная Asplanchna priodonta составляет от 5 (Топорнинский) до 21 % (Кишемский) общей биомассы зоопланктона, относительная численность Filinia longiseta достигает во 2-м Вазеринском и Поздышском каналах 6 и 8 % соответственно. Среди кладоцер относительно высоких численности и биомассы лишь в Топорнинском канале достигает Bosmina longirostris. Доля этого рачка в общей средней численности зоопланктона составляет 7 %, а в биомассе – 2 %. Особенностью зоопланктеров каналов являются малые размеры, что и обуславливает небольшие величины общей биомассы (табл. 3). Так, средняя длина половозрелых особей доминантов (Cyclops scutifer и Paracyclops affinis) не превышает 0.6–0.7 мм. Кроме того, существенную долю численности этих видов (до 10–15 % в пробе) составляют копеподиты.
Таблица 3. Показатели зоопланктона водных объектов Северо-Двинской шлюзованной системы
| Показатель | Канал* | Озера водной системы | |||||
| 1 | 2 | 3 | 4 | 5 | 6 | ||
| Индекс Шеннона – Уивера (бит/экз.) | 3.2 | 2.8 | 3.1 | 2.1 | 1.8 | 3.2 | 2.8 |
| Индекс Шеннона – Уивера (бит/г) | 2.9 | 2.3 | 2.5 | 3.1 | 2.4 | 2.7 | 2.5 |
| Отношение численностей Cladocera и Copepoda | 0.36 | 0.10 | 0.03 | 0.14 | 0.06 | 0.19 | 0.22 |
| Средняя индивидуальная масса, мг | 0.006 | 0.005 | 0.007 | 0.002 | 0.002 | 0.004 | 0.012 |
Примечание. * – обозначения каналов аналогично табл. 1.
Обсуждение
Зоопланктон каналов Северо-Двинской шлюзованной системы представлен эврибионтными, широко распространенными в регионе организмами. Основу сообществ составляют пелагические и придонные виды. Неразвитость зарослей макрофитов обуславливает малую долю фитофильных организмов. Зарегистрированные фитофильные организмы были представлены единичными особями, вероятнее всего, занесенными в каналы из близлежащих озер. Сходство морфологических характеристик и истории формирования обуславливают значительное сходство таксономического состава зоопланктона всех каналов, что подтверждается величинами индексов Чекановского – Серенсена (более 0.7). Наиболее специфичным, по сравнению с другими каналами, является состав зоопланктона Топорнинского канала. На отдельных участках этого водного объекта сформированы заросли макрофитов, в том числе погруженных, благоприятные для обитания некоторых кладоцер. Кроме того, происходит обогащение фауны этого канала из близлежащих участков Шекснинского водохранилища.
После создания искусственных водных объектов наблюдается их заселение организмами, способными адаптироваться к новым условиям обитания (Caley, Schluter, 1997; Louette et al., 2008; Hadasova, Kopp, 2014). Немногочисленные данные о структуре сообществ каналов Вологодской области свидетельствуют, что видовой состав и структура доминирующего комплекса зоопланктона этих водных объектов определяются водоемами-водоисточниками. Так, для Белозерского обводного канала отмечается доминирование Kellicottia longispina, Bosmina longirostris, Eudiaptomus gracilis, присутствующих в составе комплекса доминантов в озере Белом (Думнич, Лобуничева, 2016). Структура зоопланктона каналов Северного склона Волго-Балтийской водной системы также в значительной степени сходна с таковой в водохранилищах, которые они соединяют (Лобуничева, 2013). Подобные закономерности отмечаются и для других каналов (Гордеева и др., 1978, Akopian et al., 1999).
Заселение каналов Северо-Двинской водной системы происходило в первую очередь организмами из малых озер, которые и являются водоемами-водоисточниками. Для этих озер свойственен богатый зоопланктон, характерный для мезо-эвтрофных водоемов (см. табл. 2). Различия в структуре зоопланктона озер связаны с их морфологическими особенностями. Так, зоопланктон сравнительно глубоководного озера Сиверское, в котором выражен гиполимнион и отмечается самая высокая прозрачность воды (1.5–2 м), характеризуется чертами олиготрофии, в частности низкими величинами биомассы (0.3-0.4 г/м3). Прочие озера мелководны, имеют выраженную зарослевую зону, прозрачность воды в них не превышает 1.5 м. В этих водоемах регистрируются более высокие значения биомассы зоопланктона (оз. Кишемское – 0.4 г/м3, оз. Зауломское – 1.4 г/м3 (Ривьер, 1978; собственные неопубликованные данные)). Структура доминирующего комплекса зоопланктона во всех естественных водоемах водной системы сходна.
Доминирующей группой организмов во всех каналах являлись веслоногие ракообразные. Преобладание циклопов среди зоопланктеров каналов непосредственно связано с особенностями формирования их сообществ. В малых озерах Северо-Двинской системы основу зоопланктона составляют преимущественно циклопы, что связано с повышенной мутностью воды (Ривьер, 1978, 1982). Кроме того, отсутствие в каналах мелководной прибрежной зоны, занятой макрофитами, не позволяет (даже в наиболее теплый летний период) массово развиваться многим видам кладоцер, характерным для большинства водных объектов региона.
Численность и биомасса планктонных животных изученных каналов несколько выше, чем в сходных по происхождению водных объектах, а также в малых реках региона. Согласно собственным исследованиям, средняя биомасса зоопланктона в Белозерском обводном канале и каналах Северного склона Волго-Балтийской водной системы в летний период равна 0.2 и 0.4 г/м3 соответственно. По-видимому, некоторое увеличение уровня развития зоопланктона в каналах Северо-Двинской шлюзованной системы связано с более выраженным озерным режимом. Численность и биомасса зоопланктона в каналах, имеющих водоисточниками реки, значительно ниже (Гордеева и др., 1978, Akopian et al., 1999). Для водоемов северного склона Беломорско-Балтийского канала указывается, что средняя биомасса зоопланктона озерной части трассы канала составляет 0.4 г/м3, тогда как в речной части – лишь 0.1 г/м3 (Гордеева и др., 1978).
Заключение
Всего в составе зоопланктона каналов Северо-Двинской шлюзованной системы обнаружено 33 вида (Rotifera – 11, Cladocera – 15, Copepoda – 7). Большинство видов являются эврибионтными. Таксономический состав зоопланктона каналов сходен (коэффициент Чекановского – Серенсена более 0.7). Первоначально заселение этих водных объектов происходило за счет организмов из малых озер и Шекснинского водохранилища, с которыми каналы непосредственно соединены. В настоящее время также происходит проникновение планктонных животных в каналы. Однако морфологические особенности каналов (упрощенный рельеф дна, отсутствие выраженной мелководной зоны) и волновое воздействие определяют структуру формирующихся в них сообществ. Практически полное отсутствие зарослей высших водных растений неблагоприятно для развития многих зоопланктеров. В результате зоопланктон каналов характеризуется низким видовым богатством с преобладанием пелагических и придонных видов.
По сравнению с малыми озерами зоопланктон каналов характеризуется невысокими численностью и биомассой, доминированием небольшого числа видов, преобладанием особей мелких размеров. При этом структура доминирующего комплекса сходна. При сравнительно невысоких величинах численности и биомассы основу зоопланктона составляют веслоногие ракообразные. Выявленные особенности зоопланктона каналов непосредственно связаны с их антропогенным происхождением и эксплуатацией как транспортных путей. Специфические морфологические и гидрологические характеристики этих водных объектов обуславливают упрощенную структуру и низкий уровень развития зоопланктона.
Библиография
Андроникова И. Н. Структурно-функциональная организация зоопланктона озерных экосистем: Монография . СПб.: Наука, 1996. 189 с.
Антипов Н. П., Жаков Л. А., Лебедев В. Г., Шевелев Н. Н. Озера ландшафтов холмисто-моренных равнин // Озерные ресурсы. Вологда, 1981. С. 38–93.
Балушкина Е. В., Винберг Г. Г. Зависимость между длиной и массой тела планктонных ракообразных // Экспериментальные и полевые исследования биологических основ продуктивности озер. Л.: ЗИН АН СССР, 1979. С. 58–79.
Гидротехнические сооружения: Учебное пособие для вузов . М.: Стройиздат, 1978. 647 с.
Гордеева Л. И., Соколова В. А., Макаров В. П. Гидробиологический режим Беломорско-Балтийского канала // Гидробиология Выгозерского водохранилища. Петрозаводск, 1978. С. 134–156.
Думнич Н. В., Лобуничева Е. В. Динамика зоопланктона озерной части Шекснинского водохранилища (Вологодская область) // Современное состояние биоресурсов внутренних водоемов и пути их рационального использования: Материалы докладов Всерос. конф. с междунар. участием, посвящ. 85-летию Татарского отделения ГосНИОРХ (Казань, 24–29 октября 2016 г.). Казань, 2016. С. 338–349.
Константинов А. С. Общая гидробиология: Учеб. для студентов биол. спец. вузов . М.: Высшая школа, 1986. 472 с.
Краснова А. Н. Структура гидрофильной флоры техногенно трансформированных водоемов Северо-Двинской водной системы . Рыбинск: ОАО «Рыбинский Дом печати», 1999. 200 с.
Крючкова Н. М. Структура сообществ зоопланктона в водоемах разного типа // Продукционно-гидробиологические исследования водных экосистем (Тр. ЗИН АН СССР). Л., 1987. Т. 165. С. 184–198.
Лобуничева Е. В. Зоопланктон водохранилищ северного склона Волго-Балтийского водного пути // Биология внутренних вод: Материалы XV Школы-конференции молодых ученых (Борок, 19–24 октября 2013 г.). Кострома, 2013. С. 246–250.
Методика изучения биогеоценозов внутренних водоемов . М.: Наука, 1975. 240 с.
Мильков Ф. Н. Человек и ландшафты: очерки антропогенного ландшафтоведения . М.: Мысль, 1973. 224 с.
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 1. Зоопланктон . М.: Товарищество научных изданий КМК, 2010. 495 с.
Песенко Ю. А. Принципы и методы количественного анализа в фаунистических исследованиях . М.: Наука, 1982. 287 с.
Пидгайко М. Л. Зоопланктон водоемов Европейской части СССР . М.: Наука, 1984. 207 с.
Ривьер И. К. Зоопланктон водоемов трассы переброски северных рек // Биологические ресурсы водоемов Вологодской области, их охрана и рациональное использование: Тезисы к науч.-практ. конф. Вологда, 1978. С. 26–28.
Ривьер И. К. Особенности функционирования зоопланктонных сообществ водоемов различных типов // Структура и функционирование пресноводных экосистем. Л.: Наука, 1988. С. 80–111.
Ривьер И. К. Современное состояние зоопланктона водоемов Волго-Балтийской и Северо-Двинской водных систем // Труды ИБВВ АН. Вып. 43 (46). Экологические исследования водоемов Волго-Балтийской и Северо-Двинской водных систем. Л.: Наука, 1982. С. 90–103.
Ривьер И. К. Холодноводный зоопланктон озер бассейна Верхней Волги . Ижевск: Издатель Пермяков С. А., 2012. 390 с.
Ривьер И. К. Экология ветвистоусых ракообразных в зимних водоемах // Современные проблемы изучения ветвистоусых ракообразных. СПб.: Гидрометеоиздат, 1992. С. 65–80.
Родионова Н. В. Суточное вертикальное распределение зоопланктона в двух мезотрофных озерах // Интенсификация рыбного хозяйства на внутренних водоемах: Сб. науч. тр. ГосНИОРХ. Вып. 288. Л., 1988. С. 104–105.
Akopian M., Garnier J., Pourriot R. A large reservoir as a source of zooplankton for the river: structure of the populations and influence of fish predation // Journal of Plankton Research. 1999. Vol. 21. P. 285–297.
Caley M. J., Schluter D. The relationship between local and regional diversity // Ecology. 1997. Vol. 78. P. 70–80.
Hadasova L., Kopp R. Monitoring of the initial succession of zooplankton communities in newly created ponds within the Territorial System of Ecological Stability // MendelNet 2014. Proceedings of International PhD Students Conference. Mendel University in Brno, Czech Republic, November 19th and 20th. 2014. P. 245–249.
Louette G., De Meester L., Declerck S. Assembly of zooplankton communities in newly created ponds // Freshw. Biol. 2008. Vol. 53 (11). P. 2309–2320.
Zhao Kun, Wang Lizhu, Riseng Catherine, Wehrly Kevin, Pan Yangdong, Song Kun, Da Liangjun, Pang Wanting, You Qingmin, Tian Hua, Liu Shuiqin, Wang Quanxi. Factors determining zooplankton assemblage difference among a man-made lake, connecting canals, and the water-origin river // Ecological Indicators. 2018. Vol. 84. P. 488–496.
Благодарности
Автор выражает признательность сотрудникам Вологодского филиала ФГБНУ «ВНИРО» за помощь в сборе полевого материала.

 © 2011 - 2026
© 2011 - 2026