



Введение
В период формирования экологического направления в паразитологии В. Н. Беклемишев (1956) привлек внимание к структурообразующей роли паразитов в биоценозах и ввел в научный оборот понятие «паразитарная система». Автор рассматривал паразитарную систему как совокупность популяции паразита вместе со всеми популяциями хозяев, непосредственно поддерживающими ее существование. В наибольшей степени изучены паразитарные системы гельминтов рыб, птиц и млекопитающих (Контримавичус, Атрашкевич, 1982; Фортунато, 1985; Батькаев, Ваккер, 1985, 1988, 1992; Аникиева, 2003; Русинек, 2003; Галактионов, Атрашкевич, 2015). Паразиты земноводных и пресмыкающихся в этом аспекте практически не изучены.
Одним из обычных видов, населяющих тонкий кишечник амфибий и рептилий, является нематода Os. filiformis. Ее ареал простирается от Якутии и Забайкалья на востоке Евразии и до Испании на западе (Однокурцев, Седалищев, 2008; Щепина, Балданова, 2010; Щепина, Дугаров, 2008; Sanchis et al., 2000). Хозяевами нематоды являются: обыкновенный тритон (Triturus vulgaris), серая (Bufo bufo), монгольская (B. raddei) и зеленая (B. viridis) жабы, обыкновенная квакша (Hyla arborea), обыкновенная чесночница (Pelobates fuscus), травяная (Rana temporaria), остромордая (R. arvalis), сибирская (R. amurensis), озерная (Pelophylax ridibundus) и прудовая (P. lessonae) лягушки (Рыжиков и др., 1980). Из пресмыкающихся Os. filiformis установлена у прыткой (Lacerta agilis), полосатой (L. strigiata), зеленой (L. viridis) и живородящей (Zootoca vivipara) ящериц, веретеницы (Anguis fragilis), желтопузика (Pseudopus apodus), долгохвостки амурской (Tachydromus amurensis), обыкновенного (Natrix natrix) и водяного (N. tessellata) ужей, медянки (Coronella austriaca), кошачьей змеи (Telescopus fallax), гадюки обыкновенной (Vipera berus) (Шарпило, 1976). У названных видов амфибий и рептилий Os. filiformis найдена в Западной Сибири (Жигилева и др., 2002; Буракова, 2010; Жигилева, Кирина, 2014; Жигилева, 2017), на Урале и в Зауралье (Буракова, Вершинин, 2016), в Волжском бассейне (Кириллова и др., 2008; Лукиянов и др., 2008; Ручин, Чихляев, 2013), в Черноземье (Равковская и др., 2015) и других регионах России (Радченко, Шабунов, 2008). Os. filiformis констатирована в Белоруссии и Украине (Шарпило, 1976; Рыжиков и др., 1980; Шималов, 2002, 2009; Schimalov et al., 2000, 2001a, b). Данный представитель круглых червей проник в Скандинавию (Кудинова и др., 1985; Новохацкая, 2008б; Cedhagen, 1988) и Ирландию (Griffin, 1989). Os. filiformis встречается у хозяев не только в лесных, лесостепных и степных биотопах, но и в стациях лесо-лугово-степного пояса горных систем, например Карпат (Kozak, 1973; Sharpilo et al., 2001), Альп (Singer, Sattmann, 2007), Анатолийского плоскогорья и Армянского нагорья в Турции (Yildirimhan et al., 2009). На территории Республики Казахстан вид найден у обыкновенной жабы, травяной, остромордой и озерной лягушек и у прыткой ящерицы (Соболева, 1975; Ваккер, 1984; Ваккер и др., 1990; Тарасовская, 2009). Обширный ареал, широкий спектр хозяев делают нематоду Os. filiformis перспективной для изучения паразитарных систем.
Цель представляемой работы – установление наличия и описание паразитарных систем Os. filiformis в степной зоне Казахстана. Для достижения цели решались следующие задачи: 1) исследовали на зараженность Os. filiformis представителей батрахо- и герпетофауны в биотопах степной зоны Казахстана; 2) изучали развитие личинок из яиц, полученных от самок нематоды из разных видов хозяев; 3) анализировали характеристики различных типов паразитарных систем Os. filiformis и установливали роль отдельных видов хозяев; 4) определяли гостальный статус Os. filiformis.
Материалы
В течение 1973–1997 гг. в северо-восточной части степной зоны Республики Казахстан (Павлодарская область) на зараженность Os. filiformis исследованы: остромордая лягушка (2704 экз.), обыкновенная жаба (1 экз.), такырная круглоголовка (Phrynocephalus helioscopus, 3 экз.), разноцветная ящурка (Eremias arguta, 14 экз.), прыткая (344 экз.) и живородящая ящерицы (6 экз.), обыкновенный уж (5 экз.), узорчатый полоз (Elaphe dione, 7 экз.), степная (Vipera ursinii, 13 экз.) и обыкновенная (5 экз.) гадюки, обыкновенный щитомордник (Gloydius halys, 6 экз.). Номера пунктов ручного отлова хозяев приведены на рис. 1, в табл. 1 и 2 или в тексте. На изучаемой территории господствуют три основных типа ландшафтов – ландшафты междуречных равнин (левобережная и правобережная сухая степь), Казахского мелкосопочника (Сарыарка), речных долин и озерных впадин (Чупахин, 1973). Мелкосопочник включает лесной ландшафт гранитных гор и ландшафт сглаженных невысоких сопочных гряд. Кроме названных ландшафтов на северо-востоке представлена колочная лесостепь, на юго-востоке – боровая лесостепь (см. рис. 1). На юге левобережья находится территория, занятая полупустыней. Река Иртыш с ее интразональными ландшафтами прорезает район исследования почти точно с юга на север.
Наибольшее число хозяев собрано в ходе гельминтологического мониторинга остромордой лягушки в 1984–1997 гг. на притеррасной площадке в правобережной пойме р. Иртыш. Подопытная площадка (под названием Усолка) длиной около 500 м и шириной 0.5–7.0 м расположена на правом берегу протоки Иртыша Усолки, в 15 км выше г. Павлодар (52 12' 32.15'' СШ и 77 01' 05.70'' ВД). Берега Усолки покрыты луговым разнотравьем, тростником (Phragmites communis) и аиром (Acorus calamus). Встречаются отдельно стоящие деревья и кустарники. Побережье протоки увлажнено, а местами слегка заболочено за счет многочисленных родников, через которые на неогеновых глинах ложа реки идет выклинивание грунтовых вод. Вверху площадки размещается нерестовый водоем, заросший хвощом (Equisetum arvense), а по берегам – тростником. Внизу находится еще один нерестовый водоем, вокруг которого хозяев не отлавливали. Симпатрично с остромордой лягушкой обитает прыткая ящерица. Убежища последней находятся преимущественно на поверхности первой надпойменной террасы. Однако пресмыкающиеся совершают регулярные кормовые миграции на берег протоки. Относительная численность лягушек и ящериц на подопытной площадке, подсчитанная в июне маршрутным методом, равна 5 : 1.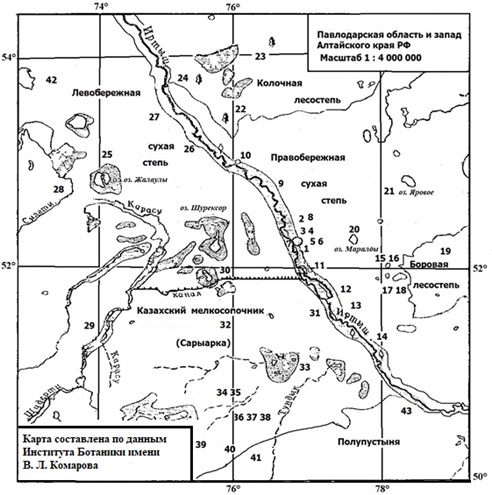
Рис. 1. Карта района исследования. Условные обозначения: боровая лесостепь – сосновые и сосново-березовые остепненные леса; колочная лесостепь – богаторазнотравно-ковыльные степи с березовыми колками; право- и левобережная степь – в северной части с разнотравно-ковыльной, в южной части и мелкосопочнике – типчаково-ковыльной растительностью; полупустыня – с полынно-ковыльными ассоциациями; озерные впадины Жалаулы, Шурексор, Маралды, Малыбай и др. – с лугово-галофитной и галофитной растительностью на солонцах и солончаках; в поймах рек Иртыш, Селеты, Шидерты, Карасу и др. – пойменные луга и кустарники, в пойме р. Иртыш – тополевые леса. Арабскими цифрами обозначены номера выборок хозяев (см. табл. 1 и 2). Границы, отделяющие основные ландшафты, показаны сплошной линией
Fig. 1. Map of the study area. Legend: forest-steppe – pine and pine-birch steppe-heath forests; forest-steppe with kolkis – birch grove with motley and feather-grass -steppe; the motley and feather grass steppe – the fescue and feather grass dry steppe; semi-desert – wormwood and feather grass deserted steppe; landscapes of lake basins – meadow-halophytic and halophytic vegetation on saline soil and salt-marsh; landscapes of river valleys. Arabic numerals indicate the numbers of the hosts samples (see Tables 1 and 2). The boundaries separating the main landscapes are shown by a solid line
Второе место сбора обоих видов хозяев для изучения паразитарной системы Os. filiformis находится в заросшей травой и кустарником долине ручья Кафар в 5 км от места его впадения в оз. Сабандыколь. Пункт расположен в мелкосопочнике в Баянаульском горном поднятии (50° 47' 20'' СШ 75° 41' 44'' ВД) и назван Баянаул. В июне отношение числа лягушек к количеству ящериц на маршруте равнялось 1 : 2. Три исследованных антропогенных биотопа (Яблонька, Карьер и АБС) расположены на территории Большого Павлодара. В двух километрах восточнее Усолки, в зеленом поясе г. Павлодар, находится надпойменный пункт Яблонька. Амфибий в нем отлавливали около заполненного водой понижения рельефа, оставшегося от строительства шоссе. В надпойменной точке с названием Карьер хозяев собирали на берегу озерца, возникшего на месте отработанного песчаного карьера. Биотоп расположен в промышленной зоне города, в 3 км северо-восточнее Яблоньки. Третий антропогенный пункт – АБС – находится рядом с агробиологической станцией Павлодарского государственного педагогического института и представляет собой водоем, образовавшийся в понижении рельефа за счет талых вод и инфильтрационной воды из поливных земель Черноярской агропромышленной фирмы.
Методы
Хранение и вскрытие хозяев, сбор и обработку гельминтов проводили по общепринятым в гельминтологии методикам (Боев и др., 1962; Pritchard, Kruse, 1982).
Экстенсивность инвазии хозяина j-й выборки (P, prevalence) определяли по формуле:
P = (Nij/Nj) · 100 ± √((P·(100 – P))/Nj),
где Nij – число хозяев j-й выборки, зараженных i-м видом гельминтов, Nj – объем j-й выборки, √ – квадратный корень, ∙ – знак умножения.
При нулевой или стопроцентной зараженности экстенсивность инвазии хозяина подсчитывали по формуле Ван дер Вардена:
Р = ((N0 + 1) / (Nj + 2))·100 ± √((P·(100 – P))/(Nj + 3)),
где N0 – число хозяев j-й выборки, свободных или стопроцентно зараженных i-м видом гельминтов (Рокицкий, 1973). Индекс обилия гельминта i-го вида (M, mean abundance) рассчитывали по формуле:
M = nij/Nj ± σ/√Nj,
где σ – среднее квадратичное отклонение, √ – квадратный корень (Плохинский, 1970).
Достоверность различий между выборками по экстенсивности инвазии определяли при помощи критерия Стьюдента (tφ), полученного после преобразования долей зараженных амфибий в углы φ (Ивантер, Коросов, 2010). Сравнение проб хозяина по индексу обилия можно проводить с использованием критерия Стьюдента (tst), но этот метод приемлем только при нормальном распределении числа паразитических организмов. В изученных биотопах отношение дисперсии к индексу обилия (s2/M) гельминта превышает единицу (см. табл. 1). Это говорит о перерассеянном распределении численности паразита у R. arvalis (Бреев, 1972). Поэтому отличие выборок хозяина по численности i-го вида гельминтов мы устанавливали, сравнивая фактическую численность i-го вида гельминтов с теоретической. Подсчет последней проводили, исходя из нулевой гипотезы (Н0) о равномерном распределении обилия гельминта относительно проб, по формуле:
nij* = (ni / N) ∙ Nj,
где ni – общее число гельминта i-го вида во всех пробах хозяина, N – число хозяев во всех выборках, nij* – теоретическая численность гельминта i-го вида в j-й выборке хозяина (Песенко, 1982). Достоверность различия между фактическим и теоретическим значениями численности устанавливали по критерию Пирсона (χ2) в соответствии с формулой:
χ2 = (nij – nij*)2/nij*.
Значения χ2, подсчитанные для отдельных выборок, суммировали и сравнивали с критическим при данном числе степеней свободы. Последнее определяли по формуле: df = k – 1, где k – число выборок.
Уровень смещения численности гельминта i-го вида относительно сравниваемых проб хозяина вычисляли с помощью показателя степени относительной биотопической приуроченности вида по формуле (Песенко, 1982):
Fij = ((nij/Nj) – (ni – nij) / (N – Nj)) / ((nij/Nj) + (ni – nij) / (N – Nj)).
Величина Fij изменяется в интервале от +1 и до –1. При Fij = +1 i-й вид гельминтов полностью предпочитает, а при Fij= –1 полностью избегает j-ю выборку хозяина. При Fij = 0 i-й вид относится к j-й выборке нейтрально, т. е. не отвергает, но и не предпочитает ее.
Результаты
1. Распространение Oswaldocruzia filiformis и параметры заражения хозяев
Нематода зарегистрирована в выборках остромордой лягушки из колочной лесостепи, поймы р. Иртыш, озерных котловин правобережной сухой степи, межсопочных озер и водотоков Казахского мелкосопочника, антропогенных биотопов г. Павлодар (см. рис. 1). По экстенсивности заражения хозяина и численности гельминта биотопы распадаются на три группы (см. табл. 1). В первой (Фёдоровка, оз. Кожа, Усолка, Черноярка, Баянаул) оба (или один) параметра зараженности превышают их среднее значение, рассчитанное по всем исследованным лягушкам. Так, значение экстенсивности инвазии хозяина из Фёдоровки, оз. Кожа, Усолки и Черноярки достоверно выше, чем среднее значение показателя (tφ = 2.24–7.54). Лишь в Баянауле зараженность R. arvalis равна среднему значению показателя (tφ = 0.05). В каждой пробе данной группы фактическая численность Os. filiformis у хозяина достоверно выше теоретической в 1.2–2.5 раза. Величина критерия χ² превышает критическую, равную 3.84, при α = 0.05 и df = 1.
| Таблица 1. Параметры инвазии и размещение численности Oswaldocruzia filiformis у Rana arvalis в степной зоне Республики Казахстан (Павлодарская область)
Table 1. Parameters of invasion and allocation of Oswaldocruzia filiformis in Rana arvalis from the steppe of the Republic of Kazakhstan (Pavlodar region) |
||||||||
| Название (номер) выборки 1 | Nj | P | M | s2/M | nij | nij* | χ² | Fij |
| Фёдоровка (22) | 25 | 96.0 ± 3.9 | 8.3 ± 1.5 | 7.0 | 207 | 82.5 | 188.0 | +0.44 |
| Оз. Кожа (15) | 25 | 80.0 ± 8.0 | 5.2 ± 1.5 | 11.3 | 129 | 82.5 | 26.2 | +0.22 |
| Усолка (1) | 1269 | 71.8 ± 1.3 | 4.0 ± 0.2 | 7.7 | 5082 | 4186.5 | 191.5 | +0.21 |
| Черноярка (2) | 274 | 67.2 ± 2.8 | 4.7 ± 0.4 | 9.7 | 1288 | 903.9 | 163.2 | +0.20 |
| Баянаул (35) | 49 | 59.2 ± 7.0 | 4.0 ± 0.9 | 9.4 | 198 | 161.7 | 8.17 | +0.10 |
| Оз. Сабандыколь (34) | 67 | 62.7 ± 5.9 | 2.4 ± 0.5 | 6.6 | 163 | 221.0 | 15.2 | -0.15 |
| Карьер (4) | 345 | 51.9 ± 2.7 | 2.3 ± 0.2 | 7.6 | 788 | 1138.2 | 107.7 | -0.20 |
| Яблонька (3) | 284 | 46.8 ± 3.0 | 2.2 ± 0.2 | 6.1 | 623 | 936.9 | 105.2 | -0.22 |
| Оз. Борли (16) | 18 | 44.4 ± 11.7 | 1.3 ± 0.5 | 2.9 | 24 | 59.4 | 21.1 | -0.43 |
| Оз. Тлеуберды (17) | 35 | 28.6 ± 7.6 | 2.3 ± 0.7 | 8.4 | 80 | 115.5 | 10.9 | -0.18 |
| Оз. Малыбай (18) | 39 | 7.7 ± 4.3 | 0.2 ± 0.1 | 2.9 | 6 | 128.7 | 116.9 | -0.91 |
| АБС 2 (5) | 147 | 7.5 ± 2.2 | 0.1 ± 0.0 | 1.5 | 16 | 485.0 | 453.5 | -0.94 |
| Сарыолен (37) | 31 | 3.0 ± 2.93 | 0.0 ± 0.0 | 0 | 102.3 | 102.3 | -1.00 | |
| Всего: | 2608 | 59.5 ± 1.0 | 3.3 ± 0.1 | 8.5 | 8604 | 8604.0 | ||
| Примечание. 1 – расположение пунктов исследования, см. рис. 1; Nj – объем выборки, экз.; P – экстенсивность инвазии, %; M – индекс обилия, экз.; s2/M – отношение дисперсии к индексу обилия; 2 – агробиологическая станция пединститута; 3 – значение получено по формуле Ван дер Вардена. | ||||||||
Показатель Fij имеет положительный знак, т. е. гельминт достоверно предпочитает биотопы данной группы. Во вторую группу выборок входят оз. Сабандыколь, антропогенные биотопы Карьер и Яблонька, оз. Борли и оз. Тлеуберды (см. табл. 1). Экстенсивность инвазии хозяина из Карьера, Яблоньки и оз. Тлеуберды достоверно меньше (tφ = 2.71–4.12), а из озер Борли и Сабандыколь – равна средней величине (tφ = 1.29 и 0.06 соответственно). Ожидаемое при равномерном размещении число Os. filiformis в каждом из биотопов этой группы, в отличие от первой, достоверно больше фактического в 1.4–2.5 раза. Отрицательный знак при показателе Fij указывает на то, что нематода избегает выборки R. arvalis из данных пунктов. В сравнении с третьей группой амфибий (оз. Малыбай, антропогенный биотоп АБС и Сарыолен), в которой величина Fij приближается или равна единице, можно считать, что степень избегания гельминтом второй группы проб хозяина – средняя. Теоретическое обилие Os. filiformis у лягушек из оз. Малыбай в 21, а из АБС в 30 раз меньше фактического. Это на порядок больше, чем во второй группе проб. Значение экстенсивности инвазии нематодой земноводных из АБС и Малыбая значительно ниже средней величины (tφ = 14.3 и 7.45). Очевидно, что данную группу R. arvalis нематода избегает сильнее, чем вторую. Особняком стоит выборка – Сарыолен, в которой Os. filiformis вообще отсутствует.
В биотопах первой группы места обитания и кормовые участки R. arvalis приурочены к хорошо увлажненной, относительно широкой прибрежной полосе водоемов и водных потоков. Побережья в них покрыты густой мезофитной растительностью, часто затенены отдельными деревьями, кустарником или тростником. Для второй группы биотопов характерны более узкие прибрежные полосы, покрытые менее плотным растительным покровом, изредка затененные тальником (Salix sp.) или тростником. Две пробы лягушек – Малыбай и АБС – отловлены на узкой, покрытой относительно редкой луговой растительностью, быстро переходящей в зону ксерофитов. Хозяева из пункта Сарыолен добыты в конце августа среди ила почти высохшего руслового водоема в одноименной степной речке. В Сарыарке подобные водные потоки обычно к осени (часто и летом) пересыхают и представляют собой цепочку отдельных плесов.
Обнаруженные различия в зараженности R. arvalis связаны, по нашему мнению, с тем, что в трех группах биотопов существуют различные условия для развития яиц, миграционной активности инвазионных личинок нематоды, следовательно, для заражения хозяев. Развитие сравнительно крупных яиц освальдокрузии происходит в течение 8–10 дней во влажной почве при температуре +14...+15 °С (Griffin, 1988). Из яиц выходят личинки, которые после двух линек становятся инвазионными (Л3). Заражение хозяев происходит перорально, при неудачных попытках схватить объект питания на почве (Hendrikx, 1983). Кроме того, мы полностью принимаем предположение Б. Шайкенова (устное сообщение), что личинки Os. filiformis мигрируют на растения по водяной пленке, образующейся после дождя или выпадения росы. Миграция инвазионных личинок на растения, облегчающая контакт с хозяином, является одной из важных адаптаций, приведших к эволюционному расцвету всей группы стронгилят (Шульц, Гвоздев, 1972). Опытным путем также показано, что личинки Os. filiformis могут распространяться, используя дождевых червей в качестве транзитных хозяев (Свiтiн, 2016). Широкая, хорошо увлажненная прибрежная полоса, плотный растительный покров на ней, наличие создающих тень отдельно стоящих деревьев, кустарников и тростника, характерные для первой группы биотопов, предоставляют оптимальные условия для развития яиц и передвижения личинок Os. filiformis и, следовательно, для инвазии хозяев. Менее подходящие условия наблюдаются во второй группе биотопов, с чем связаны пониженные параметры заражения R. arvalis нематодой. Наихудшие условия для успешного развития яиц и личинок, заражения хозяев представлены в третьей группе биотопов. В районе Сарыарки, где находится пункт Сарыолен, средняя температура июля равна +22...+25 °С, максимальная доходит до +42 °С (Гвоздецкий, Михайлов, 1963). Очевидно, температурные условия на открытых, слабо заросших ивняком песчаных берегах степных речек типа Сарыолен полностью препятствуют развитию яиц и личинок освальдокрузии.
| Таблица 2. Зараженность рептилий Oswaldocruzia filiformis (Павлодарская область)
Table 2. Infection of reptiles with Oswaldocruzia filiformis (Pavlodar region) |
||||
| Название (номер) выборки2 | Ландшафт | Вид хозяина | Nj | Р |
| С-з Техникум (11) | Пойма р. Иртыш | Lacerta agilis | 17 | 47.1 ± 12.1 |
| С-з Заря (12) | Пойма р. Иртыш | Lacerta agilis | 14 | 14.3 ± 9.4 |
| С. Ямышево (13) | Пойма р. Иртыш | Lacerta agilis | 24 | 20.8 ± 8.3 |
| С. Белогорье (31) | Пойма р. Иртыш | Natrix natrix | 5 | 20.0 ± 17.9 |
| R. arvalis | 5 | 20.0 ± 17.9 | ||
| Акшиман (33) | Мелкосопочник, ручей | Lacerta agilis | 48 | 8.3 ± 4.0 |
| Rana arvalis | 3 | 33.3 ± 27.2 | ||
| Оз. Сабандыколь (34) | Мелкосопочник | Lacerta agilis | 16 | 12.5 ± 8.3 |
| Rana arvalis | 61 | 62.7 ± 5.9 | ||
| Кызылтау (40) | Мелкосопочник, родник | Lacerta agilis | 5 | 60.0 ± 21.9 |
| Rana arvalis | 5 | 80.1 ± 7.9 | ||
| Примечание. 1 – для сравнения добавлены выборки остромордой лягушки из тех пунктов, где встречены инвазированные рептилии; 2 – расположение пунктов исследования, см. рис. 1; Nj – объем выборок, экз.; Р – экстенсивность инвазии хозяев, %. | ||||
Кроме остромордой лягушки Os. filiformis обнаружена в трех выборках L. agilis, взятых в правобережной пойме р. Иртыш (табл. 2). Симметричные пробы лягушек из этих пунктов отсутствуют. Кроме того, нематода найдена у прытких ящериц, обитающих около озер, ручьев и родников в межсопочных понижениях Сарыарки. На оз. Сабандыколь экстенсивность инвазии амфибии паразитом достоверно выше, чем у рептилии (tφ = 3.97). Выборки R. arvalis и L. agilis из горных поднятий Акшиман и Кызылтау также отличаются по зараженности Os. filiformis, но надежно (tφ = 1.09 и 0.70 соответственно). Очевидно, это связано с малым объемом выборок. Следует отметить, что при обнаружении Os. filiformis в пробах ящерицы всегда отмечается перекрывание ее ареала с таковым остромордой лягушки, обязательно зараженной обсуждаемым видом гельминтов (см. табл. 2). В пунктах из мелкосопочника, где в пробах обоих видов хозяев установлена Os. filiformis, буквально в 50–100 м выше по травянистому склону сопок, куда из-за сухости лягушки не поднимаются, ящерицы всегда свободны от освальдокрузии. В аллопатричных с R. arvalis выборках L. agilis, взятых у подножья горных поднятий Каракуз (№ 36; 48 экз.), Баянаул (№ 35, 31 экз.), Акшиман (№ 33, 22 экз.) и Кызылтау (№ 40, 33 экз.), на канале Иртыш-Караганда (№ 30; 25 экз.) и в окр. г. Павлодар (№ 5 и 6; 31 экз.) Os. filiformis не обнаружена. Нематода также отсутствует в аллопатричных с R. arvalis пробах прыткой (30 экз.) и живородящей (6 экз.) ящериц, взятых в долине ручья, впадающего в оз. Маралды (№ 20). Остромордая лягушка здесь не обитает. В целом экстенсивность заражения L. agilis нематодой в наших сборах (12.9 ± 1.4 %, N = 582 экз., tφ = 1.03) близка к величине показателя в биотопах Волыно-Подольской возвышенности и Приднепровской равнины Украины (15.5 ± 2.3 %, N = 252 экз.) (Sharpilo et al., 2001).
Кроме ящериц освальдокрузия установлена у одной особи обыкновенного ужа в пункте Белогорье (№ 31) из левобережной поймы р. Иртыш. У остальных видов рептилий и обыкновенной жабы в наших сборах Os. filiformis не найдена.
2. Экспериментальное изучение развития личинок Oswaldocruzia filiformis из яиц, происходящих от самок нематоды из разных видов хозяев
Половозрелые самки Os. filiformis, извлеченные в июне из R. arvalis и L. agilis (Усолка) и отсаженные по отдельности в чашки Петри с отстоявшейся водопроводной водой и физраствором, выделяли яйца. В трех случаях у самок, извлеченных из ящериц, мы, вслед за В. П. Шарпило (1976), в дистальных частях маток наблюдали яйца со сформированными личинками. Число яиц, выметанных нематодами, собранными у лягушек, колебалось между 14 и 128, у ящериц – между 11 и 107 экз. Очевидно, что плодовитость самок Os. filiformis, паразитирующих в лягушках и ящерицах, приблизительно одинакова. Из яиц, независимо от вида хозяина, служившего источником самок нематоды, на вторые сутки при комнатной температуре развивались личинки. Активность личинок в чашках Петри с водой сохранялась 22, с физраствором – 25 суток. При подсушивании личинки становились неподвижными, после увлажнения их движения возобновлялись. Первая линька личинок наблюдалась на третий день после начала опыта. Вторая – между 8-м и 20-м днем. Трехдневные личинки, выплодившиеся из яиц Os. filiformis, собранных у лягушек и ящериц, достигали статистически одинаковой длины, соответственно 128.7–308.0 (293.5 ± 23.4) и 120.7–310.1 (233.3 ± 34.6) мкм. Критерий tst, равный 1.44, меньше критического значения, которое при α = 0.05 и df = 53 равно 2.02. У отдельных личинок кутикулярные чехлики от линьки сохранялись до 20-го дня после выхода из яиц. К этому времени длина личинок, полученных из яиц от самок Os. filiformis из лягушек, равнялась 372.6 ± 97.2 мкм (N = 25), из ящериц – 312.7 ± 89.5 мкм (N = 23). Отличия не достоверны tst = 0.45, df = 46, tкрит = 2.02. При посеве яиц нематоды на питательную среду (культура кишечной палочки на агаре) уже на следующий день появлялись отдельные подвижные личинки Os. filiformis первой стадии развития (Л1). На 8-й день их число достигало 70 экз. При комнатной температуре личинки сохраняли подвижность. Первую линьку личинок на агаре наблюдали на 8-й день. До второй линьки гельминты в этом опыте не доживали и погибали через 8–10 дней. Очевидно, агар слишком плотная для них среда. В целом личинки из яиц, полученных от cамок Os. filiformis, извлеченных из R. arvalis и L. agilis, одинаково успешно развивались при комнатной температуре в физрастворе и водопроводной воде. В темпах роста и длительности развития в искусственных условиях, размерах одновозрастных Л1 и Л2, развившихся из яиц, полученных от самок Os. filiformis из двух видов хозяев, статистически существенных отличий мы не установили.
| Таблица 3. Характеристики Oswaldocruzia filiformis из разных видов хозяев (по литературным и нашим данным)
Table 3. Characteristics of Oswaldocruzia filiformis from different host types (according to literature and our data) |
|||||
| Признаки | Виды хозяев | ||||
| Rana temporaria (Moravek, Voitkova, 1975) | Lacerta agilis (Шарпило, 1976) | Bufo viridis (Рыжиков с соавт., 1980) | Rana arvalis (Ваккер, Тарасовская, 1988) | Vipera berus (Новохацкая, 2008б) | |
| Самки Oswaldocruzia filiformis, мм. | |||||
| Длина тела | 9.75–24.2 | 10.0–15.2 | 12.0–20.0 | 8.38–15.9 | 5.90–9.20 |
| Ширина тела | 0.177–0.408 | 0.16–0.21 | 0.23–0.28 | 0.16–0.36 | 0.12–0.20 |
| Длина везикулы | 0.090–0.105 | 0.077–0.082 | 0.040–0.116 | 0.07–0.08 | |
| Длина передней части везикулы | 0.033–0.045 | 0.032–0.060 | 0.03–0.04 | ||
| Ширина передней части везикулы | 0.011 | 0.049–0.55 | 0.048–0.064 | ||
| Длина пищевода | 0.516–0.612 | 0.42–0.47 | 0.45–0.49 | 0.32–0.54 | 0.33–0.46 |
| Длина хвоста | 0.267–0.367 | 0.23–0.33 | 0.26–0.33 | 0.14–0.32 | 0.14–0.28 |
| Вульва-терминус | 3.47–8.5 | 4.1–5.5 | 4.2–7.1 | 3.36–6.52 | 2.60 –3.40 |
| Размеры яиц | 0.096–0.117 х 0.057–0.060 | 0.082–0.104 х 0.055–0.060 | 0.099–0.104 х 0.055 | 0.068–0.100 х 0.036–0.056 | 0.06–0.08 х 0.03–0.05 |
| Самцы Oswaldocruzia filiformis, мм. | |||||
| Длина тела | 5.47–11.8 | 6.4–8.9 | 5.9–6.2 | 6.00–10.4 | 3.50–6.70 |
| Ширина тела | 0.109–0.231 | 0.11–0.17 | 0.13–0.16 | 0.14–0.26 | 0.10 –0.24 |
| Длина везикулы | 0.060–0.117 | 0.077–0.099 | 0.077–0.082 | 0.063–0.087 | 0.06–0.07 |
| Длина передней части везикулы | 0.033–0.039 | 0.036–0.048 | 0.03–0.03 | ||
| Ширина передней части везикулы | 0.049 | 0.049 | 0.040–0.056 | ||
| Длина пищевода | 0.420–0.598 | 0.37–0.42 | 0.38–0.42 | 0.22–0.52 | 0.32–0.40 |
| Длина спикулы | 0.201–0.240 | 0.187–0.203 | 0.193–0.231 | 0.184–0.232 | 0.16–0.18 |
В линейных размерах взрослых Os. filiformis из R. arvalis и L. agilis, по нашим и литературным данным, значительных отличий не наблюдается (табл. 3). Так, длина самок Os. filiformis от прыткой ящерицы практически (7.7–14.5 мм) такая же, что у остромордой лягушки. Нет существенных отличий по длине везикулы (0.030–0.087 мм), положению вульвы от терминуса (3.20–6.10 мм) и размерам яиц (0.05–0.90 х 0.04–0.05 мм). Величина самцов из ящериц приближается к таковым у R. temporaria. Согласно нашим данным, длина самцов от прыткой ящерицы равна 6.20–12.8 мм. Спикулы по цвету, количеству и характеру отростков такие же, как у остромордой лягушки. Длина спикул – 0.22–0.24 мм.
3. Паразитарная система Oswaldocruzia filiformis
По числу хозяев, необходимых для осуществления цикла развития паразитических организмов, В. Н. Беклемишев (1956) выделил простые паразитарные системы: двухчленные (1 вид окончательного хозяина), трехчленные (2 вида окончательных хозяев и 1 вид промежуточных) и многочленные (3 или более видов окончательных хозяев и 2 или более промежуточных). Включение в простую паразитарную систему одного или более видов хозяев, выполняющих ту же функцию, переводит ее в ранг сложной. Например, вместо одного окончательного и одного промежуточного хозяев при простой трехчленной системе в сложную трехчленную систему акантоцефалы Arhythmorhynchus petrochenkoi входят 5 облигатных, 2 вида факультативных, 20 видов абортивных окончательных хозяев и один промежуточный хозяин (Контримавичус, Атрашкевич, 1982). Разнообразные способы заражения, сложные трофические связи паразита и хозяина (или хозяев) придают паразитарным системам статус особых экосистем, обладающих собственными пространственными и временными границами, способных к самовоспроизведению и саморегуляции (Балашов, 2011).
| Таблица 4. Параметры инвазии Rana arvalis и Lacerta agilis нематодой Oswaldocruzia filiformis в пойме р. Иртыш (Усолка) и Казахском мелкосопочнике (Баянаул)
Table 4. Parameters of the invasion of Rana arvalis and Lacerta agilis by the nematode Oswaldocruzia filiformis in River Irtysh (Usolka) and Kazakh hilly steppe (Bayanaul) |
||||||||
| Виды хозяев | Nj | nij | nij* | χ² | Fij | P | M | s²/M |
| р. Иртыш (Усолка) | ||||||||
| Rana arvalis | 1269 | 5082 | 4808.3 | 15.6 | +0.48 | 71.7 ± 1.3 | 4.0 ± 0.2 | 7.7 |
| Lacerta agilis | 115 | 162 | 435.7 | 172.0 | -0.48 | 33.9 ± 4.4 | 1.4 ± 0.3 | 6.1 |
| Всего: | 1384 | 5244 | 5244.0 | 187.6 | tφ =7.96 | tst = 7.21 | ||
| Казахский мелкосопочник (Баянаул, долина ручья Кафар) | ||||||||
| Rana arvalis | 49 | 198 | 97.1 | 104.9 | +0.87 | 59.2 ± 7.0 | 4.0 ± 0.9 | 9.4 |
| Lacerta agilis | 59 | 16 | 116.9 | 87.1 | -0.87 | 6.8 ± 3.3 | 0.3 ± 0.2 | 12.0 |
| Всего: | 108 | 214 | 214.0 | 192.0 | tφ = 6.35 | tst = 4.06 | ||
| Примечание. Nj – объем выборок, экз.; nij – фактическая численность нематоды, экз.; nij* – теоретическая численность нематоды, экз.; χ² – критерий согласия; Fij – показатель степени приуроченности относительного обилия нематоды; P – экстенсивность инвазии, %; M – индекс обилия, экз.; tφ – критерий Фишера; tst – критерий Стьюдента при сравнении параметров инвазии хозяев из Усолки и Баянаула. | ||||||||
На исследованной территории Os. filiformis образует двухчленную паразитарную систему двух типов: простую и сложную. В пункте Черноярка, где единственным хозяином нематоды является популяция R. arvalis, функционирует простая двухчленная паразитарная система. Параметры инвазии хозяина приведены в табл. 1. В биотопах р. Иртыш (Усолка) и Казахского мелкосопочника (долина ручья Кафар, Баянаульское горное поднятие), где заражены два вида хозяев – R. arvalis и L. agilis, образовались и существуют сложные двухчленные паразитарные системы. Значение экстенсивности инвазии лягушки на Усолке достоверно выше, чем ящерицы (табл. 4). Критическая величина критерия Стьюдента равна 1.96. Распределение численности нематоды на Усолке и Кафаре относится к перерассеянному типу, так как отношение дисперсии к индексу обилия у обоих видов хозяев больше единицы (см. табл. 1). Размещение гельминта относительно выборок хозяев неравномерно (см. табл. 4). Теоретическое число гельминта в Усольской популяции лягушки в один раз достоверно меньше эмпирического. Величина критерия χ² превышает критическую, равную 3.84, при одной степени свободы и α = 0.05. Ожидаемое в соответствии с Н0 количество нематод у прыткой ящерицы, в отличие от лягушки, больше эмпирического в 2.7 раза. Положительный знак при Fij свидетельствует о том, что численность Os. filiformis достоверно и в значительной степени смещена к выборке амфибии. В Баянауле величина экстенсивности инвазии лягушки также достоверно превышает таковую ящерицы (см. табл. 4). Ожидаемое число освальдокрузий в выборке амфибии в 0.45 раза меньше, а рептилии – в 7.3 раза больше, чем найдено фактически. Для этого биотопа характерна высокая степень приуроченности обилия Os. filiformis к пробе R. arvalis.
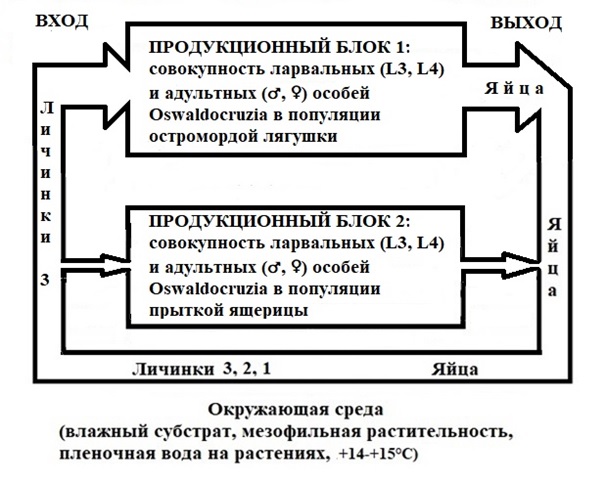
Рис. 2. Схема сложной двухчленной паразитарной системы Os. filiformis в биотопах степной зоны Республики Казахстан (объяснения в тексте)
Fig. 2. Scheme of the complex two-member parasitic system Os. filiformis in biotopes of the steppe of the Republic of Kazakhstan (explained in the text)
Сложная двухчленная паразитарная система Os. filiformis, опирающаяся на популяции R. arvalis и L. agilis, состоит из двух подсистем (рис. 2). Одна представлена освальдокрузиями, населяющими популяцию остромордой лягушки. Другая включает червей, паразитирующих в популяции прыткой ящерицы. Каждая подсистема состоит из трех обязательных частей – вход, продукционный блок (трофико-репродуктивный блок по Грановичу, 2009, с. 333) и выход. Вход образуют инвазионные личинки 3, продукционный блок – ларвальные (Л3, Л4) и адультные особи гельминта, выход – яйца, продуцируемые самками нематоды (см. рис. 2). Значение остромордой лягушки и прыткой ящерицы в паразитарной системе Os. filiformis подсчитывали по методике В. Л. Контримавичуса и Г. И. Атрашкевича (1982). Величина потока инвазии, поступающего из окружающей среды в популяцию хозяина, согласно названным авторам, равна произведению индекса обилия гельминта на относительную численность вида хозяина.
| Таблица 5. Потоки инвазии в cложных двухчленных паразитарных систем Oswaldocruzia filiformis на р. Иртыш (Усолка) и в Казахском мелкосопочнике (Баянаул)
Table 5. Flows of invasion in complex two-membered parasitic systems Oswaldocruzia filiformis on the river. Irtysh (Usolka) and in the Kazakh hilly steppe (Bayanaul) |
|||||||
| Вид хозяина | Ni:Nj | M | M♀ | Потоки инвазии в паразитарной системе | |||
| на входе | на выходе | ||||||
| экз. | % | экз. | % | ||||
| Пойма р. Иртыш (Усолка) | |||||||
| Rana arvalis | 5:1 | 4.00 | 2.10 | 20.0 | 93.5 | 10.5 | 93.1 |
| Lacerta agilis | 1:5 | 1.40 | 0.78 | 1.40 | 6.5 | 0.78 | 6.9 |
| Всего: | 21.4 | 100.0 | 11.28 | 100.0 | |||
| Казахский мелкосопочник (Баянаул, долина ручья Кафар) | |||||||
| Rana arvalis | 1:2 | 4.00 | 2.16 | 4.00 | 88.1 | 2.16 | 88.5 |
| Lacerta agilis | 2:1 | 0.27 | 0.14 | 0.54 | 11.9 | 0.28 | 11.5 |
| Всего: | 4.54 | 100.0 | 2.44 | 100,0 | |||
| Примечание. Ni : Nj – отношение численности остромордая лягушка (i): прыткая ящерица (j); M – индекс обилия, экз.; M♀ – индекс обилия половозрелых самок Os. filiformis, экз. | |||||||
Умножением индекса обилия половозрелых самок на их плодовитость и на относительную численность вида хозяина авторы определяли размер потока инвазии на выходе из популяции хозяина во внешнюю среду. По данным табл. 5, на Усолке поток инвазии Os. filiformis на входе в подсистему «остромордая лягушка» равен 4.0 · 5 = 20.0, а в подсистему «прыткая ящерица» – 1.4 · 1 = 1.4. Если принять величину всего потока инвазии на входе за единицу (20.0 + 1.4 = 21.4), то на подсистему «лягушка» приходится (20.0 / 21.4) · 100 = 93.5 %, а на подсистему «ящерица» – (1.4 / 21.4) · 100 = 6.5 % потока инвазии. При подсчете величины потоков инвазии на выходе из подсистем мы использовали два сомножителя: индекс обилия зрелых самок нематоды и относительную численность видов хозяев. Третий сомножитель – плодовитость самок гельминта – мы приняли равным единице, т. к. надежных различий в числе яиц у самок, добытых из R. arvalis и L. agilis, не установили. На Усолке доля потока инвазии на входе из продукционного блока «популяция R. arvalis» в 14.3, а на выходе в 13.5 раза больше, чем из «популяции L. agilis» (табл. 5). В Баянауле в паразитарной системе Os. filiformis доля инвазионного потока на входе в подсистему «остромордая лягушка» почти 7.4 раз, а на выходе в 7.7 раз больше, чем из подсистемы «прыткая ящерица» (см. табл. 5). Относительная численность рептилии в Баянауле немного больше, чем на Усолке. С этим связана чуть большая доля потока инвазии в паразитарной системе Os. filiformis, зависящая от прыткой ящерицы в мелкосопочнике, сравнительно с припойменным биотопом р. Иртыш.
Таким образом, в степной зоне Казахстана в зависимости от биотопических условий популяции остромордой лягушки могут обеспечивать от 88 до 93 % потока инвазии на входе и выходе из паразитарной системы Os. filiformis. Это вместе с более высокой экстенсивностью инвазии и повышенной степенью приуроченности обилия нематоды к пробам R. arvalis указывает на ведущее значение популяций остромордой лягушки в сложной паразитарной системе Os. filiformis. Прыткая ящерица заражена достоверно меньше остромордой лягушки, показатель степени приуроченности относительного обилия (Fij) имеет отрицательное значение и в сложной паразитарной системе нематоды играет второстепенную роль. Как было показано, инвазированность рептилии Os. filiformis наблюдается только в условиях симпатрии с зараженными популяциями R. arvalis. Исходя из этого мы считаем, что в степной зоне Казахстана L. agilis не может, подобно R. arvalis, самостоятельно поддерживать существование простой двухчленной паразитарной системы Os. filiformis.
Обыкновенный уж в пойме р. Иртыш, наряду с остромордой лягушкой и прыткой ящерицей, видимо, также может вовлекаться в паразитарную систему нематоды. Его вероятную роль предстоит изучить. Так же, как обыкновенной жабы и озерной лягушки, у которых зараженность установлена в окрестностях Риддера и Усть-Каменогорска (Ваккер и др., 1990).
Одним из важных результатов межвидовых взаимодействий в паразитарных системах является формирование гостальной специфичности. По степени естественной восприимчивости к паразитам выделяются следующие основные категории хозяев: облигатные, факультативные, резервуарные (транспортные) и абортивные (тупиковые, каптивные) (Шульц, Давтян, 1954; Догель, 1962; Щульц, Гвоздев, 1972; Балашов, 2001; Нигматуллин, 2004). Обычно определение гостального статуса гельминтов решается путем сравнения морфологических и физиологических признаков, величины экстенсивности инвазии хозяев, реже – индекса обилия (Нигматуллин, 2004). Применение последнего ограничено редко встречающимся в природе нормальным типом распределения численности паразитов. В биотопах степной зоны половозрелые формы Os. filiformis, в том числе самки со зрелыми яйцами, найдены у зараженных остромордых лягушек, прытких ящериц и обыкновенного ужа. Следовательно, к последним применимы категории облигатных или факультативных хозяев, поскольку только у таких хозяев гельминты проходят полное, без существенных морфологических и физиологических изменений, развитие всех органов и достигают половой зрелости. Согласно нашим данным, заметных морфологических отличий у половозрелых форм и личинок нематоды не отмечено. Ведущая роль в паразитарной системе Os. filiformis, высокий уровень заражения, широкое распространение гельминта у остромордой лягушки в биотопах лесостепи, степи, мелкосопочника, поймы Иртыша и озерных впадин говорят в пользу облигатного статуса этого вида хозяев. Второстепенная роль прыткой ящерицы в сложной паразитарной системе гельминта, заражение рептилии только при симпатрии с инвазированной нематодой амфибией, невысокая величина экстенсивности инвазии, отсутствие приуроченности относительного обилия Os. filiformis к L. agilis свидетельствуют о том, что в степной зоне Казахстана этот вид хозяев является факультативным хозяином нематоды. Мы полагаем, что, являясь, по сравнению с R. arvalis, менее гигрофильным видом (Прыткая ящерица, 1976; Банников и др., 1977), L. agilis в ранге факультативного хозяина могла способствовать проникновению Os. filiformis из экологических желобов (реки Иртыш и Урал) в биотопы озерных впадин колочной лесостепи и степи.
В. П. Шарпило (1976) относит прыткую ящерицу к облигатным хозяевам Os. filiformis. И действительно, в лесо-луговых стациях Карпат значение экстенсивности инвазии L. agilis в среднем равно 80 %, что указывает на ее возможный облигатный гостальный статус. Однако в 18 выборках (342 экз.), сделанных из южной части степной зоны Украины, лишь три ящерицы (0.88 ± 0.03 %) заражены Os. filiformis (Sharpilo et al., 2001). Это, по-видимому, свидетельствует о том, что на юге Украины статус прыткой ящерицы меняется на факультативный. Смена гостального статуса у паразитических организмов – обычное явление. Так, форма Echinococcus granulosus, вызывающая у домашних овец цистный гидатидоз, установлена у необычных хозяев: у девяти вынужденно оперированных детей и у одной забойной коровы (Лукманова и др., 2006). На птичьих базарах Земли Франца-Иосифа в паразитарную систему скребня Polymorphus phippsi, кроме облигатного хозяина обыкновенной гаги (Somateria mollissima), как факультативный хозяин включается полярная крачка (Sterna paradisaea) и обеспечивает до 1 % потока инвазии (Галактионов, Атрашкевич, 2015). Указанные авторы также установили, что люрик (Alle alle) на острове Врангеля сменил по отношению к P. phippsi каптивно-абортивный статус на факультативный. Свидетельством потенциальной смены гостальности в самом начале процесса служат постциклические паразиты (Odening, 1974). Например, освальдокрузия, найденная у совершенно необычного хозяина – налима, очевидно, съевшего зараженную лягушку (Новохацкая, 2008а). Экспериментально показана приживаемость нематоды у обыкновенного ужа и обыкновенной гадюки (Свiтiн, 2016). У последнего вида на Кижских островах в Онежском озере установлен стабильно высокий уровень зараженности Os. filiformis (Новохацкая, 2008б). Феномен связан с тем, что ведущим пищевым объектом рептилии являются травяные лягушки, в том числе инвазированные нематодой.
Заключение
В северо-восточной части степной зоны Республики Казахстан паразитарной системой Os. filiformis охватываются самые массовые виды местной батрахо- и герпетофауны – остромордая лягушка и прыткая ящерица. Заметных морфологических отличий у нематод из названных видов хозяев не отмечено. У обоих видов гельминт достигает половой зрелости, плодовитость самок, обитающих у них, приблизительно одинакова. Самки Os. filiformis, добытые из лягушек и ящериц, откладывают яйца в чашках Петри с водой или физраствором. На вторые сутки из яиц выходят личинки. Их первая линька происходит на 8-й день, вторая – на 20-й. Активность личинок в физрастворе наблюдается до 25 дней. Личинки выносят подсушивание, при увлажнении их движения возобновляются. В условиях эксперимента особых отличий в выживаемости, темпах роста и развития, линьках личинок I и II стадий, развившихся из яиц, полученных от самок Os. filiformis из лягушек и ящериц, не наблюдалось.
В пойменных биотопах р. Иртыш Os. filiformis формирует простую двухчленную паразитарную систему, опирающуюся только на популяцию R. arvalis. В притеррасных участках поймы р. Иртыш и в ряде биоценозов мелкосопочника, где ареалы популяций остромордой лягушки и прыткой ящерицы перекрываются, формируется сложная двухчленная паразитарная система. Популяции остромордой лягушки обеспечивают на входе и выходе из сложной паразитарной системы от 88 до 95 % потока инвазии. Прыткая ящерица контролирует лишь 5–12 % потока. Оптимальные условия для образования и длительного функционирования паразитарных систем Os. filiformis существуют на увлажненных, заросших густой мезофитной растительностью прибрежных частях ряда озер, рек и ручьев в колочной лесостепи, право- и левобережной степи, Казахского мелкосопочника.
Границы паразитарных систем Os. filiformis зависят от нескольких факторов. Во-первых, пространство, занимаемое паразитарными системами Os. filiformis в степной зоне, ограничивается температурой и влажностью субстрата, которые обеспечивают (или нет) нормальные условия для развития яиц и свободного передвижения инвазионных личинок Os. filiformis. В связи с этим простые и сложные двухчленные паразитарные системы гельминта приурочены к увлажненным, покрытым густым растительным покровом прибережным частям водоемов и водотоков. Во-вторых, необходимым условием существования паразитарных систем освальдокрузии является расположение на указанных побережьях кормовых участков остромордой лягушки и прыткой ящерицы. В-третьих, площади, занимаемые в биотопах сложными паразитарными системами Os. filiformis, зависят от величины симпатричной зоны хозяев нематоды.
Относительно высокая зараженность, ведущая роль в сложной паразитарной системе Os. filiformis говорят в пользу облигатного гостального статуса остромордой лягушки в степной зоне Казахстана. Подчиненное значение в паразитарной системе нематоды, относительно невысокий уровень заражения, наблюдаемый лишь при симпатрии с инвазированной Os. filiformis остромордой лягушкой, указывают на факультативный гостальный статус прыткой ящерицы. Несмотря на такой ранг, ящерица благодаря меньшей гигрофильности могла способствовать выходу нематоды из экологических желобов (реки Иртыш и Урал) в биотопы озерных впадин колочной лесостепи и степи.
Библиография
Аникиева Л. В. Гостальность и внутривидовое фенотипическое разнообразие цестод рода Proteocephalus // Материалы конф., посвящ. памяти д. б. н, проф. Б. И. Купермана. Борок, 2003. С. 10.
Балашов Ю. С. Паразитизм и экологическая паразитология // Паразитология. 2011. Т. 45. № 2. С. 81–93.
Балашов Ю. С. Специфичность паразито-хозяинных связей членистоногих с наземными позвоночными // Паразитология. 2001. Т. 35. № 6. С. 473–489.
Банников А. Г., Даревский И. С., Ищенко В. Г., Рустамов А. К., Щербак Н. Н. Определитель земноводных и пресмыкающихся фауны СССР . М.: Просвещение, 1977. 415 с.
Батькаев А. И., Ваккер В. Г. Паразитарная система трихинеллы в ландшафтах Среднего Прииртышья // Материалы докл. к 4-й Всес. конф. по проблеме трихинеллеза, человека и животных / Ред. A. C. Бессонов. Ереван, 1985. С. 35–36.
Батькаев А. И., Ваккер В. Г. Роль диких хищных млекопитающих в паразитарной системе трихинеллы в ландшафтах Среднего Прииртышья // Материалы докл. к 5-й Всесоюзной конф. по проблеме трихинеллеза человека и животных / Ред. А. С. Бессонов. Новочеркасск, 1988. С. 8–11. Научная библиотека диссертаций и авторефератов: http://www.dissercat.com/content/posleuboinaya-diagnostika-trikhinelleza-zhivotnykh#ixzz5Nrq2ke38
Батькаев А. И., Ваккер В. Г. Роль корсака в циркуляции трихинелл в Среднем Прииртышье // Материалы докл. 6 науч. конф. по проблеме трихинеллеза человека и животных. Киров, 1992. С. 24–26. Научная библиотека диссертаций и авторефератов: http://www.dissercat.com/content/epizootologiya-epidemiologiya-trikhinelleza-usovershenstvovanie-mer-borby-s-nim-v-uralskom-p#ixzz5NrpILNdy
Беклемишев В. Н. Возбудители болезней как члены биоценозов // Зоологический журнал. 1956. Т. 35. Вып. 12. С. 1765–1779.
Боев С. Н., Соколова И. Б., Панин В. Я. Гельминты копытных животных Казахстана . Алма-Ата, 1962. Т. 1. 377 с.
Бреев К. А. Применение негативного биномиального распределения для изучения популяционной экологии паразитов. . Л.: Наука, 1972. 72 p. (Методы паразитологических исследований. № 6).
Буракова А. В. Эколого-генетические особенности и гельминтофауна остромордой лягушки (Rana arvalis) в градиенте урбанизации: Автореф. дис. ... канд. биол. наук . Екатеринбург, 2010. 22 с.
Буракова А. В., Вершинин В. Л. Анализ паразитофауны синтопически обитающих представителей бесхвостых амфибий // Вестник СПбГУ. Сер. 3. 2016. Вып. 3. С. 31–36. DOI: 10.21638/11701/spbu03.2016.306.
Ваккер В. Г. Популяционные особенности гельминтов у прыткой ящерицы в Среднем Прииртышье // Тезисы докл. 8 Всес. совещ. зоологов пединститутов / Ред. С. П. Наумов, Н. М. Чернова, Л. С. Шалдыбин. Витебск: ВГПИ, 1984. Ч. 2. С. 56–58.
Ваккер В. Г., Брушко З. К., Тарасовская Н. Е. К гельминтофауне амфибий Казахстана // Материалы Всесоюзного научно-методич. совещания зоологов пед. вузов СССР / Ред. С. П. Наумов, А. Г. Юсуфов. Махачкала: ДГУ, 1990. С. 59–61.
Ваккер В. Г., Тарасовская Н. Е. Биология Oswaldocruzia. filiformis (Goeze, 1782) (Trichostrongylidae) в Среднем Прииртышье . Деп. в ВИНИТИ. 1988. № 4149 – В8. С. 1–21; Анн. в РЖБ. 1988. 6К 122, М.
Галактионов К. В., Атрашкевич Г. И. Специфика циркуляции паразитов морских птиц в высокой Арктике на примере паразитарной системы скребня Polymorphus phippsi (Palaeacanthocephala, Polymorphidae) // Паразитология. 2015. Т. 49. № 6. С. 393–411.
Гвоздецкий Н. А., Михайлов Н. И. Физическая география СССР. Азиатская часть . М.: Географгиз, 1963. 572 с.
Гранович А. И. Паразитарная система как отражение структуры популяции паразитов: концепция и термины // Труды ЗИН РАН. 2009. Т. 313. № 3. С. 329–337.
Догель В. А. Общая паразитология . Л., 1962. 464 с.
Жигилева О. Н. Взаимосвязь зараженности гельминтами и генетического разнообразия популяций животных: Дис. ... д-ра биол. наук . Тюмень, 2017. 327 с.
Жигилева О. Н., Кирина Ю. И. Заражение гельминтами сибирской лягушки (Rana amurensis Boulenger, 1886) на западной границе ареала // Паразитология. 2014. Т. 47. № 2. С. 165–169.
Жигилева О. Н., Сурель О. В., Злобина Л. С. Паразитарные сообщества остромордой лягушки на юге Западной Сибири // Вестник экологии, лесоведения и ландшафтоведения. Тюмень, 2002. Вып. 3. С. 63–68.
Ивантер Е. В., Коросов А. В. Биометрия . Петрозаводск: Изд-во ПетрГУ, 2010. 104 с.
Кириллова Н. Ю., Кириллов А. А., Чихляев И. В. Анализ гельминтофауны позвоночных животных юга Среднего Поволжья и факторы ее формирования // Материалы IV Всероссийского Съезда Паразитологического общества при Российской академии наук «Паразитология в XXI веке – проблемы, методы, решения». СПб., 2008. Т. 2. С. 313–317.
Контримавичус В. Л., Атрашкевич Г. И. Паразитарные системы и их изучение в популяционной биологии гельминтов // Паразитология. 1982. Т. 16. № 30. С. 177–187.
Кудинова М. А., Жерихова Г. В., Петрова О. Е. Гельминтофауна лягушки травяной в Южной Карелии // Гельминты и их промежуточные хозяева / Ред. Л. С. Шалдыбин. Горький, 1985. С. 24–27.
Лукиянов Л. В., Ручин А. Б., Чихляев И. В., Рыжов М. К. Гельминтофауна остромордой лягушки Rana arvalis (Amphibia, Anura) // Материалы IV Всероссийского съезда Паразитологического общества при Российской академии наук «Паразитология в XXI веке – проблемы, методы, решения». СПб., 2008. Т. 2. С. 149–151.
Лукманова Г. И., Гумеров А. А., Туйгунов М. М., Викторова Т. В. Геномное типирование изолятов Echinococcus granulosus из районов Южного Урала // Паразитология. 2006. Т. 40. № 5. С. 479–484.
Нигматуллин Ч. М. К теории жизненных циклов паразитов. Терминология и классификация хозяев по их роли в жизненных циклах гельминтов // Современные проблемы паразитологии, зоологии и экологии: Материалы I и II международных чтений, посвященных памяти и 85-летию со дня рождения С. С. Шульмана (март 2002 г. и февраль 2003 г., Калининград) / Ред. Ч. М. Нигматуллин. Калининград: Изд-во КГТУ, 2004. С. 96–119.
Новохацкая О. В. Паразитофауна рыб эвтрофируемых озер (на примере Сямозера): Автореф. дис. ... канд. биол. наук . СПб., 2008а. 26 с.
Новохацкая О. В. К вопросу о встречаемости нематоды Oswaldocruzia filiformis (Strongylida: Molineidae) в Карелии // Паразитология. 2008б. Т. 42. № 3. C. 204–210.
Однокурцев В. А., Седалищев В. Г. Гельминтофауна сибирской лягушки Якутии, ее половозрастная и сезонная изменчивость // Поволжский экологический журнал. 2008. № 2. С. 112–119.
Песенко Ю. А. Принципы и методы в фаунистических исследованиях . М.: Наука, 1982. 288 с.
Плохинский Н. А. Биометрия . М.: Изд-во МГУ, 1970. 359 с.
Прыткая ящерица / Ред. А. В. Яблоков. М.: Наука, 1976. 376 с.
Равковская Е. А., Хворова И. А., Пятова М. В., Лада Г. А. Гельминты обыкновенной чесночницы (Pelobates fuscus) и озерной лягушки (Pelophylax ridibundus) на востоке Центрального Черноземья // Вестник ТГУ. 2015. Т. 20. Вып. 1. С. 185–188.
Радченко Н. М., Шабунов А. А. Эколого-гельминтологические исследования амфибий Вологодской области // Материалы IV Всероссийского съезда Паразит. об-ва РАН «Паразитология в XXI веке – проблемы, методы, решения». СПб., 2008. Т. 3. С. 72–75.
Рокицкий П. Ф. Биологическая статистика . Минск: Вышэйша школа, 1973. 320 с.
Русинек О. Т. Паразитарная система Diphyllobotrium dendriticum (Cestoda: Pseudophyllidea) и оценка ее возраста в озере Байкал // Паразиты рыб: современные аспекты изучения: Материалы конф., посвящ. памяти д. б. н., проф. Б. И. Купермана. Борок, 2003. С. 46–47.
Ручин А. Б., Чихляев И. В. Изучение гельминтофауны остромордой – Rana arvalis Nilsson, 1842 и травяной – Rana temporaria L., 1758 лягушек (Amphibia: Anura) при совместном обитании // Современная герпетология. 2013 Т. 13. № 3/4. С. 130–136.
Рыжиков К. М., Шарпило В. П., Шевченко Н. Н. Гельминты амфибий фауны СССР . М.: Наука, 1980. 279 с.
Соболева Т. Н. К гельминтофауне водных амфибий и рептилий Казахстана // Экология водных животных / Ред. Е. В. Гвоздев. Алма-Ата: АН Каз. ССР, 1975. С. 186–192.
Тарасовская Н. Е. Значение бесхвостых амфибий в оздоровлении пастбищных и околоводных биотопов от гельминтов // Экология. Вестник Алтайского государственного аграрного университета. 2009. № 10 (60). С. 76–79.
Фортунато Э. Связь гостальной специфичности паразита с популяционной структурой вида хозяина. // Журнал общей биологии. 1985. Т. XLVI. № 3. С. 625–632.
Чупахин В. М. Основы ландшафтоведения . М.: Мысль, 1973. 224 с.
Шарпило В. П. Паразитические черви пресмыкающихся фауны СССР . Киев: Наукова Думка, 1976. 287 с.
Шималов В. В. Гельминтофауна земноводных открытых каналов в мелиорированных районах Белорусского Полесья // Паразитология. 2002. Т. 36. № 2. С. 304–309.
Шималов В. В. Гельминтофауна амфибий (Vertebrata: Amphibia) в Республике Беларусь // Паразитология. 2009. Т. 43. № 2. С. 118–129.
Шульц Р. С., Давтян Э. А. О формах хозяинно-паразитных отношений в гельминтологии // Зоологический журнал. 1954. Т. 6. С. 1201–1205.
Шульц Р. С., Гвоздев Е. В. Основы общей гельминтологии. Биология гельминтов . М.: Наука, 1972. Т. 2. 517 с.
Щепина Н. А., Балданова Д. Р. Гельминтофауна монгольской жабы Bufo raddei Strauch, 1876 в Забайкальи // Паразитология. 2010. Т. 44. № 2. С. 153–159.
Щепина Н. А., Дугаров Ж. Н. Трематоды сибирской лягушки бассейна оз. Байкал // Материалы IV Всероссийского съезда Паразитологического общества при Российской академии наук «Паразитология в XXI веке – проблемы, методы, решения». СПб., 2008. Т. 3. С. 732–734.
Свiтiн Р. С. Нематоди роду Oswaldocruzia Travassos, 1917 захiдной Палеарктик: Автореф. дис. ... канд. бiологiчних науки . Киiв, 2016. 20 c.
Cedhagen T. Endoparasites in some Swedish amphibians // Acta Parasitologica Polonica. 1988. Vol. 33. № 2. P. 107–113.
Griffin C. T. The effect of constant and changing temperatures on the development of the eggs and larvae Oswaldocruzia filiformis (Nematoda: Trichostrongyloidea) // J. of Helminthology. 1988. Vol. 62. Issue 4. P. 281–292.
Griffin C. T. Oswaldocruzia filiformis (Nematoda: Trichostrongyloidea) in frogs (Rana temporaria) from three locations in Ireland. // J. Helminthol. 1989. Vol. 63. P. 53–62.
Hendrikx W. M. L. Epidemiological aspects of the infection with Oswaldocruzia filiformis (Goeze, 1782) Travassos 1917 (Nematoda, Trichostrongylidae) in the common toad (Bufo bufo L.,1785) in the Netherlands // Neth. J. Zool. 1983. Vol. 33 (2). P. 99–124.
Kozak A. Die Nematodenfauna der Frösche in dem Karpathengebiet der CSSR // Biologia Bratislava. 1973. Vol. 28. P. 325–334.
Moravec F., Vojtkova L. Variabilität von zwei Nematodenarten – Oswaldocruzia filiformis (Goeze, 1782), und Oxysomatium brevicaudatum (Zeder, 1800), der gemeinsamen Parasiten der europäischen Amphibien und Reptilien // Scripta Fac. Sei. Nat. UJEP Brunensis, Biologia 2. 1975. 5. S. 61–76.
Odening К. Parasitismus. Grundfragen und Grundbegriffe. WTW, 1974. Band 113. Reihe Biologie. 170 s.
Pritchard M. H., Kruse G. O. W. The collection and preservation of animal parasites. Nebraska, 1982. P. 19–30, 41–44.
Sanchis V., Roig J. M., Carretero M. A., Roca V., Llorence G. Host-parasite relationships of Zootoca vivipara (Sauria: Lacertidae) in the Pyrenees (North Spain) // Folia Parasitologia. 2000. Vol. 47. P. 118–122.
Sharpilo, V. P., Biserkov, V., Kostadinova, A., Behnke, J. M., Kuzmin, Y. I. Helminths of the sand lizard, Lacerta agilis (Reptilia, Lacertidae), in the Palaearctic: faunal diversity and spatial patterns of variation in the composition and structure of component communities // Parasitology. 2001. Vol. 123 (4). Р. 389–400.
Schimalov V. V., Schimalov V. T., Schimalov A. V. Helminth fauna of lizards (Reptilia, Sauria) in sousern part of Belarus // Parasitol. Res., Short Communication. 2000. Vol. 86. P. 343.
Schimalov V. V., Schimalov V. T. Helminth fauna of toads in Belarussian Polesie // Parasitol. Res., Short Communication. 2001а. Vol. 87 (1). P. 84.
Schimalov V. V., Schimalov V. T., Schimalov A. V. Helminth fauna of newts in Belarussian Polesie // Parasitol. Res., Short Communication. 2001b. Vol. 87 (4). P. 356.
Singer E., Sattmann H. Zur Kenntnis der Parasiten Österreichischer Amphibien // Helminthologische / Parasitologische Fachgespräche. Naturhistorisches Museum, 3. Zoologische Abteilung, Wien, 2007. S. 1–3.
Yildirimhan H. S, Goldberg S. R., Bursey R. C. Helminth Parasites of the Banded Frog Rana camerani (Ranidae) from Turkey. // Comparative Parasitology. 2009. Vol. 73 (2). July. P. 222–236. URL: http://www.bioone.org/doi/full/10.1654/4229.1. DOI: http://dx.doi.org/10.1654/4229.1.
Благодарности
Выражаю мою искреннюю признательность профессору, доктору биологических наук Шайкенову Блоку Шайкеновичу за его ценные советы и дружескую поддержку. Отдаю глубокую дань памяти моим научным руководителям – академику Боеву Сергею Николаевичу и профессору Маркову Георгию Сергеевичу. В моем сердце жива благодарность Анвару Идрисовичу Батькаеву, которого уже нет с нами. Его профессиональные советы и помощь неоценимы.

 © 2011 - 2026
© 2011 - 2026