



Введение
Согласно классическому взгляду на территориальность (Howard, 1920; Noble, 1939; Nice, 1941), территориальные животные – это животные, обладающие собственными разграниченными территориями, охраняемыми от возможных конкурентов за какие-либо ресурсы. Большое число работ, проведенных со времени указанных исследований, свидетельствует о том, что территориальность животных какого-либо вида наблюдается в определенных условиях и может быть заменена альтернативными способами снижения конкуренции за ресурсы – в других условиях среды (Maher, Lott, 1995; Newton, 1998; Both, Visser, 2003). Вариация территориального поведения, включая вариацию в формируемых пространственных структурах, может проявляться не только среди разных видов, но и внутри одного вида (Maher, Lott, 1995; Newton, 1998). Последнее может быть особенно актуальным для видов с обширными ареалами, охватывающими различные местообитания.
Мы изучали территориальное поведение птиц, обладающих обширными ареалами, в разных условиях среды (разные части гнездовых ареалов, разные местообитания, разная плотность населения) на примере черноголовой Sylvia atricapilla, садовой S. borin, серой S. communis славок и славки-завирушки S. curruca. Славок обычно считают строго территориальными птицами, которые демонстрируют выраженное территориальное поведение (Simms, 1985; Shirihai et al., 2010). Птицы таких видов маркируют границы территорий пением, самцы защищают территории от других самцов своего вида, территории конспецифичных самцов разобщены.
В условиях острой социальной напряженности, возникающей в поселениях птиц в результате дефицита мест, пригодных для гнездования, и высокой плотности населения в центральных частях гнездовых ареалов мы наблюдали варианты территориального поведения, необычные для строго территориальных птиц. Эти данные частично были представлены в наших прежних публикациях (Зубцовский и др., 2006; Матанцева, Симонов, 2008). Подобные результаты, полученные в аналогичных условиях, публиковали и ранее (Зубцовский и др., 1988; Titus, Hans, 1990; Зубцовский, Матанцев, 1992).
Позже мы изучали поведение славок в регионе с относительно низкой их численностью – на севере видовых ареалов, где на фоне в целом типичного поведения обнаружили некоторую редукцию территориальности (снижение агрессивности, активности и продолжительности пения), наблюдаемую при низкой плотности населения в обширных местообитаниях с более или менее однородными условиями (Матанцева, Симонов, 2012).
В ходе последующей работы было обнаружено, что даже на севере видовых ареалов (при низкой численности изучаемых видов в регионе) птицы могут формировать скопления в ограниченных по площади биотопах, расположенных среди обширных зон с менее благоприятными условиями. Локальная плотность населения птиц в таких скоплениях может намного превышать среднюю популяционную плотность в регионе. В таких местах славки демонстрируют поведение, сходное с наблюдаемым в плотных поселениях в центральных частях их ареалов, включая некоторые формы, необычные для строго территориальных животных.
В этой статье мы представляем основные результаты сравнительного анализа данных, полученных нами в ходе всех перечисленных исследований, и обсуждаем внутривидовую вариацию территориального поведения славок в условиях разной плотности населения и разных местообитаний в центре и на севере гнездовых ареалов.
Материалы
Исследования были проведены в разных частях гнездовых ареалов птиц изучаемых видов на территории европейской части России: в Мурманской области, Карелии, Мордовии и Калининградской области (табл. 1). На Мурманскую область приходится крайняя северная периферия ареалов изучаемых видов, заселяемая не ежегодно, причем с очень низкой плотностью. Карелия также является северной периферией ареалов славок, однако относится к зоне регулярного гнездования птиц всех четырех видов, при этом условия их обитания здесь относительно неблагоприятны ввиду сравнительно короткого периода, пригодного для размножения, и частых возвратов холодов в начале репродуктивного сезона. Следует отметить, что в Костомукшском заповеднике, расположенном севернее других участков исследований в Карелии, численность славок также очень низка.
В Мурманской области исследования проводили в прибрежной полосе Кандалакшского залива Белого моря и на удаленных от берега территориях, в Костомукшском заповеднике – в поймах рек и по берегам озер, а также во внутренних зонах, занятых типично таежными биотопами. В этих районах участки исследований были заложены в открытых местообитаниях с травянистой растительностью и кустарниками; хвойных (сосняки, ельники) и хвойно-лиственных лесах; в населенных пунктах и их окрестностях, на зарастающих древесно-кустарниковой растительностью пустырях.
Более высоких показателей численности в регионе славки достигают в южной Карелии. Исследования здесь были проведены в Юго-Восточном Приладожье. Местообитания славок в данном районе можно разделить на два типа: предпочитаемые биотопы (участки, занятые луговой и кустарниковой растительностью, с подростом ели по опушкам леса, в поймах рек и ручьев) и обширные сплошные массивы хвойно-лиственных лесов. Более подробное описание климатических условий и биотопов дано в статье по нашим ранним исследованиям в южной Карелии (Матанцева, Симонов, 2012).
Калининградскую область и Мордовию следует отнести к центральным частям гнездовых ареалов модельных видов, заселяемых регулярно и с относительно высокой плотностью. В Калининградской области исследования проводили на Куршской косе Балтийского моря. Основной участок исследований занимал территорию стационара «Рыбачий» Зоологического института РАН. Эта территория, расположенная на берегу Куршского залива, представляет собой мозаику выделов биотопов разных типов: участки лиственного леса разной степени влажности; открытые места, занятые прибрежной луговой растительностью; заросли ивняка; поляны, заросшие ежевикой; тростниковые заросли; культурные насаждения. Более подробное описание участков исследований в этом районе представлено в нашей прежней публикации (Зубцовский и др., 2006).
В Мордовии исследования проводили на территории национального парка «Смольный» и в его окрестностях. Обследованные здесь биотопы также можно классифицировать как предпочитаемые славками местообитания (участки, занятые луговой и кустарниковой растительностью, по опушкам леса и в поймах рек) и обширные сплошные массивы смешанных лесов (сосново-лиственных и преимущественно лиственных с небольшим участием сосны и ели).
Ежедневно под наблюдением находилось до 7 птиц каждого изучаемого вида. За сезон контролировали до 19 территориальных самцов (settled males) каждого вида и до 18 самцов, находящихся на контролируемых участках не более 3 дней после индивидуального мечения в первые сутки после прилета и так и не занявших территории (unsettled males). В целом проведено около 15 000 часов наблюдений в природе; отмечено свыше 54 000 точек, маркирующих территории; найдено 288 гнезд славок.
Методы
Основное понятие в работе – понятие территории, и поскольку нет единства мнений по тому, что именно следует считать территорией, мы полагаем необходимым дать обозначения терминам, используемым в статье. Мы придерживаемся наиболее распространенного концептуального определения территории как охраняемого пространства, подразумевающего исключение возможных конкурентов с определенной площади (Maher, Lott, 1995). Однако у певчих птиц препятствием для конспецифичных особей к проникновению на занятый участок могут быть не только факты активной охраны его границ, но и демонстрирование границ пением, а охраняемый и демонстрируемый участки, как правило, пространственно совпадают (Fretwell, Lucas, 1969). Кроме того, именно факты пения на территории можно наиболее четко регистрировать, в то время как случаи охраны границ посредством визуальных демонстраций и прямой агрессии происходят реже и менее заметны, в связи с чем могут быть упущены в ходе наблюдений. Поэтому в качестве оперативного определения территории целесообразно использовать понятие территории как участка, границы которого маркируются пением (Maher, Lott, 1995).
Территории регистрировали методом точечного картирования участков, маркируемых пением, как минимальных выпуклых полигонов (Odum, Kuenzler, 1955). Этот метод был частично модифицирован с целью получения более подробных данных по размерам и конфигурации территорий на разных этапах репродуктивного периода. В частности, мы регистрировали все наблюдаемые перемещения самца, демонстрирующего территорию, на протяжении всего времени его пребывания на контролируемом участке, что позволяло определить точные контуры демонстрируемых территорий. Размер территории оценивали как площадь ее проекции на поверхность земли. Расстояние между соседними территориями определяли как минимальную дистанцию между ближайшими границами этих территорий. Кроме того, для формирования представления о трехмерных территориальных пространствах, помимо картирования территории как проекции на поверхность земли, в каждой точке отмечали высоту, на которой поет птица.
Поскольку территориальное поведение, как форма поведения социального, должно зависеть от уровня социальной напряженности в популяции и, следовательно, от плотности населения и величины пригодных для обитания площадей, мы оценивали плотность населения птиц на заселяемых ими площадях. В качестве адекватного показателя плотности населения в контексте изучения территориальности рассматривали плотность населения самцов – отношение числа самцов на определенной площади к размеру этой площади.
Находящиеся под наблюдением птицы были отловлены и помечены наборами номерных алюминиевых и цветных пластиковых колец в уникальных комбинациях, распознаваемых дистанционно. Наблюдения проводили ежедневно в течение всего сезона. Кроме того, проводили поиск гнезд и набросов (типичных для славок «заготовок» гнезд или негнездовых построек, по-видимому, выполняющих демонстрационную функцию) и отслеживали их судьбу.
Работа с картами и статистический анализ
В 2003–2005 гг. работали с картами, выполненными на бумаге. Позднее в полевых условиях использовали карманные и планшетные персональные компьютеры с программами OziExplorer 3.95.4m (D & L Software Pty Ltd, 2005) и OruxMaps v.7.0.10 (Jose Vázques, 2012). Площади территорий и расстояния между ними рассчитывали с применением программ Arc View GIS 3.2 (Ivironmental Systems Reseach Institute, Inc., 1992–1999) и Google Earth Pro 7.1.8.3036 (Google, 2016, 2017).
При статистическом анализе ряды исходных данных проверяли на форму распределения, применяя критерий Андерсона – Дарлинга, на равенство дисперсий – с помощью F-теста. Оценку связи и зависимости между величинами проводили с использованием корреляционного анализа: при нормальном распределении данных в двух независимых выборках и равных дисперсиях вычисляли коэффициент корреляции Пирсона, при прочих условиях – коэффициент ранговой корреляции Спирмена. Последний тип анализа был выбран на основании таких его качеств, как независимость от формы распределения и низкая чувствительность к выбросам.
При проверке статистических гипотез использовали 5 % уровень значимости (при доверительном интервале 95 %). Обработку данных и статистический анализ проводили с помощью пакетов программ Microsoft Excel (Microsoft Corporation, 2002–2010) и MaxStat Lite 3.6 (MaxStat, 2015).
Результаты
Параметры плотности населения славок, зарегистрированные на участках исследований, представлены в табл. 1. Следует отметить, что этот показатель существенно изменялся в пределах видовых ареалов и в зависимости от занимаемых местообитаний (см. Обсуждение).
Таблица 1. Средняя плотность населения славок на участках исследований, ♂♂/га
Table 1. The average population density of Sylvia warblers in different habitats, ♂♂/ha
| Вид | A | B.1 | B.2 | С.1 | C.2 | D | E |
| S. atricapilla | 1.72 | 0.10 | 0.01 | 0.07 | 0.02 | – | – |
| S. borin | 0.72 | 0.27 | 0.08 | 0.31 | 0.08 | 0.01 | 0.01 |
| S. communis | 0.85 | 0.22 | 0.07 | 0.89 | – | 0.03 | 0.56 |
| S. curruca | 0.71 | 0.03 | 0.02 | 0.23 | 0.09 | 0.02 | ˂ 0.01 |
| Примечание. «–» - вид не отмечен, в остальных случаях плотность населения указана только по заселяемым местообитаниям (с ненулевой численностью).
Участки исследований в центральных частях гнездовых ареалов: A – стационар «Рыбачий», Куршская коса Балтийского моря, Калининградская область (55°12’N, 20°50’E; 2003–2005): предпочитаемые биотопы. B – Национальный парк «Смольный», Мордовия (54°44’N, 45°30’E; 2016–2017): B.1 – предпочитаемые биотопы ограниченной площади; B.2 – обширные сплошные массивы смешанного леса. Участки исследований на северной периферии гнездовых ареалов, в порядке удаления от их центров: С – Юго-Восточное Приладожье, южная Карелия (60°46’N, 32°48’E; 2007–2015): С.1 – предпочитаемые биотопы ограниченной площади; С.2 – обширные сплошные массивы смешанного леса. D – Костомукшский заповедник, центральная Карелия (64°27’N, 30°17’E; 2015–2016). E – побережье Кандалакшского залива, Мурманская область (67°06’N, 32°42’E; 2015). |
|||||||
Показатели размеров территорий по тем участкам исследований, по которым удалось получить наиболее полные данные по биотопам разных типов, отражены на рис. 1. В каждом районе территории разных самцов одного вида значительно варьировали по размеру. При этом в условиях заселения обширных массивов сплошных лесов территории, в большинстве случаев, были крупнее, чем на других участках исследований (где благоприятные местообитания имели ограниченную площадь), и могли достигать больших величин. В целом размер территорий обычно уменьшался с ростом локальной плотности населения, что было статистически подтверждено для всех модельных видов, кроме серой славки (табл. 2). При этом тенденция к уменьшению площади территорий при увеличении плотности населения отмечена и для этого вида.
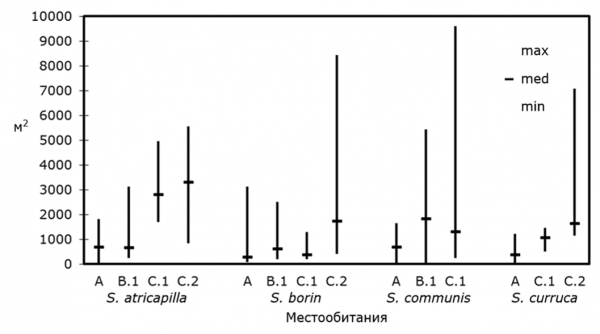
Рис. 1. Размеры территорий самцов славок в разных местообитаниях
(расшифровка буквенных обозначений в табл. 1)
Fig. 1. The territory size of Sylvia warblers in the study areas (letter symbols are explained under table 1)
Таблица 2. Параметры ранговой корреляции Спирмена между плотностью населения и размерами территорий славок
Table 2. Spearman's rank correlation between the population density and territory size of Sylvia warblers
| Вид | N | r | P |
| S. atricapilla | 40 | - 0.702 | ˂ 0.0001 |
| S. borin | 42 | - 0.438 | 0.0037 |
| S. communis | 34 | - 0.238 | 0.1746 |
| S. curruca | 26 | - 0.745 | ˂ 0.0001 |
Различия в характере формируемых территориальных структур оказались наиболее заметны также при их сравнении в наиболее благоприятных, плотно заселяемых местообитаниях и менее привлекательных для птиц участках, заселяемых с меньшей плотностью. В биотопах, заселяемых с низкой плотностью, пространственное размещение территорий было типичным для территориальных животных: территории самцов одного вида были разобщены во всех случаях (рис. 2). При этом для всех видов обнаружена обратная зависимость между плотностью населения и кратчайшим расстоянием между соседними территориями (табл. 3). Характеристики данных, полученных при изучении территориальности серой славки, позволили рассчитать коэффициент Пирсона, который также подтвердил указанную закономерность (r = -0.518, P = 0.0478, N = 15) при уровне значимости 5 %.
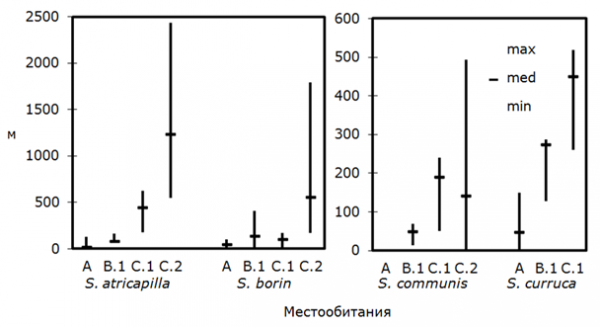
Рис. 2. Расстояние между соседними территориями самцов славок в разных местообитаниях (расшифровка буквенных обозначений в табл. 1)
Fig. 2. The distances between territories of Sylvia warblers in the study areas (letter symbols are explained under table 1)
Таблица 3. Параметры ранговой корреляции Спирмена между плотностью населения и дистанциями между территориями славок
Table 3. Spearman's rank correlation between the population density and distances between territories of Sylvia warblers
| Вид | N | r | P |
| S. atricapilla | 21 | - 0.789 | ˂ 0.0001 |
| S. borin | 31 | - 0.628 | 0.0002 |
| S. communis | 15 | - 0.630 | 0.0118 |
| S. curruca | 11 | - 0.820 | 0.0341 |
В наиболее плотных поселениях наблюдали частичное или полное перекрывание территорий у представителей всех видов, кроме славки-завирушки (рис. 2, 3). При этом в некоторых случаях перекрывания территорий по горизонтали (как проекций на поверхность земли) происходило разобщение территориальных пространств по вертикали: один из самцов-соседей пел на большей высоте от земли, другой – на меньшей (рис. 3: 4.1). Подобное перекрывание территорий было охарактеризовано нами ранее при исследовании поведения птиц в центральных частях ареалов (Матанцева, Симонов, 2008) и отмечено в наиболее плотно заселяемых ограниченных по площади биотопах на северной периферии ареалов.
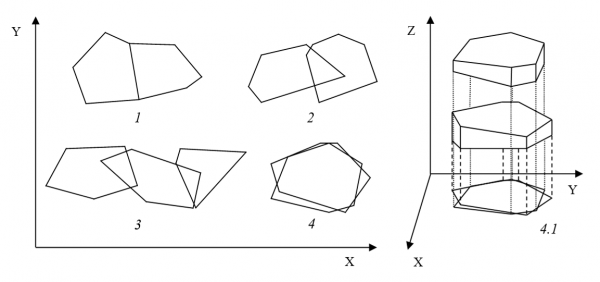
Рис. 3. Типы наблюдаемого перекрывания территорий черноголовых славок:
1 – совпадение границ; 2 – частичное перекрывание двух территорий; 3 – частичное перекрывание трех территорий; 4 – полное перекрывание двух территорий; 4.1 – разделение территориальных пространств по вертикали при перекрывании их проекций на поверхность земли
Fig. 3. Types of the observed overlap of blackcap territories:
1 – contiguity of borders; 2 – partial overlap of two territories; 3 – partial overlap of three territories; 4 – full overlap of two territories; 4.1 – vertical separation of the territories overlapped at their projections on the ground surface
На участках, где территории особей одного вида были разобщены, не было зарегистрировано случаев долгого пребывания самца на чужой территории птиц своего вида. Однако при этом не было отмечено и обычных для территориальных животных фактов агрессивного поведения на границах территорий.
Редукция территориальности в северных регионах также была заметна в отношении сокращения продолжительности пения относительно южных районов: многие самцы в северных регионах пели только в самом начале периода размножения. Некоторые из них прекращали петь уже к началу откладки яиц (особенно характерно для славок-завирушек и чернологовок), другие пели на начальных стадиях гнездования, а позже их песенная активность угасала.
Сходную редукцию строгой территориальности (снижение, вплоть до полного отсутствия, взаимной агрессии конспецифичных самцов) наблюдали и в местообитаниях с самыми высокими показателями плотности населения, но, очевидно, это было обусловлено другими причинами. В таких местах, где соседние территории перекрывались, самцы-соседи могли находиться на общих частях их территорий длительное время без видимой агрессии по отношению друг к другу. При этом все же в большинстве случаев происходило разделение их территориальной активности другими путями: в пространстве (разобщение территориальных пространств по вертикали) или во времени (временное разделение фаз активного демонстрирования территорий самцов-соседей с помощью пения, замена типичной территориальности системой иерархических отношений). При иерархических отношениях, отмеченных для садовых и черноголовых славок, самец-рецессив мог проявлять свою песенную активность только тогда, когда самец-доминант молчал. В крайне редких случаях отмечали гнездящиеся пары с самцами, для которых вообще не было зафиксировано проявления песенной активности (Матанцева, 2010). Варианты территориального поведения славок, которые могут проявляться в условиях высокой и низкой популяционной плотности, в сравнении с характеристиками типичного территориального поведения представлены в табл. 4.
Таблица 4. Характеристики типичного территориального поведения славок и его варианты, наблюдаемые в условиях высокой (ВПН) и низкой плотности населения (НПН)
Table 4. The characteristics of the typical territorial behavior of Sylvia warblers and some variants observed in conditions of high and low population density
| Типичное поведение | Варианты территориального поведения | |
| при ВПН | при НПН | |
| Строгая горизонтальная изоляция территорий конспецифичных самцов | Горизонтальное перекрывание территорий конспецифичных самцов. Разделение территориальных пространств по вертикали при значительном перекрывании их проекций на поверхность земли | Рассредоточенные в пространстве территории больших размеров. Концентрация птиц одного вида в предпочитаемых биотопах с формированием локальной ВПН в пределах обширных площадей с НПН – переход к вариантам поведения, наблюдаемым при ВПН (см. столбец 2) |
| Гнездование только при наличии собственной территории, маркируемой пением | Участие в размножении самца без собственной территории, маркируемой пением | Размножение пары на территории, маркируемой пением только в самом начале гнездования |
| Длительное пребывание на чужой территории самца своего вида невозможно вследствие агрессивной охраны границ территории | Мирное пребывание самца и даже гнездование пары в пределах чужой территории конспецифичного самца. Замена типичной территориальности системой иерархических отношений | Значительная редукция (вплоть до полного отсутствия) агрессивной защиты территорий |
Кроме указанных особенностей проявления территориальности славок в разных условиях следует также отметить тот факт, что во всех районах исследований на контролируемых участках наблюдали присутствие самцов, вообще не занявших стабильных территорий и покинувших эти участки спустя 2–3 дня после прилета. Помимо них, также во всех районах исследований были отмечены самцы, успешно занявшие территории, однако не привлекшие самок (Матанцева, Симонов, 2012; новые данные). Обсуждение отмеченных фактов представлено в следующем разделе.
Обсуждение
Плотность населения
Максимальную локальную плотность населения наблюдали на стационаре «Рыбачий» ЗИН РАН на Куршской косе Балтийского моря (см. табл. 1). Такие высокие показатели могут быть объяснены в целом высокой численностью птиц этих видов в регионе, а также их особой концентрацией на Куршской косе и непосредственно на территории стационара «Рыбачий». Куршская коса представляет собой узкую полосу суши между Балтийским морем и Куршским заливом и таким образом в определенной степени является изолированным местообитанием ограниченной площади, что способствует скоплению птиц. Многолетние данные по плотности населения славок в разных биотопах Куршской косы представлены в статье В. А. Паевского (2010). Кроме того, сама территория стационара «Рыбачий», занятая пригодными для гнездования славок биотопами, относительно изолирована от других пригодных биотопов ввиду того, что с двух из четырех сторон ограничена заливом, а с двух других – поселком.
В северных регионах, на периферии видовых ареалов, для которых характерны низкие температуры воздуха, возвраты холодов весной и в начале лета и общая нестабильность погодных условий, численность и плотность населения славок в среднем по региону были сравнительно низки. При этом в условиях в целом низкой популяционной плотности птицы одного вида могли образовывать скопления в наиболее предпочитаемых биотопах с формированием локально высокой плотности населения (см. табл. 1).
Таким образом, за исключением крайней северной периферии ареалов (заселяемой с крайне низкой плотностью), плотности населения значительно варьировали по биотопам – птицы концентрировались в наиболее предпочитаемых местообитаниях, избегая менее характерных для них сплошных массивов леса (см. табл. 1). Полученные данные свидетельствуют о том, что плотность населения птиц в зоне регулярного гнездования зависит от характера местообитаний и степени их пригодности для обитания птиц определенного вида, что согласуется с теорией идеально свободного распределения (Fretwell, Lucas, 1970; Fretwell, 1972). В соответствии с указанной теорией, вероятность заселения каждого местообитания обусловлена степенью его «благоприятности» (которую каждая особь оценивает на основе врожденных возможностей) и числом птиц, желающих его заселить (Fretwell, Lucas, 1970; Pulliam, 1988; Pulliam, Danielson, 1991). При этом обилие претендентов на заселение определенного местообитания также зависит от того, занимает ли оно значительную часть ландшафта или представлено отдельными фрагментами (Beletsky, Orians, 1987; Orians, 1991; Hanski, 2005).
Территориальное поведение птиц в местообитаниях, заселяемых с низкой плотностью
В местообитаниях с низкой локальной плотностью населения славки, как правило, занимали сравнительно крупные по размеру территории (см. рис. 1), находящиеся на значительном расстоянии друг от друга (см. рис. 2). Эти данные согласуются с положением концепции Дж. Л. Брауна (Brown, 1969) о том, что при очень низкой популяционной плотности размер территории не должен ограничиваться конкуренцией за территории. Результаты корреляционного анализа (см. табл. 2, 3) подтверждают наблюдения, свидетельствующие, что у славок с ростом плотности населения обычно происходит сокращение размеров территорий и расстояний между ними. Обратную зависимость между размером территории и популяционной плотностью отмечали для птиц многих видов (Watson, 1965; Holmes, 1970; Morse, 1976; Зимин и др., 1978; Myers et al., 1979; Yamagishi, Ueda, 1986; Both, Visser, 2003; Pons et al., 2008; и др.), хотя такая зависимость наблюдается не всегда (Рябицев, 1993; Both, Visser, 2003). Согласно известному обобщению (Both, Visser, 2003), с увеличением популяционной плотности затраты на защиту территории возрастают, что также приводит к сокращению площади охраняемой территории.
В свою очередь, локальная плотность населения также зависит от пригодности занимаемого местообитания (Fretwell, Lucas, 1970; Fretwell, 1972), которая во многом определяется его структурой, в частности структурой растительности. Многие исследователи (Simms, 1985; Cody, 1985; Van der Hut, 1986; Prescott, 1987; Smith, Shugart, 1987; DeGraaf, 1989; Pons et al., 2008; и др.) указывали на зависимость размеров территорий от структуры растительности (и связанных с ней факторов, включая пищевые ресурсы, выходящие за рамки обсуждения в данной статье). Действительно, в обширных более или менее однородных биотопах районов исследований птицы занимали крупные территории, рассредоточенные по большим площадям.
Также логично предположить, что в более жестких условиях севера птицы вынуждены увеличивать размеры своих территорий в связи с необходимостью обеспечения достаточным количеством пищи. В то же время на севере плотность населения птиц не достигает предельных значений, при которых дальнейшее увеличение размеров территорий становится невозможным. Обратное отношение между количеством корма и размером территории известно для птиц разных видов (Cody, 1985; Newton, 1998; Both, Visser, 2003; и др.), хотя прослеживается не всегда (например, об этом свидетельствуют результаты специальных экспериментов (Yom-Tov, 1974; Franzblau, Collins, 1980)). Кроме того, у многих видов участки, используемые для кормодобывания, по площади превышают демонстрируемые территории. Территориальное поведение воробьиных, как правило, позволяет им добывать корм за пределами своих территорий (Newton, 1998) и даже в пределах чужих территорий конспецифичных особей без агрессивных конфликтов (Рябицев, 1993), что мы также неоднократно наблюдали.
В целом в местообитаниях с низкой плотностью населения славки демонстрировали поведение, типичное для территориальных птиц (Howard, 1920; Ильичев, 1972). Самцы демонстрировали территории посредством пения, соседние территории конспецифичных самцов не перекрывались (рис. 2), случаев длительного пребывания самца на чужой территории своего вида не отмечали.
Однако при этом также не было зарегистрировано фактов агрессивных столкновений на границах территорий. Возможно, такой элемент поведения, как защита территориальных границ посредством нападения на чужого самца, был замещен акустической и, вероятно, визуальной сигнализацией. Такое замещение может быть допустимо в малонасыщенных поселениях с достаточным количеством мест гнездования. Также возможно, что рассредоточенное (не плотное) распределение территорий в пространстве ведет к уменьшению числа встреч и агрессивных столкновений конспецифичных самцов друг с другом и в результате способствует общему снижению затрат времени и энергии птиц. Последнее наиболее актуально в сравнительно неблагоприятных условиях среды (частые дожди и низкие температуры), характерных для северных районов.
С другой стороны, логично предположить, что редукция строгой территориальности допустима в малонасыщенных поселениях, формируемых на севере, где столкновения и взаимное влияние друг на друга у конспецифичных самцов наблюдаются реже ввиду их низкой численности.
Помимо редукции агрессивности и степени защиты территорий на северных участках исследований было отмечено снижение активности и продолжительности пения птиц изучаемых видов. Последнее было характерно для большинства территориальных самцов, привлекших самок и образовавших пары. Самцы, остававшиеся холостыми, как в случае их длительного пребывания на контролируемых участках, так и тогда, когда они покидали эти участки спустя 2–3 дня после появления, как правило, сохраняли более высокие показатели песенной активности. Наличие холостых взрослых самцов в изучаемых малонасыщенных поселениях северных регионов обычно связывают с наблюдаемым здесь дефицитом самок, типичным и для местных славковых (Лапшин, 2012; Лапшин и др., 2012, 2015).
В целом подобную общую редукцию территориальности здесь также отмечали у пеночек (Лапшин, 1978, 2016; Lapshin, 2005; Matantseva et al., 2015). Предположительно, редукция территориальности в условиях низкой социальной напряженности в разреженных северных поселениях направлена на оптимизацию бюджета времени и энергии птиц, обитающих в сравнительно жестких условиях севера.
Территориальное поведение птиц в местообитаниях, заселяемых с высокой плотностью
Наиболее компактные пространственно-территориальные структуры были сформированы в самых населенных участках наиболее благоприятных биотопов, имеющих ограниченную площадь (ограничены водными пространствами и нетипичными местообитаниями) (см. рис. 1, 2). Размеры самых маленьких территорий в ряде случаев, особенно в условиях высокомозаичной среды, были ограничены небольшой площадью занимаемой стации. В таких случаях границы территории и стации пространственно совпадали. Совмещение территориальных границ с естественными ландшафтными маркерами было отмечено для птиц разных видов (например, Рыжановский, 1977; Виноградова, 1986).
В таких плотных поселениях, особенно в условиях дефицита гнездопригодных мест, отмечали нетипичные для строго территориальных видов формы поведения: перекрывание соседних территорий по горизонтали; возможное разделение территориальных пространств по вертикали; значительная редукция активности пения и взаимной агрессии конспецифичных самцов; замена типичной территориальности системой иерархических отношений (см. табл. 4). Подобные примеры замещения типичной территориальности системой иерархических отношений в определенных ситуациях были отмечены для животных разных видов (Davis, 1958; Dewsbury, 1978; Рябицев, 1993; Maher, Lott, 1995; Newton, 1998; и др.), в том числе и для воробьиных птиц. В частности, сравнительно редкие случаи перекрывания территорий в различной степени наблюдали у славок (Pons, 1996; Зубцовский, Матанцев, 2004; Bas et al., 2005), коньков (Ryzhanovsky, 1977), овсянок (Колпакова, 2006). Частичное перекрывание территорий было описано даже для таких строго территориальных птиц, как пеночки рода Phylloscopus, в условиях высокой локальной плотности населения, особенно при наличии дефицита мест, пригодных для гнездования (Рябицев, 1993; Matantseva et al., 2015). Факты защиты территорий с одной стороны и отсутствия такой защиты с других сторон известны для каменок (Панов, Иваницкий, 1975; Панов, 1978, 1983). Для разных видов воробьиных отмечены случаи, когда конспецифичные самцы-соседи (владельцы соседних территорий) не нападали друг на друга и даже совместно защищали свои территории от чужих самцов (например, Благосклонов, 1977; Рябицев, 1993). К заключениям о том, что защита территорий у птиц может угасать в условиях дефицита гнездопригодных мест, приходили и другие исследователи (Both, Visser, 2003).
Данные специальных исследований показывают, что, поскольку самцы кормятся преимущественно на своих территориях (хотя и не всегда, как уже было отмечено), защита территорий может проявляться только в условиях умеренного обилия корма и редуцироваться в случаях слишком больших или слишком малых его количеств (Gill, Wolf, 1975; Carpenter, MacMillen, 1976; Kodric-Brown, Brown, 1978; Van Riper, 1984). У многих воробьиных территориальная агрессивность может снижаться (вплоть до нуля) в периоды выкармливания птенцов и слетков и даже раньше, вскоре после формирования пар (например, Borowiec, 1992; Рябицев, 1993), что наблюдали и мы. Кроме того, сравнительно плохая видимость в условиях плотной вегетации также способствует снижению взаимной агрессии между соседями (Рябицев, 1993). В. В. Иваницкий (1989) писал о сложности структуры территориальности и выражал мнение о том, что только в поселениях с очень высокой плотностью территория обладает характеристиками, приписываемыми ей типологической моделью территориальности (несжимаемость, четко определяемые контуры, равномерное использование).
Крайним проявлением наблюдаемой нами редукции территориальности являются отмеченные факты гнездования пар, в которых у самцов вообще не было зафиксировано проявления песенной активности (и агрессивного поведения), и, следовательно, они не обладали собственными территориями. Известны и другие данные о том, что у птиц некоторых видов самцы-доминанты занимают территории, а рецессивы – нет, при этом нетерриториальные самцы в редких случаях могут участвовать в размножении, хотя это крайне не характерно для территориальных животных (обзор: Newton, 1998). Согласно классическим представлениям (Brown, 1969), особи защищают территории тогда, когда выгоды от территории превышают затраты на ее охрану.
Особо следует обсудить и тот факт, что в условиях высокой плотности населения некоторые самцы занимали стации, нетипичные для гнездования птиц этих видов, и не привлекали самок. Небольшая их часть оставалась холостыми, задерживаясь на долгое время на занятых территориях. Однако большинство таких особей, спустя несколько дней пения на контролируемых участках без привлечения самки, эти участки покидали. По-видимому, таких особей следует рассматривать как «блуждающих» (non-breeding floaters) согласно модели Дж. Л. Брауна (Brown, 1969). «Блуждающие» особи также вносили вклад в повышение пластичности формируемых территориальных структур.
Редукция территориальности и пластичность территориальных структур в условиях высокой плотности населения, по-видимому, обусловливают возможность принять участие в репродукции большему числу особей популяции. Действительно, в условиях жестких территориальных структур, несжимаемых контуров территорий и предельно малых расстояний между ними, вселение новых самцов с собственными территориями на участки с уже распределенными территориями «типичных» размеров и на «типичном» расстоянии друг от друга было бы невозможно. Однако мы неоднократно наблюдали вселение новых самцов в биотопы с уже распределенными территориями в результате смещения этих территорий в пространстве и сокращения их размеров, а также изменения поведения птиц, в т. ч. и за счет смены типичной территориальности системой иерархических отношений.
Заключение
Полученные результаты свидетельствуют о том, что территориальное поведение славок пластично и зависит от плотности населения, которая, в свою очередь, во многом определяется характером занимаемых местообитаний. В частности, размеры территорий и расстояния между соседними территориями конспецифичных самцов, как правило, уменьшаются с ростом плотности населения, а также могут быть ограничены площадью занимаемых стаций.
В ограниченных по площади предпочитаемых биотопах территориальные структуры становятся более компактными и лабильными. Максимальное разнообразие вариантов проявления территориального поведения наблюдается в самых плотных поселениях птиц.
В условиях очень высокой и очень низкой популяционной плотности происходила редукция территориальности (сокращение периода демонстрирования территорий посредством пения, снижение активности пения, редукция агрессивности и степени защиты территорий). Несмотря на внешнее сходство, причины этого были различны. Редукция территориальности и пластичность территориальных структур, наблюдаемые при высокой плотности населения, по-видимому, обусловливают возможность принять участие в репродукции большему числу особей популяции.
Редукция строгой территориальности в условиях очень низкой плотности населения, наблюдаемая на севере видовых ареалов, по-видимому, допустима в низко насыщенных поселениях, где столкновения и взаимное влияние друг на друга конспецифичных самцов наблюдаются реже в виду их низкой численности. Таким образом, возможная в условиях низкой социальной напряженности в разреженных северных поселениях редукция территориальности, предположительно, направлена на экономию бюджета времени и энергии птиц, обитающих в сравнительно более жестких условиях.
На основании полученных данных можно заключить, что внутривидовые различия в территориальном поведении птиц способствуют максимальной реализации репродуктивного потенциала особей в конкретных условиях обитания, а пластичность территориального поведения является одним из механизмов адаптации птиц к обитанию в различных условиях среды, что позволяет им иметь обширные ареалы.
Библиография
Благосклонов К. Н. Экспериментальный анализ территориального поведения птиц // Адаптивные особенности и эволюция птиц. М.: Наука, 1977. С. 128–134.
Виноградова Н. В. Гнездовая территориальность ястребиной славки на Куршской косе // Орнитология. 1986. № 21. С. 24–30.
Зимин В. Б., Лапшин Н. В., Хохлова Т. Ю. Биология размножения садовой славки в Карелии // Фауна и экология птиц и млекопитающих таежного Северо-Запада СССР. Петрозаводск, 1978. С. 5–16.
Зубцовский Н. Е., Матанцев В. А. Итоги орнитологических исследований кафедры зоологии за последнее десятилетие // Вестник Удмуртского университета. 1992. № 3. C. 101–106.
Зубцовский Н. Е., Матанцев В. А. Стационарные популяционно-экологические исследования в островных местообитаниях как перспективный подход к решению некоторых задач популяционной биологии // Методы популяционной биологии: Материалы докл. VII Всерос. популяц. семинара. Сыктывкар, 2004. С. 131–136.
Зубцовский Н. Е., Матанцев В. А., Тюлькин Ю. А. Некоторые итоги изучения структуры и экологии птиц малых сообществ в УАССР // Экология птиц Волжско-Уральского региона (Информационные материалы). Свердловск: Изд-во УрО АН СССР, 1988. С. 33–35.
Зубцовский Н. Е., Матанцев В. А., Матанцева М. В. Этологическая лабильность птиц рода Sylvia как механизм обеспечения стабильности локальных популяций // Развитие современной орнитологии в Северной Евразии: Труды XII Междунар. орнитол. конф. Сев. Евразии. Ставрополь: Изд-во СГУ, 2006. С. 587–600.
Иваницкий В. В. Индивидуальное пространство у птиц: структурно-функциональные и экологические аспекты // Зоол. журн. 1989. Т. 68. № 4. С. 23–94.
Ильичев В. Д. Биоакустика птиц . М.: Изд-во МГУ, 1972. 286 с.
Колпакова Т. Ю. Пространственные взаимоотношения обыкновенных овсянок (Emberiza citrinella L.) в лесостепной зоне Омской области // Популяционная экология животных. Томск: Изд-во Томского ун-та, 2006. С. 135–137.
Лапшин Н. В. Поведение пеночки-веснички в предгнездовый период // Фауна и экология птиц и млекопитающих таежного Северо-Запада СССР. Петрозаводск, 1978. С. 32–39.
Лапшин Н. В. Соотношение полов у пеночек (Phylloscopus) в Карелии // Экология, эволюция и систематика животных: Материалы Междунар. науч.-практ. конф. Рязань: НП «Голос губернии», 2012. С. 293–295.
Лапшин Н. В. Особенности предгнездового поведения пеночек рода Phylloscopus в северной части ареала // Вестник Санкт-Петербургского университета. 2016. Сер. 3. Биология. Вып. 1. С. 100–115.
Лапшин Н. В., Топчиева Л. В., Матанцева М. В., Симонов С. А., Малышева И. Е., Токарева Н. П. Особенности экологии мигрирующих воробьиных птиц (анализ с применением методов молекулярной биологии) // Поволжский экологический журнал. 2012. № 1. С. 53–64.
Лапшин Н. В., Матанцева М. В., Симонов С. А. Особенности популяционно-демографической структуры популяций пеночек-весничек (Phylloscopus trochilus) в Карелии: дисбаланс в соотношении полов и полигамия // Энергетика и годовые циклы птиц (памяти В. Р. Дольника): Материалы Междунар. конф. Звенигородская биологич. станция МГУ, 24–29 сентября 2015 г. М.: Товарищество научных изданий КМК, 2015. С. 170–176.
Матанцева М. В. Эколого-этологические механизмы поддержания стабильности поселений славок Sylvia и пеночек: Дис. … канд. биол. наук. Phylloscopus . Екатеринбург, 2010. 282 с.
Матанцева М. В., Симонов С. А. Эколого-этологическая характеристика поселений славок (Sylvia) в мозаичных местообитаниях Куршской косы Балтийского моря // Экология. 2008. N 5. С. 373–378.
Матанцева М. В., Симонов С. А. Особенности территориального поведения славок (Sylvia) на северной периферии ареала (южная Карелия) // Экология. 2012. № 3. С. 204–209.
Паевский В. А. Биология размножения, морфометрия и популяционная динамика пяти видов славок рода Sylvia на Куршской косе (Восточная Прибалтика) // Русский орнитологический журнал. 2010. Т. 19. Экспресс-выпуск 602. С. 1783–1799.
Панов Е. Н. Механизмы коммуникации у птиц . М.: Наука, 1978. 306 с.
Панов Е. Н. Поведение животных и этологическая структура популяций . М.: Наука, 1983. 424 с.
Панов Е. Н., Иваницкий В. В. Межвидовые территориальные отношения в смешанной популяции чернобокой каменки Oenanthe finschii и каменки-плешанки O. pleshanka на полуострове Мангышлак // Зоол. журн. 1975. Т. 54. № 9. С. 1357–1370.
Рябицев В. К. Территориальные отношения и динамика сообществ птиц в Субарктике . Екатеринбург: Наука, Уральское отделение, 1993. 296 с.
Рыжановский В. Н. Экология лугового и краснозобого коньков на Ямале // Сравнительная экология фоновых видов птиц Ямальской тундры. Свердловск, 1977. С. 3–35.
Bas J. M., Pons P., Gómez C. Home range and territory of the Sardinian Warbler Sylvia melanocephala in Mediterranean shrubland // Bird Study. 2005. Vol. 52. P. 137–144.
Beletsky L. D., Orians G. H. Territoriality among Male Red-Winged Blackbirds: II. Removal Experiments and Site Dominance // Behavioral Ecology and Sociobiology. 1987. Vol. 20. № 5. P. 339–349.
Borowiec M. Breeding ethology and ecology of the Reed Warbler, Acrocephalus scirpaceus (Hermann, 1804) at Milicz, SW Poland // Acta Zoologica Cracoviensis. 1992. Vol. 35. P. 315–350.
Both C., Visser M. E. Density dependence, territoriality, and divisibility of resources: from optimality models to population processes // The American Naturalist. 2003. Vol. 161. № 2. P. 326–336.
Brown J. L. Territorial Behavior and Population Regulation in Birds: A Review and Re-Evaluation // The Wilson Bulletin. 1969. Vol. 81. № 3. P. 293–329.
Carpenter F. L., MacMillen R. E. Threshold model of feeding territoriality and test with a Hawaiian Honeycreeper // Science. 1976. Vol. 194. P. 639–642.
Cody M. L. (Ed.). Habitat selection in birds. Orlando, Florida: Academic press, 1985. 558 p.
Davis D. E. The role of density in aggressive behaviour of house mice // Animal Behaviour. 1958. Vol. 6. P. 207–210.
DeGraaf R. M. Territory sizes of Song Sparrows, Melospiza melodia, in rural and suburban // Canadian Field Naturalis. 1989. Vol. 103. № 1. P. 43–47.
Dewsbury D. A. Comparative animal behavior. New York: McGraw Hill, 1978. 452 p.
Franzblau M. A., Collins J. P. Test of a hypothesis of territory regulation in an insectivorous bird by experimentally increasing prey abundance // Oecologia. 1980. Vol. 46. № 2. P. 164–170.
Fretwell S. D. Populations in a seasonal environment. Princeton, NJ: Princeton University Press, 1972. 217 p.
Fretwell S. D., Lucas H. L. On territorial behaviour and other factors influencing habitat distribution in birds // Acta Biotheoretica. 1969. Vol. 19. № 1. P. 16–36.
Fretwell S. D., Lucas H. L. On territorial behavior and other factors influencing habitat distribution in birds. I. Theoretical Development // Acta Biotheoretica. 1970. Vol. 19. P. 16–36.
Gill F. B., Wolf L. L. Economics of feeding territoriality in the Golden-Winged Sunbird // Ecology. 1975. Vol. 56. P. 333–345.
Hanski I. The shrinking world: ecological consequences of habitat loss // Excellence in Ecology. Book 14 / Ed. Kinne O. Oldendorf. Luhe: Internat. Ecol. Inst., 2005. 307 p.
Holmes R. T. Differences in population density, territoriality, and food supply of dunlin on Arctic and Subarctic tundra // Anim. Pop. Relat. Food Resour. 1970. P. 303–317.
Howard H. E. Territory in Bird Life. London: John Murray, Albemarle Street, W., 1920. 386 p.
Kodric-Brown A., Brown J. H. Influence of economics, interspecific competition and sexual dimorphism on territoriality of migrant Rufous Hummingbirds // Ecology. 1978. Vol. 59. P. 285–296.
Lapshin N. V. Biology of the Wood Warbler Phylloscopus sibilatrix (Berchst.) in the taiga zone of north-western Russia // Avian Ecol. Behav. 2005. Vol. 13. P. 25–46.
Maher C. R., Lott D. E. Definitions of territoriality used in the study of variation in vertebrate systems // Animal Behaviour. 1995. Vol. 49. № 6. P. 1581–1597.
Matantseva M. V., Lapshin N. V., Simonov S. A. Ethological aspects of biodiversity within and between Phylloscopus species: behavioural variation among birds from the centre and periphery of breeding ranges // Acta Biologica Universitatis Daugavpiliensis. 2015. Vol. 15. № 1. P. 273–283.
Morse D. H. Variables affecting the density and territory size of breeding spruce-woods warblers // Ecology. 1976. Vol. 57. № 2. P. 290–301.
Myers J. P., Connors P. G., Pitelka F. A. Territory size in wintering sanderlings: the effects of prey abundance and intruder density // Auk. 1979. Vol. 96. P. 551–561.
Newton I. Population Limitation in Birds. London: Academic Press, 1998. 597 p.
Nice M. The role of territory in bird life // Amer. Midl. Nat. 1941. № 26. P. 441–448.
Noble G. K. Dominance in the life of birds // Auk. 1939. Vol. 56. Р. 263–273.
Odum E., Kuenzler R. Measurement of territory and home range size in birds // Ari. 1955. Vol. 72. № 2. P. 128–137.
Orians G. H. Preface // Amer. Nat. 1991. Vol. 137. Suppl. P. S1–S4.
Pons P. Dynamique de l’avifaune après incendie et brûlage dirigé en zone méditerranéenne: importance de la territorialité. Thèse de Doctorat, Spécialité Ecologie. Paris, 1996. 293 p.
Pons P., Bas J. M., Prodon R., Roura-Pascual N., Clavero M. Territory characteristics and coexistence with heterospecifics in the Dartford warbler Sylvia undata across a habitat gradient // Behavioral Ecology and Sociobiology. 2008. Vol. 62. P. 1217–1228.
Prescott D. R. C. The effects of habitat density and the spatial distribution of food on the social behaviour of captive wintering American tree sparrows // Canadian Journal of Zoolog. 1987. Vol. 65. P. 522–526.
Pulliam H. R. Sources, sinks, and population regulation // American Naturalist. 1988. Vol. 132. № 5. P. 652–661.
Pulliam H. R., Danielson B. J. Sources, sinks, and habitat selection: A landscape perspective on population dynamics // American Naturalist. 1991. Vol. 137. Supple. P. S50–S66.
Simms E. British Warblers. London: Collins, 1985. 432 p.
Shirihai H., Gargallo G., Helbig A. J., Harris A. Sylvia warblers: identification, taxonomy and phylogeny of the genus Sylvia. London: A & C Black, 2010. 576 p.
Smith T. M., Shugart H. H. Territory size variation in the Ovenbird: the role of habitat structure // Ecology. 1987. Vol. 68. P. 695–704.
Titus R. C., Hans C. A. Singing behavior of American Robins in linear and non-linear habitats // Wilson Bull. 1990. Vol 102. № 2. P. 325–328.
Van der Hut R. Habitat choice and temporal differentiation in reed passerines of a Dutch marsh // Ardea. 1986. Vol. 74. P. 159–176.
Van Riper C. 3rd. The influence of nectar resources on nesting success and movement patterns of the Common Amakihi (Hemignathus virens) // The Auk. 1984. Vol. 101. № 1. P. 38–46.
Watson A. A population study of ptarmigan (Lagopus mutus) in Scotland // Journal of Animal Ecology. 1965. Vol. 34. № 1. P. 135–172.
Yamagishi S., Ueda K. Simultaneous territory mapping of male Fan-tailed Warblers (Cisticola juncidis) // Journal of Field Ornithology. 1986. Vol. 57. P. 193–260.
Yom-Tov Y. The effect of food and predation on breeding density and success, clutch size and laying date of the Crow (Corvus corone L.) // Journal of Animal Ecology. 1974. Vol. 43. № 2. P. 479–498.
Благодарности
Выражаем искреннюю благодарность руководству и сотрудникам Института биологии КарНЦ РАН, биологической станции «Рыбачий» ЗИН РАН, станции «Лувеньга» Кандалакшского заповедника и национального парка «Смольный» за обеспечение возможности проведения исследований и создание благоприятных условий проживания на их базах и территориях. Мы также очень признательны рецензентам за рецензирование статьи и ценные рекомендации. Исследования проведены частично в рамках темы № 0221-2014-0037 «Динамика фауны птиц и млекопитающих Европейского Севера России в условиях интенсивных антропогенных и климатических изменений: мониторинг, управление и сохранение биоразнообразия» (№ г.р. АААА-А17-117022850042-8), частично – при поддержке РФФИ (проекты №№ 06-05-64368_а, 12-04-31872_мол_а и 15-05-03493_а).

 © 2011 - 2026
© 2011 - 2026