



Введение
В Карелии большая часть работ по биологии сига, в том числе и по питанию рыб, выполнена на крупных промысловых водоемах, в частности Ладожском и Онежском озерах, Сямозере, Водлозере, озерно-речной системе р. Каменная (Первозванский, 1986; Дятлов, 2002; Петрова, Кудерский, 2006; Стерлигова и др., 2016). Установлено, что в больших и глубоких озерах, как правило, сосуществуют несколько экологических форм сига, приспособленных к использованию различных кормовых объектов – от планктона до бентоса, от икры и мелких рыб до летающих взрослых насекомых. В более однородных условиях встречается лишь одна форма сига, а в малых водоемах, имеющих низкую продуктивность и бедную кормовую базу, конкуренция может обостряться, и не только на внутривидовом уровне, но и с другими видами рыб. Так известно, что в условиях недостатка пищевых ресурсов такой широко распространенный хищник, как речной окунь (Perca fluviatilis Linnaeus, 1758), мельчает и переходит к питанию беспозвоночными, составляя конкуренцию видам-бентофагам (Биология речного окуня, 1993) – в карельских условиях, прежде всего малотычинковому сигу. Возникающая конкурентная напряженность может негативным образом сказываться на численности популяций последнего и приводить к снижению его доли в уловах, снижая тем самым привлекательность небольших водоемов для любительского рыболовства.
В данной работе предпринята попытка оценить степень перекрывания пищевых ниш у сига и окуня, на примере одного из малых лесных озер Северной Карелии определить возможность возникновения конкурентных условий в условиях конкретного водоема.
Материалы
Питание рыб изучали в малом лесном оз. Падашулкаярви, на северо-западе Республики Карелия. Данный водоем относится к бассейну Белого моря и входит в озерно-речную систему реки Писта, его протяженность составляет около 1.5 км, ширина не превышает 0.6 км, а глубина – 6 м. По своим характеристикам это типичное олиготрофное озеро с прозрачной водой, каменистым дном и развитыми илистыми отложениями. Берега обрывистые, песчано-галечные, сплошь покрытые лесом. Рыбу вылавливали в мае 2016 года, использовали сети с размером ячеи от 20 до 40 мм. Для уменьшения влияния на данные смены сезонных аспектов у пищевых объектов, прежде всего у насекомых, для анализа была взят только один, наиболее крупный, улов, полученный в течение суток (16 мая). Исследуемая выборка включала 52 окуня и 20 сигов.
Методы
У выловленных рыб измеряли длину и массу тела. Извлеченные желудки фиксировали 4 % раствором формалина. В лабораторных условиях определяли массу пищевого комка, подсчитывали число особей, встречаемость и долю кормовых объектов. В качестве индекса наполнения желудка использовали отношение массы пищевого комка к массе рыбы, умноженной на 10 (Методическое пособие..., 1974).
Для полученных выборок вычисляли основные статистические показатели и проверяли эмпирические частоты на соответствие нормальному распределению (критерий Шапиро – Уилкса). Межвидовые отличия средних устанавливали по критерию Стьюдента (t) с поправкой Уэлча для неравных дисперсий, дисперсии сравнивали с помощью критерия Фишера (F). Поскольку распределение частот соответствовало нормальному закону не во всех случаях, для оценки уровня значимости отличий (p) использовали один из методов численного ресамплинга – перестановочный тест Монте-Карло. При таком подходе p определяется не теоретическим (нормальным) распределением, а эмпирическим распределением статистик t (или F), полученных при попарном сравнении множества выборок (10 тыс.), сгенерированных путем случайных перестановок имеющихся вариант.
Для каждого объекта питания рассчитывали индекс относительной значимости (Попова, Решетников, 2011):
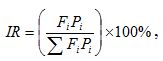
где Fi – встречаемость i-го объекта (% пищевых комков, содержащих данный объект), Pi – доля i-го объекта по массе (% от общей массы пищевых комков). Межвидовую дифференциацию индекса оценивали методом углов φ (Плохинский, 1970). В основу данного подхода положено угловое преобразование долей и использование критерия Стьюдента (t) в качестве меры статистической значимости отличий. Общность пищевых ниш у окуней и сигов измеряли с помощью индекса Мориситы – Хорна (Horn, 1966):
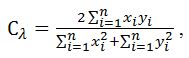
где xi – доля i-го корма у одного вида, уi – доля того же корма у другого вида. Данный индекс принимает значения в интервале от 0 до 1. Равенство нулю означает отсутствие совпадений, единица – это полное перекрывание ниш. В качестве биологически значимого показателя принято расценивать величину Сλ > 0.6.
Изменчивость питания рыб исследовали методом главных компонент (Коросов, Горбач, 2010). Исходная матрица состояла из n объектов (здесь – рыб), каждый из которых охарактеризован m признаком – предварительно прологарифмированным долям объектов питания по массе (Pi). Характер отличия объектов раскрывает их взаимное расположение на плоскости главных компонент (Ивантер, Коросов, 2014). Полученные значения главных компонент использовали в качестве интегральных характеристик питания рыб. Межвидовые отличия устанавливали по критерию Уилкоксона – Манна – Уитни, зависимость состава пищи от размера рыб – с помощью коэффициента корреляции Спирмена (rs). Данный коэффициент принимает значения от –1 до +1. Знак перед коэффициентом вскрывает направленность связи, о силе связи судят по величине коэффициента: значение rs > |0.6| указывает на тесную связь, rs > |0.3| – на среднюю и rs < |0.3| – на слабую корреляцию. Значения критериев представлены в асимптотическом формате (z), уровень их значимости оценивали с помощью перестановочного теста Монте-Карло.
Статистическую обработку данных проводили в средах Past 3.15 (Hammer et al., 2001) и R 3.3.1 (Hothorn et al., 2010).
Результаты
Выловленные сиги превосходили окуней по длине и массе тела (табл. 1). Число рыб с пустыми желудками составило 7 и 1 у окуней и сигов соответственно. В желудках других рыб обнаружены представители двух групп зоопланктона – ветвистоусые и веслоногие ракообразные, шести групп зообентоса – моллюски, личинки и куколки комаров-звонцов, личинки стрекоз, ручейников и мокрецов, нимфы поденок, а также остатки взрослых, летающих насекомых. Установлено, что и окуни, и сиги употребляют в пищу все перечисленные организмы (табл. 2, рис. 1). Вместе с тем сравнение индексов относительной значимости IR выявило значимую межвидовую дифференциацию в трофических предпочтениях. Так, основную часть рациона окуней составляют личинки крупных насекомых, прежде всего ручейников, а сиги специализируются на моллюсках. Полученные результаты подтверждаются и значением индекса Мориситы – Хорна Сλ = 0.5, указывающего на слабое перекрывание пищевых ниш окуней и сигов в оз. Падашулкаярви.
Таблица 1. Основные статистические показатели исследованных выборок
Table 1. The main statistical parameters of the samples
| Показатель | n | min | max | M | S | Me | W | p |
| Речной окунь – Perca fluviatilis | ||||||||
| Длина тела, см | 52 | 10 | 27 | 18.1 | 4.8 | 18.6 | 0.94 | 0.017 |
| Масса тела, г | 52 | 45 | 179 | 89.8 | 23.6 | 89.0 | 0.94 | 0.006 |
| Масса пищевого комка, мг | 45 | 32 | 940 | 352.7 | 236.6 | 320.0 | 0.94 | 0.018 |
| Индекс наполнения желудка, ‰ | 45 | 3 | 85 | 40.1 | 23.5 | 37.0 | 0.96 | 0.092 |
| Возраст, лет | 52 | 2+ | 8+ | |||||
| Обыкновенный сиг – Coregonus lavaretus | ||||||||
| Длина тела, см | 20 | 24 | 35 | 30.2 | 3.1 | 31.0 | 0.94 | 0.251 |
| Масса тела, г | 20 | 100 | 300 | 217.0 | 56.9 | 217.5 | 0.95 | 0.330 |
| Масса пищевого комка, мг | 19 | 70 | 2980 | 629.4 | 632.7 | 430.0 | 0.61 | <0.001 |
| Индекс наполнения желудка, ‰ | 19 | 7 | 117 | 27.1 | 23.7 | 21.0 | 0.67 | <0.001 |
| Возраст, лет | 20 | 2+ | 4+ | |||||
Примечание. n – объем выборки, min и max – минимальное и максимальное значения вариант, M – среднее арифметическое, S – стандартное отклонение, Me – медиана, W – значение критерия Шапиро – Уилкса, p – значимость отличий от нормального распределения частот, значимые отличия указаны жирным шрифтом.
Таблица 2. Спектры питания рыб
Table 2. Spectra of fish nutrition
| Вид рыб | Окунь | Сиг | Отличия IR | |||||
| Состав пищи | F | P | IR | F | P | IR | t | p |
| Ветвистоусые ракообразные – Cladocera | 17 | 12 | 8 | 30 | 18 | 15 | 1.60 | 0.116 |
| Веслоногие ракообразные – Copepoda | 8 | 5 | 2 | 30 | 11 | 9 | 2.30 | 0.034 |
| Личинки комаров-звонцов – Chironomidae | 25 | 5 | 5 | 25 | 8 | 6 | 0.31 | 0.374 |
| Куколки комаров-звонцов – Chironomidae | 38 | 11 | 17 | 25 | 8 | 6 | 2.51 | 0.023 |
| Нимфы поденок – Ephemeroptera | 17 | 8 | 5 | 15 | 3 | 1 | 1.77 | 0.085 |
| Личинки мокрецов – Heleidae | 80 | 4 | 13 | 20 | 2 | 1 | 3.80 | 0.001 |
| Моллюски – Mollusca | 6 | 4 | 1 | 55 | 36 | 56 | 10.54 | <0.001 |
| Личинки стрекоз – Odonata | 25 | 17 | 17 | 15 | 10 | 4 | 3.16 | 0.006 |
| Личинки ручейников – Trichoptera | 27 | 20 | 21 | 10 | 2 | 1 | 5.32 | <0.001 |
| Летающие имаго насекомых – Insecta | 21 | 14 | 12 | 10 | 2 | 1 | 3.59 | 0.002 |
Примечание. F – встречаемость остатков кормового объекта в пищевом комке (%), P – их доля от массы пищевого комка (%), IR – индекс относительной значимости объекта в питании рыб (%), t – значение критерия Стьюдента, p – значимость отличий долей, значимые отличия указаны жирным шрифтом.
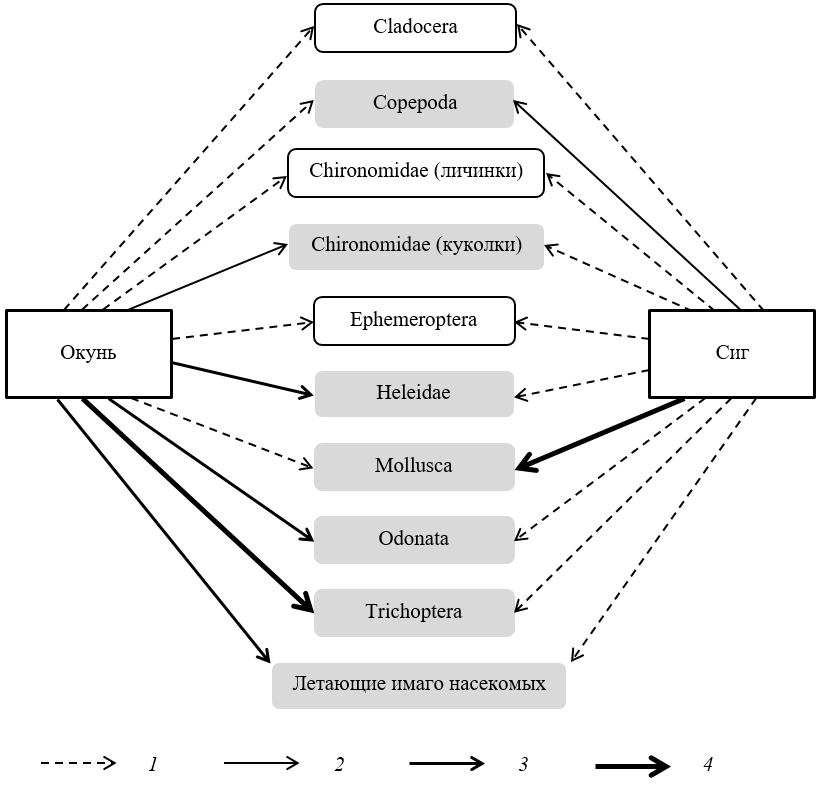
Рис. 1. Пищевые связи окуня и сига в оз. Падашулкаярви. Стрелками указан спектр пищевых связей: 1 – возможные объекты питания, 2–4 – основные виды корма, классифицированные по мере увеличения предпочтительности
Fig. 1. Food connections between perch and whitefish in Padashulkayarvi lake. The arrows indicate the range of food links: 1 – possible objects of nutrition, 2–4 – the main types of food classified with increasing preference
Компонентный анализ выявил три значимые главные компоненты, описывающие основные направления изменчивости трофических связей окуней и сигов. Эти компоненты, учитывающие 54 % общей дисперсии выборки (табл. 3), отражают самые существенные отличия в питании рыб. Первая компонента отвечает за общее смещение рациона рыб по мере их роста. Максимальные отрицательные значения факторных нагрузок имеют мелкие ракообразные и личинки мокрецов – корм молодых рыб, тогда как объекты питания крупных рыб – преимагинальные стадии комаров звонцов и личинки ручейников – получили максимальные положительные значения коэффициентов. Дифференциация значений главных компонент между окунями и сигами здесь достаточно высока, но статистически не значима. Вместе с тем результаты корреляционного анализа, характеризующие связь этих значений с размерами рыб, позволяют заключить, что определяющий вклад в дисперсию первой компоненты внесли окуни: изменчивость именно их рациона выявляется здесь более отчетливо. Вторая компонента обозначила группу рыб, в содержимом желудков которых преобладали личинки стрекоз и нимфы поденок. Межвидовые отличия здесь не выражены совсем, но для окуней остается важной размерная составляющая – с этим видом корма в состоянии справиться только достаточно крупные рыбы. Порядок расположения рыб в осях первых двух компонент (рис. 2) иллюстрирует два обозначенных выше тренда смены зоопланктона на зообентос в рационе питания окуней и сигов. Межвидовая дифференциация рыб наиболее отчетливо проявилась в третьей компоненте (табл. 3, рис. 2). Здесь моллюски – основной корм сигов, получивший максимальное в абсолютном выражении значение факторной нагрузки, противопоставлен летающим насекомым – одному из предпочитаемых объектов питания окуней. Таким образом, эти кормовые объекты выступили в качестве важнейших индикаторов расхождения трофических ниш окуней и сигов в оз. Падашулкаярви.
Таблица 3. Факторные нагрузки и дисперсии значимых главных компонент (ГК), межвидовая дифференциация и корреляция с длиной рыб значений главных компонент
Table 3. The factor loalings and dispersion of significant principal components, the differentiation between species and correlations of fish length with the values of the principal components
| № | Факторные нагрузки | ГК-1 | ГК-2 | ГК-3 |
| 1 | Ветвистоусые ракообразные – Cladocera | -0.401 | -0.348 | 0.131 |
| 2 | Веслоногие ракообразные – Copepoda | -0.433 | -0.238 | 0.052 |
| 3 | Личинки комаров-звонцов – Chironomidae | 0.458 | -0.315 | -0.117 |
| 4 | Куколки комаров-звонцов – Chironomidae | 0.468 | -0.205 | 0.138 |
| 5 | Нимфы поденок – Ephemeroptera | 0.151 | 0.403 | 0.133 |
| 6 | Личинки мокрецов – Heleidae | -0.282 | -0.090 | 0.053 |
| 7 | Моллюски – Mollusca | -0.043 | 0.193 | -0.787 |
| 8 | Личинки стрекоз – Odonata | 0.019 | 0.498 | -0.088 |
| 9 | Личинки ручейников – Trichoptera | 0.344 | -0.277 | 0.050 |
| 10 | Летающие имаго насекомых | 0.016 | 0.385 | 0.545 |
| Дисперсия | 2.33 | 1.91 | 1.22 | |
| Дисперсия, % | 23 | 19 | 12 | |
| Комулята, % | 23 | 42 | 54 | |
| Межвидовая дифференциация значений компонент Критерий Уилкоксона – Манна – Уитни (z) | 1.65 | 0.07 | 2.82 | |
| Значимость отличий (p) | 0.101 | 0.948 | 0.004 | |
| Связь значений компонент с длиной окуней | ||||
| Коэффициент корреляции Спирмена (rs) | 0.45 | 0.37 | -0.01 | |
| Асимптотический критерий z | 2.99 | 2.44 | 0.06 | |
| Значимость связи (p) | 0.002 | 0.016 | 0.950 | |
| Связь значений компонент с длиной сигов | ||||
| Коэффициент корреляции Спирмена (rs) | 0.45 | 0.02 | -0.21 | |
| Асимптотический критерий z | 1.91 | 0.08 | 0.87 | |
| Значимость связи (p) | 0.532 | 0.940 | 0.399 | |
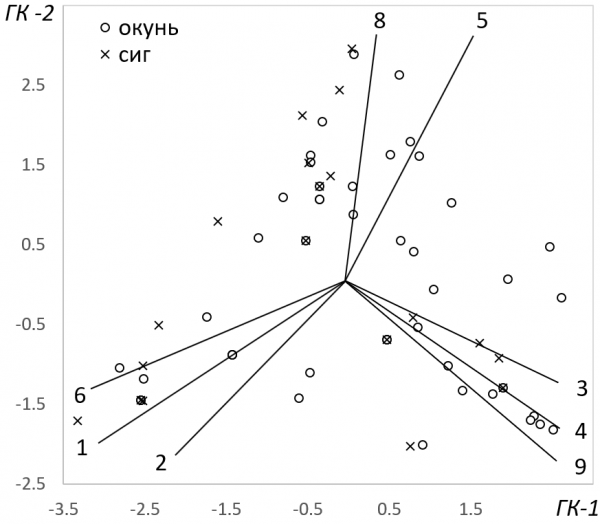
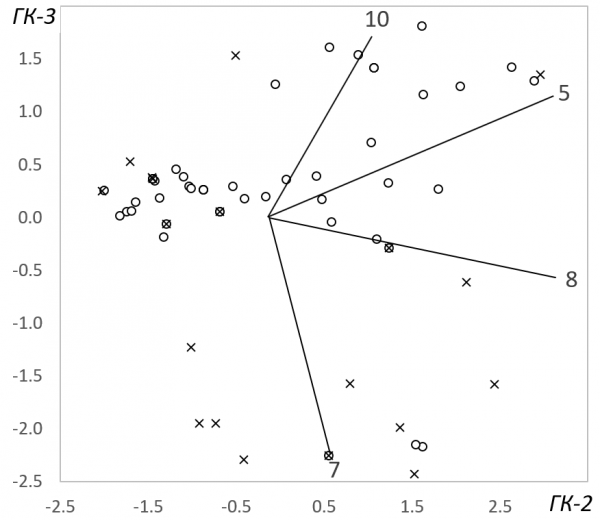
Рис. 2. Биплот ординации рыб по спектру питания в осях значимых главных компонент. Оси факторных нагрузок обозначены в сответствии с нумерацией объектов питания, принятой в табл. 3
Fig. 2. Biplot of fish ordination according to the feeding spectrum in the axes of significant principal components. The аxes of the factor loadings are indicated in accordance with the numbers of feed objects as in Table. 3
Обсуждение
В водоемах Карелии сиги растут быстрее, чем окуни, например, в возрасте 4+ среднестатистический малотычинковый сиг из оз. Каменное при длине в 23.4 см весит 143 г, а такой же по возрасту окунь имеет размер 14.5 см и массу тела 52 г (Первозванский, 1986). Этим обстоятельством объясняется размерная и весовая дифференциация рыб в наших пробах. Соответствие размерно-весовых характеристик сигов нормальному распределению указывает на относительную однородность выборки и позволяет заключить, что отловленные рыбы принадлежат к одной совокупности, в данном случае – единственной, подошедшей к месту вылова стае. Значимое отклонение частот от гауссовой кривой у окуней можно интерпретировать как суперпозицию распределений, возникающую в результате объединения нескольких, существенно отличающихся по возрасту кластеров – по-видимому, в сети попадали рыбы из разных стай. Другим показателем отличий может служить коэффициент вариации длины тела, значение которого у сигов не достигает и 10 %, тогда как у окуней находится на уровне 26 %. Размерный показатель в данном случае является ключевым индикатором изменчивости, поскольку масса тела зависит от наполнения желудка и, следовательно, менее постоянна.
Состав пищи сигов и окуней в исследованном озере типичен – они, как и везде в Карелии, питаются всеми представителями зообентоса, подходящими по размеру, в рационе мелких рыб преобладает зоопланктон, а в желудках крупных особей встречаются рыбы (Первозванский, 1986). Устойчивой видовой специфики пищевых связей на региональном уровне не прослеживается, предпочтения сигов и окуней варьируют в широких пределах в зависимости от водоема и времени года. Рыбы, по-видимому, в каждом конкретном случае используют в пищу наиболее доступные объекты, и выявленные нами в оз. Падашулкаярви предпочтения в питании сига и окуня (моллюски и ручейники соответственно) можно считать спецификой данного водоема в весенний период. Еще одной важной особенностью является также то, что в отличие от других водоемов летающие имаго насекомых как объект питания более характерны для окуней, чем для сигов.
Результаты компонентного анализа подтверждают хорошо известный возрастной тренд изменчивости трофических связей: молодые рыбы питаются в основном зоопланктоном, а затем, по мере роста, переходят на более крупные объекты, в содержимом желудков существенно возрастает доля бентосных форм. Эта закономерность выявляется в двух первых компонентах, отражая дифференциацию бентоса на две группы – хирономид и ручейников, личинок стрекоз и поденок. Подобное разделение обычно связывают с преобладанием тех или иных кормовых объектов в различных биотопах. Важная роль при этом отводится сезонным аспектам в жизни насекомых, когда, например, определенный вид корма становится недоступным вследствие массового вылета имаго. Показанная дифференциация трофических ниш характеризует, прежде всего, окуней, рацион сигов менее разнообразен. Принадлежность последних к одной стае предполагает, что все рыбы, в отличие от окуней, кормились в одинаковых условиях, а отсутствие значимой связи структуры рациона с длиной их тела обусловлено небольшой изменчивостью вариант в исследуемой выборке. Основное отличие сигов от окуней – использование ими в качестве основного кормового объекта моллюсков – также может быть обусловлено особенностями питания конкретной стаи непосредственно перед отловом.
Заключение
Таким образом, использование одной и той же кормовой базы сигами и окунями в условиях небольшого озера потенциально может приводить к межвидовой конкуренции. Однако эксплуатация различных ресурсов, распределенная в пространстве и во времени, способствует динамическому разделению трофических ниш не только между видами, но и между стаями. Кормовая база оз. Падашулкаярви, по-видимому, позволяет это делать, – на слабое перекрывание ниш сигов и окуней указывает индекс Мориситы – Хорна. Отсутствие острой конкуренции обеспечивает возможность сосуществования в одном небольшом водоеме популяций этих видов рыб.
Библиография
Биология речного окуня . М.: Наука, 1993. 188 с.
Дятлов М. А. Рыбы Ладожского озера . Петрозаводск: КарНЦ РАН, 2002. 281 с.
Ивантер Э. В., Коросов А. В. Введение в количественную биологию . Петрозаводск: Изд-во ПетрГУ, 2014. 298 с. URL: http://elibrary.karelia.ru/book.shtml?levelID=003&id=21497&cType=1 (дата обращения: 01.07.2017).
Коросов А. В., Горбач В. В. Компьютерная обработка биологических данных . Петрозаводск: Изд-во ПетрГУ, 2010. 86 с. URL: http://elibrary.karelia.ru/book.shtml?levelID=031&id=15287&cType=1 (дата обращения: 01.07.2017).
Методическое пособие по изучению питания и пищевых отношений рыб в естественных условиях . М.: Наука, 1974. 254 с.
Первозванский В. Я. Рыбы водоемов района Костомукшского железорудного месторождения (экология, воспроизводство, использование) . Петрозаводск: Карелия, 1986. 216 с.
Петрова Л. П., Кудерский Л. А. Водлозеро: природа, рыбы, рыбный промысел . Петрозаводск: КарНЦ РАН, 2006. 198 с.
Плохинский Н. А. Биометрия . М.: Изд-во МГУ, 1970. 367 с.
Попова О. А., Решетников Ю. С. О комплексных индексах при изучении питания // Вопросы ихтиологии. 2011. Т. 51. № 5. С. 712–717.
Стерлигова О. П., Ильмаст Н. В., Савосин Д. С. Круглоротые и рыбы пресных вод Карелии . Петрозаводск: Карельский научный центр РАН, 2016. 224 с.
Hammer Ø., Harper D. A. T., Ryan P. D. PAST: Paleontological statistics software package for education and data analysis // Palaeontologia Electronica. 2001. Vol. 4. № 1. P. 1–9. URL: https://folk.uio.no/ohammer/past/ (дата обращения: 15.06.2017).
Horn H. S. Measurement of "Overlap" in comparative ecological studies // The American Naturalist. 1966. Vol. 100. № 914. P. 419–424.
Hothorn T., Hornik K., van de Wiel M. A., Zeileis A. Package ‘coin’. Conditional inference procedures in a permutation test framework. Ver. 1.0-11. 2010. URL: http://cran.opensourceresources.org/index.html (дата обращения: 20.06.2017).

 © 2011 - 2026
© 2011 - 2026