



Введение
Совокупность процессов жизнедеятельности организма, направленных на размножение, служит залогом существования надорганизменных комплексов во времени. В то же время пластичность репродуктивной стратегии можно рассматривать в качестве адаптивной возможности, которая может проявляться на популяционном уровне (Шмальгаузен, 1983). В частности, варианты репродуктивной специфики пойкилотермных животных как r-стратегов, как правило, канализированы в двух направлениях:
1) увеличение числа яиц при уменьшении их средних размеров;
2) увеличение размеров яйца на фоне снижения плодовитости.
При этом оба сценария часто можно наблюдать у представителей одного вида. Подобное проявление вариантов в репродукции некоторые авторы именуют принципом «адаптивной игры в орлянку» («adaptive coin flipping») (Kaplan, Cooper, 1984). Исходя из общих представлений о репродуктивном потенциале популяций (Darwin, 1874; Shine, 1988; Reeve, Fairbairn, 1999) имеет место существенная связь размеров производителей с их плодовитостью. Выбор в пользу конкретного варианта может быть обусловлен экологической спецификой популяции на фоне локальных условий среды. Биологический смысл «адаптивной игры в орлянку» заключается в оптимизации затрат энергии на процессы, связанные с размножением (MacArthur, Wilson, 1967; Pianka, 1970). При любом сценарии вероятность успеха воспроизводства на популяционном уровне должна оставаться высокой (Шмальгаузен, 1983).
В связи с этим цель данной работы – анализ репродуктивных особенностей прыткой ящерицы (на примере северных популяций в западных и восточных предгорьях Среднего Урала) в контексте оптимизации энергозатрат на размножение.
Материалы
Была выполнена работа по изучению плодовитости самок прыткой ящерицы. Районы проведения исследований находятся на территории Среднего Урала: в западных и восточных предгорьях Уральского хребта на юге Свердловской области. Полевой материал собран в 2011–2013 гг. в трех локалитетах (рисунок). В данном контексте понятие «локалитет» означает географическую точку на карте Свердловской области, где находятся сообщества с многолетним обитанием прыткой ящерицы.
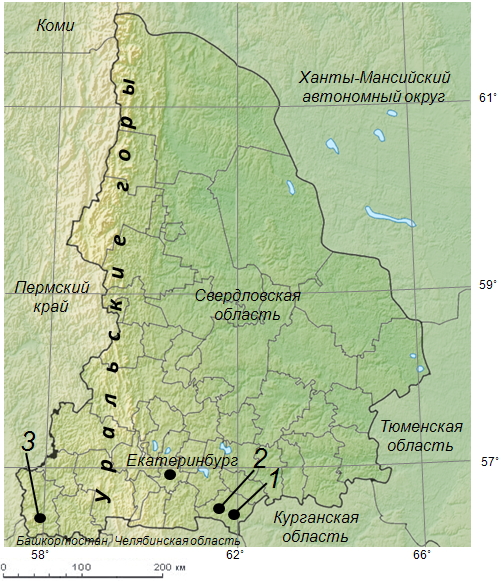
Картосхема Свердловской области с указанием местообитаний прыткой ящерицы Lacerta agilis: 1 – зона многоэтажной застройки г. Каменска-Уральского, 2 – окрестности железнодорожной станции Кунавино, 3 – Красноуфимская лесостепь
Schematic map of Sverdlovsk region and sand lizard Lacerta agilis habitats: 1 – area with blocks of flats in Kamensk-Uralsky town, 2 – Kunavino railway station neighborhoods, 3 – forest-steppe in Krasnoufimsk neighborhood
1. Зона многоэтажной застройки в черте г. Каменска-Уральского. Координаты: 56º24´ с. ш., 61º55´ в. д. Данное местообитание расположено в подтаежной зоне в восточных предгорьях Уральского хребта на границе двух административных районов г. Каменска-Уральского: Красногорского и Синарского, на расстоянии нескольких десятков метров от многоэтажных жилых зданий, малых хозяйственных корпусов и Байновского моста через р. Исеть с двусторонним пешеходным и транспортным движением. Участки обитания прыткой ящерицы сосредоточены в пойме р. Исеть, включающей транспортные коммуникации и опушку соснового леса. Типичный ландшафт в зоне поселения ящериц представляет собой хорошо прогреваемые остепненные склоны западной и восточной экспозиции с уклоном 35–56 градусов. Антропогенный фактор выражен прежде всего в загрязнении транспортными выхлопными газами и твердыми бытовыми отходами, фрагментации ландшафта за счет наличия действующих коммуникаций, а также большой рекреационной нагрузке. Площадь локалитета 9.2 га.
2. Зона малоэтажной застройки в окрестностях железнодорожной станции Кунавино Каменского района. Координаты: 56º27´ с. ш., 61º49´ в. д. Локалитет находится в подтаежной зоне на восточном макросклоне Уральских гор на территории Каменского района в 17 км к северо-западу от центра г. Каменска-Уральского (в качестве центра города принято местоположение центрального почтамта) и в 9 км от городской популяции. Данный ландшафт представляет собой характерный пример сельского поселения человека с немногочисленными одноэтажными зданиями и садово-огородными хозяйствами. Находки L. agilis были отмечены на открытых пространствах, примыкающих к лесозащитным полосам и железнодорожной линии. Антропогенный фактор выражен в присутствии действующих железнодорожных путей и наличия малых агроландшафтов. Площадь локалитета 12.5 га.
3. Участок Красноуфимской лесостепи в окрестностях пос. Новый Бугалыш. Координаты: 56º18´ с. ш., 57º56´ в. д. Локалитет расположен на западном макросклоне Уральского хребта в 37 км к югу от г. Красноуфимска и в 175 км к юго-западу от г. Екатеринбурга. Расстояние до городской популяции L. agilis в восточных предгорьях – 245 км. Общий облик данного локалитета формируют лесостепные сообщества, типичные для обитания прыткой ящерицы. Антропогенный фактор выражен в сельскохозяйственном использовании ландшафта, наличии пастбищ и полезащитных насаждений. Площадь локалитета 23.1 га.
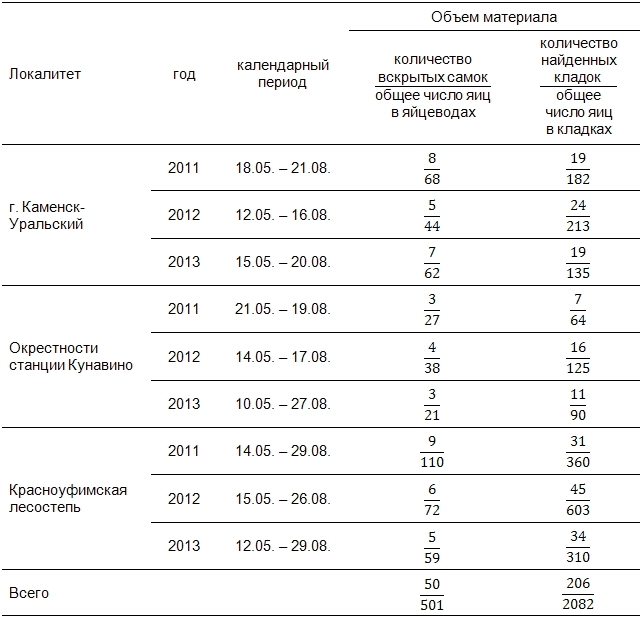
Измеряли длину тела (L.) беременных самок, отмечали количество и размеры зрелых яиц в их яйцеводах, число яиц в найденных кладках рептилий (размер кладки), особенности местоположения кладок в стациях. Также определяли процент погибших кладок в качестве косвенного индикатора прессинга на ранних стадиях онтогенеза прыткой ящерицы. Общий объем материала, включенного в исследование, представлен в табл. 1.
Таблица 1. Объем исследованного материала
Методы
Измерения длины тела самок и размеров яиц в их яйцеводах выполнены с помощью цифрового штангенциркуля ШЦЦ-1-125 (Россия) с точностью до 0.1 мм. Были сняты промеры длины (большого диаметра, D) и ширины (малого диаметра, d) яиц из яйцеводов для расчета объема яйца, согласно формуле для вытянутого эллипсоида вращения (Бронштейн, Семендяев, 1964):
V = 4/3πa2b,
где a и b – малая и большая полуоси соответственно.
Анализ и статистическую обработку данных проводили по общепринятым методикам (Лакин, 1990) при помощи программных пакетов MS Excel 2007 и Statistica 8.0. Для проверки нормальности распределения применяли тест Шапиро – Уилка. Сравнительный анализ данных для выявления возможных различий проводили с помощью дисперсионного анализа. Для определения корреляционных отношений размера самки, объема яйца и плодовитости использовали линейный коэффициент (R).
Полученные результаты описывали с точки зрения репродуктивных стратегий L. agilis в местных условиях. Макросклоны Уральской горной страны обладают своей климатической спецификой. Так, восточный склон расположен в барьерной тени хребта, поэтому, несмотря на преобладание западных ветров, приносящих осадки, климат здесь более сухой и континентальный, чем на западном склоне (Алисов и др., 1954; Дьяченко, 1997). Гипотеза исследования базируется на том, что климатическая специфика подобного рода может влиять на стратегию размножения прыткой ящерицы.
Результаты
В результате анализа репродуктивных характеристик L. agilis установлена общая тенденция и определенная специфика популяций на западном и восточном макросклоне Уральских гор (табл. 2).
Таблица 2. Репродуктивные показатели самок прыткой ящерицы Lacerta agilis в уральских популяциях
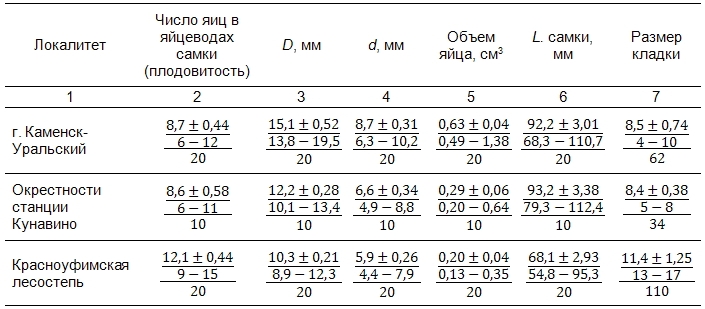
Примечание. Верхний ряд – средние значения ± ошибки среднего; средний ряд – минимальные и максимальные значения; нижний ряд – объем выборки: 2–6-й столбцы – число вскрытых самок, 7-й столбец – число найденных кладок.
В качестве общей закономерности установлена отрицательная корреляция (R = -0.432) объема яйца и плодовитости (F = 11.04; p = 0.0017). В то же время по репродуктивным показателям самок в изучаемых популяциях выявлены существенные различия. Например, средний объем яйца значимо уменьшается (F(2.47) = 28.747; p = 0.00001) в направлении от восточного макросклона к западному. При этом для плодовитости наблюдается противоположная тенденция. Кроме того, средние размеры самок в данных популяциях также отличаются (F(2.158) = 98.434; p = 0.00001): в популяциях восточного склона длина тела самок больше, чем западного.
Результаты анализа репродуктивных характеристик указывают на снижение плодовитости прыткой ящерицы в направлении с запада на восток. Подобная тенденция может быть сопряжена с сонаправленным возрастанием фрагментации территорий. Наибольший объем яйца отмечен в городской популяции (0.63 см3), где число яиц в яйцеводах самки невелико – 8.7. Для рептилий восточных предгорий отмечена положительная корреляция между размером самки и объемом яйца (R = 0.484, p = 0.0012), в то время как в популяции западных предгорий такой связи не выявлено.
Результаты хронологических наблюдений показали, что самки откладывают яйца в период с середины по конец июня на западном макросклоне и с конца июня по середину июля на восточном. При этом средняя продолжительность периода откладки яиц для рептилий западных и восточных предгорий составляет 11 дней. Большинство кладок были найдены в неглубоких норах и в основании ветвей мелких кустарников (чаще всего это дрок красильный Genista tinctoria Linnaeus, 1758; ракитник русский Chamaecytisus ruthenicus (Fish. ex Wol.) Klask., 1958). Реже кладки встречались непосредственно в подстилочно-войлочном горизонте почвы либо в небольших углублениях под крупными одиночными деревьями или скальными выступами.
Далеко не все кладки были обнаружены в жизнеспособном состоянии: часть из них погибла в результате механических повреждений и/или разорения хищниками. Встречаемость поврежденных кладок составляет 37.5 % на восточном склоне и 27.6 % на западном. Следовательно, определенная доля рептилий не выживает еще до выхода во внешнюю среду. Доля поврежденных кладок может служить косвенной оценкой прессинга в популяциях. В увеличении этой доли может отражаться действие стрессовых факторов, в особенности для городской популяции в восточных предгорьях. Относительная плотность кладок в городской популяции (6.92 ± 0.74) существенно выше, чем в других локалитетах (0.88 ± 0.19 и 4.84 ± 0.34 в окрестностях станции Кунавино и Красноуфимской лесостепи соответственно), что может быть обусловлено меньшей площадью местообитания и высокой плотностью городской популяции (Галицын, 2013).
Обсуждение
Среди множества характеристик популяции репродуктивные возможности являются наиболее важными, т. к. именно они определяют динамику численности (Солбриг, Солбриг, 1982). Прогнозируемая динамика населения, в т. ч. оценка риска вымирания, в значительной мере обусловлена приростом популяции, что, в свою очередь, сильно зависит от плодовитости самок. В данной статье в качестве показателя плодовитости L. agilis использовали число яиц в яйцеводах, что означало безвозвратное изъятие экземпляров из репродуктивного ядра популяций (см. Материалы). Несмотря на инвазивность подхода (как следствие – определенный ущерб для популяций), его использование в данном случае представляется вполне оправданным. Прежде всего потому, что использование числа яиц в найденных кладках в качестве показателя плодовитости может быть сопряжено с существенным риском смещения оценки как в сторону завышения (Власова и др., 2013), так и снижения (Кидов и др., 2012). Это происходит из-за способности самок прыткой ящерицы делать так называемые порционные и групповые кладки. В то же время размеры обнаруженных кладок и количество яиц внутри тела самок могут коррелировать между собой, что наблюдается в изучаемых группах. В анализе размеров и объема яиц также применяли соответствующие показатели, полученные в результате вскрытия беременных самок. Привлечение к анализу размеров яиц во внешней среде с целью снижения ущерба для популяций было нецелесообразным, поскольку размеры яиц L. agilis в течение инкубационного периода существенно возрастают (Кидов, Матушкина, 2017), а точная дата откладки яиц в природных популяциях рептилий остается неопределенной.
В городской популяции L. agilis отмечена наименьшая плодовитость и наибольший объем яйца. Однако в популяции, обитающей в окрестностях станции Кунавино, при сопоставимой плодовитости средний объем яйца значительно меньше. Возможно, в данном случае для более четкого отражения связи размера яйца и плодовитости необходима выборка большего объема. Однако численность и плотность данной популяции существенно ниже в сравнении с городской и лесостепной; по этой причине было вскрыто вдвое меньше самок. В восточных предгорьях подходящие участки для обитания L. agilis встречаются фрагментарно, следовательно, популяции в значительной степени изолированы и могут обладать своей репродуктивной спецификой. Показано (Siliceo, Díaz, 2010), что кладки небольшого размера характерны для угрожаемых видов лацертидных ящериц с фрагментированными участками ареалов.
Сопоставление сведений по размерам кладок в изучаемых популяциях L. agilis со сводными монографическими данными (Баранов и др., 1976) выявляет сходство городской популяции с популяциями лесостепной зоны, в то время как группировка в окрестностях станции Кунавино ближе к некоторым популяциям лесной зоны. Число яиц в кладках рептилий Красноуфимской лесостепи оказывается существенно выше и сравнимо с представителями популяций степной зоны юга России, для которых описана наибольшая плодовитость (Кидов и др., 2011).
В рамках основной гипотезы исследования состояние репродуктивных параметров прыткой ящерицы отражается в контексте микроклиматических условий западного и восточного макросклона Уральского хребта. Известно, что в формировании и динамике климата Урала велика роль западного переноса воздуха. Уральские горы не слишком высоки, но тем не менее заметно ослабляют движение воздушных масс с запада на восток, вследствие чего количество осадков на западных склонах увеличивается, а в направлении на восток уменьшается (Дьяченко, 1997). Об этом свидетельствуют более высокие значения гидротермического коэффициента на западном склоне – 1.5-1.6 в сравнении с восточным – 1.26. Вероятно, более резко континентальные климатические условия в восточных предгорьях (Алисов и др., 1954; Дьяченко, 1997) могут способствовать преимущественной выживаемости крупных производителей и формированию крупных яиц в городской популяции. Более крупные яйца обладают большей удельной теплоемкостью, а соотношение их объема и площади поверхности обеспечивает меньшую потерю влаги. К тому же показано, что размер яйца ящериц обладает определенной степенью термопластичности (Lai-Gao et al., 2010). В то же время формирование большого количества крупных яиц будет неизбежно сопряжено со значительным расходом энергии, которую организм должен восполнить. В связи с этим увеличение объема яиц сопровождается компенсаторным снижением плодовитости. Поскольку размеры сеголетков, как правило, хорошо соответствуют размерам яиц (Roitberg et al., 2015), необходимый уровень выживаемости новой генерации рептилий в городской популяции должен обеспечиваться также за счет крупных сеголетков.
Говоря о вкладе плодовитости рептилий в фактический прирост природных популяций, важно учитывать показатель сохранности яиц в течение инкубационного периода, а также процент выхода молоди. Так, на примере популяции L. agilis Центрального Предкавказья показано (Кидов, Матушкина, 2017), что при инкубации в лабораторных условиях средняя сохранность яиц составляет 39.4 %. В природных условиях на фоне совокупного действия факторов следует ожидать более низкие показатели. На увеличение или снижение процента гибели яиц может влиять климатическая характеристика репродуктивного сезона (Епланова, 2014). Поэтому в качестве направления дальнейшего поиска следует выделить сравнительное изучение размножения представителей уральских популяций L. agilis в природных и модельных условиях. Это дает возможность оценить процент выхода и выживаемость сеголетков, а также апробировать практические методы контроля уязвимых популяций с искусственной инкубацией яиц (Corbett, 1988) и/или зимовкой сеголетков в неволе (Caughley, Gunn, 1996) с последующим их возвращением на участки обитания.
Заключение
Таким образом, при сопоставлении полученных тенденций с принципом «адаптивной игры в орлянку» (см. Введение) можно отметить реализацию обоих направлений в репродуктивной стратегии прыткой ящерицы на популяционном уровне. Первый вариант с большей плодовитостью реализуется в лесостепной популяции на западном склоне, в то время как второй вариант с большим размером яйца – в городской популяции восточных предгорий. В условиях нарастания континентальности климата репродуктивная пластичность подобного рода может быть проявлением одного из регуляторных механизмов адаптивности локальных популяций прыткой ящерицы.
Библиография
Алисов Б. П., Берлин И. А., Михель В. М. Курс климатологии. Ч. 3 . Л.: Гидрометеоиздат, 1954. 320 с.
Баранов А. С., Стрельцов А. Б., Тертышников М. Ф. Размножение // Прыткая ящерица. Монографическое описание вида. М.: Наука, 1976. С. 214–226.
Бронштейн И. Н., Семендяев К. А. Справочник по математике для инженеров и учащихся втузов . М.: Наука, 1964. 604 с.
Власова О. П., Власов Е. А., Власов А. А. Пресмыкающиеся Центрально-Черноземного заповедника // Вестник Тамбовского университета. Сер.: Естественные и технические науки. 2013. Т. 18. № 6–1. С. 2988–2991.
Галицын Д. И. Территориальность и характер поселения прыткой ящерицы (Lacerta agilis Linnaeus, 1758) на трансформированных территориях Среднего Урала // Известия Уфимского научного центра РАН. 2013. № 3. С. 7–10.
Дьяченко А. П. Флора листостебельных мхов Урала. Ч. 1: История изучения. Конспект. Таксономический анализ . Екатеринбург: Изд-во Урал. ун-та, 1997. 264 с.
Епланова Г. В. О различиях репродуктивных характеристик двух популяций прыткой ящерицы из Самарской области // Известия Самарского научного центра РАН. 2014. Т. 16. № 5 (1). С. 421–423.
Кидов А. А., Тимошина А. Л., Матушкина К. А., Пыхов С. Г., Ливадина Л. В., Жиримес В. Г. Материалы к изучению репродуктивной биологии настоящих ящериц (Reptilia, Sauria, Squamata: Lacertidae) Кавказа // Научные исследования в зоологических парках. М.: Московский Зоологический Парк, 2011. Вып. 27. С. 100–113.
Кидов А. А., Тимошина А. Л., Коврина Е. Г., Матушкина К. А., Пыхов С. Г. Характеристика репродуктивных показателей восточной прыткой ящерицы (Lacerta agilis exigua Eichwald, 1831) (Reptilia, Squamata, Sauria: Lacertidae) в Кумо-Манычской впадине // Естественные и технические науки. 2012. № 1 (57). С. 81–83.
Кидов А. А., Матушкина К. А. Размножение прыткой ящерицы (Lacerta agilis L.) на Ставропольской возвышенности // Известия Тимирязевской сельскохозяйственной академии. 2017. Вып. 4. С. 56–69.
Лакин Г. Ф. Биометрия . М.: Высшая школа, 1990. 352 с.
Солбриг О., Солбриг Д. Популяционная биология и эволюция . М.: Мир, 1982. 488 с.
Шмальгаузен И. И. Пути и закономерности эволюционного процесса. Избранные труды . М.: Наука, 1983. 360 с.
Caughley G., Gunn A. Conservation Biology in Theory and Practice. Cambridge: Blackwell Science, 1996. 459 p.
Corbett K. F. Verbreitung und Status der Zauneidechse Lacerta agilis in GrobBritannien // Mertensiella. 1988. № 1. P. 92–100.
Darwin C. The Descent of Man and Selection in Relation to Sex. 2nd edn. Appleton, New York, 1874.
Kaplan R. H., Cooper W. S. The evolution of developmental plasticity in reproductive characteristics: An application of the adaptive coin-flipping principle // Amer. Natur. 1984. № 123. P. 393–410.
Lai-Gao Luo, Guo-Hua Ding, Xiang Ji. Income breeding and temperature-induced plasticity in reproductive traits in lizards // The Journal of Experimental Biology. 2010. Vol. 213. P. 2073–2078.
MacArthur R. H., Wilson E. O. The theory of island biogeography. New York: Princeton Univ. Pres, 1967. 203 p.
Pianka E. R. On ”r” and ”k” selection // Amer. Natur. 1970. № 104. P. 592–597.
Reeve J. P., Fairbairn D. J. Change in sexual size dimorphism as a correlated response to selection on fecundity // Heredity. 1999. Vol. 83. P. 697–706.
Roitberg E. S., Eplanova G. V., Kotenko T. I., Amat F., Carretero M. A., Kuranova V. N., Bulakhova N. A., Zinenko O. I., Yakovlev V. A. Geographic variation of life-history traits in the sand lizard, Lacerta agilis: testing Darwin’s fecundity-advantage hypothesis // J. Evol. Biol. 2015. Vol. 28. P. 613–629.
Shine R. The evolution of large body size in females: a critique of Darwin’s ”fecundity advantage” model // Amer. Natur. 1988. Vol. 131. P. 124–131.
Siliceo I., Díaz J. A. A comparative study of clutch size, range size, and the conservation status of island vs. mainland lacertid lizards // Biological Conservation. 2010. Vol. 143. P. 2601–2608.
Благодарности
Работа выполнена в рамках государственного задания Института экологии растений и животных Уральского отделения РАН, а также частично поддержана Комплексной программой Уральского отделения РАН (проект № 18-4-4-28) и программой 211 Правительства Российской Федерации, соглашение № 02.A03.21.0006.
Авторы выражают благодарность заведующему музеем Института экологии растений и животных УрО РАН Н. Г. Ерохину за возможность работы с коллекционным материалом.

 © 2011 - 2026
© 2011 - 2026