



Введение
Каким образом можно перейти от современного экологического кризиса к гармоничному устойчивому существованию человека в биосфере?
Ответ на этот вопрос позволил бы конкретизировать пути возможного перехода биосферы в ноосферу – проблему, которую поставил еще В.И. Вернадский (Вернадский, 1944). В общей форме ответ ясен – человек должен перейти от стихийного влияния на биосферу к управлению антропогенными факторами, опасно нарушающими структуру и функцию биосферы (Lapenis, 2002; Rockstrom, Klum, 2015 и мн. др.). Это означает переход к «управляемой эволюции» биосферы (Яблоков и др., 2015).
Аналитический обзор
Предпосылкой эффективного управления является понимание движущих сил и структур системы, которой предстоит управлять (т.е. биосферы).
Из четырех уровней организации живого (молекулярно-генетического, онтогенетического, популяционно-видового и биосферного) наибольшая методологическая ясность в выделении таких сил и структур существует на популяционно-видовом уровне (Тимофеев-Ресовский и др., 1969).
После работ Ч. Дарвина в классической концепции эволюции было ясно, что главным фактором возникновения видов является естественный отбор. Шагнуть дальше в познании механизма эволюции позволило выделение в разномасштабном процессе эволюции (от популяций до царств) процессов микроэволюции (Dobzansky, 1937; Timofeeff-Ressovsky, 1939). Следующим шагом этого теоретического прорыва оказалось выделение элементарных структур и их взаимодействий.
Этот подход элементаризации позволил выделить на популяционно-видовом уровне организации жизни элементарную структуру (популяцию), элементарное явление (необратимое изменение генетического состава популяции), элементарный материал (мутации разных типов), элементарные факторы (мутационный процесс, волны численности, изоляция и естественный отбор), качественный этап (возникновение новых признаков и свойств - адаптаций, - приводящих к образованию нового вида). Эта элементаризация позволила успешно моделировать микроэволюционные ситуации (рис. 1).
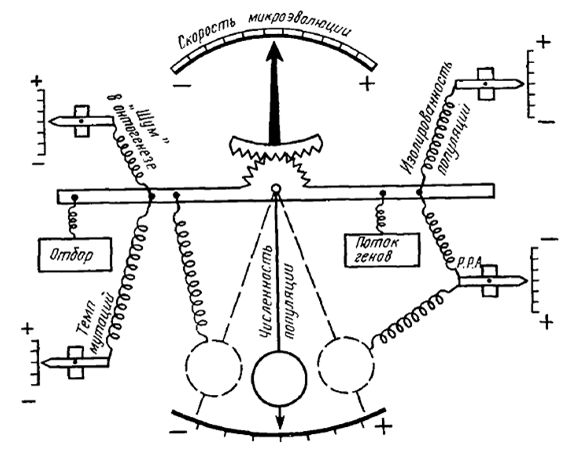
Рис 1. Схема взаимодействия эволюционных сил в процессе микроэволюции на популяционно-видовом уровне. Скорость возникновения адаптации (необратимого изменения генотипического состава популяции) является кумулятивным результатом. Р.Р.А. – радиус репродуктивной активности (дистанция переноса генов), «шум» в онтогенезе – обобщенное выражение эпигенетических процессов (из: Yablokov, 1986, по Sperlich, 1972 с изменениями).
Fig.1. The scheme of the interaction of evolutionary forсes in the process of microevolution at the population-species level.
Уровень современных знаний не позволяет пока сделать аналогичную модель для биосферного уровня. Г. М. Длусский, В. В. Жерихин, В. Ф. Левченко, R. М. May, R. Odum, Т. Н. Работнов, А. С. Раутиан, С. М. Разумовский, Я. И. Старобогатов, Ю. И. Чернов, С. С. Шварц, И. И. Шмальгаузен, Э. И. Колчинский и мн. др. в разное время предпринимали попытки выделения различных составляющих в процессе эволюции на биогеоценотическом уровне (обзоры см. Колчинский, 1990, Северцов, 2013; Пучковский, 2014, Yablokov et al., 2015).
Эволюционные события на популяционно-видовом и биосферном уровнях тесно взаимосвязаны. Возникновение любой адаптации на популяционно-видовом уровне обязательно отразится, в конце концов, в выполняемых видом биогеохимических функциях и изменении метаболизма экосистем. Возникновение нового вида означает возникновение новой экологической ниши, то есть использование ресурсов, которые не использовались ранее, это - шаг в освоении живыми организмами биосферы (Gauze, 1934, Тимофеев-Ресовский, 1958). Эволюция видов, с одной стороны, определяется трофико-энергетическими связями, а с другой стороны, определяет эволюцию всей биосферы. Эта связь – один из аргументов за то, чтобы попытаться перенести подход элементаризации, успешно «сработавший» на популяционно-видовом уровне, к процессам эволюции на биосферном уровне.
Первый шаг на этом пути – выделение среди всего многообразия процессов эволюции биосферы процессов макроэволюции (эволюции всей биосферы в целом), и микроэволюции – процессов на уровне биогеоценозов.
Минимальной, далее неразложимой без потери качества, единицей на биосферном уровне оказывается биогеоценоз (Сукачев, Дылис, 1964, Тимофеев-Ресовский, 1970, Работнов, 1995, Тюрюканов, 2001 и мн. др.).
Биогеоценоз (БГЦ) — исторически сложившаяся, относительно стабильная в пространстве и времени (на протяжении, по меньшей мере, одного сукцессионного цикла – см. далее) экосистема на основе конкретного растительного сообщества. БГЦ занимает сравнительно крупный участок поверхности суши Земли и включает почву, растительный покров и приземный слой атмосферы (экотоп), с обитающими в этом пространстве всеми живыми существами (биоценоз). Компонентами биоценоза являются фитоценоз и зооценоз. БГЦ включает постоянно или периодически обитающих представителей множества видовых популяций практически всех царств живых организмов. Пространственные границы БГЦ могут быть размыты и подвижны во времени. Принято, что они определяются границами популяций растений-эдификаторов (т.е. тех, которые создают облик данного сообщества и существенно изменяют среду). Ареал популяций некоторых видов может включать разные БГЦ. Внутри БГЦ не проходит популяционных границ для принадлежащих ему видовых популяций. Непременной особенностью любого БГЦ является сукцессия – циклическая смена биоценозов. На каждой фазе сукцессионного цикла происходит накопление специфических для нее веществ и энергии и, тем самым, создаются условия для формирования последующей фазы и сообщества с другой композицией видовых популяций. На последнем этапе сукцессии возникает относительно устойчивый и сравнительно долго существующий биоценоз (климаксное сообщество), после деградации которого весь сукцессионный цикл повторяется.
Именно биогеоценоз оказывается элементарной эволюционной структурой биосферы (Тимофеев-Ресовский, 1970, и др.). Предположение (Колчинский, 1990), что видовая популяция выступает не только элементарной единицей эволюции на популяционно-видовом уровне, но и как элементарная биогеохимическая, энергетическая и трофическая единица на уровне биогеоценоза, не оправдано уже потому, что ареалы видовых популяций могут охватывать пространство многих биогеоценозов.
Есть некоторая ясность и в выделении элементарного эволюционного события в биосфере. В. В. Жерихин (1997) считал таковым необратимое изменение набора реализованных экологических ниш, то есть фактически - изменение видового состава биогеоценоза. В качестве рабочей гипотезы можно принять, что элементарным эволюционным событием на этом уровне является необратимое изменение видового состава биогеоценоза, означающее изменение (метаморфоз) метаболизма биогеоценоза или группы биогеоценозов.
Качественный этап микроэволюции на биосферном уровне – образование нового биогеоценоза (метаморфоз метаболизма прежнего БГЦ). Возникновение нового биогеоценоза должно быть связано с возникновением новых эволюционных дифференцировок. Образование нового БГЦ (и, по-видимому, биома на макро-эволюционном биосферном уровне) определяет, скорее всего, процесс группового (ценотического) естественного отбора. Только в процессе такого отбора может пройти апробация бесчисленных сочетаний разных видовых популяции, сочетания потоков вещества и энергии в троичекой системе «продуцент – консумент – редуцент», и материализованные в мутуалистических (кооперативных) отношениях хищник-жертва, комменсализма, паразитизма, симбиотизма.
Естественный отбор является и на этом уровне (в форме группового отбора) направляющим эволюцию элементарным эволюционным фактором. Является ли он единственным элементарным фактором микроэволюции на биосферном уровне? По аналогии с процессами на популяционно-видовом уровне, эволюционными фактором может быть изоляция (возникновение каких-то преград в потоках вещества и энергии).
Пока нет ясности в выделении элементарного эволюционного материала на биосферном уровне. Возможно, на этом уровне элементарным эволюционным материалом являются видовые популяции.
В процессе микроэволюции на биосферном уровне точками приложения группового отбора могут быть либо особенности (оптимизация) потоков вещества и энергии (и информации?) в процессах метаболизма экосистем, либо, отражающий эти же процессы, видовой состав БГЦ.
Заключение
Предложенная выше схема элементаризации на биосферном уровне, включающая выделение микроэволюционного уровня эволюции биосферы и выделения на этом уровне элементарных эволюционных единиц, материала, факторов, событий и качественных этапов, конечно, требует критического анализа и развития с привлечением фактического материала по конкретным биоценозам. Эта работа трудоемкая и потребует участие многих специалистов, но она должна быть сделана, если мы хотим перейти к управляемой эволюции биосферы.
Библиография
Вернадский В. И. Несколько слов о ноосфере. // Усп. Совр. Биол. 1944. Том 18. Вып. 2. С. 113–120.
Жерихин В. В. Избранные труды по палеоэкологии и филоценогенетике. М.: КМК, 2003. . 542 с.
Колчинский Э. И. Эволюция биосферы. Историко-критический очерк исследований в СССР. Л.: «Наука», 1990. 236 с.
Пучковский С. В. Эволюция биосистем. Факторы микроэволюции и филогенеза в эволюционном пространстве-времени. Ижевск: изд. Удмуртского университета, 2013. 444 с.
Pаботнов Т. А. История фитоценологии. М.: «Аргус», 1995. 158 с.
Северцов А. С. Эволюционная экология позвоночных животных. М.: КМК, 2013. 347 с.
Сукачев В. Н., Дылис Н. В. Основы лесной биогеоценологии. М.: Наука, 1964. 574 с.
Тимофеев-Ресовский Н. В. Микроэволюция. Элементарные явления, материал и факторы микроэволюционного процесса. // Бот. Журн. 1958. Том 43. № 3. С. 317–336.
Тимофеев-Ресовский Н. В. Структурные уровни биологических структур. В кн.: Системный подход в экологии. Системные исследования АН СССР. Институт истории естествознания и техники. М. 1970. С. 80–136.
Тимофеев-Ресовский Н. В., Воронцов Н. Н., Яблоков А. В. Краткий очерк теории эволюции. М.: «Наука», 1969. 408 с.
Тюрюканов А. Н. Избранные труды. М.: РЭФИА, 2001. 308 с.
Яблоков А. В. Популяционная биология. М.: Высшая Школа, 1986. 304 с.
Dobzhansky Th. Genetics and the Origin of Species. N.Y.: Columbia Univ. Press., 1937. 364 p.
Gauze G. F. The Struggle for Existence. Bultimore: Williams & Wilkins, 1934. 163 p.
Lapenis A. G. Directed Evolution of the Biosphere: Biogeochemical Selection or Gaia? // Profes. Geograph. 2002. Vol. 54. N 3. pp. 379–391.
Rockström J., Klum M. Big World, Small Planet: Abundance within Planetary Boundaries. New Haven: Yale University Press, 2015. 208 p.
Sperlich D. Populationgenetik. Grundlagen und Experimentelle Ergebnisse. Stuttgart, 1973. XI+ 197 p.
Timofeeff-Ressovsky N. W. Genetik und Evolution (Bericht einer Zoologen). Z. Indukt. Abstammungs-Vererbungsl. 1939. Vol. 76. P. 158–218.
Yablokov A.V. Population Biology. Progress and Problems of Studies on Natural Populations. M.: MIR, 1986. 304 p.
Yablokov A., Levchenko V., Kerzhentsev A. The Decision Exists: Transition to Controlled Evolution of the Biosphere // Philosophy & Cosmology. 2015. Vol. 14. P. 91–117.

 © 2011 - 2026
© 2011 - 2026