



Статья-рецензия написана по поводу того, что в редакцию «Принципов экологии» поступили две рукописи, выражающие два подхода к исследованиям по термобиологии рептилий, которые опубликованы в настоящем выпуске. Это своего рода «верхушки айсбергов» над обширными списками публикаций каждого из авторов и их коллег, отражающие образ их мыслей. Работы В. А. Черлина и др. ориентированы на изучение проблем терморегуляции в контексте прочих жизненных отправлений рептилий, которые обслуживаются различными формами поведения; таким образом, это «этолого-термобиологическое» направление. В исследованиях Н. А. Литвинова и др. делается попытка дать полномасштабную температурную характеристику полевой экологии рептилий по всех ее аспектах (органном, индивидуальном, сезонном, географическом); условно говоря – это «эколого-термобиологическое» направление. С обеих сторон можно услышать взаимную критику проблемных мест противоположного направления. Эта критика частично оправдана (кто не без греха?), а частично неконструктивна, поскольку оба направления, на наш взгляд, имеют существенный общий недостаток – отсутствие в своей основе теории физиологической регуляции. Цель нашего сообщения состоит в том, чтобы предложить общее теоретическое основание и новое общее направление совместных исследований в этой области.
В поисках точек соприкосновения обоих направлений исследований необходимо кратко остановиться на их основных достоинствах и недостатках. Поскольку собственные суждения не могут быть свободными от субъективизма, пришлось ориентироваться и на мнения рецензентов. Должно быть понятно также, что в нашу задачу не входит оценка вклада того или иного автора в термобиологическую науку.
Уже на этом этапе обсуждения следует определиться с тематикой собственно термобиологических исследований. Поскольку стоит задача объединения усилий исследователей разной направленности, то следует искать ясно обозначенные точки соприкосновения. Если считать, что к «термобиологии рептилий» должны относиться все явления жизни, связанные с температурой, то это понятие станет почти равным понятию «биология рептилий». По этой причине примем, что предметом нашего рассмотрения будет только проблема терморегуляции.
В «этолого-термобиологических» исследованиях одним из важнейших моментов оказывается теоретическое положение, согласно которому каждая форма поведения рептилий проявляется в своем диапазоне температуры тела, т.е. при каждой форме поведения у рептилий проявляется специфическая стратегия отношения к температуре. Таким образом, обобщенные количественные термальные характеристики имеют биологический смысл только относительно той формы поведения, при которой они собирались (по этой причине в работах В. А. Черлина так много внимания уделяется формам поведения). Сбор полевых данных осложняется тем, что диапазоны температуры тела, характерные для некоторых форм поведения, могут перекрываться. Отсюда следует важный методический момент термобиологических наблюдений: сначала определяем форму поведения животного, потом наблюдаемую температурную характеристику относим только к этой форме и используем в расчетах обобщенных показателей (средних, дисперсий и пр.). С этих позиций строится критика распространенного способа описания термальных свойств рептилий путем объединения в общую выборку всей массы полученных в поле значений для расчета статистических показателей (средней, дисперсии и т. п.), а также последующее сравнение полученных средних между регионами, сезонами, цветовыми формами и пр. В такой «многофакторной каше» одной выборки показатели отдельных форм поведения обязательно будут представлены в случайных пропорциях, которые никогда не совпадут с пропорциями в другой выборке из другого региона или за другой отрезок времени. Следовательно, различия между выборочными параметрами наблюдаться будут, но всегда по непонятной причине. Смысл имеет только сравнение термальных характеристик для отдельных форм поведения порознь.
Второй принципиальный аспект «этолого-термобиологических» исследований – разделение термобиологических показателей на «термофизиологические» и «термоэкологические». Если метрики из первой группы характеризуют физиологические потребности организма, то вторые – экологические возможности среды. Например, термофизиологические предпочитаемые температуры, исходя из определения («диапазон температур тела при полной активности, который рептилии направленно избирают и поддерживают у себя в данных экологических условиях»), будут наблюдаться только в условиях, когда может проявиться «полная активность» животного.
Однако в разных частях ареала и в разные сезоны погодные условия могут и не предоставить рептилии выбора из всего возможного диапазона температур, и ей приходится довольствоваться тем, что есть, и не проявлять полный диапазон температур при «полной активности». Соответственно, те температуры тела, которые имеет данная особь, не будут соответствовать видоспецифическим потребностям, но лишь текущим возможностям, уровню теплообеспечения среды. Следовательно, оцененные в поле температурные показатели не всегда можно воспринимать как соответствующие потребностям организма, т. е. их нельзя считать однозначно физиологическими, но, скорее, экологическими. С этих позиций опять же нельзя оправдать практику сравнивать даже репрезентативные выборки значений температуры тела из разных регионов, в разные сезоны и пр. В конечном итоге такой анализ имеет смысл сравнения разных температурных условий среды и ничего не может сообщить собственно о биологических свойства вида в данной части ареала или в данный сезон. Термобиологический смысл имеет только сравнение термофизиологических, но не термоэкологических показателей животного.
Эти положения развиваются в представленной выше статье В. А. Черлина и позволяют ему сделать вывод о стабильности важнейших видовых термофизиологических характеристик – о стабильности температурного диапазона для «полной активности» на всем протяжении видового ареала. К сожалению, эта интересная тема находится вне проблематики собственно терморегуляции и мы больше не будем ее касаться.
Здесь следует обсудить некоторые противоречия, подмеченные оппонентами. Первый упрек – в избыточном объеме теории. В рамках своего этолого-термобиологического направления автор рассматривает практически все формы поведения животных, хотя многие из них имеют очень косвенное отношение к терморегуляции или вовсе не имеют его (например, «термонейтральное поведение» или «добровольный перегрев»). Многообразие этих форм, а также переклассификации и уточнение их названий в последних публикациях не вносят определенности в предмет обсуждения. Важной задачей здесь видится однозначное выявление только тех форм поведения, которые имеют терморегуляционный смысл. На наш взгляд, это качество несет только треть из названных форм.
Вторая проблема состоит в том, что разграничение форм поведения рептилий зависит, как пишет автор, от «умения» (Черлин, 2012), что не может не настораживать. Например, без большого навыка трудно разобраться, когда (в состоянии «полной активности») движения рептилий «…быстры и… точны» (с. 35), а когда (в состоянии «неполной активности») их «движения недостаточно быстры, точны и результативны» (с. 44). Однако здесь существует еще и опасность логического круга, когда температуры тела и среды привлекаются для определения формы поведения рептилии. Рассмотрим ситуацию – на поверхности субстрата найдена неподвижная рептилия, к какой форме отнести ее поведение? Наблюдение утром (когда температура тела ниже температуры субстрата, Тт < Тсуб.) говорит о «нагревании»; днем (Тт > Тсуб.) имеем «баскинг», вечером (Тт = Тсуб.) – «остывание». Такое деление выполнить можно, но тогда нельзя охарактеризовать эти формы поведения через наблюдаемые значения Тт! Когда, определив с помощью показателя температуры тела форму поведения, мы будем характеризовать эту форму с помощью найденной Тт, то получим бессмысленную тавтологию, не более. Если мы хотим дать температурную характеристику той или иной форме поведения, то их разграничение должно выполняться по другим критериям. Разобраться в этом вопросе мешает то обстоятельство, что последние публикации В. А. Черлина в этом пункте имеют ссылки на те свои работы, в которых есть ссылки на оригинальные исследования. Оказывается довольно сложно выйти на источники с исходными данными.
Третий сложный момент состоит в критерии разграничения термофизиологических и термоэкологических показателей. Здесь важен не столько методический, сколько теоретический вопрос. В широком смысле любое значение термоэкологического показателя (например, Тт = 10 °С) имеет смысл физиологического, поскольку будет определять интенсивность обменных и прочих физиологических процессов. В узком же смысле термофизиологический параметр должен учитывать феномен терморегуляции – «полные потенциальные предпочтения» указывают именно на активный выбор определенных температур. Рассмотрим такие «понятные» термофизиологические показатели, как «диапазон оптимальных температур тела». Здесь возникает вопрос: почему физиологическая характеристика представлена не одним значением, но диапазоном? Не будем рассматривать вопросы точности определения термальных показателей в полевых условиях, но сойдемся на том, что значений оптимальной температуры может быть много. Какой причине приписать существование этих значений? Видимо, температурным условиям среды. Получается, что диапазон термофизиологических характеристик в строгом смысле является одновременно и термоэкологической характеристикой. Смысл термина размывается, что недопустимо для поиска точек соприкосновения разных исследовательских направлений. Можно предложить еще одну интерпретацию примера. Есть две видоспецифические «чисто» термофизиологические характеристики – верхний и нижний пороги оптимальных температур. Однако такое соображение означает, что следует выяснить физиологические механизмы поддержания (или хотя бы «стремления» к поддержанию) обеих этих границ. Поскольку у автора нет на этот счет содержательных предложений, видимо, предстоит существенная ревизия набора термофизиологических показателей.
Итак, в работах В. А. Черлина мы находим методологию поиска стабильных параметров терморегуляции – ориентируясь на поведение, оценивать температурные характеристики животных, которые при определенном угле рассмотрения можно интерпретировать как физиологические, обеспечивающие регуляцию состояния животных.
Направление эколого-термобиологических исследований, проводимых Н. А. Литвиновым, существенно отличается от представленного выше современной инструментальной базой. Термисторы для моментального определения температуры в любой части тела, имплантируемые в тело животного микрорегистраторы для долговременной регистрации температуры тела, логгеры для долговременной регистрации температуры субстратов – весь этот арсенал позволяет поднять количественные наблюдения по термобиологии рептилий на качественно иной уровень. Легкость слежения за индивидуальной температурой тела животного дает возможность авторам увидеть структуру ее динамики во всей простоте – именно как результат терморегуляторного поведения. Ориентируясь на диаграмму (см. рис. 2 в статье из настоящего сборника), они говорят всего о двух существенных формах терморегуляторного поведения – «нагревание до уровня оптимальной температуры» и «контроль за своей температурой»), а также о менее существенной форме – остывании. На этих эколого-эмпирических позициях основано скептическое отношение к более обширным классификациям форм термального поведения рептилий как не поддающихся точному определению и поэтому не имеющих термобиологического содержания.
Авторы «эколого-термобиологического» направления исследований используют довольно узкий общепринятый набор температурных показателей, ориентированных на определение в полевых условиях. Достаточно строгое и простое разделение форм терморегуляционной активности рептилий на две категории позволяет авторам подойти к статистическим оценкам выборок. Для характеристики дневного варьирования температуры тела принят типичный статистический набор – средняя арифметическая, стандартное отклонение, ошибка. Эти параметры довольно обширных выборок, полученные для разных условий, цветовых форм, размеров особей и пр., авторы сравнивают с помощью параметрических статистических критериев. Для характеристики процессов нагревания вполне обоснованно выбрана регрессия, правда, в отличие от средней, линии регрессии не снабжаются доверительными зонами и статистические сравнения коэффициентов регрессии не выполняются. Хотя авторы явно не проводят разделения термобиологических показателей на «физиологические» и «экологические», но в тексте принимается, что какие-то температуры могут быть оптимальными, какие-то нет. Поскольку термин «оптимальный» имеет в данном разделе науки только физиологический смысл, фактически авторы вводят указанное подразделение. Более того, сделана попытка количественно определить некое единственное значение температуры, названное «абсолютным температурным оптимумом», которое имеет смысл именно видоспецифической физиологической характеристики. Таким образом, авторы принимают, что терморегуляция (изученных видов) рептилии устроена не очень сложно и для ее описания не требуется большого количества регулирующих параметров. Практически достаточно только три – средняя (оптимальная) температура тела, «скорость нагревания» и «абсолютный температурный оптимум». Из контекста публикаций также следует, что указанные параметры могут меняться в соответствии с тем, какому «виду, подвиду, популяции, определенному времени года, возрасту, полу» эти параметры принадлежат. Естественно, это означает, что рассматриваемые характеристики (как термофизиологические, так и термоэкологические) могут широко варьировать. Судя по публикациям, авторы собрали довольно обширные материалы по 13 видам рептилий Волжского бассейна. Авторы не понаслышке знают о сложностях сбора полевого материала и некоторые из них высказывают явное удивление, когда читают, что представители другого из рассмотренных направлений исследовали основные термобиологические показатели у 50 видов рептилий.
Рассмотрим критику теперь уже второго направления термобиологических исследований, которая, как представляется, позволит выявить точки возможного соприкосновения с первым.
Можно назвать похвальным стремление максимально упростить схему терморегуляции, выявив в чистом виде минимально необходимое число поведенческих актов, направленных только на терморегуляцию. На наш взгляд, однако, авторы в этом переусердствовали. Во-первых, хоть и называли в статьях, но практически не рассмотрели некоторые важные формы поведения терморегуляции, как, например, «выход на поверхность» для первичного нагревания или же «сворачивание в клубок» при частичном остывании и др. Во-вторых, такая форма поведения, как «контроль за своей температурой», остается совершенно не расшифрованной, хотя по факту состоит из целой серии разнонаправленных поведенческих актов. Даже простой уход с открытой поверхности в легкие укрытия состоит из двух актов – собственно уход с прежнего места и выбор подходящего нового. И первый, и второй свершившиеся акты, видимо, соответствуют каким-то физиологическим потребностям (критериям), для описания которых их нужно сначала хотя бы назвать. Таким образом, для объяснения феноменологии, наблюдаемой с высокой точностью, следует расширить арсенал терморегуляторных типов поведения.
Разнообразные формы терморегуляторного поведения характерны для дневного времяпровождения. Очевидно, что авторы знают об этом скрытом разнообразии. Однако, рассчитывая средние дневные температуры тела, они фактически признают эти формы как неизбежный набор факторов случайного варьирования и регистрируют их только в форме дисперсии. В таком подходе нет методической ошибки, однако он накладывает определенные обязательства на авторов. Во-первых, статистическую характеристику должны получать все статистические параметры (включая коэффициенты регрессии и АТО), а не только средние. Во-вторых, сознательно вводя в поле рассмотрения множество разнородных случайных факторов, необходимо давать адекватную интерпретацию статистическим параметрам. Например, при сравнении двух регионов следует иметь в виду, что средняя арифметическая температура тела выражает не только и не столько характеристики вида, сколько условия среды, в которых этот вид обитает. К сожалению, авторы экологического направления обычно дают иную интерпретацию средним температурам тела, а именно – как видоспецифическим характеристикам.
Здесь мы вновь выходим на тему отделения экологических терморегуляционных характеристик от физиологических и их точного количественного оценивания. Отрадно, что авторы данного направления подошли к решению вопроса с помощью вариационной статистики, но решили задачу, на наш взгляд, неправильно. «Абсолютный температурный оптимум» определяется как «довольно нестабильный уровень температуры тела, достигаемый путем инсоляции, когда температура тела животного становится равной внешней температуре (обычно температуре субстрата)» (Литвинов, см. настоящий выпуск, с. ??). Уже первая оценка («довольно нестабильный») ставит под сомнение ценность этого показателя – если доверительные интервалы для температуры субстрата и тела будут иметь слабую трансгрессию, значит, никакой АТО и нет (однако в статьях авторы никогда не приводят статистических параметров этого показателя). Возникает и теоретическое возражение – как можно физиологически значимое состояние («оптимум»!) гелиотермных животных оценивать по температуре внешней среды? Нам кажется очевидным, что при разных температурных условиях среды рептилии (здесь в основном речь идет о видах умеренных широт) могут достичь физиологически оптимальных температур тела. Кроме этого, реальные наблюдения противоречат обсуждаемому феномену – сами же авторы неоднократно писали о том, что температура тела в среднем всегда выше температуры субстрата на 6 °С и больше. И тем не менее они с помощью «решения системы уравнений» пытаются получить какую-то характеристику. Несмотря на то, что данный обсуждаемый показатель нам кажется некорректным, сам подход – расчет ненаблюдаемого показателя – следует признать правильным.
Итак, в работах Н. А. Литвинова и др. мы находим инструментарий для эффективного поиска ключевых термобиологических характеристик – точные и адресные методы оценки температуры тела, опыт разнообразной статистической обработки больших многофакторных массивов данных и технологию расчета термофизиологических показателей, которые в полевых условиях могут и не наблюдаться.
После нашего обзора двух подходов их различия кажутся существенными. Однако, на наш взгляд, сильнее отличаются заблуждения их авторов, тогда как многие аспекты проблемы они улавливают одинаково. Во-первых, одни явно, другие контекстуально признают существование видоспецифического «оптимума» температур (как бы этот диапазон ни называли). Во-вторых, и те и другие почему-то не рассматривают процесс поддержания этого оптимума, собственно процесс регуляции, с позиций физиологии. Вот с этой базы и может стартовать синтез.
Наше предложение состоит в том, чтобы добавить строгости в выбор показателей терморегуляции на базе физиологической теории. Во-первых, необходимо найти строгий критерий для разделения термофизиологических (терморегулирующих) параметров и термоэкологических (терморегулируемых) переменных состояния животных. Во-вторых, научиться оценивать термофизиологические параметры как характеристики видоспецифичного способа терморегуляции.
Физиологическая теория регуляции в своей основе использует кибернетическую конструкцию – схему отрицательной обратной связи (см. обзор: Коросов, 2008): «...информация, поступившая обратно в управляющий центр, стремится противодействовать отклонению управляемой величины от управляющей» (Винер, 1983, с. 165); «...отрицательная обратная связь уменьшает расхождение между заданным и фактическим значением переменной... позволяет поддерживать стационарное состояние...» (Шмид-Ниельсен, 1982, с. 705). Если авторы обсуждаемых направлений согласны с тем, что рептилии осуществляют терморегуляцию, то на первом этапе становления консенсуса им необходимо признать существование некой «управляющей» величины, «штатного» значения параметра терморегуляции, например некоего значения температуры тела, выше которого животное «не хочет» нагреваться. Это есть в прямом смысле «термофизиологический показатель», который имеет смысл видоспецифичной физиологической константы, неизменной для группы животных одного статуса. Стоит вопрос: каким образом ее померить?
Текущую температуру тела рептилии нужно рассматривать как переменную, которая может принимать самые разные значения в зависимости от теплообмена с внешней средой (включая инсоляцию, тепловое излучение от предметов и от тела гада, теплообмен, конвекцию). Температура тела почти всегда представляет собой термозависимую, термоэкологическую переменную терморегуляции. Эта величина связана с проявлением двух начал – биологической потребностью рептилии в росте температуры тела до «желательного» уровня, обеспечивающего обмен веществ, и возможностью среды предоставить животному тот или иной объем нужной теплоты.
В общем случае самая богатая статистика полевых измерений температуры не в состоянии оценить уровни термофизиологических параметров.
Средние арифметические или медианы, рассчитанные по большому массиву данных, не годятся по той причине, что они характеризуют весь диапазон изменчивости внешних термальных условий и имеют к характеристикам термопреферендума косвенное отношение. Максимальные и минимальные значения по своему смыслу гораздо ближе к термофизиологическим порогам, но также не годятся для их оценки, поскольку имеют очень плохое статистическое обоснование. Таких значений, во-первых, всегда мало, во-вторых, они могут быть связаны с неучтенными патологическими состояниями животных, в-третьих, рост точности измерений постоянно раздвигает границы варьирования. Например, долгое время считалось, что максимальная температура обыкновенной гадюки составляет 34° (Saint-Giron, 1975, 1978), потом – 34.5° (Spaendonk et al., 1990; Дробенков, 1990), большие выборки дали величину 35° (Коросов, 2010), а телеметрия показала, что возможно кратковременное повышение до 37.5° (Литвинов, Четанов, 2014). Всего 15 крайних вариант дали размах в 3.5°С! Однако физиологический параметр должен быть определен гораздо точнее, поскольку организм обычно имеет очень высокий порог чувствительности. Например, терморецепторы гипоталамуса, запускающие реакцию терморегуляции у млекопитающих, чувствительны к отклонениям в доли градуса (Физиология…, 2003; Бачериков и др., 2006). Высокая чувствительность к перепадам температуры нужна для того, чтобы на фоне большой инерционности обратной связи (когда еще запустятся и осуществлятся реакции терморегуляции…) колебания температуры тела не были слишком большими. Ведь даже у млекопитающих при пороговой чувствительности 0.41 °С варьирование собственно температуры тела может составлять несколько градусов.
Чтобы выйти на оценку штатных параметров терморегуляции, необходимо понять, для чего они нужны и как сказываются на состоянии животных. Эти константы призваны менять функционирование особи при неблагоприятном уровне теплообеспечения, вызывая приспособительные реакции для улучшения ситуации с теплообменом. Можно грубо подразделить реакции терморегуляции на биохимические (акклимация), физиологические (полипноэ, вазомоторика, пульс), морфологические (смена позы), этологические (смена форм поведения). В полевых условиях удается наблюдать только два последних типа реакций, но и их достаточно, чтобы найти методику измерения термофизиологических параметров. Что означает с физиологической точки зрения смена форм поведения, например переход с открытой поверхности в легкое укрытие? Прямая инсоляция ведет к росту температуры тела до того момента, пока она не станет равной физиологически заданному порогу, т. е. внутренняя переменная произвольно меняется только до того момента времени, пока не станет равной внутреннему пороговому параметру. В этот момент осуществляется тот или иной поведенческий акт. Отсюда выводится предлагаемый нами методический прием (смысл всей публикации):
Значение температуры тела, которое наблюдается в момент смены форм терморегуляторного поведения, можно принять как оценку термофизиологического параметра терморегуляции.
Технологически это означает постоянное наблюдение за животным с вживленными датчиками температуры и фиксация моментов времени (значит, и температуры), когда происходит смена форм активности. Несмотря на кажущуюся простоту этой методики, нам неизвестны случаи ее применения (кроме примера, рассмотренного ниже). Причина состоит в том, что для проведения таких измерений нужны: а) теория физиологической регуляции, б) теория смены форм поведения, в) точный инструментарий и опыт измерения текущей температуры тела рептилий. На наш взгляд, теперь появляются предпосылки для синтеза указанных компонентов и выхода на новый уровень совместных термобиологических исследований.
Первой задачей коллективных действий может быть обсуждение минимального списка форм терморегуляционного поведения, для которых следует в первую очередь определять «управляющие» параметры, а также планирование такого рода полевых наблюдений. На втором плане просматриваются вопросы изучения зависимости физиологических констант от статуса особи (пола, возраста, акклимации, упитанности, стадии питания, места обитания, региона, вида и др.).
В качестве близкого примера термобиологических исследований нового образца приведем наш опыт оценки температурной чувствительности обыкновенной гадюки в лабораторных условиях (Коросов и др., 2003). В целях большей иллюстративности мы по-иному обработали фрагмент исходных данных.
Цель опытов состояла в том, чтобы определить уровень излучения, который обыкновенная гадюка воспринимает как сигнал к переходу к новой форме поведения – прекращения остывания в норе и началу нагревания на поверхности почвы под лучами солнца. Хотелось понять термальную причину, по которой змея покидает убежище. Эту работу не удалось закончить по техническим причинам, но она годится для иллюстрации смысла предлагаемой методологии термобиологических исследований.
В лаборатории полевого стационара на протяжении 2 месяцев выполнен 21 опыт над 8 животным с длиной тела от 11 до 62 см. С момента питания животных проходило разное время; в 7 случаях животные питались несколько дней назад (1, 5, 7, 8, 11), в 14 опытах – более 14 дней назад.
Опыты проходили в камере размером 60*30*40 см с клеенчатым полом с нанесенными линиями измерительной сантиметровой шкалы. У одной из меньших стенок помещали термометр для оценки температуры пола (в разных опытах от 15 до 27 °С). Эту температуру принимали равной температуре тела змеи. У противоположной стенки был смонтирован «излучатель» – закопченная жестяная банка с лампой накаливания внутри и закрытым ртутным термометром снаружи. В сеть электропитания лампы входил реостат, регулировка которого позволяла контролировать температуру излучателя. Во время опыта животное помещали в камеру и через час (когда животное успокаивалось) начинали постепенно повышать температуру излучателя примерно на 1–2 градуса в минуту. В тот момент, когда змея начинала двигаться к источнику излучения («положительный термотаксис»), фиксировали температуру пола (тела, Тт), излучателя (Ти) и расстояние (L) ближайшей части тела змеи до излучателя. В соответствии с законами физики мощность теплового потока от источника составит: R = s × (Ти4 – Тт4), где s = 5,6687 × 10-8 (Дж × м-2 × с-1 × К-4) постоянная Стефана — Больцмана (Процессы…, 2015). На удалении от источника мощность будет падать пропорционально расстоянию. Приняв диаметр излучателя за 5 см, а сечение луча теплового излучения за 1 см2, несложно показать (из соотношения подобных треугольников), что на расстоянии L от источника сечение теплового луча составит L/10, т. е. мощность теплового потока уменьшится в L/10 раз. Очевидно, ситуация с переносом тепла сложнее, но на наших дистанциях (5–35 см) нужно было учесть только эффект уменьшения мощности излучения с расстоянием, а не рассчитать истинные ее значения. Исходя из этих пропорций по результатам опыта были рассчитаны показатели «тепловой чувствительности» обыкновенной гадюки (табл., рис.).
Таблица. Характеристики и результаты опыта по оценке «тепловой чувствительности» обыкновенной гадюки (juv – сеголетки, ad- – взрослые голодные, ad+ – взрослые сытые)
| Т тела | Т излучателя | Расстояние | Теплота (q, дж / кВ.м×с) | ||||||||||
| n | мин. | макс. | мин. | макс. | мин. | макс. | мин. | макс. | M | Me | S | Cv | |
| juv | 4 | 15 | 25 | 38 | 63 | 5 | 28 | 0.09 | 0.29 | 0.16 | 0.14 | 0.089 | 54 |
| ad- | 10 | 19 | 27 | 28 | 50 | 10 | 35 | 0.01 | 0.11 | 0.05 | 0.05 | 0.034 | 65 |
| ad+ | 7 | 14 | 26 | 22 | 48 | 5 | 26 | 0.00 | 0.14 | 0.04 | 0.01 | 0.05 | 144 |
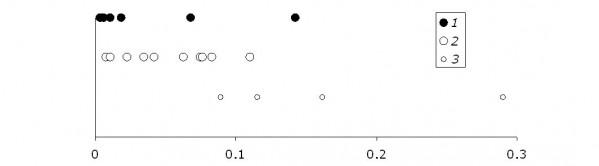
Рис. Изменчивость показателя «тепловая чувствительность» обыкновенной гадюки: 1 – взрослые сытые, 2 – взрослые голодные, 3 – сеголетки
Иллюстрации показывают, что для гадюк разных категорий условия опытов (диапазоны температур тела, излучателя и расстояний от источника) не различались. Тем не менее показатели «тепловой чувствительности» для гадюк с разным статусом отличаются. Самыми «инертными» оказались сеголетки (qjuv> qad), самыми реактивными – недавно поевшие гадюки (qad+> qad-), правда, о значимости различий пока говорить не приходится. Само отличие в реактивности говорит о том, что «штатный параметр» «желательной тепловой нагрузки» не остается постоянным, но меняется в зависимости от состояния животных. Кроме того, широкая изменчивость замеров указывает на необходимость существенного совершенствования методики как оценки тепловой нагрузки, так и оценки температуры тела животных.
Напоследок необходимо обсудить вопрос о значении тех новых материалов по термобиологии, которые могут быть получены в результате совместной работы, – не разрушит ли это знание имеющиеся авторские наработки? Конечно, без появления и разрешения противоречий не обойтись, ведь предлагаемая технология уже указывает на недостатки ранее использованных методик. Однако, на наш взгляд, никакого отказа от прежних наработок не произойдет. Температурные диапазоны форм активности животных, изучаемые В. А. Черлиным, получат фактические, а не статистические границы. Выявленные Н. А. Литвиновым отличия статистических термобиологических характеристик разнообразных групп животных получат более определенную факторную интерпретацию.
Для начала же следует провести хотя бы одно совместное наблюдение и прийти к согласию при его интерпретации.
Библиография
Бачериков А. Н., Кузьминов В. Н., Ткаченко Т. В., Назарчук А. Г. Современные представления о системе терморегуляции // Вісник психіатрії та психофармакотерапії. 2006. № 1. С. 178–182.
Винер Н. Кибернетика. М.: Наука, 1983. 342 с.
Дробенков С. М. Терморегуляторное поведение гадюки обыкновенной (Vipera berus L.) в условиях Белоруссии / Деп. ред. журн. Известия АН БССР. Сер. биол. наук. Минск, 1990. 18 с. Деп. ВИНИТИ 03. 01. 91. № 50–В91.
Коросов А. В. Экология обыкновенной гадюки (Vipera berus L.) на Севере (факты и модели) . Петрозаводск: Изд-во ПетрГУ, 2010. 264 с.
Коросов А. В., Сапунова О. В., Хилков Т. Н. Реакция обыкновенной гадюки на тепло // Змеи Восточной Европы. Тольятти, 2003. С. 34.
Литвинов Н. А., Четанов Н. А. Непрерывная регистрация температуры тела обыкновенной гадюки летом // Известия Самарского НЦ РАН. 2014. Т. 16. № 5(1). С. 430–435.
Литвинов Н. А. К вопросу об оценке температуры тела рептилий, ее соотношении с внешней температурой и адаптивности размеров и окраски // Принципы экологии. 2015. Т. 4. № 1. С. 1.
Петерсдорф Р. Г., Рут Р. К. Глава 8. Нарушения терморегуляции // Внутренние болезни / ред. Е. Браунвальд. М.: Медицина, 1993. 446 с.
Процессы переноса тепла // Политех в сети. URL: http://www.webpoliteh.ru/ subj/fizika/100-lekciya-3-processy-perenosa-teploty.html (дата обращения 5.03.2015)
Физиология человека / Под ред. В. М. Покровского, Г. Ф. Коротько. 2-е изд., перераб. и доп. М., 2003. 656 с.
Черлин В. А. Термобиология рептилий. Общая концепция . СПб.: Русско-балтийский информационный центр БЛИЦ, 2012. 362 с.
Черлин В. А. Тепловые адаптации рептилий и механизмы их формирования // Принципы экологии. 2015. Т. 4. № 1. С. 1.
Четанов Н. А., Литвинов Н. А., Бакиев А. Г. Возможности применения показателей вариации в исследовании термобиологии рептилий // Поволж. экол. журн. 2009. № 3. С. 263–269.
Шмидт-Ниельсен К. Физиология животных. Приспособление и среда. М.: Мир, 1982. Т. 1–2. 800 с.
Saint-Girons H. Observations preliminaires sur la thermoregulation des Viperes d'Europe // Vie et Milieu. 1975. Vol. 25. Fasc. 1. Ser. C. P. 137.
Saint-Girons H. Thermoregulation comparee des viperes d'Europe. Etude biotelemetriaue // Terre et vie. 1978. 32. № 3. P. 417–440.
Spaendonk V. G., Damme V. R., Verheyen R. F. Thermal ecology of the adder (Vipera berus L. ) // Belg. J. Zool. 1990. Suppl. 120. № 1. P. 62–63.
Благодарности
Автор признателен В. А. Черлину и Н. А. Литвинову за обсуждение рукописи.

 © 2011 - 2026
© 2011 - 2026