



Введение
В литературе имеется большой объем материалов, описывающих различные стороны адаптации рептилий к температурным условиям среды. В свое время было опубликовано несколько крупных статей и сводок по этой теме (Cowles, Bogert, 1944; Brattstrom, 1965; Tempelton, 1970; Cloudsley-Thompson, 1971). На том этапе исследований они касались в основном феноменологии явления – чисто описательных признаков, констатирующих некоторые внешние проявления терморегуляции рептилий, но в которых не были учтены эндогенные характеристики, связанные у пресмыкающихся, в частности, с механизмами регуляции температуры тела. Такое описание адаптаций рептилий, конечно, имеет право на существование, но оно мало помогает в понимании термобиологии этих животных.
Несколько лет назад вышла книга, посвященная непосредственно тепловым адаптациям рептилий (Angilletta, 2009). В ней подробно рассматриваются вопросы, связанные с температурной изменчивостью, чувствительностью, терморегуляцией, акклимацией, коадаптациями, эволюцией термобиологической сферы, сравниваются различные математические модели, описывающие разные стороны тепловых адаптаций, и т. п.
В последнее время в литературе стали встречаться два термина, описывающие степень проявленности терморегуляции у рептилий и/или формы их адаптации к термальным условиям среды: thermoregulators (терморегуляторы – животные, которые активно и эффективно регулируют температуру тела) и thermoconformers (термоконформеры – животные, которые не проявляют активных усилий по регуляции температуры тела, т. е. оперативная температура почти совпадает с реальной температурой тела живых животных в данных условиях. Оперативная температура – температура тела, которая устанавливается как проявление «термального равновесия» между телом животного и «термальным фоном» среды в случае, если бы у животного не существовало никаких проявлений физиологической и поведенческой терморегуляции. Наиболее правильно было бы определять ее как температуру мертвого тела, помещенного в ту же среду, где находится и живое животное. Но на практике, по понятным причинам, ее определяют не так, а как температуру макета тела животного, сделанного в натуральную величину из специально подобранных материалов, сходных по теплоемкости и теплопроводности с телом животного, часто – из меди и ее сплавов (Bakken, 1992; и мн. др.). Другими словами, температура тела в этом случае практически пассивно следует за температурами среды. Нужно признать, что относительные термоконформеры среди рептилий, безусловно, существуют (Alcala, Brown, 1966; Hertz, 1974; Huey, 1974; Huey, Webster, 1976; Rand, Humphry, 1968; Ruibal, 1961; Ruibal, Philibosian, 1970; Stebbins et al., 1967; и др.). Но они именно относительные, поскольку, чаще всего, такие виды почти не регулируют температуру тела в границах тех диапазонов температур, в которых они оказываются в природных условиях и которые при этом почти соответствуют эндогенно потребным температурам тела данных видов рептилий, и/или динамика температур среды обеспечивает эндогенно потребную динамику температуры тела без активного применения специальных механизмов оперативной регуляции (оперативный – приспособленный для правильного и быстрого практического выполнения тех или иных задач, умеющий справляться с практическими задачами – Толковый словарь…, 1938). Сам факт того, что терморегуляционные реакции у них не всегда четко наблюдаются, – это не проявление отсутствия терморегуляции или наличия какого-то специфического ее механизма, а лишь большая или меньшая степень редукции существующего, стандартного механизма терморегуляции, связанная с особенностями физиологии и обитанием в тропических климатах, где температуры среды не опускаются слишком низко и/или не поднимаются слишком высоко и где сезон таких «теплых» условий имеет в году большую продолжительность (теплый сезон с температурами около 28–32° длится практически весь год).
Кроме того, используется еще одна пара терминов: thermal specialists (рептилии, которые удерживают температуру тела в узком диапазоне; близко к понятию «стенотермы») и thermal generalists (рептилии, температура тела которых находится в широком диапазоне, т. е. они не так четко регулируют температуру тела; близко к понятию «эвритермы»). Однако и это разделение не дает сколь-нибудь важной информации, поскольку если, например, два разных вида рептилий удерживают температуру тела в достаточно узком диапазоне, но делают это за счет сильно различающихся эндогенных механизмов или вследствие того, что температуры среды в месте обитания подвержены слабым колебаниям, то эти ситуации надо четко различать.
Несмотря на большое количество публикаций, касающихся термальных адаптаций рептилий, нам кажется возможным и необходимым еще раз обратиться к этой теме. Рассматривая способы адаптации пресмыкающихся к термальным условиям, важно, видимо, учесть, кроме прочего, эндогенные характеристики этих животных, связанные, в частности, с терморегуляцией, во всей сложности их внутренних и внешних взаимосвязей. Выявление общих эколого-физиологических и физиолого-экологических закономерностей позволит описать механизмы этих адаптаций, а уже как следствие – создать естественную физиолого-экологическую модель адаптации рептилий к температурным условиям, т. е. выявить основные пути формирования и развития этих адаптаций.
Материалы
Материалом для данной статьи служат результаты сорокалетней работы по термобиологии рептилий (Черлин, 1989, 2012а, 2014). Сбор первичной информации в полевых условиях происходил в пустынях и горах Средней Азии на территориях заповедников – Бадхызского (Кушка, южная Туркмения), Репетекского (восточные Каракумы, Туркмения), Сюнт-Хасардагского (Кара-Кала, юго-западная Туркмения), Кугитангского (хребет Кугитанг, южная Туркмения), Кызылкумского (восточные Кызылкумы, Узбекистан), Нуратинского (Нуратинский хребет, Узбекистан), во многих других точках – на Красноводском плато (западная Туркмения), в окрестности озера Айдаркуль (Узбекистан), на хребте Бабатаг (Узбекистан), на Гиссарском хребте (Узбекистан), в окрестностях и на территории гор. Ташкента (Узбекистан), на хребте Копетдаг (южная Туркмения). Также исследования велись в других природных зонах: в Ленинградской области (на Карельском перешейке), в Карелии (заповедник «Кивач»), на острове Кунашир (Дальний Восток), а также в болотах дельты реки Миссисипи (штат Луизиана, юг США), в низинных тропических лесах национального парка «Уджун-Кулон» (западная оконечность острова Ява и близлежащие острова, Индонезия), в горных лесах у пос. Сите-Гунунг (северо-западная часть о. Ява, Индонезия). В полевых условиях была исследована термобиология 50 видов рептилий фауны бывшего СССР и 8 тропических видов. Работы проводились в течение 22 полевых сезонов (в общей сложности более 40 месяцев). Серии измерений по каждому из изученных видов в зависимости от их доступности составляли от 30–40 до нескольких сотен. В отдельных случаях число измерений могло превышать тысячу. В общей сложности были обработаны материалы 6372 встреч рептилий разных видов в природе, более 20000 измерений среды и тела рептилий при этих встречах и около 90000 измерений, полученных на наших метеорологических площадках.
Более 250 экземпляров пойманных в разное время рептилий интересующих видов привозили в места, где возможно было их удовлетворительно содержать в неволе, с ними проводили специальные исследования по термобиологии. Такими местами в разное время были террариумные отделы Ленинградского и Ташкентского зоопарков, Зоологический институт АН СССР, герпетарий АОЗТ «Биопрепарат» и домашние террариумы в Санкт-Петербурге.
Кроме того, в течение более 30 лет в этих организациях мы вели наблюдения за другими содержавшимися в неволе видами рептилий фауны бывшего СССР – за 63 видами (более 2,5 тысячи экземпляров), из которых 38 видов змей (13 видов ядовитых), 20 видов ящериц и 4 вида черепах, а также за 115 тропическими видами (около 760 экземплярами), включая 6 видов сухопутных и 11 видов водяных черепах, 9 видов крокодилов, 44 вида ящериц и 45 видов змей (13 видов ядовитых).
Методы
Основной отличительной чертой авторской методологии сбора информации по термобиологии рептилий является выделение статусов и форм их активности (Черлин, 1991, 2010, 2013б), в которых стратегии отношения к температуре различны (раздел «Статусы и формы активности у рептилий»).
Наиболее полный материал по какому-либо виду рептилий мы получали, если изучали его в полевых условиях и дополняли наблюдениями, измерениями и опытами при содержании их в неволе. Эти варианты исследований гармонично дополняют друг друга: с одной стороны, в хорошо организованном и оборудованном террариуме, где контроль и наблюдение за животными во всех фазах активности облегчается, легко наблюдается то, что в поле можно не заметить в связи со скрытностью и осторожностью животных, с другими осложняющими обстоятельствами; с другой стороны, в поле поведение животных более полное, демонстрирующее в естественном состоянии сложные связи активности и поведения рептилий с условиями среды, «завуалированные» или неявно проявляющиеся в условиях неволи, где параметры среды задает человек.
Кроме того, в полевых условиях в одних и тех же «базовых» географических точках стационарные исследования проводили по-возможности в течение 3–5 лет в разные сезоны года, а также в разных частях ареала и в разных экологических условиях.
Окончательный вариант метода сбора информации в поле и в лаборатории, сформировавшийся в результате многолетних исследований, представляет собой комплекс из трех взаимосвязанных блоков: 1) подробного изучения климатических условий биотопов и микростаций в местах обитания рептилий данного вида, 2) различных вариантов наблюдений за поведением пресмыкающихся с выявлением форм терморегуляционного поведения и форм активности, 3) изучения суточной и сезонной динамики температуры тела животных в связи с выявленными формами активности.
Обработка материалов встреч и наблюдений производилась по специально разработанным методикам отдельно по формам поведенческой активности (Черлин, 2010, 2013б, 2014).
Результаты
ОСНОВНАЯ НАПРАВЛЕННОСТЬ ТЕРМОРЕГУЛЯЦИИ РЕПТИЛИЙ
Начать разговор о таком комплексном понятии, как тепловые адаптации, нужно с принципов организации терморегуляции у рептилий.
Рептилии – эктотермные животные, температура их тела поддерживается в основном за счет факторов внешней среды, они не имеют более или менее значимого эндогенного механизма для повышения температуры тела. Практически все аспекты жизнедеятельности рептилий оказываются температурно зависимыми: биохимия, физиология, поведение, экология и т. п. (Van Damme et al., 1991; Shine, Harlow, 1993; Bauwens et al., 1995; Черлин, 2014; и мн. др.), поэтому регулирование температуры тела для пресмыкающихся – одна из самых главных задач в обеспечении их жизнедеятельности.
Какова основная направленность регуляции температуры тела у рептилий?
Один из аспектов терморегуляции направлен на реализацию принципа стабилизации высокой температуры тела в эволюции позвоночных животных: в прогрессивной эволюции позвоночных животных наблюдается направленность на стабилизацию высокой температуры тела (Рюмин, 1939, 1940; Черлин, 1990, 2012в). Этот принцип необходим, в частности, для энергетической оптимизации работы ферментных систем организма (Черлин, 1990, 2012в), способствует более интенсивной работе внутренних органов, анализаторов, а также энергообеспеченности активности на клеточном уровне (Черлин, 1988в, 2014). Эта стабилизация может осуществляться либо за счет тепловых факторов внешней среды, либо за счет эндогенной теплопродукции. Рептилии демонстрируют максимальное среди позвоночных животных развитие эффективной терморегуляции за счет экзогенных тепловых факторов благодаря, прежде всего, прекрасно развитому комплексу терморегуляционного поведения. Отчасти поэтому они – высшие эктотермы.
Следовательно, важная направленность терморегуляции рептилий – поддержание определенной суточной и сезонной динамики температуры тела, в рамках которых должен иметься промежуток времени с высокой температурой тела. Эта необходимость периодически нагреваться наблюдается практически у всех пресмыкающихся. Бывают, правда, и крайне редкие исключения. Например, высокоспециализированные виды, населяющие «холодные» географические области, которые вообще крайне редко прогреваются солнцем, – бирманская гадюка Azemiops feae (Kardong, 1986; Orlov, 1997; и др.), или живущая в перманентно «холодных» стациях бразильская змеиношейная черепаха Hydromedusa maximiliani (Souza, Martins, 2006), или мадагаскарские гекконы рода Uroplatus и др. Хотя и здесь не все так однозначно. По устному сообщению известного террариумиста и герпетолога С. Рябова (Московский зоопарк), при содержании в неволе, например, бирманских гадюк, которые, действительно, в природе практически никогда не греются (во всяком случае, никто и никогда этого не наблюдал у этих облигатно ночных змей), выясняется следующее: когда в террариуме им предоставляется возможность нагреваться, они делают это с большим удовольствием и подолгу. Правда, при этом они в 100 % случаев через короткое время погибают в результате бурного развития у них гельминтов.
Другой аспект терморегуляции у рептилий направлен на реализацию закона автономизации жизнедеятельности живой системы: эволюционное развитие живых систем направлено на уменьшение зависимости их жизнедеятельности от внешних для них факторов (Черлин, 2012б).
Реализация этого закона выражается в своеобразной форме связи температуры тела рептилий со средой: благодаря активному использованию и развитию специфических комплексов физиологических и поведенческих регуляторных реакций рептилии
1) усиливают оперативную связь температуры тела с микроклиматическими термальными факторами среды, но за счет этого одновременно
2) ослабляют зависимость своей жизнедеятельности от климатических условий географического региона обитания.
Таким образом, еще одна важная направленность терморегуляции рептилий – удовлетворение эндогенных потребностей рептилий во временной стабилизации температуры тела в течение некоторой части суток и в реализации суточной и сезонной динамики температуры тела за счет максимально эффективной, оперативной связи температуры тела с тепловыми факторами среды (тепловые факторы среды – факторы, влияющие на температуру тела животного путем теплового излучения, конвекции и теплообмена при контакте).
КЛЮЧЕВЫЕ ПОНЯТИЯ ТЕРМОБИОЛОГИИ РЕПТИЛИЙ
Для того чтобы понять, как происходит процесс адаптации рептилий к тепловым условиям внешней среды, необходимо иметь представление о том, как организована структура термобиологических характеристик у рептилий, в чем состоят их термальные потребности, за счет чего и как они могут удовлетворяться. Именно это в сжатой форме мы описываем в данном разделе (подробно см.: Черлин, 2014).
Статусы и формы активности у рептилий
При изучении термобиологии рептилий выяснилось, что одним из важных ее элементов являются понятия активности, ее статусы и формы (Черлин, 2014).
Активность, sensu lato, – всеобщая характеристика живых существ, их собственная динамика как источник преобразования или поддержания ими жизненно значимых связей с окружающим миром (Дудьев, 2008. Стр. 17);
– фундаментальное свойство живых организмов, выражающееся в их способности, преодолевая и используя внешние воздействия для реализации своих потребностей, целесообразно реагировать на факторы внешней среды (Покровский, 2005. Стр. 24).
Первое упоминание о разделении активности рептилий на неактивное и активное состояния, последнее из которых делится на полную и неполную активность, отмечено в работе С. С. Либерман и Н. В. Покровской (1943), хотя в ней авторы не называли описанные ими состояния «статусами активности» или какими-то другими специальными терминами. Статусы активности – относительно однородные состояния животных, в которых они могут в той или иной мере полноты осуществлять комплексы поведенческих актов, а) реализующих жизнеобеспечение особей (индивидуальная безопасность, терморегуляция, питание и пр.), б) активно включающих их в жизнь популяций (поддержание социальной структуры, охрана, размножение и пр.) и в) активно включающих их в жизнь биоценозов (пищевые цепи, межвидовые взаимодействия и пр.).
Но прежде чем говорить о статусах и формах активности, нужно определить «вспомогательный» термин, который будет встречаться в статье.
Рутинная активность – весь комплекс ежедневной, обычной, нормальной активности (normal activity – Cowles, Bogert, 1944; Huey, 1982; и др.) животного (в том числе и двигательной), направленный на его жизнеобеспечение, включающий регуляционные поведенческие реакции, а также поведение, связанное с питанием, коммуникациями, размножением, режимами активности и отдыха и т. п. Поскольку рутинная активность «связывает» жизнедеятельность животного с абиотическим и биотическим окружением, регуляция оптимальных (или хотя бы просто адекватных, допустимых) условий реализации рутинной активности для рептилий – задача первостепенной важности.
Состояние полной активности – животные находятся в состоянии, в полной мере обеспечивающем эффективное протекание всех основных поведенческих актов, характерных для рутинной активности (локомоции, терморегуляция, питание, сексуальность, территориальность, защита, коммуникации и т. п.). В этом состоянии движения рептилий активны, быстры (адекватно видовым особенностям) и точны. Состояние неполной активности – животные находятся при температурах тела, недостаточно высоких для перехода в состояние полной активности; при этом рептилии могут совершать ряд поведенческих актов, характерных для полной активности (локомоции, даже питание), но набор их неполный, реактивность, скорость и точность движений понижены. Терморегуляторные поведенческие реакции в переходных состояниях проявляются, но они направлены почти исключительно на повышение температуры тела до уровня перехода в состояние полной активности, на ускорение нагревания тела или замедление его остывания.
Формы активности – это комплекс поведенческих реакций, с помощью которых реализуется специфическая функциональная нагрузка каждого из статусов активности, осуществляется связь жизнедеятельности животных с тепловым фактором. У всех групп пресмыкающихся можно выделить стандартные формы активности. Они чрезвычайно важны тем, что, во-первых, в каждой из них рептилии поддерживают какую-то одну из сторон отношения к температуре: либо имеют возможность не обращать на нее внимания, либо удерживают на каком-то определенном уровне, либо направленно ее повышают или понижают, либо противодействуют избыточному нагреванию или слишком быстрому остыванию. Во-вторых, в каждой из форм активности рептилии имеют возможность реализовать и реализуют вполне конкретный набор поведенческих актов, важных для жизнедеятельности индивидуального животного, популяции и вида в целом: активные локомоции, питание, эффективная защита, проявление полового поведения и т. п. Выявление форм активности, таким образом, является одновременно и описанием важнейших принципов связи жизнедеятельности рептилий с температурой, что, в свою очередь, позволяет более полно и эффективно исследовать температурные предпочтения и границы, физиологические закономерности, связанные с температурой, и т. п.
Формы активности рептилий в неактивном состоянии – ночной покой, анабиотические состояния (гибернация, эстивация).
Формы активности рептилий в состоянии полной активности – термостабилизирующее поведение, термонейтральное поведение, терминг, добровольный перегрев, дневной отдых.
Термостабилизирующее поведение – форма активности, при которой особи реализуют комплекс поведенческих актов, направленных на удовлетворение своих индивидуальных потребностей, видоспецифичных потребностей жизнеобеспечения популяции и взаимодействие с биотическим окружением, т. е. выполняют активную функцию в биоценозе (питание, внутривидовые коммуникации, связанные с размножением, территориальностью, поддержанием социальной структуры популяции и т. п., межвидовые коммуникации – охрана территории и т. п.); температура их тела при этом находится (активно удерживается или пассивно оказывается) в определенном нешироком диапазоне, внутри которого стремится оказаться в еще более узких рамках. Если у животных в середине дня активность прерывается, например, в связи со слишком высокими температурами среды, то можно выделить первичное термостабилизирующее поведение (до ухода в убежище днем) и вторичное термостабилизирующее поведение (после выхода из дневного убежища во второй половине дня). Термостабилизирующее поведение характерно только для игуаноморфных, сцинкоморфных, веритеницевых и варановых ящериц.
Термонейтральное поведение – форма активности, при которой рептилии, как и при термостабилизирующем поведении, реализуют ряд поведенческих актов, направленных на удовлетворение своих индивидуальных потребностей, видоспецифичных потребностей жизнеобеспечения популяции и взаимодействие с биотическим окружением; но, в отличие от термостабилизирующего поведения, температура тела при этом может направленно не поддерживаться в узком диапазоне, оказываться часто существенно ниже модальной (предпочитаемой) и поэтому варьировать в довольно широких пределах. Термонейтральное поведение характерно только для гаттерии, гекконовых ящериц, змей, крокодилов и черепах.
Терминг (therming) – форма активности, при которой рептилии греются, крайне редко отвлекаясь на другие формы активности (питание, социальные контакты и т. п.). Температура тела при этом удерживается примерно на одном, достаточно высоком уровне в диапазоне термостабилизации. Терминг наблюдается только у гаттерии, гекконовых ящериц, змей, крокодилов и черепах. Терминг – чрезвычайно важная форма поведения, поскольку рептилии, для которых она характерна, именно при ней активно и направленно поддерживают температуру тела в диапазоне термостабилизации и набирают необходимое «количество теплоты» для протекания важнейших физиологических процессов (переваривания корма, гаметогенеза, беременности). При этом необходимо различать термины «терминг» и «баскинг». Баскинг (basking) – форма терморегуляционного поведения (а не форма активности), при которой рептилии греются, т. е. тем или иным способом получают тепло; температура тела при этом может подниматься, оставаться на одном уровне или опускаться. Он может наблюдаться при разных формах активности и у всех групп пресмыкающихся.
Добровольный перегрев – форма активности, при которой животные добровольно находятся в зоне интенсивного поступления тепла, температура их тела при этом выше диапазона термостабилизации. Животные при добровольном перегреве специальными (поведенческими и физиологическими) приемами противодействуют повышению температуры тела, замедляют скорость ее повышения до верхнего предела температуры активного состояния. Эта форма поведения в норме встречается не так уж часто и связано это обычно с некими особыми случаями. Например, самец степной агамы весной в период половой активности очень серьезно относится в охране своей территории и пытается как можно дольше, даже в жаркую середину дня, оставаться на ветках кустов. Беременные самки или животные, проглотившие большую добычу, иногда задерживаются в зоне сильного нагрева, что вызвано соответствующими, специфическими физиологическими потребностями. Могут быть и другие случаи. Эта форма поведения характерна только для игуаноморфных, сцинкоморфных, веретеницевых и варановых ящериц.
Формы активности рептилий в состоянии неполной активности – формы активности, при которых температура тела животных ниже температуры, обеспечивающей полную активность; при этом рептилии стремятся либо повысить ее до необходимого высокого уровня и перехода в состояние полной активности, либо замедлить остывание. К таким формам активности относятся: нагревание, остывание, псевдотермостабилизирующее поведение.
Псевдотермостабилизирующее поведение – форма активности, которая наблюдается у рептилий в условиях, когда термальные факторы среды достаточно длительное время не позволяют температуре тела повыситься до температуры полной активности. В этих прохладных условиях рептилии проводят иногда продолжительное время, ожидая появления солнца или начала влияния другого источника активного поступления тепла. При этой форме поведения животные, как и при термостабилизирующем поведении, реализуют ряд важных поведенческих функций (питание, локомоции), но набор их неполный, делается это не так активно, как при термостабилизирующем поведении, движения недостаточно быстры, точны и результативны. Их основное внимание направлено, прежде всего, на отслеживание возможности нагреться. Мы наблюдали такое поведение у рептилий, живущих в условиях с прохладной и изменчивой погодой: например, у живородящих ящериц Zootoca vivipara в Ленинградской области и в Карелии, у агам Чернова Laudakia chernovi на высоте более 2000 м над уровнем моря на хребте Кугитанг, ранней весной в Каракумах у песчаных круглоголовок Phrynocephalus interscapularis. Эта форма поведения характерна только для игуаноморфных, сцинкоморфных и веритеницевых и варановых ящериц.
Возможные суточные последовательности форм активности у разных видов рептилий приведены на рис. 1.
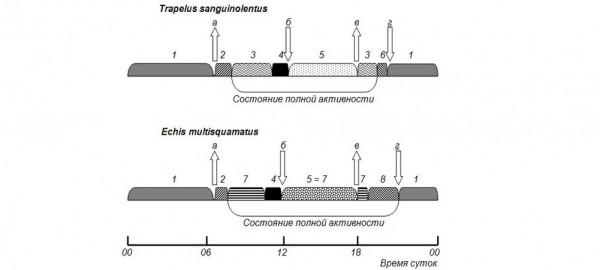
Рис. 1. Суточная последовательность поведенческих реакции, связанных с температурой, у разных видов рептилий: а – выход на поверхность утром; б – уход с поверхности днем; в – выход на поверхность вечером; г – уход с поверхности вечером; 1 – ночной покой; 2 – нагревание; 3 – термостабилизирующее поведение; 4 – добровольный перегрев; 5 – дневной отдых; 6 – остывание; 7 – терминг; 8 – термонейтральное поведение
Fig. 1. The daily sequence of behavioral reactions connected with the temperature in different reptile species: а – travel to the surface in the morning; б – leaving the surface at midday; в - travel to the surface in the evening; г – leaving the surface in the evening; 1 – night rest; 2 – heating; 3 – thermal stabilizing behavior; 4 – voluntary overheating; 5 – day rest; 6 – cooling; 7 – therming; 8 – thermal neutral behavior
Термобиологические показатели рептилий, их изменчивость и стабильность
Говоря о термобиологических показателях, необходимо обратить внимание на очень важное обстоятельство. Существуют физиологически обусловленные термобиологические показатели, а есть те диапазоны температур тела в рамках физиологически стабильных границ, которые рептилии имеют возможность выбрать и проявить лишь в конкретных климатических условиях среды. Если внешние условия не позволяют рептилиям проявить весь потенциальный диапазон своих физиологических возможностей, то можно наблюдать заметные различия в результатах исследований одного и того же вида, например, в лаборатории и в природе, в разных биотопах в естественных условиях. Но это будут не физиологически, а лишь экологически, экзогенно обусловленные различия. Поэтому мы разделили термобиологические показатели рептилий на термофизиологические и термоэкологические (Черлин, Музыченко, 1988).
Термофизиологические показатели описывают полные потенциальные предпочтения и границы, а термоэкологические показатели чаще всего демонстрируют лишь более узкую часть термофизиологического диапазона, которая имеет возможность проявиться в данных климатических и экологических условиях. Смысл и происхождение этих искажений проиллюстрированы на рис. 2.
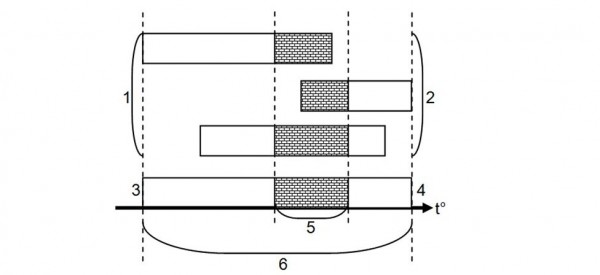
Рис. 2. Термобиологические показатели рептилий одного и того же вида, корректно полученные в различных полевых условиях и в лаборатории: 1 – в полевых условиях, 2 – термоэкологические показатели, 3 – в лаборатории, 4 – термофизиологические показатели, 5 – диапазон термостабилизации, 6 – диапазон полной активности
Fig. 2. Thermobiological features of the same species of reptiles in different field and laboratory conditions. 1 – field conditions, 2 – thermoecological indicators, 3 – in laboratory, 4 – thermophysiological indicators, 5 – the thermostabilization range, 6 – full activity range
Основные термофизиологические показатели, которые имеет смысл в данном контексте рассматривать: диапазон температур полной активности ограничивается минимальной и максимальной температурами полной активности; диапазон термостабилизации (довольно узкий диапазон высоких температур тела, который рептилии активно, направленно и оперативно поддерживают у себя в течение более или менее продолжительного времени); критические минимум и максимум температуры тела (температуры тела при наступлении холодового и теплового шоков); температуры ночного покоя (температуры тела, которые рептилии активно и направленно поддерживают у себя в состоянии ночного покоя и которые необходимы им для поддержания суточного градиента температуры тела); диапазон суточных колебаний температуры тела – диапазон температур между максимальной температурой тела при полной активности и минимальной температурой ночного покоя; «запас температурной прочности вида» – диапазон температур тела между максимальной температурой полной активности и критическим максимумом (Черлин, 2014).
Основные термоэкологические показатели, которые имеет смысл в данном контексте рассматривать: диапазон добровольных температур тела – диапазон температур тела, зарегистрированный у рептилий данного вида, находящихся в состоянии полной активности в данных экологических условиях (аналог диапазона полной активности в разделе термофизиологических показателей), минимальные и максимальные добровольные температуры тела ограничивают диапазон температур полной активности в данных экологических условиях (аналоги минимальной и максимальной температур полной активности в разделе термофизиологических показателей), предпочитаемые температуры – диапазон температур тела при полной активности, который рептилии направленно избирают и поддерживают у себя в данных экологических условиях (аналог диапазона термостабилизации в разделе термофизиологических показателей), ночные температуры – температуры тела в период ночного покоя, характерные для данных экологических условий (аналог температур ночного покоя в разделе термофизиологических показателей), суточный перепад температур – разница между предпочитаемой и средней ночной температурами тела; этот показатель очень важен для жизнедеятельности рептилий, поскольку он определяет возможность протекания у них ряда важнейших физиологических процессов (аналог диапазона суточных колебаний температуры тела в разделе термофизиологических показателей).
Поскольку термофизиологические и термоэкологические показатели характеризуют разные (эндогенные и экзогенные) стороны процесса терморегуляции, их варьирование у рептилий одного вида связано с разными причинами.
Полевые и лабораторные авторские исследования восьми видов ящериц в разных регионах и в разные сезоны, а также анализ литературных материалов показали следующие результаты (Черлин, 2014):
1) термоэкологические показатели у рептилий одного вида из разных популяций или из одной популяции, но в разные сезоны года или в разные годы могут заметно различаться, поскольку являются результатами взаимодействия термофизиологических характеристик с очень изменчивыми ландшафтными, климатическими, биотопическими и другими экологическими условиями-ограничениями;
2) термофизиологические показатели, связанные с регуляцией температуры тела в неактивном состоянии (температуры тела неактивного состояния) и в состоянии ночного покоя (или дневного покоя у ночных видов), различаются в разных популяциях одного вида, географически и сезонно. Они проявляют циркадную и сезонную эндогенные ритмики и, скорее всего, имеют, кроме прочего, определенную экологическую привязку;
3) термофизиологические показатели, связанные с биохимическими основаниями, – показатели термальной выносливости (критические минимальная и максимальная температуры, а также температурная выносливость тканей) – могут быть стабильными, могут немного различаться в разных популяциях одного вида, но экологическое значение этих различий неопределенно и неоднозначно, на них влияют некоторые особые физиологические состояния: беременность, возраст и т. п.; эта изменчивость требует дальнейшего изучения;
4) на изменчивость различных термофизиологических показателей (температурные предпочтения, температурная выносливость) также влияют некоторые особые физиологические состояния: голод, беременность, возраст, стресс, акклимация, вирусные заболевания и т. п.;
5) термофизиологические показатели, являющиеся базовыми, «штатными» характеристиками, закрепленными в системе терморегуляции как терморегулирующие константы в период полной активности (минимальная и максимальная температуры полной активности, диапазоны термостабилизации), демонстрируют внутривидовую стабильность. У них нет значимой географической, сезонной или пр. изменчивости.
Таким образом, можно констатировать очень важное обстоятельство: у рептилий имеется видоспецифичный комплекс базовых, стабильных, термофизиологических характеристик, детерминированных, прежде всего, структурой механизма регуляции температуры тела в состоянии полной активности (минимальная и максимальная температуры полной активности, диапазон термостабилизации). Этот комплекс не подвержен внутривидовой (географической, популяционной и сезонной) изменчивости и формирует видоспецифичную, стабильную, физиолого-экологическую матрицу (Cherlin, 1991; Черлин, 2012а, 2014). По сути физиолого-экологическая матрица – это описание видоспецифичного комплекса характеристик термального гомеостаза и основных механизмов его поддержания в различных условиях среды.
Видоспецифичная физиолого-экологическая матрица обуславливает связь жизнедеятельности данного вида рептилий с климатическими условиями среды. Она является основой, ключевым элементом механизма адаптации рептилий к условиям среды.
Неоднозначность связи термобиологии рептилий и условий среды
Какое влияние оказывают описанные выше особенности терморегуляции и термобиологические характеристики рептилий на процессы их термальной адаптации?
Логично было бы предположить, что термофизиологические показатели рептилий играют исключительно важную роль в их адаптации к различным климатическим условиям. Но связь термобиологических показателей рептилий с условиями среды оказывается неожиданно неоднозначной…
Как уже упоминалось ранее, диалектика связи температуры тела с термальными факторами среды весьма неоднозначная: благодаря активному использованию и развитию специфических комплексов физиологических и поведенческих регуляторных реакций рептилии стремятся одновременно усилить оперативную связь температуры тела с микроклиматическими термальными факторами среды, но за счет этого одновременно ослабить зависимость своей жизнедеятельности от макроклиматических условий мест обитания. Эта закономерность может быть выражена и другими словами: за счет сильного развития оперативной связи температуры тела с микроклиматическими параметрами среды рептилии получают возможность адаптироваться к широкому спектру макроклиматических условий географических регионов обитания.
В качестве примера можно рассмотреть герпетофауну западных Кызылкумов. В ее составе имеются следующие основные виды: песчаная круглоголовка Phrynocephalus interscapularis, ушастая круглоголовка Phrynocephalus mystaceus, степная агама Trapelus sanguinolentus, серый варан Varanus griseus, сетчатая ящурка Eremias grammica, быстрая ящурка Eremias velox, линейчатая ящурка Eremias lineolata, средняя ящурка Eremias intermedia, серый геккон Cyrtopodion russowi, каспийский геккон Cyrtopodion caspius, сцинковый геккон Teratoscincus scincus, гребнепалый геккон Crossobamon eversmanni, пустынный гологлаз Ablepharus deserti, стрела-змея Psammophis lineolatus, чешуелобый полоз Spalerosophis diadema, гюрза Чернова Macrovipera lebetina černovi и степная черепаха Agrionemys horsfieldi.
Даже беглого взгляда на табл. 1 достаточно, чтобы заметить значительные различия в термобиологических показателях у этих видов. За счет чего в таких жестких пустынных условиях это становится возможным?
Здесь необходимо обратить внимание на ряд важных обстоятельств. Во-первых, на одной и той же площади существует значительное разнообразие основных биотопов: дневная пустыня с ячеистыми песками; ночная пустыня; обрывы коренного берега Амударьи днем; то же ночью; тугайный лес днем; то же ночью и т. п. Все эти биотопы обладают специфическими климатическими условиями по всем показателям: температурно-влажностному режиму, освещенности и т. п.
Таблица 1. Термобиологические показатели некоторых рептилий западных Кызылкумов
| Виды | Mint°
полной активности |
Max t°
полной активности |
Диапазон термостабилизации | Источник |
| Phrynocephalus interscapularis | 37.5 | 45.0 | 40.5–41.5 | Черлин, Музыченко, 1988 |
| Ph. mystaceus | 36.0 | 44.2 | 38.8–39.8 | Черлин, Музыченко, 1988 |
| Trapelus
sanguinolentus |
37.0 | 44.5 | 38.0–42.0 | Черлин, 1988б |
| Varanus griseus | 34.0 | 40.6 | 36.0–39.0 | Целлариус и др., 1991 |
| Eremias grammica | 34.0 | 42.5 | 38.6–39.5 | Черлин, Музыченко, 1988 |
| E. velox | 36.0 | 41.0 | 39.5 | Черлин, Чикин, 1991 |
| E. lineolata | 34.0 | 41.0 | 35.0–37.5 | Черлин, 2012а |
| Psammophis
lineolatus |
32.0 | 40.4 | 34.0–40.0 | Черлин, 1991; Черлин, 2013в |
| Cyrtopodion
russowi |
11.0 | 42.5 | 37.0–40.0 | Черлин, 1988б |
| E. intermedia | 32.0 | 39.0 | 32.5–37.5 | Черлин, 2012а |
| Agrionemys
horsfieldi |
27.0 | 36.5 | 32.0–34.0 | Черлин, 2012а |
| Spalerosophis
diadema |
28.0 | 34.5 | 28.8–32.4 | Черлин, 1991; Черлин, 2013в |
| Ablepharus deserti | 25.0 | 33.0 | 27.0–31.0 | Черлин, 2012а |
| Cyrtopodion
caspius |
23.0 | 36.0 | 26.0 –32.0 | Черлин, 2012а |
| Teratoscincus scincus | 16.0 | 33.0 | 27.5–33.0 | Черлин и др., 1983; Черлин, 2013а |
| Crossobamon eversmanni | 18.0 | 32.0 | 28.0–32.0 | Черлин, 2013а |
| Macrovipera
lebetina černovi |
17.0 | 34.0 | 27.0–30.0 | Черлин, Шепилов, 2014; Cherlin, Shepilov, 2014 |
Во-вторых, между разными видами рептилий существуют порой весьма значительные различия в массе тела (от 1 до 3 порядков) и других особенностях, очень сильно отражающихся на процессах теплообмена со средой.
В-третьих, огромное нивелирующее влияние на связь температур тела и среды оказывают терморегуляционное и другие формы поведения (способность или неспособность залезать на ветки, активность использования нор и т. п.).
В-четвертых, сами термобиологические показатели активности, как выяснилось, не всегда напрямую связаны с климатом района обитания (Черлин, 1989). Так, в одних и тех же биотопах в одно и то же время в Каракумах и Кызылкумах живут ушастая и песчаная круглоголовки, которые имеют немного различные модальные диапазоны термостабилизации: соответственно 38.8–39.8° и 40.5–41.5° (Черлин, Музыченко, 1988). Критические максимумы температуры тела при этом у данных видов примерно одинаковые (48.0–50.0°). У серого геккона, живущего на стволах деревьев и не уходящего в убежища даже в самое жаркое время суток, критический максимум 43.5–44.0°, а у сцинкового геккона, обитающего там же, но встречающегося на поверхности только ночью, этот показатель 41.0–44.0° (Черлин, Музыченко, 1984), то есть эти цифры вполне сопоставимы. Но диапазоны термостабилизации двух этих видов соответственно 36–40° (Черлин, 1983 б) и 27.5–33° (Черлин и др., 1983), то есть значительно различаются. При сравнении ушастых круглоголовок и сетчатых ящурок видно, что их диапазоны термостабилизации очень сходны (38.8–39.8°), а критические максимумы различны (соответственно 49.0–49.5° и 44.0–45.0°) (Черлин, Музыченко, 1983, 1988). Все это приводит к тому, что в одном и том же месте могут обитать виды как очень высокотемпературные, так и типичные мезофилы. И действительно, почти половина видов (кроме, очевидно, группы наиболее термофильных), обладая своими показателями активности, могли бы обитать и в значительно более северных районах с более холодным и мягким климатом, включая степи и даже широколиственные леса. Но их там нет. Нетрудно такую же ситуацию экстраполировать и на другие районы, и на другой набор видов рептилий.
Исходя из этого можно сделать первый вывод: в одном и том же месте даже в достаточно экстремальных аридных условиях вполне благополучно живут рептилии с очень разными термобиологическими показателями, описывающими полную активность.
Принадлежность к группам термонейтрально или термостабильно активных рептилий также мало помогает в понимании специфики адаптации к разным климатическим зонам. Рептилии той или другой групп вполне успешно приспосабливаются к самому широкому спектру природно-климатических условий. Так, даже в жарких пустынях можно встретить представителей тех или других пресмыкающихся. В приведенном выше примере с герпетофауной Кызылкумов все агамиды, лацертиды и сцинциды относятся к термостабильно активным, а геккониды и змеи – к термонейтрально активным рептилиям. В таежной зоне, где климат холодный, также обитают, с одной стороны, термостабильно активные живородящие (Zootoca vivipara) и прыткие (Lacerta agilis) ящерицы, веретеница (Anguis fragilis), а с другой – термонейтрально активные обыкновенные гадюки (Pelias berus) и обыкновенный уж (Natrix natrix). И в пустыне, и в тайге обе группы пресмыкающихся вполне успешно приспосабливаются к природно-климатическим условиям, используя микробиотопическое и микроклиматическое разнообразие и формируя различные пространственно-временные структуры активности. Следовательно, и эти достаточно глубокие физиологические различия также не являются определяющими ни в адаптации к различным условиям среды, ни в географическом распространении пресмыкающихся.
Но можно сделать и еще одно чрезвычайно интересное сравнение. В табл. 2 приведены термофизиологические показатели ряда видов змей и ящериц. В строчках без заливки помещены мезофильные виды, обитающие в жарких пустынных регионах, а в залитых светло-серым цветом – обитатели лесных, таежных и даже приполярных регионов. В этой связи поразительно то, что подавляющая часть термофизиологических показателей, связанных с проявлением полной активности, у обеих групп почти одинаковая! Существенно различаются минимальные температуры полной активности (частично это может быть связано не только с физиологическими различиями, но и с методическими некорректностями). Стабильно более низкими в подавляющую часть года оказываются и температуры ночного покоя. Максимальные же температуры полной активности и диапазоны термостабилизации у обеих групп очень сходны.
Таблица 2. Термобиологические показатели некоторых видов змей и ящериц западных Кызылкумов и северных регионов
| Виды | Min t°
полной актив- ности |
Max t°
полной активнос- ти |
Диапазон термоста- билизации | Температуры ночного покоя | Источник |
| Macrovipera
lebetina černovi |
17.0 | 34.0 | 27.0–30.0 | 10–32 в разные сезоны | Черлин, Шепилов, 2014; Cherlin, Shepilov, 2014 |
| Echis multisquamatus | 25.0 | 34.0 | 30–32 | 10–32 в разные сезоны | Черлин, Целлариус, 1981 |
| Spalerosophis diadema | 28.0 | 34.5 | 28.8–32.4 | 10–32 в разные сезоны | Черлин, 1991; Черлин, 2013в |
| Pelias berus | ̴ 30 | Leeuwen, 2009 | |||
| -«- | 32–33 | 10–20 | Lourdais et al., 2013 | ||
| -«- | 9 | 31 | Чан-Кьен, 1967 | ||
| -«- | 13–14 | 32–34 | Юмашев, 1995 | ||
| -«- | 5.6 | 33.2 | Литвинов, 2004 | ||
| -«- | 34 | 32–34 | Saint-Grons, 1980 | ||
| -«- | 9 | 34 | ~32 | Коросов, 2010 | |
| Ablepharus deserti | 25.0 | 33.0 | 27.0–31.0 | 15–25 | Черлин, 2012а |
| Teratoscincus scincus | 16.0 | 33.0 | 27.5–33.0 | 10–32 в разные сезоны | Черлин и др., 1983; Черлин, 2013а |
| Crossobamon eversmanni | 18.0 | 32.0 | 28.0–32.0 | 10–32 в разные сезоны | Черлин, 2013а |
| Zootoca vivipara | 26.8 | 33.0 | 28.8–32.5 | 10–15 | Черлин, 2014 |
| Lacerta agilis | 35.2–35.8 | 10–15 | Amat et al., 2003 |
Различаются и суммарные количества теплоты (STe), получаемой разными видами рептилий за год: несмотря на то, что диапазоны полной активности у обеих групп сходны, но продолжительность периода высоких температур и уровни температур ночного покоя у пустынных видов существенно больше (см. табл. 3).
Таблица 3. Суммы эффективных температур (STe) для некоторых видов рептилий за год (средние по нескольким регионам). (По: Черлин, 2014)
| Вид рептилий | Min | Max | Среднее | %% | |
| За весь год | |||||
| Pelias berus | 45960 | 85137 | 69458 | 100 | 43.8 |
| Echis multisquamatus | 179697 | 180969 | 180213 | 323.8 | 113.6 |
| Macrovipera lebetina černovi | - | - | 158655 | 285.1 | 100 |
| Montivipera raddei | - | - | 160234 | 288.0 | 101.0 |
| Тропические удавы (Boidae) | - | - | 240900 | 346.8 | 151.8 |
| Только за период активности (без зимовки) | |||||
| Pelias berus | 34392 | 71052 | 55649 | 100 | 46.6 |
| Echis multisquamatus | 148041 | 156033 | 151245 | 271.8 | 126.7 |
| Macrovipera lebetina černovi | - | - | 119415 | 214.6 | 100 |
| Montivipera raddei | - | - | 128290 | 230.5 | 107.4 |
| Тропические удавы (Boidae) | - | - | 240900 | 432.9 | 201.7 |
Исходя из сказанного, можно сделать следующие выводы.
Вывод второй: в контрастно разных климатических условиях могут успешно обитать рептилии, обладающие сходными термобиологическими показателями, описывающими полную активность.
Вывод третий: сами по себе термобиологические и другие показатели, описывающие полную активность, не являются специфическими адаптациями к климату данного региона.
Закономерности формирования пространственно-временной структуры активности, в свою очередь, базируются также и на видоспецифичных физиологических, морфологических, поведенческих и других генетически фиксированных характеристиках. У близкородственных групп эти закономерности и многие характеристики часто бывают сходными (Черлин, 1989; и др.). Значит, если сравнивать близкородственные виды со сходными закономерностями, но с несколько различающимися абсолютными значениями указанных характеристик, то можно понять и объяснить разницу в их пространственно-временном, а также частично в биотопическом и географическом размещении (Черлин, 1989). Так, на адырах Нуратинского хребта (Узбекистан) в полосе шириной всего около 1 км встречаются совместно степная и туркестанская агамы. Температуры полной активности и диапазоны термостабилизации у степных агам существенно выше, чем у туркестанских (табл. 4).
Таблица 4. Термобиологические показатели трех видов агам
| Виды | Температуры полной активности, °С | Диапазон термостабилизации, °C | |
| min | max | ||
| Trapelus sanguinolentus | 37.0 | 44.5 | 38.0–42.0 |
| Laudakia lehmanni | 35.0 | 39.0 | 35.0–37.5; Mo = 37.3 |
| Laudakia chernovi | 32.0 | 37.0 | 32.0–34.0; Mo = 33.6 |
В соответствии с этим степная агама населяет более жаркие остепненные открытые пространства низин около хребта, а туркестанская поднимается по ущельям и склонам в горы, где температуры среды более изменчивы, контрастны и к тому же их основной фон заметно ниже (Черлин, Чикин, 1991; Cherlin, 1989).
Подобное же сравнение возможно и для туркестанской агамы и агамы Чернова, с которыми мы работали на хребте Кугитанг в Туркмении. Агамы Чернова выбирают заметно более низкие температуры тела и поэтому поднимаются намного выше в горы (более 2000 м над у. м.), чем туркестанские агамы (ниже 2000 м над у. м.). Агамы Чернова оказываются способны выдерживать большие перепады температур, для них характерно псевдо ТСП, когда они подолгу находятся на поверхности без солнца при достаточно низких температурах, располагаются на гребнях каменных выходов, чтобы лучше использовать ненадолго появляющееся солнце, и ожидают коротких периодов солнечной погоды для нагрева тела до температуры полной активности. Ниже по склонам, где живут туркестанские агамы, им будет явно жарко; при постоянно высокой температуре воздуха и горячих камнях им грозит там регулярный перегрев.
Можно рассматривать целые герпетокомплексы на одной территории, внутри которых у разных видов различны терморегуляторные стратегии и разнообразны пространственно-временные структуры активности. Один пример такого рода герпетофауна Кызылкумов уже приведен в табл. 1.
Другой пример – герпетофауна горных ущелий Нуратинского хребта, где можно встретить набор фоновых видов: степная и туркестанская агамы, быстрая ящурка, туркестанский геккон Cyrtopodion fedtschenkoi и желтопузик Pseudopus apodus (Черлин, Чикин, 1991; Cherlin, 1989). Их термобиологические показатели приведены в табл. 5.
Таблица 5. Термобиологические показатели некоторых горных рептилий хребта Нурата
| Виды | Температура полной активности, °С | Диапазон термостабилизации, °C | Температуры ночного покоя, °С | ||
| min | max | Весна | Лето | ||
| Trapelus sanguinolentus | 37.0 | 44.5 | 38.0–42.0 | ||
| Laudakia lehmanni | 35.0 | 39.0 | 35.5–37.5; Mo = 37.3 | 18.0-20.0 | 25.0-27.0 |
| Cyrtopodion fedtshenkoi | 25.0 | 36.0 | 32.0–34.5; Mo = 33.3 | 18.0-20.0 | 25.0-27.0 |
| Eremias velox | 36.0 | 41.0 | 39.0–41.0; Mo = 39.5 | 17.0-19.0 | 25.0-27.0 |
| Pseudopus apodus | 25.0? | 30.0 | 27.0–30.0; Mo = 28.0–29.0 | 17.0-19.0 | 20.0-22.0 |
Хорошо прогреваемые участки дна ущелий занимали в основном быстрые ящурки и степные агамы, имеющие самые высокие температуры полной активности и модальные диапазоны термостабилизации. Туркестанские агамы, способные свободно лазать по скалам и вертикальным поверхностям и имеющие немного более низкие температурные показатели активности, занимали в основном вертикальные склоны и отвесные поверхности камней и скал, тянущиеся намного выше в горы, которые хорошо прогревались солнцем, но не так длительно и регулярно, как дно ущелий внизу. А еще менее теплолюбивые туркестанские гекконы, которые мельче и легче агам, способные свободно передвигаться по вертикальным каменным поверхностям и потолку небольших пещер и пустот в скалах, ограниченно использовали открытые солнечные участки скал, в основном получая тепло от разогретых камней на склонах (как снаружи, так и внутри полостей, пустот и пещер). Желтопузики же, крупные, не умеющие лазать и ограниченно подвижные, с низким температурным предпочтением, могли населять только поймы саев и пологие глинисто-щебнистые поросшие травой склоны. Часто они держались недалеко от густых зарослей мяты и другой травы, растущей около родников. В качестве убежищ желтопузики использовали густые заросли травы, трещины скал и пустоты под камнями. Таким образом, распределение пресмыкающихся по стациям вполне соответствовало их термобиологическим показателям.
В результате оказывается, что в одних и тех же местах могут жить рептилии с очень разными термобиологическими характеристиками.
Из сказанного выше можно сделать четвертый вывод: если ориентироваться только на термобиологические характеристики рептилий, описывающие показатели их полной активности, то невозможно объяснить закономерности их приуроченности к различным природно-климатическим зонам.
Это показывает, что причинно-следственные связи между термофизиологией и природными условиями обитания у рептилий неоднозначны и многоплановы. Между ними существует важный посредник – крайне эффективный комплекс в основном поведенческой терморегуляции, который чутко меняет только пространственно-временную структуру активности. А это, в свою очередь, очень сильно нивелирует влияния условий внешней среды на термофизиологию рептилий, ослабляет давление на нее отбора, противодействует накоплению адаптивно значимых изменений комплекса термофизиологических характеристик в генофонде популяций.
Все приведенные выше материалы говорят о том, что связь между термофизиологическими показателями рептилий, описывающими условия реализации полной активности, и характеристиками природных условий среды – не всегда однозначна. Комплекс в основном поведенческой и физиологической терморегуляции существенно нарушает (размывает) однозначность связи между температурами среды и тела. Благодаря терморегуляции (в основном поведенческой) рептилии как группа, имея потребность в нагреве тела хотя бы на недолгое время до уровней примерно от 28 до 42° в зависимости от видов, могут обитать как в экстремально жарких пустынях, так и в не менее экстремальных для их физиологии приполярных районах и высокогорьях, т. е. в регионах с широким спектром географических и климатических условий. Таким образом, термофизиологические характеристики, описывающие условия протекания у рептилий полной активности, часто могут не очень соответствовать характеристикам географических регионов обитания, т. е., например, мезофилы могут успешно обитать в регионах с жарким климатом, а в холодных климатах могут жить рептилии, которым необходим нагрев тела до 30–34˚. Таким образом, термофизиологические характеристики, описывающие условия протекания у рептилий полной активности, могут и не являться для них специфически адаптивными к условиям данных природно-климатических зон.
Фактор количества теплоты в экологии рептилий
Хотелось бы особо остановиться на такой важной характеристике, как «эффективные температуры» (STe), или «дни-градусы», как ее называли раньше. Чаще всего такие исследования проводили на насекомых (Иванов, 1959), особенно при изучении продолжительности отдельных стадий развития (Parker, 1930; и др.), но были и публикации, касающиеся рыб (Кашкаров, 1938), земноводных (Наумов, 1963). Были, конечно, и более поздние публикации. В англоязычной литературе этот показатель называют «градусо-днями» – degrees days (DD), или «тепловыми единицами» – heat units (Begon et al., 1986; Valenzuela et al., 1997; Baskerwille, Emin, 1969; Whillams, Crossman, 1977; Parker, Andrews, 2006). Думаю, такой подход весьма продуктивен и для пресмыкающихся. Так, на примере черепахи тартаруги Podocnemis expansa показана высокая негативная корреляция (-0.85) между количеством градусо-дней и продолжительностью инкубации яиц (Valenzuela et al., 1997), а исследования восточной заборной игуаны Sceloporus undulatus – значение количества дней-градусов для завершения эмбриогенеза и, таким образом, определение северной границы распространения (Parker, Andrews, 2006). Этот показатель успешно используется для научного исследования и практической деятельности по сохранению морских черепах (Matsuzawa et al., 2004; и др.).
Обладая вполне определенным набором стабильных видоспецифичных морфологических, физиологических и поведенческих характеристик, рептилии данного вида в условиях определенного региона с определенными климатическими характеристиками могут иметь единственно возможный вариант пространственно-временно́й структуры суточной и сезонной активности. Следовательно, рептилии данного вида в определенных природно-климатических условиях могут набрать лишь вполне определенную, конечную величину STe. Нами разработан скорректированный метод для расчета годовой STe именно для рептилий и произведены расчеты годовых сумм STe для одних и тех же видов в разных частях их ареалов и разных видов, обитающих в различных природных зонах (Черлин, 2011, 2014).
Внутри достаточно жесткой пространственно-временно́й структуры активности рептилии данного вида имеют возможность некоторой модификации STe за счет выбора различных ночных температур и доли терминга в сутках. Можно посмотреть на данный вопрос и с другой стороны: при обитании в разных природно-климатических зонах величина STe, определяющая возможности существования жизнеспособной популяции, изменяется в основном за счет объективно складывающейся пространственно-временно́й структуры активности, субъективных модификаций уровней ночных температур и продолжительности терминга.
Расчеты STe проводились в основном на разных видах змей (Черлин, 2012а, 2014), биологию которых мы изучали как в природе, так и лаборатории (среднеазиатские эфы Echis multisquamatus, обыкновенные гадюки Pelias berus, гюрза Macrovipera lebetina, и др.). Из термобиологических исследований очевидно, что практически у всех этих змей температуры термостабилизации находятся в пределах 28–32°, а диапазоны температур полной активности редко превышают 32–33°. Для каждого из этих видов змей мы производили расчеты STe для разных точек их ареалов, в которых климатические условия существенно различаются. Сравнение этих цифр показывает, что основная разница между ними сводится к двум моментам: 1) у более теплолюбивых видов ночные температуры выше и 2) у более теплолюбивых видов продолжительность периода с высокими ночными температурами в году намного больше. Таким образом, несмотря на то, что температуры тела при активности у них оказываются сходными, годовая величина STe для этих видов довольно сильно различается – для змей, населяющих более теплые регионы, она более чем в 3 раза превышает эту величину для тех, которые населяют холодные регионы. Это значит, что для завершения одних и тех же важнейших физиологических сезонных процессов теплолюбивым видам надо значительно больше времени, чем холодолюбивым.
Очевидно, у каждого вида рептилий есть критическое значение STe, ниже которого реализация, в частности, репродуктивного цикла невозможна. Это обстоятельство будет являться серьезным препятствием к распространению животных данного вида в регионы с более холодным климатом. При этом ряд видов, обитающих в горах или в северных районах, не всегда могут набрать за год достаточную для завершения, например, полового цикла STe, и размножение у них происходит неежегодно, что продемонстрировано Нильсоном на примере обыкновенной гадюки (Nilson, 1981).
Таким образом, получается, что представители данного вида рептилий, как индивидуальные животные, могут существовать на значительно большем ареале, чем реально живет вид. И это подтверждает практика содержания рептилий в неволе. Однако серьезное ограничение на распространение видов оказывает невозможность завершения у них ряда важнейших физиологических сезонных циклов, необходимых для существования вида (в частности, размножения), вследствие, в том числе, и недостаточности STe.
Имея материалы исследований по термобиологии некоторого вида рептилий и зная определенные климатические характеристики разных районов, можно с большой степенью достоверности вычислить структуру суточной и сезонной активности этих животных и сумму эффективных температур для них в данных местах. А зная критический минимум суммы эффективных температур для данного вида, можно, например, определить, в каких районах этот вид сможет жить, а в каких – нет.
Обсуждение
В научной литературе часто употребляется словосочетание «термальные адаптации». Но, говоря о термальных адаптациях, авторы чаще всего описывают и обсуждают то, что составляет предмет термобиологии как таковой, термобиологические категории, эколого-физиологические, физиолого-экологические, физиологические, экологические и прочие аспекты термобиологии рептилий. Думаю, что здесь нужно разобраться в том, чем отличаются по смыслу понятия «термобиология» и «термобиологические особенности», с одной стороны, от понятия «термальные адаптации» – с другой. Часто в литературе они используются как синонимы, т. е. принимается точка зрения, что все термобиологические показатели, процессы и структуры являются однозначно адаптивными. Но нам кажется, что это не совсем одно и то же. По крайней мере, если это так, то такое положения надо доказать, а различия между двумя группами понятий нужно тогда определить и в дальнейшем термины применять правильно.
Итак, термобиология, термобиологические особенности. Что это? Это понятия статические, т. е. набор, комплекс эндогенных характеристик, которые описывают свойства животного, зависимые от температуры, параметры термального гомеостаза, внутреннюю организацию и механизмы терморегуляции. А вот термальные адаптации – это понятие функциональное, динамическое. Это морфологические свойства, биохимические, физиологические и поведенческие реакции, направленные на приспособление, улучшение приспособенности организма животного или сообщества к данным термальным условиям среды или на приспособление к новым, меняющимся условиям термальной среды (при изменении условий или при освоении новых биотопов, что тоже есть изменение условий). Выражаются они в сохранении или организации возможностей для успешного выживания особей и популяций в данных или меняющихся условиях среды при допустимых энергетических затратах на жизнеобеспечение особей и воспроизводство популяций. И все это – при непременном сохранение показателей термального гомеостаза.
В самом общем смысле адаптация (лат. adapto – приспособляю) – процесс приспособления к условиям внешней среды.
Индивидуальные адаптации – реакции индивидуального животного (в основном поведенческие), направленные на реализацию актов жизнеобеспечения его как самостоятельной особи, на выполнение им всех необходимых физиологических и экологических функций в популяции и биоценозе в данных или меняющихся условиях среды, на безусловное сохранение основных параметров гомеостаза, улучшение адаптированности особи в различных условиях среды и получение ею конкурентных преимуществ перед другими индивидами.
Адаптированное состояние особи – это такое равновесное, устойчивое, динамическое состояние, при котором отдельное, индивидуальное животное может обеспечить выживание себя как самостоятельной особи с наименьшими возможными для данных условий (оптимальными) энергетическими затратами при сохранении основных характеристик гомеостаза.
Что представляет собой реакция индивидуальной адаптации? Ее важнейший смысл и направленность – в любых условиях среды (постоянных или меняющихся) сохранять неизменным видоспецифичный комплекс базовых, стабильных, термофизиологических, терморегулирующих показателей (характеристик гомеостаза) – физиолого-экологическую матрицу. Реализуется это благодаря двум основным направлениям индивидуальных адаптивных реакций:
1) оперативно и максимально эффективно нивелировать (нейтрализовать) влияние «возмущающих» воздействий во внутренней или внешней среде, т. е. использовать в основном наиболее простые, энергетически малозатратные, поведенческие регуляционные реакции и комплекс поведенческих мер для изменения режима своей активности (суточной и сезонной пространственно-временно́й структуры активности) с целью реализации периодических физиологических процессов в организме и сохранения характеристик гомеостаза;
2) изменить функционирование различных биохимических, физиологических и прочих систем организма, чтобы их работа была наиболее оптимальной и энергетически наименее затратной в новых условиях; этот путь более сложный, энергетически более дорогой, чем первый.
Результаты исследований показывают, что, хотя все животные (по крайней мере высшие) задействуют при адаптациях оба этих направления, но очевидно, что чем успешнее используется первое, тем менее актуальным становится второе.
Основной же смысл адаптаций, связанный с сохранением неизменности характеристик физиолого-экологической матрицы, важен всегда и в любом случае. Поэтому комплекс характеристик термального гомеостаза физиолого-экологической матрицы нужно считать центральным, ключевым понятием в механизме процесса адаптации.
Поведенческие терморегуляционные реакции
Можно выделить несколько типов поведенческих терморегуляционных реакций:
1) поведенческие терморегуляционные реакции, направленные на регуляцию температуры тела рептилий, т. е. поддерживающие ее определенный уровень или способствующие реализации необходимой суточной динамики температуры тела при обязательном сохранении параметров термального гомеостаза;
2) реакции поведенческого реагирования на различные экологические (в том числе и термальные) факторы внешней среды и их изменения с целью «тонкой подстройки», «тюнинга» структуры жизнедеятельности для сохранения постоянного динамического равновесия между: а) необходимостью реализации всех важнейших элементов жизнедеятельности особи и популяции данного вида и б) необходимостью сохранения параметров гомеостаза. Этот «тюнинг» осуществляется за счет изменений в пространственно-временно́й структуре активности животных. В наборе этих поведенческих регулирующих реакций, характерных для видов, часто имеются общие реакции, т. е. отдельные элементы этого набора видоспецифичны;
3) реакции оперативного поведенческого реагирования на динамику факторов внешней среды с целью реализации необходимых физиологических суточных циклов (сон-бодрствование и т. п.). Эти реакции выступают как «тонкая подстройка», «тюнинг» пространственно-временно́й структуры суточной активности (Черлин, 2014);
4) реакции поведенческого реагирования на динамику факторов внешней среды с целью реализации необходимых физиологических сезонных циклов (цикл размножения, цикл питания, цикл жировых тел и т. п.). Эти реакции выступают как «тонкая подстройка», «тюнинг» пространственно-временно́й структуры сезонной активности (Черлин, 2014).
При этом «тюнинг» суточных и сезонных пространственно-временных структур активности осуществляется «через сито» базовых, неизменных, видоспецифичных, физиолого-экологических, терморегулирующих характеристик, т. е. видоспецифичной, стабильной физиолого-экологической матрицы (Cherlin, 1991; Черлин, 2014).
Значение поведенческих терморегуляционных реакций в комплексе индивидуальных адаптаций
О многочисленных формах и разнообразных механизмах поведенческой терморегуляции уже было подробно написано ранее (Черлин, 2012а, 2014). В контексте данного рассмотрения важен ответ на вопрос: являются ли поведенческие, терморегуляционные реакции адаптивными, т. е. направлены ли они а) на реализацию актов жизнеобеспечения отдельных особей с сохранением основных параметров гомеостаза, б) на выполнение ими всех необходимых физиологических и экологических функций в популяции и в биоценозе, в) на создание индивидуальных конкурентных преимуществ?
Поведенческие терморегуляционные реакции являются в рамках физиолого-экологической схемы терморегуляции оперативными поведенческими ответами обратной связи на отклонения переменной (температуры тела) от неизменного «штатного», терморегулирующего значения этой переменной, хранящегося в памяти системы в виде характеристик гомеостаза (физиолого-экологической матрицы). Эти ответные реакции направлены на достижение всех тех целей, которые указаны в предыдущем абзаце как характеристики адаптивного процесса. Следовательно, поведенческие терморегуляционные реакции, безусловно, могут считаться элементами индивидуальных адаптивных реакций.
Значение поведенческих механизмов формирования пространственно-временно́й структуры активности в комплексе индивидуальных адаптаций
В подавляющем большинстве случаев среда предоставляет для рептилий достаточно большие возможности, чтобы регулировать температуру тела. Этому способствует мозаичность температурных и других экологических условий среды, т. е. разнородность (контрастность) «термального поля» и других факторов экологической среды. Разные виды рептилий используют для терморегуляции эту разнородность условий в разных вариантах: поверхность почвы – ветки деревьев и кустов, поверхность почвы – норы, солнечные участки поверхности – теневые и т. п. Но сами рептилии остаются в зоне активности настолько долго, насколько им позволяют условия среды. Более того, в ряде случаев они пытаются всеми возможными способами продлить время активности при высоких температурах, что подтверждается наличием у некоторых видов рептилий такой формы активности, как добровольный перегрев (Черлин, 2014 и др.).
Термальные условия среды используются различными видами рептилий по-разному. В любом случае, важное условие при этом – сохранить видоспецифичный комплекс стабильных, термофизиологических характеристик гомеостаза.
Очень ярко демонстрирует связь режима активности с температурой тела и климатическими факторами среднеазиатская эфа Echis multisquamatus (рис. 3), с которой мы работали в течение четырех лет во впадине Еройландуз в Бадхызском заповеднике на самом юге Туркменистана (Черлин, Целлариус, 1981).
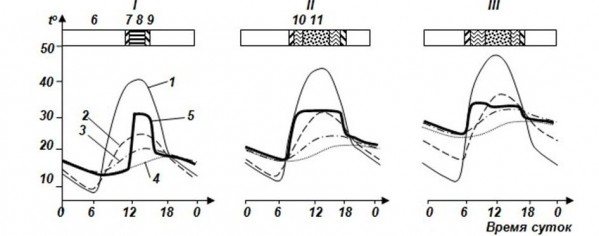
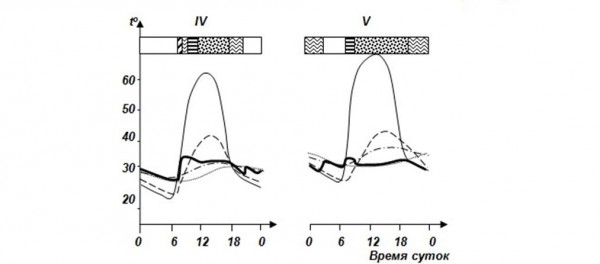
Рис. 3. Суточная и сезонная динамика температуры тела среднеазиатской эфы в зависимости от температур среды. I – начало весны, II – середина весны, III – конец весны, IV – начало лета, V – середина лета. По оси ординат (t°) – температура, °С, по оси абсцисс – время суток. 1 – температура поверхности почвы, 2 – температура воздуха на высоте 3 см, 3 – температура почвы на глубине 20 см, 4 – температура почвы на глубине 30 см, 5 – температура тела, 6 – ночной покой, 7 – нагревание, 8 – терминг, 9 – остывание, 10 – ТНП, 11 – дневной отдых = терминг (по Черлин, Целлариус, 1981)
Fig. 3. Daily and seasonal dynamics of body temperature in saw scaled viper Echis multisquamatus depending on environmental temperature. I – early spring, II – the middle of spring, III – late spring, IV – the beginning of summer, V – the middle of summer. Ordinate axis (t°) – temperature, °С, abscissa axis – day time. 1 – land surface temperature, 2 – air temperature at 3 cm height, 3 – soil temperature at 20 cm depth, 4 – soil temperature at 30 cm depth, 5 – body temperature, 6 – night rest, 7 – heating, 8 – therming, 9 – cooling, 10 – thermally neutral behavior, 11 – day rest = therming (by Cherlin, Cellarius, 1981)
Графики демонстрируют жесткую зависимость температуры тела в каждый промежуток времени от вполне определенных температурных характеристик среды. Это, в свою очередь, показывает, что такое животное, как эфа, обладающее вполне определенными термобиологическими характеристиками и особенностями терморегуляции, может быть активно только в данное время и в данном месте. Возможности для нее находиться в другом месте и быть активной в другое время, обладая своими характерными термобиологическими особенностями, потребностями и возможностями терморегуляции, просто нет. В противном случае температуры тела станут либо слишком низкими, либо слишком высокими, т. е. выйдут за пределы диапазона температур полной активности и также не смогут обеспечить физиологически необходимые суточную и сезонную динамики.
Еще один довольно жесткий пример подобной ситуации наблюдался нами в той же впадине Еройландуз, где мы работали не только с эфами, но и всеми другими рептилиями, которые там водятся (Целлариус, Черлин, Лукин, 1983). Летом температуры в норах (на глубине 20 см) вечером и в начале ночи повышались до своего суточного максимума – 32° и иногда даже немного выше (см. рис. 1V). Температуры тела эф в норах практически полностью совпадают с температурой почвы там. Уровень 32° – максимальная температура полной активности для эфы (как и для других скрытноживущих видов змей, обитающих там же, – афганских литоринхов Lytorhynchus ridgewayi, индийских бойг Boiga trigonatum, поперечнополосатых волкозубов Lycodon striatus). Поэтому в этот момент все змеи названных видов почти одновременно выходили на поверхность, где температуры в это время ниже. Другого варианта действий у них не было. Глядя на термометр нашей метеоплощадки, измерявший температуру почвы на глубине 20 см, мы точно знали: как только температура на нем поднимается до 32°, можно идти «собирать» все интересующие нас виды змей.
Однозначную зависимость структуры активности от температурных условий среды показывает серый варан, планомерное изучение которого мы вели в течение многих лет в Кызылкумском заповеднике в Узбекистане (Целлариус, Черлин, Меньшиков, 1991; Целлариус, Целлариус, 1997). Мы выяснили, что суточную динамику температуры тела этого животного в природной среде можно рассчитать, зная суточный ход температур поверхности почвы и приземного слоя воздуха на специальной метеоплощадке. Средняя температура тела варанов в состоянии полной активности в данной популяции в любой произвольно выбранный момент времени соответствовала величине:
tр = (tп + tв)/2,
где tр – средняя рассчитанная температура тела варанов в некий момент времени, tп и tв – температуры поверхности песка и приземного слоя воздуха на высоте примерно 5 см на метеоплощадке в этот же момент.
На рис. 4 видно, как однозначно определяется возможное время полной активности этих ящериц: они выходят из нор и ведут полную активность строго в такое время, когда температурные условия среды позволяют температуре их тела находиться в диапазоне полной активности.
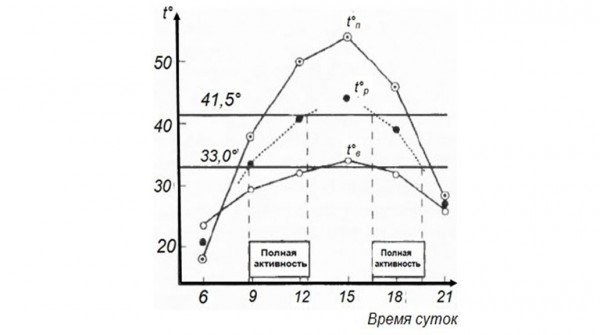
Рис. 4. Схема расчета продолжительности периода полной активности серого варана по метеоданным (по Целлариус, Целлариус, 1997). По оси ординат (t°) – температура, °С, по оси абсцисс – время суток. tп – температура поверхности песка, tв – температура приземного слоя воздуха, tр – рассчитанная температура тела
Fig. 4. The calculation of the full activity period in Varanus griseus using the meteorological data (by Cellarius, Cellarius, 1997). Ordinate axis (t°) – temperature, °С, abscissa axis – day time. tп – sand surface temperature, tв – surface air temperature, tр – calculated body temperature
Мы изучали термобиологию дальневосточного сцинка Plestiodon latiscutatus в 1983 году на острове Кунашир (Боркин и др., 2005). Наши исследования показали, что именно определенные и стабильные термофизиологические характеристики этих ящериц (температура полной активности 29.0-36.5°, диапазон термостабилизации 33.0-36.0°), их морфологическая структура (массивное тело, короткие лапки) и поведенческие особенности жестко определяют пространственно-временную структуру их активности, а также биотопическое распространение и даже географическое распространение (Боркин и др., 2005).
Мы провели сравнение экологии и термобиологии полосатой ящурки Eremias scripta и песчаной круглоголовки Phrynocephalus interscapularis в Каракумах (Черлин, 1988б; Черлин, Музыченко, 1983) при их совместном обитании (рис. 5). Оба вида – мелкие, подвижные, псаммофильные ящерицы, тяготеющие к высоким температурам. Но полосатые ящурки прекрасно лазают по веткам кустарников и деревьев, буквально перелетая с одной на другую. Лапки расположены так, что при движении по почве тело оказывается прижатым или очень близко к ней, вследствие чего их крохотное туловище с малой массой и минимальной термальной инерционностью очень быстро приобретает ту же температуру, что и поверхность почвы. Температуры полной активности и модальный диапазон термостабилизации достаточно высоки и близки к 40°. У песчаной круглоголовки лапки расположены так, что ящерицы могут легко приподниматься на них, удерживая тело приподнятым и отделенным от поверхности почвы. Температуры полной активности и диапазон термостабилизации около 40° и даже несколько выше. Это исследование продемонстрировало, что температурные условия среды вместе с особенностями терморегуляции жестко определяют единственно возможный вариант пространственно-временной структуры активности для обоих этих видов.
Рис. 5. Динамика температур среды и тела ящериц в Каракумах в летний период: 1 – полосатая ящурка; II – песчаная круглоголовка. Общие формы поведения ящериц: 1 – ночной отдых, 2 – нагревание, 3 – the thermal ТСП, 4 – остывание, 5 – добровольный перегрев, 6 – дневной отдых. Температуры: 7 – тела ящериц; 8 – поверхности почвы на открытом месте; 9 – воздуха в приземном слое (на высоте 2 см) на открытом месте; 10 – воздуха в кроне дерева на высоте 2 м; 11 – поверхности почвы в тени кустов; 12 – почвы на глубине 5 см; 13 – почвы на глубине 10 см; 14 – почвы на глубине 15 см
Fig. 5. The dynamics of lizards' body and ambient temperature in Karakum desert in summer. 1 – Eremias scripta; II – Phrynocephalus interscapularis. The activity patterns in lizards: 1 – night rest, 2 – heating, 3 – thermally stabilizing behavior, 4 – cooling, 5 – voluntary overheating, 6 – day rest. Temperatures: 7 – lizards’ body temperature; 8 – land surface temperature at an open place; 9 – surface air temperature at 2 cm height at an open place; 10 – air temperature in the tree crown at 2 m height; 11 – land surface temperature in the bush shade; 12 – soil temperature at 5 cm depth; 13 – soil temperature at 10 cm depth; 14 – soil temperature at 15 cm depth
Нами были проделаны специальные работы по сравнению термобиологических особенностей активности нескольких обитающих совместно видов рептилий: средней Eremias intermedia и линейчатой E. lineolata ящурок в Кызылкумах (Черлин, 2014), сцинковых и гребнепалых гекконов в Каракумах и Кызылкумах (Черлин, 2013а), трех видов среднеазиатских змей: среднеазиатской эфы, чешуелобого полоза и стрелы-змеи (Черлин, 2013б). Результаты всех работ однозначно показали, что именно термобиологические характеристики видов, их морфологические, физиологические и поведенческие особенности, а также разнородность «термального поля» в пустыне определяют механизмы их пространственно-временного и биотопического размещения, сезонное изменение структуры их активности, биотопическое размещение и географическое распространение, а также показывают механизм их временного и биотопического разобщения.
Большое значение в биологии рептилий имеет количество теплоты, которое получают рептилии в течение годового цикла, поскольку это обстоятельство существенно влияет на возможности адекватного протекания у них циклов питания, размножения и т. п., другими словами – на возможности выживания особей, популяций и вида в целом, их биотопического размещения и географического распространения (Baskerwille, Emin, 1969; Begon et al., 1986; Matsuzawa et al., 2004; Parker, Andrews, 2006; Valenzuela et al., 1997; Whillams, Crossman, 1977; Черлин, 2014; и др.). Пространственно-временная структура активности животных, т. е. в интересующем нас аспекте – время, которое они проводят при одинаковых температурах тела в течение суток, сезона и года, имеет для регуляции получаемого животными количества теплоты большое значение. Таким образом, модификации пространственно-временно́й структуры активности у рептилий одного вида в разных климатических и экологических условиях являются адаптивным механизмом, «согласующим» и «подгоняющим» их биологию к условиям среды.
Все описанные в данном разделе регуляционные реакции рептилий направлены на оптимизацию жизнеобеспечения особей данного вида рептилий в стабильных или меняющихся условиях среды при сохранении неизменности комплекса показателей физиолого-экологической матрицы. Они, в первую очередь, должны считаться индивидуальными адаптивными реакциями.
Регуляция основных сезонных физиологических циклов рептилий климатическими условиями
Сезонные физиологические циклы рептилий (питание, размножение) очень жестко связаны с климатическими условиями.
Циклы размножения в отношении температурных, влажностных и световых потребностей разделяются на ряд этапов. Для самцов – половая активность (когда в течение суток имеется достаточно продолжительный период термостабилизирующего или термонейтрального поведения, а суточный перепад температур тела держится около 12–15° и даже более, когда влажность повышена и возрастает длительность световой фазы активности, для дневных видов – увеличение продолжительности и интенсивности ультрафиолетового излучения), восстановительный период (когда температура тела в течение достаточно длительного времени удерживается на относительно высоком уровне и имеется продолжительный период термостабилизирующего или термонейтрального поведения для интенсивного питания и т. п.), период охлаждения (когда в течение 1–4 месяцев температура тела опускается и удерживается на низком уровне: для разных групп пресмыкающихся примерно от 20 до 0°). Для самок – половая активность (когда в течение суток повышены влажность и суточный перепад температур тела, когда возрастает длительность световой фазы активности, а для дневных видов – увеличивается продолжительность и интенсивность ультрафиолетового излучения), беременность и рождение молодняка или яйцекладка (когда уменьшается суточный перепад температур тела и когда температура тела в течение суток удерживается на высоком уровне, обычно в диапазоне 30–35°), восстановительный период (когда температура тела в течение достаточно длительного времени удерживается на относительно высоком уровне и имеется продолжительный период термостабилизирующего или термонейтрального поведения для интенсивного питания и т. п.), период охлаждения (когда в течение 1–4 месяцев температура тела опускается и удерживается на низком уровне: для разных групп пресмыкающихся примерно от 20 до 12°).
Питание возможно, только когда животное может свободно охотиться (когда в течение суток имеется достаточно продолжительный период термостабилизирующего или термонейтрального поведения) и переваривать пищу (когда температура в области желудка длительное время в течение суток может удерживаться на необходимо высоком уровне, обычно в диапазоне 30–35°).
В течение многих лет мы изучали биологию среднеазиатской эфы Echis multisquamatus (Черлин, Целлариус, 1981). Мы выяснили, что важнейшим показателем, характеризующим и регулирующим возможности реализации сезонных физиологических циклов у этих змей, является температура в норах, где змеи проводят время днем и в течение второй половины ночи (температура ночного покоя). Температура, до которой змеи нагреваются и проводят значительную часть времени днем, находится во все сезоны на уровне 28–34°. А вот температуры в норах с ранней весны к середине лета возрастают от 12–17° до 30–36°. Таким образом, именно температура почвы в норах определяет амплитуду суточных колебаний температуры тела эф и ее сезонную динамику (от весны к лету суточная амплитуда колебаний температуры тела эф уменьшается примерно с 20° почти до 0). Она обуславливает (регулирует) течение ряда важнейших сезонных физиологических процессов – состояния гонад, половой активности, течения беременности, восстановительного периода для самцов и самок, возможности переваривания пищи и т. п.
Рис. 6 и 7 показывают, как течение сезонных циклов размножения и питания определяется (регулируется) сезонной динамикой климатических (температурных) характеристик биотопов. Такая или подобная ситуация характерна и для других рептилий.
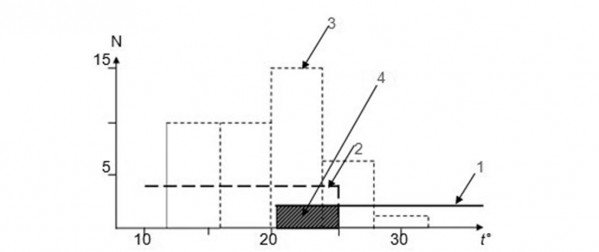
Рис. 6. Микроклиматические условия, определяющие сроки протекания половой активности у среднеазиатской эфы (по Черлин, Целлариус, 1981). N – частота встреч эф (в % от общего числа встреч), t° – температура почвы на глубине 20 см; 1 – температура почвы в норах, при которой эфы могут свободно передвигаться по поверхности (иметь длительный период термонейтрального поведения); 2 – температура почвы в норах, при которой может протекать гаметогенез; 3 – частота встреч эф на поверхности в светлое время суток (в % от общего числа встреч); 4 – интервал температур почвы в норах, при котором возможна половая активность
Fig. 6. Microclimatic conditions, determining the sexual activity period in saw scaled viper, Echis multisquamatus (by Сherlin, Cellarius, 1981). N – frequency of snakes' encounters (percentage of total encounters number), t° – soil temperature at 20 cm depth; 1 – soil temperature in holes, at which snakes can move freely over the surface (and have the long period of thermally neutral behavior); 2 – soil temperature in holes, at which gametogenesis is possible; 3 – frequency of snakes' encounters during the daytime (percentage of total snakes' encounters) ; 4 – soil temperature range in holes, at which sexual activity is possible.
Рис. 7. Микроклиматические условия, определяющие время протекания пищевой активности у среднеазиатской эфы (по: Черлин, Целлариус, 1981). t° – температура почвы на глубине 20 см; 1 – температура почвы в норах, при которой возможно переваривание пищи; 2 – температура почвы в норах, при которой возможна охота; 3 – температура почвы в норах после завершения половой активности; а – интервал температур почвы в норах, при которых возможно питание не размножавшихся в данный сезон особей, б – интервал температур почвы в норах, при котором возможно питание размножавшихся в данный сезон особей
Fig. 7. Microclimatic conditions, determining the foraging activity period in saw scaled viper, Echis multisquamatus (by Cherlin, Cellarius, 1981). t° – soil temperature at 20 cm depth; 1 – soil temperature in holes, at which digestion is possible; 2 – soil temperature in holes, at which predatory behavior is possible; 3 – soil temperature in holes after sexual activity; а – soil temperature range in holes, at which predatory behavior of snakes not breeding this year is possible; б – soil temperature range in holes, at which predatory behavior of snakes breeding this year is possible
Физиологические температурно зависимые и терморегуляционные реакции у рептилий
В ряде публикаций был подробно описан набор физиологических температурно зависимых и терморегуляционных реакций у рептилий (Черлин, 2014; и др.). В данном контексте важно ответить на вопрос: являются ли они адаптивными, и если да – то все ли они адаптивны?
Значение биохимических и физиологических температурно зависимых процессов и реакций в комплексе индивидуальных адаптаций рептилий
У рептилий описано множество биохимических и физиологических реакций, которые различным образом зависимы от температуры. Являются ли они термальными адаптациями?
Давно известно о влиянии температуры на поведение пресмыкающихся. Разные формы активности пресмыкающихся протекают в различных температурных диапазонах. Даже успех охоты за добычей в значительной степени зависит от уровня температуры тела (Greenwald, 1974). Рептилии значительную часть времени поддерживают у себя определенный диапазон температур тела, в котором успех ряда (но не всех!) важных поведенческих актов оказывается максимальным. В случаях совпадения оптимума функций с диапазоном температур термостабилизации связь с температурой можно было бы назвать адаптивной.
Температурная чувствительность биохимических соединений, отдельных тканей, органов, сложных физиологических процессов и т. п. может быть различной и связанной с разными причинами в зависимости от их функций (рис. 8).
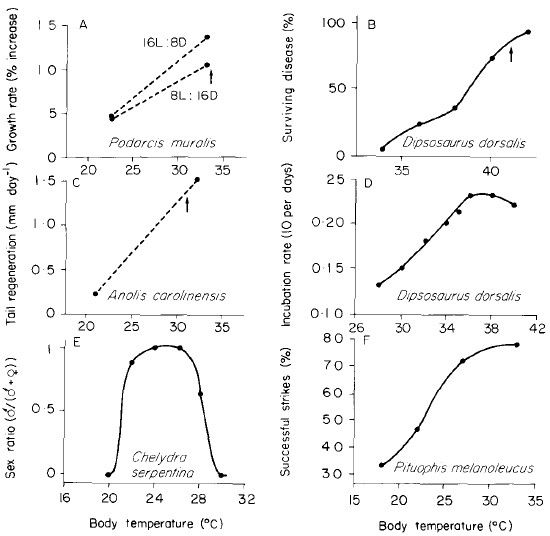
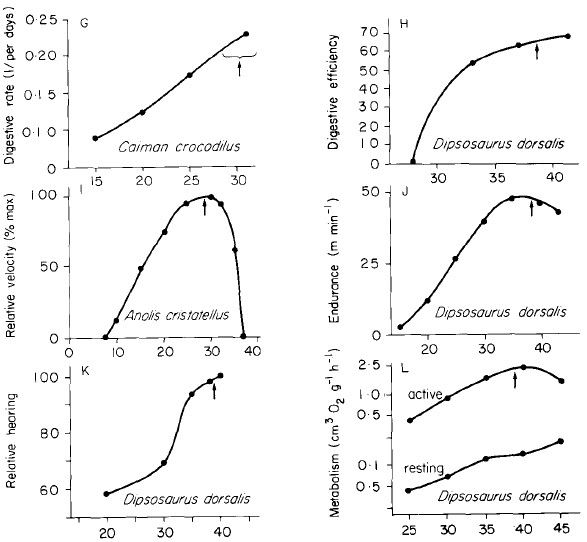
Рис. 8. Зависимость интенсивности разных функций и процессов организма рептилий от температуры тела. Вертикальная стрелка показывает предпочитаемую температуру. А (по Licht et al., 1969); B (выживание после трех дней, по Kluger, 1979), C (по Maderson, Licht, 1968), D (по Muth, 1980), E (по Bull, 1980), F (по Greenwald, 1974), G (по Diefenbach, 1975a, b), H (по Harlow et al., 1976), I (по Huey, 1982 и Huey, Webster, 1976), J (по Bennett, 1980), K (слух на частоте 1000 Hz, по Werner, 1972), L (по Bennett, Dawson, 1972)
Fig. 8. Dependence of intensity of different physiological processes on temperature of reptiles. The vertical arrow shows the preferred temperature. А (by Licht et al., 1969); B (survival after 3 days, by Kluger, 1979), C (by Maderson, Licht, 1968), D (по Muth, 1980), E (by Bull, 1980), F (by Greenwald, 1974), G (by Diefenbach, 1975a, b), H (by Harlow et al., 1976), I (by Huey, 1982 и Huey, Webster, 1976), J (by Bennett, 1980), K (hearing at the frequency of 1000 Hz, by Werner, 1972), L (by Bennett, Dawson, 1972)
Известно о связи температуры с морфологическим строением покровов (Гражданкин, 1981а, б; Черлин, 1983а; Klauber, 1941; Osgood, 1978; Fox et al., 1961; и др.). Чаще всего зависимость однозначная: при более высоких температурах количество чешуй на теле увеличивается за счет увеличения числа брюшных щитков (а значит, и числа «сегментов» туловища) и/или увеличения числа чешуй поперек тела. По нашему мнению, эта закономерность имеет адаптивное значение: увеличение числа чешуй на теле по ряду причин может усиливать транспирацию с покровов и, таким образом, увеличивать возможности охлаждения тела при высоких температурах, замедлять нагрев тела и т. п. (Гражданкин, 1981а, б; Черлин, 1983а).
Мы совместно с Р. Н. Ахмеровым изучали процессы окислительного фосфорилирования (сопряженного окисления) и окисления экзогенного НАД.Н, которое обеспечивается особой системой ферментов, отличной от основной дыхательной цепи, – редокс-цепью с цитохромом В5 (несопряженное окисление – Агуреев, Мохова, 1969; Ахмеров, 1981; Akhmerov, 1986). Исследования проводились на тканях печени животных разного уровня организации (озерной лягушки Rana ridibunda, сетчатой ящурки Eremias grammica, степной агамы Trapelus sanguinolentus и домовой мыши Mus musculus) и при разных физиологически значимых температурах (16, 25, 37 и 42°).
Фосфорилирующее, сопряженное окисление обеспечивает клетки и внутриклеточные структуры энергией для совершения определенной работы, а несопряженное, по мнению ряда авторов, может являться одним из специфических механизмов термогенеза (Ленинджер, 1966; Скулачев, 1969; Агуреев, Мохова, 1969; Арчаков, 1975; Черлин, 1988в).
К сожалению, распространенный в биохимических исследованиях вариант изучения скорости окисления в митохондриях при стандартной температуре 25° in vitro сам по себе, конечно, представляет определенный интерес, но совершенно не отражает реальные условия функционирования энергетических механизмов in vivo. И действительно, если у мышей температура тела почти все время держится в довольно узком диапазоне (близком к уровню 37° в нашем исследовании), то у интересующих нас видов рептилий ситуация совсем иная. В неактивном состоянии они находятся при температурах тела от 13–15° до 32–33° в разные сезоны года. Но активность у них всегда происходит при более высоких температурах тела: от 36–37° до 43–44°. В таком случае особый интерес представляет рассмотрение вопроса об изменении скорости окисления митохондрий в широком спектре температур тела с особым вниманием к его интенсивности при температурах нормальной активности, т. е. при физиологических температурах.
Одна из основных закономерностей, проявившаяся у всех изученных животных, заключается в возрастании интенсивности обоих видов окисления с увеличением температуры (Агзамов и др., 1992; Ахмеров и др., 1995).
Сравнение скоростей сопряженного и несопряженного окисления у представителей различных классов позвоночных животных показало, что у лягушки эти скорости при всех температурах значительно ниже, чем у пресмыкающихся и млекопитающих (рис. 9 и 10).
С ростом температуры от 16 до 42° интенсивность фосфорилирующего окисления в митохондриях у белой мыши возрастала в 3.9 раза, а у степной агамы – в 13.9 раза. Если у агамы интенсивность фосфорилирующего окисления с ростом температуры возрастала почти равномерно, то у мыши после 37° этот рост явно замедлился. Максимальный скачок в увеличении скорости фосфорилирующего окисления у мышей возник в интервале 25–37°, а у степной агамы – 37–42°. С ростом температуры от 16 до 42° интенсивность несопряженного окисления увеличилась у лягушки в 7.4, у агамы – в 15.1, у ящурки – в 4.9, у мыши – в 8.9 раза. Как и в случае сопряженного окисления, скорость роста несопряженного окисления у мышей при возрастании температуры более 37° замедлялась. У двух изученных нами видов рептилий динамика возрастания интенсивности несопряженного окисления различна. У сетчатой ящурки при переходе от 25° к 37° окисление увеличилось в 2.0 раза, а от 37° к 42° – всего в 1.1 раза. У степной агамы при переходе от 25° к 37° окисление возросло в 3.0 раза, а от 37° к 42° – в 1.8 раза.
Рис. 9. Фосфорилирующее окисление митохондрий у разных групп животных (V нг-ат О2/мин/мг белка). I – озерная лягушка, II – степная агама, III – белая мышь. Зачернен столбец, соответствующий скорости окисления при нормальной для данного вида животных активности, т. е. при «физиологической» температуре. V – в нг-ат О/мин/мг белка (скорости дыхания митохондрий печени разных животных)
Fig. 9. The mitochondrial phosphorilation in different animal groups (V ng-at О2/min/mg of albumin). I – Pelophylax ridibundus, II – Trapelus sanguinolentus, III – Mus musculus. Black column corresponds to the oxidation speed at normal activity of given species, i. e. at “physiological” temperature. V - ng-at O/min/mg of albumin (breething speed of mitochondria of the liver of different species)
Рис. 10. Несопряженное окисление НАД.Н у разных групп животных (V нг-ат О2/мин/мг белка). I - озерная лягушка, II – степная агама, III – сетчатая ящурка, IV – белая мышь. Обозначения – как на рис. 9
Fig. 10. The noncoupled mitochondrial oxidation in different animal groups (V ng-at О2/min/mg of albumin). I – Pelophylax ridibundus, II – Trapelus sanguinolentus, III – Eremias grammica, IV – Mus musculus. Black column corresponds to the oxidation speed at normal activity of given species , i.e. at “physiological” temperature.
Активность степной агамы протекает при температурах тела от 38–39° до 43–44°, оптимумом в области 40–42°. Для сетчатой ящурки этот диапазон составляет 36–43° с оптимумом 39–41°. Для белой мыши диапазон физиологических температур около 37–38°, а у озерной лягушки – около 25°.
Как уже отмечалось ранее, интенсивность сопряженного окисления у степной агамы при каждом конкретном уровне температур от 3.9 (при 16°) до 2.4 (при 37°) раза ниже, чем у мышей, а при 42° разница сократилась до 1.1 раза. Однако, если сравнить скорости фосфорилирующего окисления при уровнях физиологических температур для каждого из видов (37° для мыши и 42° для агамы), то оказывается, что они одинаковы (см. рис. 5). Поскольку фосфорилирующее окисление обеспечивает клетки и внутриклеточные структуры энергией для совершения определенной работы, одинаково высокие скорости такого окисления отмечаются именно у очень подвижных животных (ящерицы и мыши) в физиологическом диапазоне температур тела. Это, очевидно, обеспечивает высокие потенциальные возможности их активности и не зависит от таксономического положения животного. У малоподвижной лягушки эти скорости во всех случаях оказываются значительно ниже. Эта ситуация демонстрирует принципиальную невозможность для нее значительно увеличить активность даже при повышении температуры тела. Такая зависимость явно является адаптивно значимой.
Несопряженное окисление в митохондриях печени всех изученных видов животных также усиливается с возрастанием температуры (см. рис. 10). Однако при всех уровнях температур у лягушки оно оказывается в 3.4-5.6, у ящурки в 1.1 (при 16°) – 2.1, а агамы в 2.2 раза ниже, чем у мыши. При физиологических температурах (см. рис. 4) у лягушки интенсивность этого окисления в 13.3, у ящурки и агамы в 2.2 раза ниже, чем у мыши. Функциональное значение этого типа окисления для адаптаций пока неясно.
В литературе имеется много и других данных о зависимости различных биохимических процессов в организме рептилий от температуры (Баженова, 1980; Иванов и др., 1986; Кусакина, 1965; Ушаков, Даревский, 1959; Bickler, 1982; Wilhoft, 1958; Cree et al., 1990; и др).
Интересные данные получены по теплоустойчивости мышечной ткани у рептилий. Показано, что этот показатель в течение сезона у одного и того же вида изменяется: он ниже в период половой активности и беременности, выше – летом и осенью перед уходом на зимовку. С одной стороны, его изменения связаны в большей степени с гормональным фоном, чем с экологией, но с другой – он строго постоянен не только на уровне видов, но и подвидов одного вида (Ушаков, 1959, 1963). Ситуация эта сходна с таковой по критическим температурам.
Можно сказать, что температура влияет на все физиологические отправления организма: метаболизм (Проссер, 1977; Слоним, 1984; Соколова, 1980; Хозацкий, Масленникова, 1989; Шмидт-Нильсен, 1982; Al-Sadoon, 1987; Al-Sadoon, Abdo, 1991; Al-Sadoon, Spellerberg, 1985; Beaupre et al., 1993a; Bennett, Dawson, 1976; Ellis, Chappell, 1987; Niewiarowski, Waldschmidt, 1992; John-Alder et al., 1983; и мн. др.), функции иммунной системы (Mondal, Rai, 2001; Firth et al., 1989: Flatin, 1976; и др.), переваривание корма (Черномордиков, 1943; Beaupre et al., 1993 б; Chen et al., 2003; Coulson, Coulson, 1986; Deifinbach, 1975a, б; Naulleau, 1983; Pafilis, Valacos, 2004; Pafilis et al., 2007; Riddle, 1909; Saint-Girons, 1983; Skoczylas, 1982; Van Damme et al., 1991; и др.), на потребление корма (Van Damme et al., 1991; и др.), ово- и сперматогенез (Angelini et al., 1979, 1982; Action…, 1981; Callard et al., 1972; Licht, 1973; Bradshaw et al., 1980; Cowles, Bogert, 1944; Lang, 1979; Matz, 1984; Regal, 1966; Saint-Girons, 1982; Witten, Heatwole, 1978; и мн. др.), протекание беременности (Aldridge, 1979; и др.) и определение пола и морфологии потомства (Bull, 1980; Yntema, 1976; Brana, Ji, 2000; Warner, Shine, 2005; Rodríguez-Díaz et al., 2010; и др.), на сердечно-сосудистую систему (Иванов, Турдыев, 1983; Исабекова и др., 1986; Campos, 1964; Licht., 1964; Stinner, 1987; Vera, Gonzalez, 1986; и др.), дыхательную систему (Davies et al., 1982; Glass et al., 1985; Kraus, Jackson, 1980; Morris, 1984; и др.), на сенсорные системы (Campbell, 1969; Van Damme et al., 1987; Werner, 1972; и др.), подвижность (Avery, Bond, 1989; Bennett, 1980, 1990; Greenwald, 1974; и др.), защитное поведение (Christian, Tracy, 1981; Goode, Duvall, 1989; Hertz et al., 1982; Scribner, Weatherhead, 1995; и др.), на эмбриогенез, рост и развитие (Захаров и др., 1982; Chen et al., 2010; Du, Feng, 2008; Vinegar, 1974; и др.), линьку (Semlitsch, 1979; Uhiu et al., 1986; и др.), успешность охоты (Greenwald, 1974; Avery, Mynott, 1990; Diaz, 1994; и др.), образ жизни (Beaupre, 1995; Grant, Dunham, 1990; и мн. др.) и т. п. Но и физиологическое состояние может влиять на термальные предпочтения рептилий (Blouin-Demers, Weatherhead, 2001; Peterson et al., 1993; Werner, 1990; и др.).
Географическое распространение рептилий в значительной степени зависит от климатических условий и, в частности, от температурных факторов (Божанский, 1985; Динесман, 1949; Черлин, 1988а; Bustard, 1967; Monasterio et al., 2009; и др.). Также большое значение температура имеет и в выборе биотопов (Webb, Shine, 1998; Blouin-Demers, Weatherhead, 2001; Chruszcz, Barclay, 2002; Quirt et al., 2006; и др.).
Практически все аспекты жизнедеятельности рептилий оказываются температурно зависимыми (Van Damme et al., 1991; Shine, Harlow, 1993; Bauwens et al., 1995; и мн. др.). Эти и многие другие обстоятельства дают возможность считать температуру одним из важнейших, а может быть, и первостепенным по важности фактором, влияющим на все стороны жизнедеятельности рептилий.
Являются ли эти биохимические и физиологические температурно зависимые процессы и реакции термальными адаптациями? Очевидно – далеко не все. Так, скорость и эффективность пищеварения возрастают при повышении температуры. Но возникновение такого свойства не может рассматриваться как результат адаптации. Это – явление, отражающее общие биохимические и физиологические закономерности, распространяющиеся на большинство ферментов и других важных веществ в организме животных. Оно, безусловно, используется рептилиями в их ежедневной жизни. Но при этом рептилии адаптируют свою жизнедеятельность к данному свойству (в основном, за счет применения поведенческой регуляции), чтобы оптимизировать ее и сделать более энергетически эффективной, а не наоборот. Такую термозависимость физиологических функций, думаю, некорректно считать адаптивной.
Можно ли считать термальной адаптацией такое физиологическое явление, как акклимация? Акклимация – экспериментальная адаптация, приспособление организма к воздействию определенного набора внешних факторов в искусственно созданных условиях, при которых наблюдаются специфические ответные биохимические или физиологические реакции, довольно быстро возвращающиеся в прежнее состояние после прекращения действия этих искусственно созданных условий.
Так, критические температуры тела могут быть подвержены определенным сезонным изменениям. Они зависят от той температуры, при которой испытуемых животных содержали до опыта (Kour, Hutchison, 1970; Licht et al., 1966; и др.). В этих цитированных работах описаны опыты, которые были поставлены на четырех видах ящериц при трех уровнях температурной акклимации: 15, 25 и 35°. Критический максимум у одних и тех же видов в зависимости от температуры акклимации колебался в среднем в пределах 1.0–3.5°, а критический минимум – 4–6°. Наибольший размах изменчивости в опытах в первом случае был около 6°, а во втором – 10.5°. Эти реакции вполне определенно наблюдаются в условиях лабораторных экспериментов. А как обстоят дела с акклимацией у рептилий в естественных, природных условиях? Борясь с искушением назвать эту реакцию безусловно адаптивно значимой, следует напомнить, что, например, несмотря на действительное увеличение критического максимума при увеличении температуры акклимации (как это могло бы быть в природе в летний период), рептилии с температурой критического максимума в жизни никогда не сталкиваются. Хорошо развитый механизм регуляции верхней границы температуры полной активности через различные поведенческие терморегуляторные реакции этого категорически не допускает. Мало того, реальные температуры тела при активности рептилий в природе и прохладной весной, и жарким летом одинаковы (Черлин, 2014; Черлин, Музыченко, 1988), т. е. преадаптаций к низким температурам весной и высоким – летом на самом деле у рептилий в естественных условиях не происходит. Да и максимальная температура полной активности как термофизиологический показатель у рептилий внутри вида во все сезоны одинакова (Черлин, 2014). Поэтому и температуры критического максимума, изученные нами непосредственно в природе у нескольких видов пустынных ящериц в Каракумах, в разные сезоны года не различались (Черлин, Музыченко, 1984). Так что возрастание критического максимума у рептилий при возрастании температуры акклимации в лабораторных экспериментах не говорит о том, что в природе в более жарких условиях (лето или равнинные биотопы) критические температуры у обитающих там рептилий должны оказаться выше, чем при обитании в более прохладных условиях (весна или в горах). Сезонные различия в уровнях критических температур (возможно, и в других термофизиологических показателях) в лабораторных экспериментах у рептилий, скорее всего, могут быть связаны с сезонными различиями в их физиологическом состоянии.
Эффекты акклимации краткосрочные: через некоторое время после прекращения действия экспериментального фактора физиологические параметры животного возвращаются в исходное состояние. В литературе описано множество других реакций рептилий в опытах по акклимации (Du et al., 2010; Elphick, Shine, 1998; Else, Bennett, 1987; Goodman, 1971; Huang, Tu, 2009; Hochscheid et al., 2004; Huey et al., 1999; Huey, Slatkin, 1976; Garland, Adolph, 1991; Kaufmann, Bennett, 1989; Kolbe et al., 2012; Muir et al., 2010; Parker, 2014; Patterson, Davies, 1984; Qu et al., 2011; Shu et al., 2010; Scott, Pettus, 1979; Southwood et al., 2005; Stewart, 1965; Wysocki et al., 2009; Yang et al., 2008).
Таким образом, эффекты акклимации – чисто физиологические реакции, не имеющие особого адаптивного значения.
Значение физиологических терморегулирующих характеристик и реакций в комплексе индивидуальных адаптаций рептилий
По сути это комплекс географически, сезонно и популяционно неизменных физиологических констант, которые образуют стабильную физиолого-экологическую матрицу (Черлин, 2014). Параметры этой матрицы формируются видоспецифичной физиологической нормой реакции физиологических структур животного на различные внешние воздействия. Эта матрица определяет видоспецифичные параметры гомеостаза и многое другое в экологии данных видов. В рамках физиолого-этологической схемы терморегуляции эти характеристики являются терморегулирующими.
В нашем контексте имеет смысл особо обратить внимание на очень важное обстоятельство – характеристики физиолого-экологической матрицы строго видоспецифичны и неизменны ни географически, ни сезонно, т. е. в рамках комплекса терморегулирующих характеристик, составляющих физиолого-экологическую матрицу, в них не происходит внутривидовых, адаптивных, «подстроечных» изменений.
С точки зрения значения в адаптации рептилий к климатическим условиям среды термофизиологические показатели можно разделить на те, которые
1) описывают характеристики регуляции температуры тела в период полной активности (диапазон температур полной активности, минимальную и максимальную температуры полной активности, диапазон термостабилизации); эти характеристики связаны в своем проявлении с внутренней организацией механизмов терморегуляции при полной активности в нервной системе рептилий,
2) описывают характеристики регуляции температуры тела в период неактивного состояния (регуляция температуры ночного покоя, температур гибернации и эстивации),
3) описывают характеристики температурной выносливости (критические минимум и максимум температуры тела),
4) описывают характеристики, которые складываются вследствие определенного сочетания других термофизиологических характеристик (диапазон суточных колебаний температуры тела, «запас температурной прочности вида»).
Показатели первой группы, конечно, непосредственно влияют на связь температуры тела животных с климатом среды, но они видоспецифичны и неизменны в рамках всех популяций вида, где бы животные не обитали. Эти показатели надежно защищены эффективными механизмами терморегуляции (поведенческой и физиологической) от внешних влияний. Рептилии, судя по всему, не меняют их различными способами для адаптации к условиям среды, а используют механизмы терморегуляции для сохранения неизменными ключевых термофизиологических, терморегулирующих характеристик за счет тонкой подстройки пространственно-временных структур активности. Поэтому терморегуляционные реакции, безусловно, можно рассматривать как адаптивные, а вот сами термофизиологические показатели вряд ли можно считать адаптивно настроенными характеристиками.
При анализе показателей второй группы важно отметить следующее. Сами по себе показатели температуры тела в неактивном состоянии чрезвычайно важны, прежде всего для регуляции сроков и характера протекания важнейших сезонных физиологических циклов у рептилий (питания, размножения, и т. п.), т. е. для подстройки этих физиологических циклов к внешним, макроклиматическим условиям районов их обитания (Черлин, 2014а). Следовательно, эти показатели изначально имеют адаптивную направленность.
Показатели третьей группы сами по себе адаптивной направленности иметь не могут, т. к. рептилии могут столкнуться с ними в жизни только один раз – первый и последний. Подойти к температурам теплового или холодового шоков в природных условиях – это для животного смерть. Здесь намного важнее регуляция температуры тела в период полной активности (и частично – в неактивном состоянии, например на зимовках), которая призвана, кроме прочего, не подпускать температуру тела животного к опасным границам. Сами же температуры теплового и холодового шоков вряд ли имеют адаптивную составляющую, т. к. их конкретные величины, скорее всего, связаны с определенными биохимическими и физиологическими закономерностями и взаимосвязями, которые могут вообще не быть связаны с температурной выносливостью (беременность и т. п.).
Показатели четвертой группы адаптивно очень важны. Они определяют возможности рептилий данных видов выживать в данных условиях внешней среды. Так, диапазон суточных колебаний температуры тела регулирует сроки и характер протекания процессов питания, пищеварения, циклов жировых тел, проявления половой активности, гаметогенеза, созревания яиц и эмбрионов, восстановительного периода семенников и яичников и т. п.
Значение термоэкологических, терморегулируемых показателей в комплексе индивидуальных адаптаций рептилий
Термоэкологические показатели, видимо, не являются адаптивно значимыми, поскольку представляют собой лишь фактический, конечный результат взаимодействия физиологических требований организма (термофизиологических показателей) через системы терморегуляции с климатическими и экологическими условиями внешней среды, т. е. это конечные показатели, результаты разных процессов (адаптивных или неадаптивных), а не сами эти процессы. Другими словами, это показатели, которые однозначно получаются такими, какие они есть, вне зависимости от того, насколько они влияют на термальные адаптации.
Выводы о значении индивидуальных термобиологических особенностей в комплексе индивидуальных адаптаций рептилий
1) Важнейшим, ключевым феноменом в формировании адаптаций рептилий к природно-климатическим условиям среды является комплекс видоспецифичных, стабильных, географически и сезонно неизменных показателей гомеостаза (физиолого-экологическая матрица).
2) Основной смысл и направленность индивидуальных термальных адаптаций – сохранение комплекса важнейших характеристик гомеостаза (физиолого-экологической матрицы) за счет подстройки пространственно-временных структур суточной и сезонной активности.
3) Индивидуальные адаптации рептилий к термальным условиям среды обитания происходят не за счет адаптивной модификации термофизиологических характеристик, а в основном благодаря тонкой подстройке пространственно-временных структур суточной и сезонной активности, основой для которой служит физиолого-экологическая матрица.
4) Формирование, подстройка пространственно-временных структур активности у рептилий происходит посредством «отсеивания» через «сито» видоспецифичной, неизменной, физиолого-экологической, термобиологической матрицы тех временных, микроклиматических, биотопических и прочих условий, в которых активность рептилий данного вида по объективным причинам оказывается невозможной.
5) Поскольку эта матрица является видоспецифичным и неизменным комплексом терморегулирующих характеристик, то в любых биотопах с конкретными климатическими условиями рептилии данного вида могут иметь единственно возможный, «рамочный» вариант пространственно-временно́й структуры суточной и сезонной активности.
6) Физиолого-экологическая матрица через модификацию пространственно-временных структур сезонной активности также регулирует течение сезонных физиологических циклов питания и размножения (Черлин, 2014).
7) Этот единственно возможный для рептилий данного вида в данных климатических и экологических условиях «рамочный» вариант пространственно-временных структур суточной и сезонной активности, формируемый стабильной терморегулирующей физиолого-экологической матрицей, однозначно определяет в данных условиях среды количество теплоты, которое может получить организм пресмыкающегося за год.
8) Критерием адаптивных возможностей является то, насколько единственно допустимая для данного вида в данных условиях пространственно-временная структура сезонной активности позволяет или не позволяет данному виду набрать то количество теплоты, которое дает возможность реализовать все необходимые для него в течение годового цикла физиологические процессы, обеспечивающие выживание индивидуального животного и популяции.
9) Общебиологические, методологические результаты данного исследования и анализа материалов показывают необходимость сместить акцент исследований по термальным (и не только) адаптациям (по крайней мере – рептилий) с проблем биохимической, физиологической, термобиологической и пр. пластичности (хотя исследовать эти проблемы, конечно, нужно) на а) проблемы изучения комплекса показателей термального гомеостаза (физиолого-экологической матрицы) и б) различных (поведенческих и физиологических) механизмов реагирования на внешние возмущающие воздействия с целью сохранения этого гомеостатического комплекса.
На адаптации к среде, идущие через сохранение гомеостаза и развитие внешних связей, указывают примеры, касающиеся конкретных групп живых организмов (Алехин, 1938; Хаскин, 1975; и др.), а также концептуальные работы (Чернов, 1975; и др.). О необходимости сохранения гомеостаза при адаптациях биологи иногда упоминают, это положение даже периодически встречается в определениях, которые даются процессу адаптации (Краткий…, 2008), но с конкретными примерами проявления этих закономерностей, по крайней мере у эктотермных позвоночных животных, я не сталкивался. Наше исследование – пока единственная известная мне иллюстрация значения параметров гомеостаза в адаптивном процессе эктотермных позвоночных.
Заключение
Адаптация (приспособление) организма, популяции или сообщества к определенным условиям внешней среды – это процесс оптимизации необходимых и достаточных, энергетических и вещественных затрат, который благодаря комплексу поведенческих и других регуляторных реакций, биохимических, физиологических и морфологических перестроек обеспечивает благополучную и конкурентоспособную жизнедеятельность особей и популяций, процветание вида в различных условиях внешней среды.
Адаптации как биологическое явление могут иметь разную направленность и механизмы действия:
- адаптации к определенным абиотическим факторам внешней среды;
- адаптации к изменениям условий среды;
- адаптации к конкретным географическим и макроклиматическим комплексам;
- адаптации к появлению нового, сильного, определенного возмущающего фактора внешней среды;
- адаптации к условиям внешней среды в связи с появлением нового, отличного от прежнего, внутреннего состояния организма;
- адаптации в связи с появлением необходимости и/или возможности освоения новых условий внешней среды;
- адаптации в связи с необходимостью изменения в активности и направленности двигательной и психической деятельности;
- адаптации к различным условиям внешней среды, когда важным для благополучной жизнедеятельности требованием организма является сохранение стабильности комплекса ключевых физиологических параметров гомеостаза (физиолого-экологической матрицы) и т. д. и т. п.
Важнейшим, ключевым понятием в формировании адаптаций рептилий к природно-климатическим условиям среды является физиолого-экологическая матрица вида (Cherlin, 1991) – комплекс видоспецифичных, стабильных, популяционно, географически и сезонно неизменных показателей гомеостаза и видоспецифичный набор биохимических, физиологических и поведенческих приемов регуляции параметров комплекса этих показателей.
Индивидуальные адаптации к среде обитания происходят у рептилий не за счет адаптивной модификации термофизиологических характеристик, а за счет тонкой (в основном поведенческой) подстройки пространственно-временных структур суточной и сезонной активности. Подстройка пространственно-временных структур активности у рептилий происходит посредством «отсеивания» через «сито» видоспецифичной, неизменной, физиолого-экологической, термобиологической матрицы тех временных, микроклиматических, биотопических и прочих условий, в которых активность рептилий данного вида по объективным причинам оказывается невозможной. В результате получается, что данный вид рептилий в данных условиях среды может иметь единственно возможный «рамочный вариант» режима пространственно-временно́й структуры суточной и сезонной активности.
Таким образом, основной смысл и направленность индивидуальных термальных адаптаций рептилий – сохранение видоспецифичного комплекса важнейших характеристик гомеостаза (физиолого-экологической матрицы) за счет подстройки пространственно-временныхх структур суточной и сезонной активности. Критерием адаптивных возможностей является то, насколько единственно допустимая для данного вида в данных условиях пространственно-временная структура сезонной активности позволяет или не позволяет данному виду набрать то количество теплоты, которое дает возможность реализовать все необходимые для него в течение годового цикла физиологические процессы, обеспечивающие выживание индивидуального животного и популяции.
Здесь проявляется диалектическое единство двух пластов процесса адаптации у рептилий: 1) необходимость сохранения стабильности видоспецифичных характеристик гомеостаза (физиолого-экологической матрицы) в различных условиях внешней среды с помощью видоспецифичных регуляторных реакций, с одной стороны, и 2) необходимость сохранения пластичности жизнедеятельности организмов данного вида в различных условиях внешней среды с помощью модификации пространственно-временно́й структуры активности, с другой стороны.
В соответствии с этим стоит обратить внимание на то, что исследования в области адаптации животных к разным условиям внешней среды должны обязательно касаться обеих сторон этого процесса: 1) изучения стабильного, видоспецифичного комплекса характеристик и значений показателей физиолого-экологической матрицы, 2) изучения видоспецифичных регуляторных реакций, направленных на сохранение стабильности физиолого-экологической матрицы в различных условиях среды.
Это показывает необходимость сместить акцент исследований по термальным (возможно, и не только) адаптациям (по крайней мере рептилий) с проблем биохимической, физиологической, термобиологической пластичности (хотя исследовать эти проблемы, конечно, нужно) на проблемы изучения и сохранения стабильности комплекса показателей термобиологического гомеостаза и поведенческих, физиологических и прочих механизмов реагирования на внешние возмущающие воздействия с целью сохранения этого гомеостатического комплекса.
Исходя из этого можно предложить несколько определений и замечаний, связанных с понятием адаптаций, направленных на сохранение гомеостаза.
В самом общем смысле определение такой адаптации может выглядеть следующим образом.
Адаптация – биологический процесс, смысл которого заключается в сохранении комплекса видоспецифичных, ключевых, стабильных параметров гомеостаза животного (его физиолого-экологической матрицы) в различных условиях среды с помощью видоспецифичного набора биохимических, физиологических и поведенческих приемов регуляции с целью обеспечения благополучной и конкурентоспособной жизнедеятельности особей, популяций, процветания вида.
Границы адаптивных возможностей – а) границы параметров внешней среды, в рамках которых видоспецифичные приемы адаптации способны сохранять неизменным видоспецифичный комплекс физиолого-экологической матрицы; б) границы значений показателей физиолого-экологической матрицы, в рамках которых животные способны с помощью видоспецифичного набора регуляционных реакций обеспечивать благополучную и конкурентоспособную жизнедеятельность в разных условиях среды.
Очевидно, закономерности, описанные в данной статье, не являются эвристическими для всей биоты. Они проявляются в области термальной биологии и термальных адаптаций у рептилий. Так, видимо, к этому и стоит относиться. Возможно, они проявляются и у других групп живых организмов, и в других случаях, когда важным требованием сохранения успешной жизнедеятельности является сохранение комплекса стабильных физиологических характеристик гомеостаза. Но эта проблема требует отдельного, специального изучения.
Библиография
Агзамов Х., Эрматова С. М., Ахмеров Р. Н. Влияние сезонных изменений температуры окружающей среды на функциональные параметры митохондрий печени у представителей разных классов позвоночных // Журн. эволюц. биохимии и физиологии. 1998. Т. 34. № 2. С. 191–201.
Агуреев А. П., Мохова Е. Н. Регуляция живых систем и состояние энергетического обмена . Пущино, 1969. С. 27–51.
Алехин В. В. География растений . М.: Государственное учебно-педагогическое изд. Наркомпроса РСФСР, 1938. 328 с.
Арчаков А. И. Микросомальное окисление . М.: Наука, 1975. 327 с.
Ахмеров P. H. Тканевое окисление, энергопродукция и теплопродукция в разных тканях крыс // Узб. биол. ж. 1981. № 5. С. 25–27.
Ахмеров Р. Н., Гибарова С., Алламуратова Ш. И., Алматов К. Т. Соотношение дыхания митохондрий печени у млекопитающих и рептилий при разной температуре // Журн. эволюц. биохимии и физиологии. 1995. Т. 31. № 4. С. 410–415.
Баженова А. Ф. Температурные зависимости тканевого метаболизма у пустынных рептилий // Температурная компенсация и поведенческий гомеостазис. Л., 1980. С. 89–95.
Божанский А. Т. Использование климаграмм в герпетологических исследованиях на примере обыкновенной гадюки (Vípera berus) // Вопросы герпетологии. Л.: Наука, 1985. С. 32–33.
Боркин Л. Я., Черлин В. А., Басарукин А. М., Маймин М. Ю. Термобиология дальневосточного сцинка (Eumeces latiscuttatus) на острове Кунашир, южные Курильские острова // Современная герпетология. 2005. Т. 3/4. С. 5–28.
Гражданкин А. В. Роль испарительной кожной влагоотдачи в терморегуляции наземных пустынных рептилий // Вопросы герпетологии. Л.: Наука, 1981а. С. 42.
Гражданкин А. В. Кожная влагоотдача у пустынных птиц и рептилий // Зоол. ж. 1981б. Т. 60, № 2. С. 265–270.
Дедю И. И. Экологический энциклопедический словарь . Кишинев: Главная редакция Молдавской советской энциклопедии, 1989. 406 с.
Динесман Л. Г. О распространении и экологии рептилий в связи с зонами солнечной радиации // Проблемы физической географии. 1949. Т. XIV. С. 153–165.
Дудьев В. П. Психомоторика: словарь-справочник . М.: Владос, 2008. 368 с.
Захаров В. М., Баранов А. С., Валецкий А. В. Влияние температуры инкубации на продолжительность развития прыткой ящерицы – Lacerta agilis (Squamata, Lacertidae) // Зоол. ж. 1982. Т. LXL, № 6. С. 883–889.
Иванов З. В. Жизнь насекомых . М.: Сельхозгиз, 1959.
Иванов В. И., Турдыев А. А., Аббасова И. А. Липиды микросом гепатоцитов черепахи Testudo horsfieldi при различных температурах содержания // Ж. эволюц. биохим. и физиол. 1986. Т. 22. № 3. С. 240–246.
Исабекова С. Б., Чинтаева Ф. Х., Карягина Н. М. Особенности реакции кровеносной и лимфатической систем у рептилий на термовоздействия // Нейрогуморалальная регуляция вегетативных функций: Материалы науч. конф. Ин-та физиол. КазССР. Алма-Ата, 1986. С. 52–55.
Кашкаров Д. Н. Основы экологии животных . М.; Л.: Госмедиздат, 1938. 602 с.
Коросов А. В. Двухконтурная отрицательная обратная связь и модель терморегуляции гадюки // Ученые записки Петрозаводского государственного университета. Сер. «Естественные и технические науки». 2008. № 1. С. 74–82.
Коросов А. В. Экология обыкновенной гадюки на Севере . Петрозаводск: Изд-во ПетрГУ, 2010. 264 с.
Краткий толковый психолого-психиатрический словарь . М., 2008.
Кусакина А. А. Термоустойчивость гемоглобина пяти видов ящериц пустыни Каракум // Теплоустойчивость клеток животных. М.; Л.: Наука, 1965. С. 212–215.
Леках В. А. Ключ к пониманию физиологии: учебное пособие . М.: Едиториал УРСС, 2002. 360 с.
Ленинджер A. Митохондрия . М.: Мир, 1966. 167 с.
Либерман С. С., Покровская И. В. Материалы по экологии прыткой ящерицы // Зоол. ж. 1943. Т. 22. № 2. С. 247–256.
Литвинов Н. А. Термобиологические исследования // Бакиев А. Г., Гаранин В. И., Литвинов Н. А., Павлов А. В., Ратников В. Ю. Змеи Волжско-Камского края. Самара: Изд-во Самарского научного центра РАН, 2004. C. 109–146.
Меницкий Д. Н. Принципы саморегуляции функциональных систем // Системный анализ механизмов поведения / Под ред. К. В. Судакова. М.: Наука, 1979. С. 81–91.
Наумов Н. П. Экология животных . М.: Высшая школа, 1963. 618 с.
Покровский В. И. Энциклопедический словарь медицинских терминов . М.: Медицина, 2005. 1591 с.
Проссер Л. Сравнительная физиология животных . Т. 2 / Ред. Л. Проссер. М.: Мир, 1977. 84 с.
Рюмин А. В. Температурная чувствительность позвоночных животных и биологический путь происхождения теплокровных форм : Сб. студ. научных работ МГУ. М., 1939. Вып. 6. С. 55–84.
Рюмин А. В. Значение температуры в онтогенезе и филогенезе животных // Успехи современной биологии . 1940. Т. 12. № 3. С. 504–515.
Скулачев В. П. Аккумуляция энергии в клетке . М.: Наука, 1969. 440 с.
Словарь русского языка Т. Ф. Ефремовой . URL: http://enc-dic.com/efremova/Operativnyj-61489.html
Слоним А. Д. Физиология терморегуляции . Л.: Наука, Ленингр. отд-ние, 1984. 378 с.
Соколова Т. М. Энергетическая интерпретация газообмена рептилий при различных температурах и его экологическая обусловленность // Экологическая оценка энергетического баланса животных. Свердловск, 1980. С. 56–79.
Толковый словарь русского языка . Т. 2. / Под ред. Д. Н. Ушакова. М.: Советская энциклопедия: ОГИЗ: Государственное издательство иностранных и национальных словарей, 1938. 1040 с.
Ушаков Б. П. Теплоустойчивость тканей – видовой признак пойкилотермных животных // Зоол. журн. 1959. Т. 38, № 9. С. 1292–1302.
Ушаков Б. П. Изменения уровня теплоустойчивости мышечной ткани рептилий, связанные с сезоном и циклом размножения // Сб. работ Ин-та цитологии АН СССР. 1963. № 6. С. 51–61.
Ушаков Б. П., Даревский И. С. Сравнение теплоустойчивости мышечных волокон и отношение к температуре у двух симпатрических видов полупустынных ящериц // ДАН СССР. 1959. Т. 128. № 4. С. 833–835.
Хаскин В. В. Энергетика теплообразования и адаптация к холоду . Новосибирск: Наука, 1975. 200 с.
Хозацкий Л. И., Масленникова Л. С. Терморегуляция у мягкотелых черепах // Вопросы герпетологии. Киев, 1989. С. 271–272.
Целлариус Е. Ю., Целлариус А. Ю. Температурные условия активности серого варана (Varanus griseus, Reptilia, Sauria) // Зоол. ж. 1997. T. 76, № 2. С. 206–211.
Целлариус А. Ю., Черлин В. А., Лукин Ю. А. Население пресмыкающихся бессточной впадины Еройландуз (Бадхыз, Туркмения) // Известия Академии наук Туркменской ССР. Серия биологических наук. 1983. № 6. С. 63–66.
Целлариус А. Ю., Черлин В. А., Меньшиков Ю. Г. Предварительное сообщение о работах по изучению биологии Varanus griseus (Reptilia, Varanidae) в Средней Азии // Герпетологические исследования. Ленинград: ЛИСС, 1991. № 1. С. 61–103.
Чан Кьен. Систематика и экология обыкновенной гадюки Vipera berus (Linné, 1758) : Автореф. дисс. ... канд. биол. наук. Л.: ЛГУ, 1967. 14 с.
Черлин В. А. Зависимость щиткования змей рода Echis от климатических факторов // Зоологический журнал. 1983а. Т. 62. № 2. С. 252–258.
Черлин В. А. Способы адаптации пресмыкающихся к температурным условиям среды // Журнал общей биологии. 1983б. Т. 44, № 6. С. 753–764.
Черлин В. А. Географическое распространение змей рода Echis в связи с особенностями их адаптации к среде обитания // Известия Академии наук Туркменской ССР. Серия биологических наук. № 3. 1988а. С. 29–33.
Черлин В. А. К термобиологии серого геккона (Cyrtopodion russowi), полосатой ящурки (Eremias scripta) и степной агамы (Trapelus sanguinolentus) в Восточных Каракумах // Известия Академии наук Туркменской ССР. Серия биологических наук. 1988б. № 5. С. 36–43.
Черлин В. А. Интенсивность окисления митохондрий печени разных классов позвоночных животных при различных температурах // Тезисы научных сообщений IV съезда физиологов Узбекистана. Ташкент, 9–11 ноября 1988. Ташкент, 1988в. С. 140–141.
Черлин В. А. Популяционные аспекты термальных адаптаций у пресмыкающихся // Проблемы популяционной экологии земноводных и пресмыкающихся . М., 1989. С. 135–172. (Итоги науки и техники. ВИНИТИ. Серия Зоология позвоночных. Т. 17).
Черлин В. А. Стабилизация высокой температуры тела в эволюции позвоночных животных // Успехи современной биологии. 1990. Т. 109. № 3. С. 440–452.
Черлин В. А. Методы исследований по термобиологии пресмыкающихся // Экология животных Узбекистана. Ташкент, 1991. С. 70–97.
Черлин В. А. Термобиология рептилий. Общие сведения и методы исследований (руководство) . СПб.: Русско-Балтийский информационный центр «БЛИЦ», 2010. 124 с.
Черлин В. А. Биологические основы содержания пресмыкающихся в неволе: тепловой фактор . СПб.: Русско-Балтийский информационный центр «БЛИЦ», 2011. 176 с.
Черлин В. А. Термобиология рептилий. Общая концепция . СПб.: Русско-Балтийский информационный центр «БЛИЦ», 2012а. 362 с.
Черлин В. А. Термобиология рептилий и ее применение в экологических исследованиях и природоохранной деятельности // Вопросы герпетологии. Минск, 2012в. С. 344–349.
Черлин В. А. Сравнение термобиологии сцинкового (Teratoscincus scincus scincus) и гребнепалого (Crossobamon eversmanni) гекконов // Вестник Тамбовского университета. Сер. «Естественные и технические науки». 2013а. Т. 18. Вып. 6. С. 3110–3112.
Черлин В. А. Сложности и возможные ошибки при полевых исследованиях по термобиологии рептилий // Современная герпетология: проблемы и пути их решения: Статьи по материалам докладов Первой международной молодежной конференции герпетологов России и сопредельных стран (Санкт-Петербург, Россия, 25–27 ноября 2013 г.) / Зоологический институт РАН. СПб., 2013б. С. 32–39.
Черлин В. А. Сравнение термобиологии трех видов змей Средней Азии (Echis multisquamatus, Spalerosophis diadema, Psammophis lineolatum) // Працi Украïнського герпетологiчного товариства. № 4. 2013в. С. 184–189.
Черлин В. А. Рептилии: температура и экология . Saarbrücken: Lambert Academic Publishing, 2014. 442 с.
Черлин В. А., Музыченко И. В. Термобиология и экология сетчатой ящурки (Eremias grammica), ушастой (Phrynocephalus mystaceus) и песчаной (Ph. interscapularis) круглоголовок летом в Каракумах // Зоологический журнал. 1983. Т. 62. № 6. С. 897–908.
Черлин В. А., Музыченко И. В. К методике определения критического максимума температуры тела некоторых среднеазиатских ящериц // Известия Академии наук Туркменской ССР. Серия биологических наук. 1984. № 5. С. 73–76.
Черлин В. А., Музыченко И. В. Сезонная изменчивость термобиологических показателей некоторых аридных ящериц // Зоологический журнал. 1988. Т. 67. № 3. С. 406–416.
Черлин В. А., Целлариус А. Ю. Зависимость поведения песчаной эфы, Echis multisquamatus Cherlin 1981 от температурных условий в Южной Туркмении // Фауна и экология амфибий и рептилий палеарктической Азии. Л.: Наука, 1981. С. 96–108 (Труды Зоологического института АН СССР. Т. 101).
Черлин В. А., Целлариус А. Ю., Громов А. В. К температурной биологии сцинкового геккона (Teratoscincus scincus) в Каракумах // Экология. 1983. № 2. С. 84–87.
Черлин В. А., Чикин Ю. А. К термобиологии ящериц горных районов Узбекистана // Герпетологические исследования. Л.: ЛИСС, 1991. № 1. С. 119–129.
Черлин В. А., Шепилов С. А. Термобиология среднеазиатской гюрзы (Macrovipera lebetina turanica) хребта Нуратау и гюрзы Чернова (Macrovipera lebetina černovi) западных Кызылкумов // Зоологический журнал. 2014. Т. 93, № 2. С. 242–247.
Чернов Ю. И. Природная зональность и животный мир суши . М.: Мысль, 1975. 222 с.
Черномордиков В. В. О температурных реакциях пресмыкающихся // Зоологический журнал. 1943. Т. 22, № 5. С. 274–279.
Шмидт-Нильсен К. Физиология животных . Т. 1. М.: Мир, 1982. 414 с.
Эккерт Р., Рэнделл Д., Огастин Дж. Физиология животных: Механизмы и адаптация : Пер. с англ.; В 2 т. Т. 1. М.: Мир, 1991. 424 с.
Энциклопедический словарь по психологии и педагогике . 2013. URL: http://psychology_pedagogy.academic.ru// (Дата обращения 15.05.2015)
Юмашев И. Ю. Термобиологические показатели обыкновенной гадюки в бассейне Верхней Волги // Первая конференция герпетологов Поволжья: Тез. докл. Тольятти, 1995. С. 63–65.
Action des facteurs externes sur les cycles de reproduction. Les strategies de reproduction. 2 part // Bull. Soc. Zool. France. 1981. Vol. 106. № 4. P. 398–502.
Akhmerov R. N. Qualitive difference in mitohondria of endothermic and ectothermic animals // FEBS Letters. l986. Vol. 198. № 2. Р. 251–255.
Alcala А. С., Brown W. С. Thermal relation of two tropical lizards on Negros island, Phillippine islands // Copeia. 1966. № 3. P. 593–594.
Aldridge R. D. Female reproductive cycles of the snakes Arizona elegans and Crotalus viridis // Herpetologica. 1979. № 35. P. 256–261.
Al-Sadoon M. K. The influence of temperature and activity on aerobic and anaerobic metabolism in the viviparous lizard, Lacerta vivipara Jacquin // The Herpetological Journal. 1987. № 1. P. 181–185.
Al-Sadoon M. K., Abdo N. M. Temperature and body mass effects on the metabolic rate of Acanthodactylus schmidti Weigmann (Reptilia: Lacertidae) // Journal of Arid Environments. 1991. Vol. 21, № 3. P. 351–361.
Al-Sadoon M. K., Spellerberg I. F. Effect of Temperature on the Oxygen Consumption of Lizards from different Climatic Regions // Amphibia-Reptilia. 1985. Vol. 6, № 3. P. 241–258.
Amat F., Llorente G. A., Carretero M. A. A preliminary study on thermal ecology, activity times and microhabitat use of Lacerta agilis (Squamata: Lacertidae) in the Pyrenees // Folia Zool. 2003. Vol. 52. № 4. P. 413–422.
Angelini F., Brizzi R., Barone C. The annual spermatogenetic cycle of Podarcis sicula campestris De Betta (Reptilia, Lacertidae). I. The spermatogenetic cycle in nature // Monitore Zoologico Italiano, Firenze (N.S.). 1979. Vol. 13. P. 279–301.
Angelini F., Botte V., D'Alterio E. Autumn, photothermal regimes and reproductive functions in the female lizard Rodareis s. sicula Raf // Monit. Zool. Ital. 1982. Vol. 16. № 2. P. 133–148.
Angilletta M. J. Thermal Adaptation: A Theoretical and Empirical Synthesis. Oxford: Oxford University Press, 2009. 289 р.
Avery R. A., Bond D. J. Movement patterns of lacertid lizards: effects of temperature on speed, pauses and gait in Lacerta vivipara // Amphibia-Reptilia. 1989. Vol. 10, № 1. P. 77–84.
Avery R. A., Mynott A. The effects of temperature on prey handling time in the common lizard, Lacerta vivipara // Amphibia-Reptilia. 1990. Vol. 11, № 2. P. 111–122.
Bakken G. S. Measurement and application of operative and standard operative temperatures in ecology // American Zoologist. 1992. № 32. P. 194–216.
Baskerville G. L., Emin P. Rapid estimation of heat accumulation from maximum and minimum temperatures // Ecology. 1969. № 50. P. 514–517.
Bauwens D., Garland T., Castilla A. M., Van Damme R. Evolution of sprint speed in lacertid lizards: morphological, physiological, and behavioral covariation // Evolution. 1995. № 49. P. 848–863.
Beaupre S. J. Effects of geographically variable thermal environment on bioenergetics of mottled rock rattlesnakes // Ecology. 1995. № 76. P. 1655–1665.
Beaupre S. J., Dunham A. E., Overall K. L. Metabolism of a desert lizard: the effects of mass, sex, population of origin, temperature, time of day, and feeding on oxygen consumption of Sceloporus merriami // Physiological Zoology. 1993а. № 66. P. 128–147.
Beaupre S. J., Dunham A. E., Overall K. L. The effects of consumption rate and temperature on apparent digestibility coefficient and passage time in canyon lizards (Sceloporus merriami) from two populations // Functional Ecology. 1993б. № 7. P. 273–280.
Begon M., Harper J. L., Townsend C. R. Ecology: individuals, populations, and communities // Sinauer Associates. 1986. P. 876.
Bennett A. F. Thermal dependence of lizard behavior // Anim. Behav. 1980. № 28. P. 752–762.
Bennett A. F. The metabolic foundations of vertebrate behavior // Bioscience. 1980. № 30. P. 452–456.
Bennett A. F. Thermal dependence of locomotor capacity // American Journal of Physiology. 1990. № 250. P. R253–R258.
Bennett A. F., Dawson W. R. Metabolism // Biology of the Reptilia. (C. Gans and W. R. Dawson, eds). London; New York: Academic Press, 1976. Vol. 5, № 2. P. 127–223.
Bennett A. F., Dawson W. R. Aerobic and anaerobic metabolism during activity in the lizard Dipsosaurus dorsalis // J. comp. Physiol. 1972. № 81. P. 289–299.
Berk M. L., Heath J. E. An analysis of behavioral thermoregulation in the lizard, Dipsosaurus dorsalis // J. Therm. Biology. 1975. № 1. P. 15–22.
Bickler P. Intracellular pH in lizard Dipsosaurus dorsalis in relation to changing body temperatures // J. Appl. Physiol.: Respir. Environ. and Exercise Physiol. 1982. Vol. 53, № 6. P. 1466–1472.
Blouin-Demers G., Weatherhead P. J. Habitat use by Black Rat Snakes (Elaphe obsolete obsoleta) in fragmented forests // Ecology. 2001. № 83. P. 2882–2896.
Bradshaw S. D., Gans C., Saint-Girons H. Behavioral thermoregulation in a Pygopodid lizard, Lialis burtonis // Copeia. 1980. № 4. P. 738–743.
Brana F., Ji X. Influence of incubation temperature on morphology, locomotor performance, and early growth of hatchling wall lizards (Podarcis muralis) // J. Exp. Zool. 2000. Vol. 286, № 4. P. 422–433.
Brattstrom В. Н. Body temperatures of reptiles // Amer. Midl. Naturalist. 1965. Vol. 73, № 2. P. 376–492.
Bull J. J. Sex determination in reptiles // Q. Rev. Biol. 1980. № 55. P. 13–21.
Busard H. R. Gekkonid adapt fat storage to desert environments // Science. 1967. № 158. P. 1197–1198.
Callard J. P., Chan S. W. C., Potts M. The control of the reptilian gonsad // Amer. Zool. 1972. Vol. 12. № 2. P. 273–287.
Campbell H. W. The effect of temperature on auditory sensitivity of lizards // Physiol. Zool. 1969. Vol. 42, № 2. P. 183–210.
Campos V. M. Effecto de los cambios de temperature sobre las frecuencias cardiac y respiratoria del lagarto (Alligator mississipiensis) // Rev. boil. trop. Univ. Costa Rica. 1964. Vol. 12, № 1. P. 49–57.
Chen X. J., Xu X. F., Ji X. Influence of body temperature on food assimilation and locomotor performance in white-striped grass lizards, Takydromus wolteri (Lacertidae) // Journal Thermal Biology. 2003. Vol. 28, № 5. P. 385–391.
Chen Y.-H., Huang S.-P., Chang M.-H., Tu M.-C. Thermal Effects on Embryogenesis and Hatchlings of the Grass Lizard Takydromus stejnegeri (Squamata: Lacertidae) and Implications of Their Potential for Limiting Its Altitudinal Distribution in Taiwan // Zoological Studies. 2010. Vol. 49, № 3. P. 374–380.
Cherlin V. A. Thermal biology of Middle Asian reptiles // First World Congress of Herpetology. 11–19 september 1989. University of Kent at Canterbury. United Kingdom, 1989. P. 10.
Cherlin V. A. Method of Ecology-Physiological Matrix of Species in Researches on Reptile Biology // Герпетологические исследования. Л.: ЛИСС, 1991. № 1. С. 138–146.
Cherlin V. A., Shepilov S. A. Thermal Biology of the Central Asian Blunt-Nosed Viper (Macrovipera lebetina turanica) from Nuratau Crest and the Chernov Blunt_Nosed Viper (Macrovipera lebetina černovi) from Western Kysylkum // Biology Bulletin. 2014. Vol. 41. № 8. P. 639–644.
Christian K. A., Tracy C. R. The effect of the thermal environment on the ability of hatchling Galapagos land iguanas to avoid predation during dispersal // Oecologia (Berlin). 1981. № 49. P. 218–223.
Chruszcz B. J., Barclay R. M. R. Thermoregulatory ecology of a solitary bat, Myotis evotis, roosting in rock crevices // Functional Ecology. 2002. № 16. P. 18–26.
Cloudsley-Thompson J. L. The Temperature and Water Relations of Reptiles. England: Merrow Publishing Co., Ltd., 1971.
Coulson R. A., Coulson T. D. Effect of temperature on the rate of digestion, amino acid absorption and assimilation in the alligator // Comp. Biochem. And Physiol. 1986. Vol. A83. № 3. P. 585–588.
Cowles R. B., Bogert C. M. Preliminary study of the thermal requirements of desert reptiles // Bulletin of the American Museum of Natural History. 1944. № 83. P. 261–296.
Cree A., Guilette L. J., Cockrem J. F., Joss J. M. P. Effects of capture and temperature stresses on plasma steroid concentration in mail tuatara (Sphenodon punctatus) // J. Exp. Zool. 1990. Vol. 253. № 1. P. 38–48.
Davies D. G., Thomas J. L., Smith E. N. Effect of body temperature on ventilator control in the alligator // J. Appl. Physiol.: Respir. Environ. and Exercise Physiol. 1982. Vol. 52. № 1. P. 114–118.
Diaz J. A. Effects of body temperature on the predatory behaviour of the lizard Psammodromus algirus hunting winged and wingless prey // The Herpetological Journal. 1994. № 4. P. 145–150.
Diefenbach C. O. da C. Gastric function in Caiman crocodilus (Crocodylia: Reptilia) I. Rate of gastric digestion and gastric motility as a function of temperature // Comp. Biochem. Physiol. 1975a. № 51A. P. 259–265.
Diefenbach C. O. da C. Gastric function in Caiman crocodilus (Crocodylia: Reptilia) II. Effects of temperature on Ph and proteolysis // Comp. Biochem. Physiol. 1975б. № 51A. P. 267–274.
Du W.-G., Feng J.-H. Phenotypic effects of thermal mean and fluctuations on embryonic development and hatchling traits in a lacertid lizard, Takydromus septentrionalis // Journal of Experimental Zoology Part A: Ecological Genetics and Physiology. 2008. Vol. 309A, № 3. P. 138–146.
Du Wei-Guo, Ye H., Zhao B., Warner D. A., Shine R. Thermal Acclimation of Heart Rates in Reptilian Embryos // PLoS One. 2010. Vol. 5. № 12. E15308.
Elphick M. J., Shine R. Longterm effects of incubation temperatures on the morphology and locomotor performance of hatchling lizards (Bassiana duperreyi, Scincidae) // Biol. J. Linn. Soc. 1998. Vol. 63. № 3. P. 429–447.
Ellis T. M., Chappell M. A. Metabolism, temperature relations, maternal behavior, and reproductive energetic in the ball python (Python regius) // J. Comp. Physiol. 1987. Vol. B157, № 3. P. 393–402.
Else P. L., Bennett A. F. The thermal dependence of locomotor performance and muscle contractile function in the salamander Ambystoma tigrinum nebulosum // J. Exp. Biol. 1987. № 128. P. 219–233.
Firth B. T., Thompson M. B., Kennaway D. J., Belan I. Thermal sensitivity of reptilian melatonin rhythms: “cold” tuatara vs. “warm” skink // Amer. J. Physiol. 1989. Vol. 256. № 5. P. R1160–R1163.
Flatin J. Sensibilite tissularie aux hormones en function de la temperature // Bull. Soc. Zool. France. 1976. Vol. 101. P. 941–942.
Fox W., Gordon Ch., Fox M. Morphological effects of low temperature during the embryonic development of the gater snake, Thamnophis elegans // Zoologica. 1961. № 46. P. 57–71.
Garland T., Adolph S. C. Physiological differentiation of vertebrate populations // Annu. Rev. Ecol. Syst. 1991. Vol. 22. P. 193–228.
Glass M. L., Boutilier R. G., Heisler N. Effects of body temperature on respiration, blood gases and asid-base status in the turtle Chrysemys picta bellii // J. Exp. Biol. 1985. Vol. 114. P. 37–51.
Goode M. J., Duvall D. Body temperature and defensive behaviour of free-ranging prairie rattlesnakes, Crotalus viridis viridis // Animal Behaviour. 1989. № 38. P. 360–362.
Goodman D. E. Thermoregulation in the Brown Water Snake, Natrix taxispilota, with Discussion of the Ecological Significance of Thermal Preferenda in the Order Squamata: Ph. D. Dissertation. University of Florida, Gainesville, Florida, 1971.
Grant B. W, Dunham A. E. Elevational covariation in environmental constraints and life histories of the desert lizard Sceloporus merriami // Ecology. 1990. № 71. P. 1765–1776.
Greenwald O. E. Thermal dependence of striking and prey capture by gopher snakes // Copeia. 1974. № 1. P. 141–148.
Harlow J. J., Hillman S. S. and Hoffman M. The effect of temperature on digestive efficiency in the herbivorous izard Dipsosaurus dorsalis // J. comp. Physiol. 1976. № 111. P. 1–6.
Heath J. E. Behavioral regulation of body temperature in poikilotherms // The Physiologist. 1970. № 13. P. 339–410.
Hertz P. E. Thermal passivity of a tropical forest lizard, Anolis polylepis // J. Herpetol. 1974. № 8. P. 323–327.
Hertz P. E., Huey R., Nevo E. Fight versus flight: thermal dependence of defensive behaviour in a lizard // Animal Behaviour 1982. № 30. P. 676–679.
Hochscheid S., Bentivegna F., Speakman J. R. Long-Term Cold Acclimation Leads to High Q10 Effects on Oxygen Consumption of Loggerhead Sea Turtles Caretta caretta // Physiological and Biochemical Zoology. 2004. Vol. 77. № 2. P. 209–222.
Huang S. P., Tu M. C. Locomotor and Elevational Distribution of a Mountainous Lizard, Takydromus hsuehshanensis, in Taiwan // Zoological Studies. 2009. Vol. 48. № 4. P. 477–484.
Huey R. B. Behavioral thermoregulation in lizards: importance of associated costs // Science. New Series. 1974. 184(4140). P. 1001–1003.
Huey R. Temperature, physiology, and the ecology of reptiles // C. Gans and F. H. Pough (eds.). Biology of the Reptilia. Vol. 12. Physiology (C). London: Academic Press, 1982. P. 25–91.
Huey R. B., Berrigan D., Gilchrist G. W., Herron J. C. Testing the Adaptive Significance of Acclimation: A Strong Inference Approach (1) // American Zoologist. 1999. Vol. 39. № 2. P. 15.
Huey R. B., Slatkin M. Cost and benefits of lizard thermoregulation // Quarterly Review of Biology. 1976. № 51. P. 363–384.
Huey R., Webster T. P. Thermal biology of Anolis lizards in a complex fauna: the cristatellus group on Puerto Rico // Ecology. 1976. № 57. P. 985–994.
John-Alder H. B., Lowe C. H., Bennett A. F. Thermal dependence of locomotory energetic and aerobic capacity of the gila monster (Heloderma suspectum) // J. Comp. Physiol. 1983. Vol. B151. № 2. P. 119–126.
Kardong K. V. Observations on live Azemiops feae, Fea’s viper // Herpetological Review. 1986. Vol. 17. № 4. P. 81–82.
Kaufmann J. S., Bennett A. F. The Effect of Temperature and Thermal Acclimation on Locomotor Performance in Xantusia vigilis, the Desert Night Lizard // Physiological Zoology. 1989. Vol. 62, № 5. P. 1047.
Klauber L. M. Four papers on the application of the statistical methods to herpetological problems. III. The correlation between scalation and life zones in San Diego County snakes // Bull. Zool. Soc. San Diego. 1941. № 17. P. 73–79.
Kluger M. J. Fever in ectotherms: evolutionary implications // Am. Zool. 1979. № 19. P. 295–304.
Kolbe J. J., VanMiddlesworth P. S., Losin N., Dappen N., Losos J. B. Climatic niche shift predicts thermal trait response in one but not both introductions of the Puerto Rican lizard Anolis cristatellus to Miami, Florida, USA. The Authors. Published by Blackwell Publishing Ltd., 2012. P. 1503–1516.
Кour E. L., Hutchison V. H. Critical thermal tolerances and heating and cooling rates of lizards from diverse habitats // Copeia. 1970. № 2. P. 219–229.
Kraus D. R., Jackson D. C. Temperature effects on ventilation and acid-base balance of the green turtle // Amer. J. Physiol. 1980. Vol. 239. № 3. P. R254–R258.
Lang J. W. Thermophilic response of the American alligator and the American crocodile to feeding // Copeia. 1979. P. 48–59.
Leeuwen J. P. The adder (Vipera berus). Problematic isolation in Dutch populations? // Alterra. 1009. July. P. 24.
Licht P. The effect of temperature on hart rates of lizards during rest and activity // Physiol. Zool. 1964. № 38. P. 129–137.
Licht P. Photoperiodic and thermal influences on reproductive cycles in reptiles // Endocrinology. Amsterdam; New-York, 1973. P. 185–190.
Licht P., Dawson W. R., Shoemaker V. H, Main A. R. Observations on the thermal relations on Western Australian lizards // Copeia. 1966. № 1. P. 97–110.
Lourdais O., Guillon V., DeNardo D., Boulin-Demers G. Cold climate specialization: adaptive covariation between metabolic rate and thermoregulation in pregnant vipers // Physiology & Behavior. 2013. № 119. P. 149–155.
Maderson P. F. A., Licht P. Factors influencing rates of tail regeneration in the lizard Anolis carolinensis // Experientia. 1968. № 24. P. 1083–1086.
Matsuzawa Y., Sato K., Sakamoto W., Bjorndal K. A. Seasonal fluctuations in sand temperature: effects on the incubation period and mortality of loggerhead sea turtle (Caretta caretta) pre-emergent hatchlings in Minabe, Japan // Marine Biology. 2004. № 140. P. 639–646.
Matz G. La reproduction des reptiles et les facteurs de son induction // Acta zool. et pathol. Antverp. № 78. Mainten. and reprod. reptile captive. 1984. Vol. 1. P. 33–68.
Monasterio C., Salvador A., Iraeta P., Díaz J. A. The effects of thermal biology and refuge availability on the restricted distribution of an alpine lizard // Journal of Biogeography. 2009. Vol. 36. № 9. P. 1673–1684.
Mondal S., Rai U. In vitro effect of temperature on phagocytic and cytotoxic activities of splenic phagocytes of the wall lizard, Hemidactylus flaviviridis // Comp. Biochem. Physiol. 2001. № A129. P. 391–398.
Muir T. J., Costanzo J. P., Lee R. E. Brief Chilling to Subzero Temperature Increases Cold Hardiness in the Hatchling Painted Turtle (Chrysemys picta) // Physiological and Biochemical Zoology. 2010. Vol. 83. № 1. P. 1740–1781.
Muth A. Physiological ecology of desert iguana (Dipsosaurus dorsalis) eggs: temperature and water relations // Ecology. 1980. № 61. P. 1335–1343.
Naulleau G. Action de la temperature sur la digestion chez cinq espéces de vipéres européenes du genre Vipera // Bull. Soc. Zool. Fr. 1983. № 108. P. 47–58.
Niewiarowski P. H., Waldschmidt S. R. Variation in metabolic rates of a lizard: use of SMR in ecological contexts // Functional Ecology. 1992. № 6. P. 15–22.
Nilson G. Ovarian cycle and reproductive dynamics in the female Adder, Vipera berus (Reptilia, Viperidae) // Amphibia-Reptilia. 1981. № 2. P. 63–82.
Orlov N. Viperid snakes (Viperidae Bonaparte, 1840) of Tam-Dao mountain range // Russian Journal of Herpetology. 1997. Vol. 4. № 1. P. 67–74.
Osgood D W. Effect of temperature on the development of meristic characters in-Natrix fasciata // Copeia. 1978. № 1. P. 33–48.
Pafilis P., Valakos E. D. Temperature effect on the digestive efficiency of the main organic compounds in two Mediterranean lizards // Proceedings 10th MEDECOS Conference, April 25 – May 1. 2004. Rhodes, Greece. Arianoutsou & Papanastasis (eds).
Pafilis P., Foufopoulos J., Poulakakis N., Lymberakis P., Valakos D. Digestive performance in five Mediterranean lizard species: effects of temperature and insularity // Journal of Comparative Physiology B: Biochemical, Systemic, and Environmental Physiology. 2007. Vol. 177. № 1. P. 49–60.
Parker J. R. Some effects of temperature and moisture upon Melanoplus mexicanus mexicanus Saussure and Camnulapellucida Scudder (Orthoptera) // Univ. Montana. Agri. Exp. Sta. Bull. 1930. № 223. P. 1–132.
Parker S. L. Physiological ecology of the ground skink, Scincella lateralis in South Carolina: thermal biology, metabolism, water loss, and seasonal patterns // Herpetological Conservation and Biology. 2014. Vol. 9. № 2. P. 309–321.
Parker S. L., Andrews R. M. Incubation temperatures and phenotypic traits in Sceloporus undulatus: implications for the northern limits of distribution // Oecologoa. 2006. DOI 10.1007/s00442-006-0583-0. Springer. 15 p.
Patterson J. W., Davies P. M. C. The influence of temperature, sexual condition, and season on the metabolic rate of the lizard Psammodromus hispanicus // J. Comp. Physiol. B. 1984. № 154. P. 311–316.
Peterson C. R., Gibson A. R., Dorcas M. E. Snake thermal ecology: the causes and con-sequences of body-temperature variation // R. A. Seigel and J. T. Collins (Eds.). Snakes: Ecology and Behavior. McGraw-Hill, New York, U.S.A., 1993. P. 241–314.
Qu Yanfu et al. Thermal preference, thermal tolerance and the thermal dependence of digestive performance in two Phrynocephalus lizards (Agamidae), with a review of species studied // Current Zoology. 2011. Vol. 57. № 6. P. 684−700.
Quirt K. C., Blouin-Demers G., Howes B. J., Lougheed S. C. Microhabitat Selection of Five-Lined Skinks in Northern Peripheral Populations // Journal of Herpetology. 2006. Vol. 40. № 3. P. 335-342.
Rand A. S., Humphrey S. S. Interspecific competition in the tropical rain forest: ecological distribution among lizards at Belem, Para // Proc. U.S. Natl. Mus. 1968. № 125. P. 1–17.
Regal P. J. Thermophilic response following feeding in certain reptiles // Copeia. 1966. № 3. P. 588–590.
Riddle O. The rate of digestion in the cold-blooded vertebrates – the influence of season and temperature // Am. J. Physiol. 1909. № 24. P. 447–458.
Rodríguez-Díaz T., González F., Ji X., Braña F. Effects of incubation temperature on hatchling phenotypes in an oviparous lizard with prolonged egg retention: are the two main hypotheses on the evolution of viviparity compatible? // Zoology. 2010. Vol. 113. № 1. P. 33–38.
Ruibal R. Thermal relations of ve species of tropical lizards // Evolution. 1961. № 15. P. 98–111.
Ruibal R., Philibosian R. Eurythermy and niche expansion in lizards // Copeia. 1970. № 4. P. 645–653.
Saint-Girons H. Thermoregulation in reptiles with special reference to the tuatara and its ecophysiology // Tuatara. 1980. Vol. 24. № 2. P. 59–80.
Saint-Girons H. Reproductive cycles of male snakes and their relationships with climate and female reproductive cycles // Herpetologica. 1982. Vol. 38. № 1. P. 5–16.
Saint-Girons H. Regime et rations alimentaires des serpents // Bull. de la Soc. Zool. de France. 1983. Vol. 108. № 3. P. 431–437.
Scott J. R., Pettus D. Effects of seasonal acclimation on the preferred body temperature ofThamnophis elegans vagrans // J. Therm. Biol. 1979. №o 4. P. 307–309.
Scribner S. J., Weatherhead P. J. Locomotion and antipredator behavior in three species of semi-aquatic snakes // Canadian Journal of Zoology. 1995. № 73. P. 321–329.
Semlitsch R. D. The influence of temperature on ecdysis rates in snakes (Genus Natrix) (Reptilia, Serpentes, Colubridae) // Journal of Herpetology. 1979. Vol. 13. № 2. P. 212–214.
Shine R., Harlow P. Maternal thermoregulation influences offspring viability in a viviparous lizard // Oecologia. 1993. № 96. P. 122–127.
Shu L., Sun B. J., Du W. G. Effects of temperature and food availability on selected body temperature and locomotor performance of Plestiodon (Eumeces) chinensis (Scincidae) // Animal Biology. 2010. Vol. 60. P. 337–347.
Skoczylas R. Gastric motility in the grass snake (Natrix natrix L.) in different ambient temperatures // Exogeneus and Endogenous Influence Metab. and Neural Contr. Respirat., Read., Activ. and Energy Supply Muscles, Ion and Osmoregulat., Reprod., Percept., and Orientat. 3rd Congr. Eur. Soc. Comp. Physiol. And Biochem., Noordwijkerhout, 31 Aug. – 3 Sept. 1981. 1982. Vol. 3. Abstr. Oxford, 1981. P. 11–12.
Southwood A. L., Andrews R. D., Paladino F. V., Jones D. R. Effects of Diving and Swimming Behavior on Body Temperatures of Pacific Leatherback Turtles in Tropical Seas // Physiological and Biochemical Zoology. 2005. Vol. 78. № 2. P. 13.
Souza F. L., Martins F. I. Body temperature of free-living freshwater turtles, Hydromedusa maximiliani (Testudines, Chelidae) // Amphibia-Reptilia. 2006. № 27. P. 464–468.
Stebbins R. C., Lowenstain J. M., Cohen N. W. A field study of the Lava Lizard (Tropidumal bemarlensis) in the Galapagos Islands // Ecology. 1967. № 48. P. 839–851.
Stewart G. R. Thermal ecology of the garter snakes Thamnophis sirtalis concinnus (Hallowell) and Thamnophis ordinoides (Baird and Girard) // Herpetologica. 1965. № 21. P. 81–101.
Stinner J. N. Cardiovascular and metabolic responses to temperature in Coluber constrictor // Amer. J. Physiol. 1987. Vol. 253. № 2. P. 222–227.
Templeton J. R. Reptiles // The Comparative Physiology of Thermoregulation. (G. C. Whittow, ed.). New York and London: Academic Press, 1970. Vol. 1. P. 167–221.
Uhiu K. W., Sham J. S. K., Maderson P. F. A., Zucker A. H. Interaction between thermal environments and hormones affecting skin-shedding frequency in the tokay (Gekko gecko) (Gekkonidae, Lacertilia) // Comp. Biochem. and Physiol. 1986. Vol. 84, № 2. P. 245–351.
Valenzuela N., Botero R., Martinez E. Field study of sex determination in Podocnemis expansa from Colombian Amazonia // Herpetologica. 1997. Vol. 53. № 3. P. 390–398.
Van Damme R., Bauwens D., Verheyen R. F. The thermal dependence of feeding behaviour, food consumption and gut-passage time in the lizard Lacerta vivipara Jacquin // Functional Ecology. 1991. № 5. P. 507–517.
Van Damme R., Vanderstighelen D., Verheyen R. Thermal dependence of chemoreceptive predator-recognition in Lacerta vivipara // Proc. 4th Ordinary Gen. Meet. Soc. Eur. Herpetol., Nijmegen, 17–21 Aug. 1987. P. 415–419.
Vera P. L. de, Gonzalez G. J. Cardiac responses to temperature in the lizard Gallotia galliti // Comp. Biochem. And Physiol. 1986. Vol. A85. № 2. P. 389–394.
Vinegar A. Evolutionary implications of temperature induced anomalies of development in snake embryos // Herpetologica. 1974. № 30. P. 72–74.
Warner D. A., Shine R. The adaptive significance of temperature-dependent sex determination: experimental tests with a short-lived lizard // Evolution Int. J. Org. Evolution. 2005. Vol. 59, № 10. P. 2209–2221.
Webb J. K., Shine R. Using thermal ecology to predict retreat-site selection by an endangered snake species // Biological Conservation. 1998. № 86. P. 233–242.
Werner Y. L. Temperature effects on inner-ear sensitivity in six species of iguanid lizards // J. Herpetol. 1972. № 6. P. 47–177.
Werner Y. L. Do gravid females of oviparous gekkonid lizards maintain elevated body temperatures? Hemidactylus frenatus and Lepidodactylus lugubris on Oahu // Amphibia-Reptilia. 1990. № 11. P. 200–204.
Whillans T. H., Crossman E. J. Morphological parameters and Spring activities in a central Ontario population of Midland Painted Turtle, Chrysemys picta marginata (Agassiz) // Canadian field-Naturalist. 1977. Vol. 91. № 1. P. 47–57.
Wilhoft D. C. The effect of temperature on thyroid history and survival in the lizard, Sceloporus occidentalis // Copeia. 1958. № 4. P. 265–276.
Witten G. J., Heatwole H. Preferred temperature of the agamid lizard Amphibolurus nobbinobbi // Copeia. 1978. № 2. P. 362–364.
Wysocki L. E., Montey K., Popper A. N. The influence of ambient temperature and thermal acclimation on hearing in a eurythermal and a stenothermal otophysan fish // The Journal of Experimental Biology. 2009. № 212. P. 3091–3099.
Yang J., Sun Y. Y., An H., Ji X. Northern grass lizards (Takydromus septentrionalis) from different populations do not differ in thermal preference and thermal tolerance when acclimated under identical thermal conditions // J. Comp. Physiol. B. 2008. № 178. P. 343–349.
Yntema C. L. Effects of incubation temperature on sexual differentiation in the turtle (Chelydra serpentina) // J. Morphol. 1976. № 150. P. 453–461.

 © 2011 - 2026
© 2011 - 2026