



Уже более семидесяти лет назад, в 1943 г., в Зоологическом журнале была опубликована статья С. С. Либерман и Н. В. Покровской «Материалы по экологии прыткой ящерицы» (Либерман, Покровская, 1943). Под этим непритязательным названием оказалась работа совершенно замечательная, которая явно опередила свое время. В те годы термобиология рептилий еще только зарождалась. В Советском Союзе к тому времени было опубликовано всего несколько статей, касающихся этой темы (Стрельников, 1934; Родионов, 1938; Сергеев, 1939; Черномордиков, 1943). Англо- и немецкоязычных материалов было несколько больше: с 1842 г., когда появилась первая публикация, касающаяся темы теплового баланса рептилий (статья Lamarre-Pacquot, 1842 о том, что в Бенгалии насиживающие самки питонов повышают температуру тела на несколько градусов), по начало 1944 г. мне известно их всего 77 (Agassiz, Gould, 1856; Weese, 1917; Hall, 1922; Mosauer, Lazier, 1933; Herter, 1941; и др). Лишь в следующем, 1944 г., появится работа Р. Б. Коула и Ч. М. Богерта (Cowles, Bogert, 1944), от которой, в основном, и отсчитывается сейчас время существования данного направления науки. Именно эта работа заложила базис термобиологии рептилий, основы терминологии, методические подходы и т. п. Однако по своей реальной значимости статья С. С. Либерман и Н. В. Покровской ничуть ей не уступает и именно она могла бы считаться такой пионерной работой. Но, во-первых, она была опубликована во время Второй мировой войны, во-вторых, на русском языке, а значит, осталась практически неизвестной западным специалистам. К сожалению, ее и по сей день мало кто вспоминает. В англоязычных статьях она изредка цитируется, но, судя по материалам этих статей, о ней вспоминают лишь постольку, поскольку она существует. В ее глубинную суть (для чего надо ее добросовестно перевести и прочесть) авторы не вникают, из западных авторов в нее никто, похоже, серьезно не вчитывался.
Статья С. С. Либерман и Н. В. Покровской являет нам пример очень подробной, научно обоснованной, комплексной работы по изучению животных. В первой ее части авторы описали температурные условия существования прытких ящериц Lacerta agilis, во второй – особенности их размножения и развития яиц. Обе части одинаково интересные и качественные. И это не очень удивительно, поскольку, не умаляя заслуг самих авторов, нужно помнить, что научное руководство работой осуществлял замечательный советский биолог А. М. Сергеев.
Работы начала – середины XX века делались зоологами, которые были еще в значительно большей степени «натуралистами», чем современные ученые («натуралист» – человек, который любит и знает интересующий его живой объект и природу вокруг, умеет его наблюдать и видеть, понимать непосредственно, почти без привлечения серьезного аппаратного обеспечения и суперсовременных методов статистической обработки). Поэтому их наблюдения имеют большую научную ценность, к ним надо относиться с уважением и вниманием, в них можно встретить чрезвычайно ценные детали, которые, к сожалению, упускаются порой их последователями, т. е. нашими современниками.
Обращает на себя внимание очень подробное изучение суточного хода микроклиматических параметров во всей зоне обитания ящериц, включая норы. Это позволило получить полную суточную динамику температур тела ящериц. Авторы, раскопав несколько нор, получили данные о том, где находились эти рептилии ночью, и, таким образом, определили, что их минимальная температура тела за сутки составляла 14–15°.
Стоит отметить, что в этом исследовании применялось измерение температуры тела у ящериц «per os», как пишут сами авторы. Также они описывают, что измерения производились термометром от аспирационного психрометра Ассмана. Баллончик со ртутью у этого термометра достаточно мал, чтобы позволить измерять ректальную температуру тела таких рептилий среднего размера, как взрослая обыкновенная гадюка, эфа, обыкновенный уж или амурский полоз. Но он слишком велик для измерения температуры тела у животных такого размера и строения, как прыткая ящерица. К сожалению, тогда в СССР не было изготовленного промышленным способом адекватного оборудования для этих целей. Измерения «per os» с помощью термометра Ассмана у прытких ящериц могут означать только измерения во рту и верхней части пищевода, поскольку проникновение его более глубоко маловероятно. Кроме того, он слишком температурно инерционен, поэтому может дать большие ошибки при измерениях таких небольших объектов. Сейчас мы знаем, что это вообще не самый удачный вариант методики, поскольку он, скорее, показатель температуры головы рептилии, чем «ядра» ее туловища. Теперь нам также известно, что на самом деле ящерицы наиболее четко регулируют именно температуру туловища, но в те годы этой информации еще не было. Да и существует очевидная разница между температурами головы и туловища (Crawford et al., 1977; etc.). Поэтому сравнение данных, касающихся температур тела, измеренных «per os» или ректально (т. е. температур головы и туловища), показывает, что они могут заметно различаться. При этом «разброс» температур головы больше, чем туловища, т. е. температура во рту и пищеводе может в определенные моменты оказаться несколько выше или ниже ректальной температуры. Поэтому при измерениях температур тела прытких ящериц у авторов статьи получились данные, порой существенно отличающиеся от указанных другими исследователями. Так, температура тела (ректальная) прытких ящериц при активности практически никогда не превышает 36° (Amat et al., 2003; наши измерения и мн. др.), а в Заключении своей статье С. С. Либерман и Н. В. Покровская указали максимальную температуру (во рту) 41°, что очень близко к указанным ими же температурам теплового окоченения.
Но несмотря на некоторые неточности и даже, может быть, некорректности, в статье С. С. Либерман и Н. В. Покровской появился ряд очень важных новаций, которые в дальнейшем, в работах других авторов, получили свое развитие. Точнее было бы сказать, что на самом деле немногие западные герпетологи были знакомы с этой статьей (т. е. немногие ее читали, а не просто были знакомы с фактом ее существования или, в крайнем случае, с цифрами, приведенными в таблицах), но к развитию системы терминов и понятий толкала общая логика полученных в исследованиях разных авторов материалов. Фактически некоторые новации, впервые появившиеся в данной статье, потом другими авторами «переоткрывались», а затем и развивались.
Эти новации в статье С. С. Либерман и Н. В. Покровской следующие.
1. При определении термобиологических показателей авторы полностью перешли от регистрации температур среды (Herter, 1941) к измерениям температур тела.
2. В этой работе впервые был использован целый набор взаимосвязанных терминов и понятий в области термобиологии рептилий. К сожалению, авторы в дальнейшем не дали им строгих определений, но на стр. 251 своей статьи, на рис. 7 (см. рис. в моей статье) они также первый раз привели объединенную картину термобиологических показателей. Они применили их вполне корректно, четко проиллюстрировали их на рисунке, так что их смысл оказался вполне понятен.
3. Авторы впервые выявили различные статусы активности пресмыкающихся. Они выделили неактивное состояние и активность, которую, в свою очередь, разделили на неполную и полную.
Поскольку в статье, к сожалению, нет строгих определений статусов активности, то очень важно, что авторы привели описание того состояния ящериц, которое они определили как неактивное, и переход их в состояние полной активности. Эти описания приведены применительно к ящерицам, которых они достали из нор в 4 часа утра: «Из одной норки (t° – 14.8°) были выкопаны два самца, температура их тела 15 и 14°. Из другой норки (на глубине 29 см, t° 15.4°) был выкопан один самец, температура его тела 15°. Ящерицы не могли передвигаться, лишь слабо шевелили конечностями и медленно сгибали тело. Тонус мускулатуры очень понижен, дыхание редкое. На солнце температура тела постепенно повышалась, следуя за повышением температуры воздуха и все время несколько ее обгоняя. При температуре тела 25.5 и 27.5° (t° воздуха 20°) ящерицы активны, но кожа еще в складках, конечности дряблые. Полная подвижность приобреталась при температуре тела 30.5°» (стр. 249).
На рисунке явно проявляется понимание авторами смысла и места состояний неполной и полной активности. В выводах статьи (в Заключении на стр. 255) они выделили состояния активности в целом и отдельно – полной активности. Это очень важное теоретическое обстоятельство, т. к. полная активность – это состояние, резко отличающееся от состояний неактивного и неполной активности.
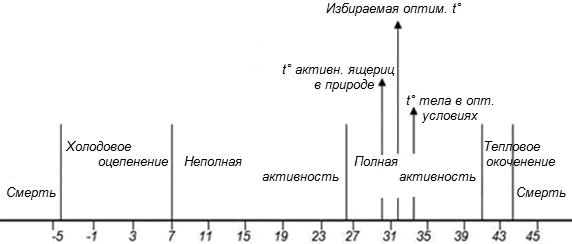
Температурные условия существования L. agilis (по: Либерман, Покровская, 1943; рис. 7 на стр. 251)
Temperature conditions of existence of L. agilis (Liberman, Pokrovskaya, 1943; fig. 7 on p. 251)
4. Выделение полной активности имеет и важное методическое значение, поскольку сам диапазон температур «полной активности» (а не просто абстрактной «активности») является важным термобиологическим показателем, а избираемую температуру (предпочитаемую, оптимальную) у таких животных, как прыткие ящерицы (термостабильно активные рептилии – Черлин, 2014), можно получить только из совокупности данных, полученных при полной активности, на что авторы также специально указали впервые.
5. Авторы определили такой важный показатель, как температура тела ящериц при полной активности. В дальнейшем все исследователи стали применять термин «добровольная температура», окончательно укоренившийся после публикации статьи Р. Б. Коула и Ч. М. Богерта (Cowles, Bogert, 1944). Следует обратить внимание на то, что в статье Р. Б. Коула и Ч. М. Богерта понятие «добровольная температура» было определено и применено очень корректно. Хотя эти авторы и не выделяли статусы активности, но они специально оговаривали на стр. 277, что «добровольные температуры» – это диапазон температур тела, который регистрируется после того, как ящерицы нагрелись и когда они уже начали «полную активность» (нормальную активность, «рутинную» активность), хотя этим последним словосочетаниям они особого значения и отдельного определения далее не давали. С. С. Либерман и Н. В. Покровская, напротив, изначально сконцентрировали внимание именно на «ключевом» факторе – на состоянии полной активности. Таким образом, смысловое значение словосочетания «температура тела при полной активности», использованное С. С. Либерман и Н. В. Покровской, и термин «добровольная (voluntary) температура», примененный Р. Б. Коулом и Ч. М. Богертом, на самом деле по сути идентичны, но невнимание их последователей к понятию «полная активность» привело в дальнейшем ко множеству методических неточностей и ошибок. В результате термин «добровольная температура» Р. Б. Коула и Ч. М. Богерта стал не совсем четким, его значение часто определялось с ошибками, и он оказался намного более «расплывчатым», чем «температура тела при полной активности» С. С. Либерман и Н. В. Покровской (хотя этот последний термин вообще у последующих исследователей потерялся и забылся). Тем не менее именно термин «добровольная температура» окончательно утвердился после публикации Р. Б. Коула и Ч. М. Богерта.
6. Очень важно, что С. С. Либерман и Н. В. Покровская четко поняли и указали на то, что после того как прыткие ящерицы переходят в состояние полной активности, «температура ящерицы, несмотря на теснейшую зависимость от температуры среды, остается относительно постоянной, составляя в период активности 30–33°» (стр. 248). Другими словами, они впервые отметили, что при полной активности температура тела прытких ящериц почти постоянно удерживается в узком и стабильном диапазоне на достаточно высоком уровне. Это показывает, что они четко поняли и проиллюстрировали своим конкретным экспериментальным материалом смысл и значение как самого понятия полной активности, так и диапазона температур полной активности.
7. С. С. Либерман и Н. В. Покровская также определили избираемую (или «оптимальную», как они ее называли) температуру тела у ящериц (29–33°). Еще раз повторюсь, что они впервые и вполне конкретно указали на два очень важных методических обстоятельства: 1) избираемую («оптимальную») температуру тела у прытких ящериц можно определять только из совокупности температур тела при полной активности; 2) для выявления диапазона предпочитаемых (оптимальных) температур более всего подходит не средняя арифметическая из совокупности данных по температурам тела при встречах ящериц в состоянии полной активности, а мода (или модальный класс) этой совокупности. Такой методический прием был впервые применен, по замечанию самих авторов, их научным руководителем А. М. Сергеевым при определении оптимальной температуры тела у Lacerta saxicola (пример приведен в статье С. С. Либерман и Н. В. Покровской на рис. 1, стр. 249). Применение моды, как биологически более обоснованного, чем средняя арифметическая, метода статистической обработки при определении предпочитаемых температур (в частности, у рыб), использовали и другие авторы (Рюмин, 1939; Ивлев, 1958; McCauley, 1977; Reynolds, Casterlin, 1978). Это важно, поскольку предпочитаемые («оптимальные») температуры, в частности, у прытких ящериц, – это уровень (или диапазон) температур тела, который наиболее часто встречается у них (избирается ими) в состоянии полной активности. Для описания этой характеристики, т. е. области совокупности с максимально плотным распределением данных, более всего подходит именно мода (или модальный класс), а не средняя арифметическая величина (Черлин, 1010, 2012, 2013, 2014).
8. Как значимые показатели температурной выносливости у прытких ящериц С. С. Либерман и Н. В. Покровская выбрали следующие: для холодовой выносливости – холодовое оцепенение (примерно +7.5°) и смерть (примерно –4.9°); для тепловой выносливости – тепловое окоченение (+41°) и смерть (+44.5°). При этом холодовое оцепенение и тепловое окоченение – состояния обратимые, т. е. если ящериц перенести в среду с нормальной температурой, то через короткое время температура их тела вернется в допустимые границы и ящерицы продолжат вести обычную активность. По всей видимости, показатели «холодовое оцепенение» и «тепловое окоченение» могут соответствовать применяющимся сейчас показателям «критический минимум» и «критический максимум», т. е. температурам холодового и теплового шока.
Большой интерес представляет и вторая часть статьи, касающаяся размножения и развития яиц прытких ящериц. Важен даже сам набор основных проблем, рассматривающихся в статье: условия откладки яиц, число и строение яиц, условия развития яиц, роль оболочки яиц. Проведенные опыты ответили, в частности, на два важных вопроса: «1) выбирает ли самка для откладки яиц определенные места и 2) насколько условия мест, избранных для откладки яиц, соответствуют оптимальным условиям развития последних» (стр. 251). Для яйцекладки самки выбирают грунт с влажным песком (в среднем это примерно 16.21 %). В сухой грунт яйца вообще не откладываются. Авторы доказывают, что влажность грунта является одним из условий, стимулирующих кладку. Также в статье определены условия влажности, необходимые для инкубации, формы поступления воды в яйца через разные типы его оболочек, необходимое количество влаги для развития яиц на разных стадиях. Это замечательная, очень корректная в методическом и практическом плане работа, особенно при учете того, что выполнена она была в конце тридцатых – начале сороковых годов XX века!
По моей просьбе заведующая отделом герпетологии Зоологического музея МГУ В. Ф. Орлова произвела в архивах Московского государственного университета поиск данных об авторах статьи. В книге об истории Зоологического музея МГУ (Любарский, 2009) на стр. 99 есть фотография 1941 г., на которой среди известных фигур кафедры зоологии позвоночных изображены С. С. Либерман и Н. В. Покровская. К сожалению, о Н. В. Покровской более никаких материалов обнаружить не удалось. А вот о С. С. Либерман кое-что нашлось. Серафима Соломоновна Либерман родилась 20 ноября 1919 г. В книге о жизни А. Н. Формозова (1980) на стр. 105 есть следующая информация: «Студенты старались как-то облегчить жизнь своего профессора. Жившая по соседству, на Остоженке, Серафима Соломоновна Либерман приходила к нему пилить дрова, а он в другие дни помогал в этом ей». Это относится к 1942–1943 гг. Впоследствии С. С. Либерман ушла в медицину, стала профессором, опубликовала с соавторами ряд книг и статей по лекарственной терапии (Сигидин и др., 1988; Гуськова, Либерман и др., 1982; Гуськова, Либерман, 1997; и др.). Я искренне благодарю В. Ф. Орлову за помощь в поиске этого исторического материала.
Отдельно несколько слов хотелось бы сказать о руководителе работы С. С. Либерман и Н. В. Покровской – замечательном биологе Алексее Михайловиче Сергееве. Он родился в Москве в 1913 г. Ходил в кружок юных биологов Московского зоопарка, где выполнил свою первую научную работу. Поступил на биологический факультет МГУ на кафедру зоологии позвоночных. Еще в студенческие годы участвовал во многих научных экспедициях. Окончил аспирантуру и представил к защите кандидатскую диссертацию, которая Ученым советом единодушно была оценена как докторская. Свободно владел тремя европейскими языками.
В 1939 г. участвовал в освобождении Западной Украины от белополяков, а в первые же дни Великой Отечественной войны прямо с полевой практики отправился добровольцем на фронт (хотя, как доктор наук, мог воспользоваться бронью). Воевал под Киевом, попал в окружение, но вывел из него свой взвод. По навету был арестован и находился в Усольском фильтрационном лагере, где в 1943 г. в возрасте 30 лет покончил с собой (Васильев, Васильева, 2012).
Он прожил короткую, но яркую жизнь. Его вклад в науку оценивается очень высоко. По работе С. С. Либерман и Н. В. Покровской, которой он руководил, его научный талант и потенциал не вызывают никаких сомнений. Его научные идеи, в частности в области биологии рептилий (понятия «термобиология» тогда еще не было), явно опережали все, что на тот момент было сделано за границей.
***
В заключении моей статьи хотелось бы обратить внимание на два важных обстоятельства. Во-первых, работа С. С. Либерман и Н. В. Покровской, выполненная под научным руководством А. М. Сергеева, – очень важная и знаковая. В ней впервые авторы обратили внимание и определили чрезвычайно важные детали, термины и понятия, продемонстрировали важные теоретические положения и методические подходы, которые, к сожалению, в дальнейшем учеными забылись: это статусы активности (полное и неполное активные состояния), температура полной активности, методика определения оптимальной температуры и др. Недостаточное внимание к этим вопросам привело, к сожалению, к серьезным некорректностям и ошибкам в дальнейшем развитии термобиологии рептилий, о чем я уже неоднократно подробно писал в своих работах (Черлин, 2012, 2013, 2014). Во-вторых, и это хотелось бы подчеркнуть особо, статья С. С. Либерман и Н. В. Покровской по праву может считаться первой основополагающей в области термобиологии рептилий, поскольку была опубликована в 1943 г., т. е. на год раньше статьи Р. Б. Коула и Ч. М. Богерта, от которой сейчас отсчитывается начало этого направления научных исследований, а по научной значимости она ничуть ей не уступает.
Библиография
Васильев Б. Д., Васильева А. Б. Жизнь и научное наследие Алексея Михайловича Сергеева (1912–1943) // Зоологический журнал. 2012. Т. 91. № 11. С. 1283–1290.
Гуськова Т. А., Либерман С. С. Хронобиологические аспекты лекарственной токсикологии (обзор состояния проблемы) // Токсикологический вестник. 1997. № 2. С. 2–7.
Гуськова Т. А., Либерман С. С. и др. Результаты экспериментального и клинического изучения преднизолонгемисукцината – отечественного водорастворимого препарата преднизолона // ЭЙ. Новые лекарственные препараты. 1982. № 7. С. 17–20.
Ивлев В. С. Эколого-физиологический анализ распределения рыб в градиентных условиях среды // Тр. совещ. Ихтиол. комис. АН СССР. Вып. 8. М., 1958. С. 288–296.
Либерман С. С., Покровская И. В. Материалы по экологии прыткой ящерицы // Зоол. журн. 1943. Т. 22. № 2. С. 247–256.
Любарский Г. И. История Зоологического музея МГУ: Идеи, люди, структуры . М.: КМК, 2009. 744 с.
Родионов В. Некоторые данные по газообмену у рептилий в состоянии переохлаждения // Бюлл. Моск. об-ва испытателей природы. Отдел биол. 1938. Т. 47. № 2. С. 182–187.
Рюмин А. В. Температурная чувствительность позвоночных животных и биологический путь происхождения теплокровных форм // Сб. студ. научн. работ МГУ. М., 1939. Вып. 6. С. 55–84.
Сергеев А М. Температура пресмыкающихся в естественных условиях // Докл. АН СССР. 1939. Т. XXII. № 1. С. 49–52.
Сигидин Я. А., Шварц Г. Я., Арзамасцев А. П., Либерман С. С. Лекарственная терапия воспалительных процессов . М.: Медицина, 1988. 240 с.
Стрельников И. Д. Свет как фактор в экологии животных. Статья первая. Действие солнечной радиации на температуру тела некоторых пойкилотермных животных // Известия Научного института имени П. Ф. Лесгафта. 1934. Том XVII–XVIII. С. 313–372.
Формозов А. А. Александр Николаевич Формозов (1899–1973) / отв. ред. д-р геогр. наук А. А. Насимович. М.: Наука, 1980. 152 с.
Черлин В. А. Термобиология рептилий. Общие сведения и методы исследований (руководство) . СПб.: Русско-Балтийский информационный центр «БЛИЦ», 2010. 124 с.
Черлин В. А. Термобиология рептилий. Общая концепция . СПб.: Русско-Балтийский информационный центр « БЛИЦ», 2012. 361 с.
Черлин В. А. Сложности и возможные ошибки при полевых исследованиях по термобиологии рептилий // Современная герпетология: проблемы и пути их решения: Статьи по материалам докладов Первой международной молодежной конференции герпетологов России и сопредельных стран (Санкт-Петербург, Россия, 25–27 ноября 2013 г.) / Зоологический институт РАН. СПб., 2013. С. 32–39.
Черлин В. А. Рептилии: температура и экология . Saarbrücken: Lambert Academic Publishing, 2014. 442 с.
Черномордиков В. В. О температурных реакциях пресмыкающихся // Зоол. журн. 1943. Т. 22. № 5. С. 274–279.
Agassiz L., Gould A. A. Principles of zoology: Touching the structure, development, distribution and natural arrangement of the races of animals, living and extinct. Part I.Comparative physiology. Sheldon, Blakeman and Co., New York, 1856. 250 p.
Amat F., Llorente G. A., Carretero M. A. Reproductive cycle of the sand lizard (Lacerta agilis) in its southwestern range // Amphibia-Reptilia. 2000. Vol. 21. P. 463–476.
Cowles R. B., Bogert C. M. Preliminary study of the thermal requirements of desert reptiles // Bulletin of the American Museumof Natural History. 1944. № 83. P. 261–296.
Crawford E. C., Palomeque J., Barber B. J. Aphysiological basis for head–body temperature differences in a panting lizard. // Comp. Biochem. Physiol. 1977. 56A, 2:161–163.
Hall F. G. The vital limit of exsiccation of certain animals // Biol. Bull. 1922. Vol. 42. P. 31–51.
Herter K. Die Vorzungstemperaturen bel Landtieren. Naturwissenschaften. Berlin, 1941. Vol. 29. P. 155–164.
McCauley R. W. Laboratory methods for determining temperature preference // Temperature preference studies in environmental impact assessments: an overview with procedural recommendations. Proceed. Symp. and Panel Discuss. Northeast Fish and Wildlife Conf. (Northeast Division, Amer. Fish Soc. Hershey. Pa. April 27. 1976 / J. Fish Res. BoardCan. 1977. Vol. 34. № 5. P. 749–452 (Ed. Richards E. P., Reynolds W. W., McCauley R. W.).
Mosauer W., Lazier E. L. Death from insolation in desert snakes // Copeia. 1933. № 3. P. 149.
Reynolds W. W., Casterlin M. E. Ontogenetic change in preferred temperature and diel activity of the bullhead, Ictahurus natalis // Comp. Biochem. Physiol. 1978. Vol. A59. № 4. P. 409–411.
Weese A. O. An experimental study of the reactions of the horned lizard, Phryrlosoma modestum Gir., a reptile of the semi–arid desert // Biol. Bull. 1917. № 32. P. 98–116.

 © 2011 - 2026
© 2011 - 2026