



Введение
Настоящая работа – обобщение данных о фитопланктоне Выборгского залива в период интенсивного антропогенного воздействия в XXI в. и оценка его структуры, количественного развития и пространственного распределения.
Выборгский залив – самый крупный (площадь 450 км²) из заливов второго порядка восточной части Финского залива, вдается в сушу в северо-восточном направлении на 60 км, его береговая линия сильно изрезана, акватория состоит из ряда мелководных бухт с большим количеством островов.
Акватория Выборгского залива делится на три части: верхнюю, среднюю и нижнюю (таблица).
Характеристики частей Выборгского залива (Терешенкова, 2006; Дроздов, Коробков, 2010)
| Часть | Площадь, км² | Преобладающие глубины, м | Доля островов в общей площади, % | Прозрачность воды в литорали, м | Соленость в верхнем горизонте, ‰ |
| Верхняя | 140 | 3.5–4.0 | 21 | до 1.0 | 0–3.0 |
| Средняя | 160 | 10 | 16 | до 2.0 | 1.5–2.0 |
| Нижняя | 120 | 25–30 | 2 | до 3.5 | 2.3–4.0 |
Верхняя часть занимает акваторию от места впадения Сайменского канала до разреза пос. Советский – пос. Подборовье (рис. 1). Сложность очертаний береговой линии обусловила замедленный водообмен акватории этого района с другими частями залива. Здесь расположен подходной фарватер Сайменского канала, связывающий оз. Сайма с Финским заливом, и верхняя часть залива испытывает значительное распресняющее воздействие от поступления его водных масс.
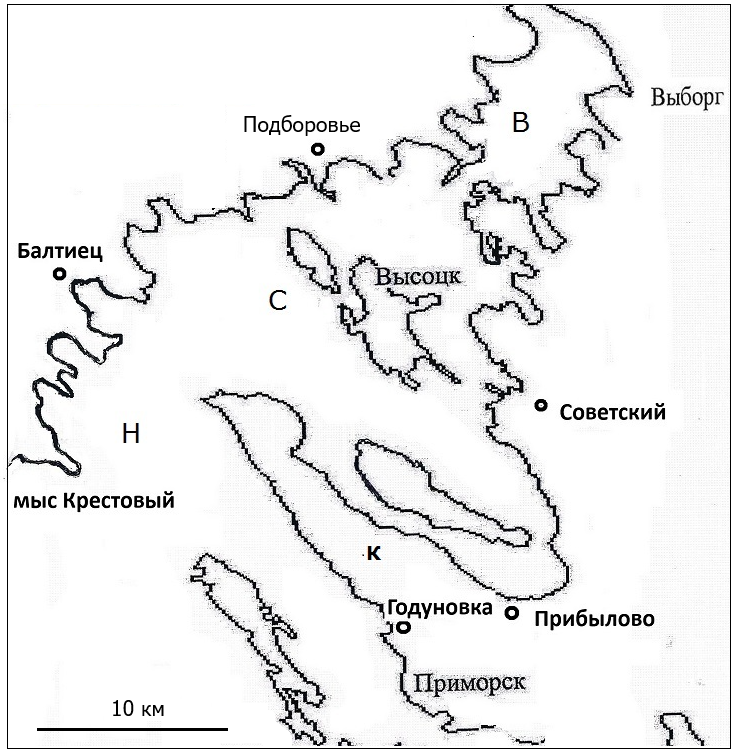
Рис. 1. Схема Выборгского залива Балтийского моря: В – верхняя часть; С – средняя часть; Н – нижняя часть; К – полуостров Киперорт
Fig. 1. Cheme of the Vyborg Bay of the Baltic Sea: В – upper part; С – middle part; Н – lower part; K – Kiperort Peninsula
Средняя часть распространяется на юг до разреза пос. Прибылово – мыс севернее залива Балтиец (см. рис. 1), что совпадает с наиболее узкой частью акватории залива (ширина 6 км). Полуостров Киперорт, вытянутый в северном направлении, перегораживает залив и ограничивает проникновение солоноватых вод из нижней части в среднюю. В то же время пресный сток здесь меньше, чем в верхней части, и соленость несколько выше (см. таблицу).
Нижняя часть ограничена от открытой акватории восточной части Финского залива разрезом пос. Годуновка – мыс Крестовый. Доля акватории, занятая островами, минимальна. В наибольшей степени испытывает влияние вод открытой части Финского залива.
Режим солености Выборгского залива не стабилен и определяется сгонно-нагонными перемещениями осолоненных вод Финского залива, а также распресняющим влиянием Сайменского канала и многочисленных притоков. В целом залив характеризуется соленостью от 0 до 4 ‰ и может быть отнесен к акваториям эстуарного типа.
Морфология и гидрологические особенности залива обусловливают значительную гетерогенность условий обитания гидробионтов, в частности планктонных водорослей, в то же время его фитопланктон недостаточно изучен по сравнению с другими заливами второго порядка восточной части Финского залива, первые данные эпизодичны и относятся к концу XX в. (Макарова, 1997; Терешенкова, 2006; Экосистема…, 2008).
Единственная станция государственного мониторинга расположена в нижней части Выборгского залива на границе с глубоководным районом Финского залива, данные о фитопланктоне основной его акватории не имеют систематического характера и получены преимущественно в связи с мониторинговыми исследованиями возможного влияния на фитопланктон крупномасштабных гидротехнических работ, проводившихся в XXI в. в различных частях акватории залива. Из наиболее масштабных можно упомянуть взрывные работы по углублению подходного фарватера Сайменского канала, работы по реконструкции причалов порта Выборг, длительные работы по развитию терминалов морского торгового порта Высоцк и другие работы по дноуглублению и помещению грунтов в подводные отвалы.
Представленная работа – первая попытка обобщения данных о фитопланктоне всей акватории Выборгского залива на основании наблюдений в разные периоды его сезонной сукцессии с начала XXI в.
Материалы
Материалом для работы послужили результаты исследования фитопланктона залива в 2018–2024 гг. Пробы отбирали на станции государственного мониторинга в нижней части залива и на участках его акватории, связанных с проведением гидротехнических работ и эксплуатацией портовых сооружений, а также фоновых станциях.
Наряду с собственными, использованы архивные данные лаборатории гидробиологии Санкт-Петербургского филиала ВНИРО за 2000–2017 гг., отобранные и обработанные по аналогичной методике.
Проанализированы результаты обработки 160 проб фитопланктона.
Методы
Пробы воды отбирали батометром Паталаса в трофогенном слое (до глубины, соответствующей утроенной прозрачности по диску Секки) через каждый метр. Взятую в равных количествах из каждого слоя воду перемешивали, помещали в пластиковую емкость, фиксировали раствором Люголя с добавлением уксусной кислоты и формалина, концентрировали фильтрационным методом с использованием мембранных фильтров с диаметром пор 0.8–1.2 мкм. Фитопланктоном считали всю фотосинтезирующую часть планктона, подсчет его организмов проводили в счетной камере объемом 0.01 мл, биомассу определяли счетно-объемным методом на основании размеров водорослей, измеренных в процессе камеральной обработки (Методика изучения..., 1975; Методические рекомендации…, 1981). Таксономический состав фитопланктона определяли в процессе обработки количественных проб. Доминирующими считали систематические группы или отдельные таксоны рангом ниже рода, составляющие не менее 10 % общей численности или биомассы фитопланктона. Статистическую обработку проводили в программе MS Excel.
Результаты
Фитопланктон Выборгского залива в 2000–2024 гг. формировали водоросли 8 систематических групп: Cyanobacteria (цианобактерии), Chrysophyceae (золотистые), Bacillariophyta (диатомовые), Cryptophyceae (криптофитовые), Dinophyceae (динофитовые), Euglenophyceae (эвгленовые), Chlorophyta (зеленые), Charophyta (харовые). В составе фитопланктона отмечалась также фотосинтезирующая инфузория Myrionecta rubra Jank. (Ciliophora).
В безледный период 2000–2024 гг. диапазон изменения биомассы фитопланктона залива составил 0.12–23.16 г/м3, т. е. ее вариативность достигала двух порядков, численность 1643–533270 млн кл/м3 изменялась более чем на три порядка.
Биомасса весеннего фитопланктона изменялась от 0.23 до 12.18 г/м3. В нижней части залива численно преобладали цианобактерии и зеленые водоросли, преимущественно рода Monoraphidium. Основу биомассы составляли диатомовые и динофлагеллаты, среди которых основными доминантами были Diatoma tenuis C. Ag. и Apocalathium aciculiferum (Lemm) Crav. Daugb. Maestr. & Calad., в отдельные годы – эвгленовые, представленные Eutreptiella gymnastica Tronds., и фотосинтезирующая инфузория Myrionecta rubra, доля которой в общей биомассе в начале июня 2020 г. достигала 43 %. В целом для весенних данных более половины биомассы составляли диатомовые, более трети – динофлагеллаты.
Весенний фитопланктон средней части был представлен преимущественно диатомовыми, составлявшими до 90 % его биомассы, основными доминантами были D. tenuis, Tabellaria fenestratа (Lyngb.) Kütz., Skeletоnema subsalsum (A. Cleve) Bethge, S. сostatum (Grev.) Cleve. В верхней части численно и по биомассе также преобладали диатомовые, представители родов Aulacoseira, Cyclotella и T. fenestrata. В состав доминирующего комплекса входили криптофитовые и эвгленовые (рис. 2).
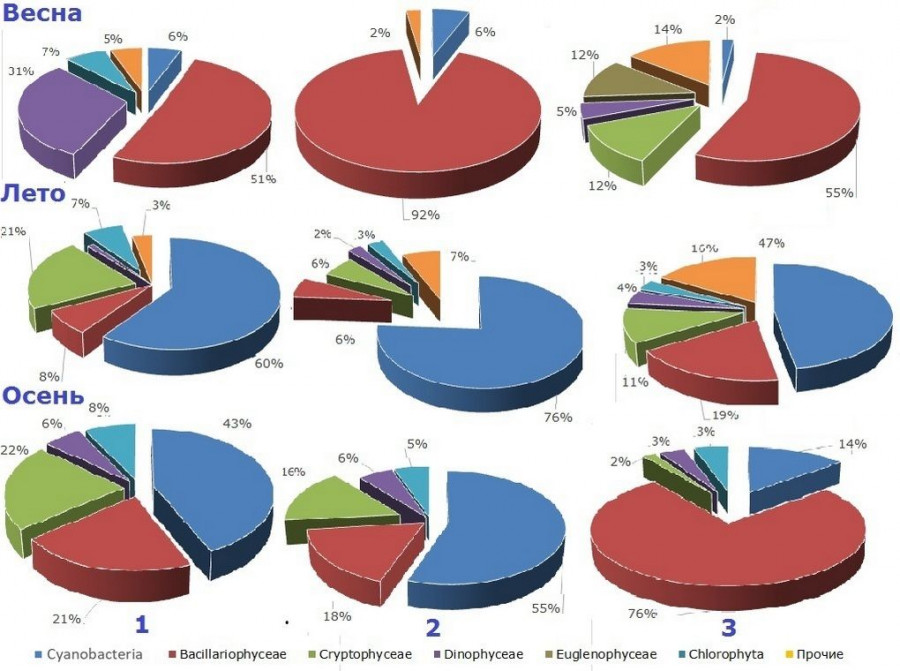
Рис. 2. Доля водорослей основных систематических групп в биомассе фитопланктона различных частей Выборгского залива: 1 – нижняя, 2 – средняя, 3 – верхняя
Fig. 2.The proportion of algae of the main systematic groups in the phytoplankton biomass of various parts of the Vyborg Bay : 1 – lower part, 2 – middle part, 3 – upper part
Летом биомасса изменялась от 0.33 до 17.66 г/м3, в нижнем районе ее основу составляли цианобактерии, доминировали Aphanizomenon flosaquae Ralfs ex Born. & Flah., Planktothrix agardhii (Gom.) Anagn. & Kom., Pseudanabaena limnetica (Lemm.) Kom., в доминирующий по биомассе комплекс также входили криптофитовые (виды р. Cryptomonas), диатомовые и зеленые водоросли (см. рис. 2).
Цианобактерии составляли основу численности и биомассы фитопланктона в среднем районе, доминировали Aphanizomenon gracile Lemm., A. flosaquae, P. agardhii, P. limnetica.
В верхней части залива доминирующий комплекс фитопланктона был более разнообразен. Наряду с цианобактериями значительную долю биомассы составляли диатомовые и криптофитовые (см. рис. 2), основные доминанты – P. agardhii, Limnothrix planktonica (Wolosz.) Meff., Planktolyngbya subtilis (West) Anagn. & Kom, A. gracile, водоросли рода Cryptomonas, T. fenestrata, Cyclotella sp., Aulacoseira sp.
Осенью биомасса фитопланктона залива составляла от 0.12 до 3.35 г/м3, количественные показатели развития фитопланктона его нижней части определяли цианобактерии (P. agardhii, P. limnetica, A. flosaquae). По биомассе, кроме цианобактерий, доминировали диатомовые и криптофитовые (водоросли родов Fragilaria и Cryptomonas).
В среднем участке основу численности, как и летом, составляли цианобактерии – P. agardhii, P. limnetica, A. flosaquae. По биомассе наряду с ними доминирующий комплекс формировали диатомовые c преобладанием Aulacoseira islandica (O. Müll.) Sim. и S. costatum и криптомонады – представители рода Cryptomonas, доля которых в биомассе к концу осени увеличивалась.
В верхнем участке при доминировании по численности цианобактерий (преимущественно L. planktonica и P. subtilis) диатомовые – c преобладанием Aulacoseira italica, (Eher.) Sim., A. granulata (Eher.) Sim., S. subsalsum составляли основу биомассы (см. рис. 2).
Специфику различных частей залива наглядно иллюстрирует характер изменения биомассы фитопланктона в сезонном аспекте: верхняя часть характеризуется максимальными величинами в середине лета, тогда как средняя и нижняя – в весенний период, что характерно для восточной части Финского залива. В целом весной диатомовые доминируют во всех частях залива, в нижней в состав доминирующего комплекса входят динофлагеллаты. Средние величины весенних биомасс в нижней и верхней частях залива были сходны, а в средней части почти в 5 раз выше, чем в остальных двух. Летние биомассы в верхней части залива были в среднем в 3 раза выше, чем в нижней, а в средней – более чем в 2 раза выше, чем в верхней (рис. 3).
Минимальные биомассы во всех частях залива отмечены осенью (см. рис. 3), при этом в нижней части доминировали цианобактерии, свойственные летнему фитопланктону, что подтверждает представление об отсутствии специфического осеннего комплекса планктонных водорослей в восточной части Финского залива. В то же время в верхней части основу биомассы осеннего фитопланктона формировали диатомовые (см. рис. 3), что характерно для фитопланктона озерно-речного комплекса и, вероятно, обусловлено влиянием вод Сайменского канала. В осеннем фитопланктоне средней части залива доминировали цианобактерии и (с меньшей долей в биомассе) диатомовые.
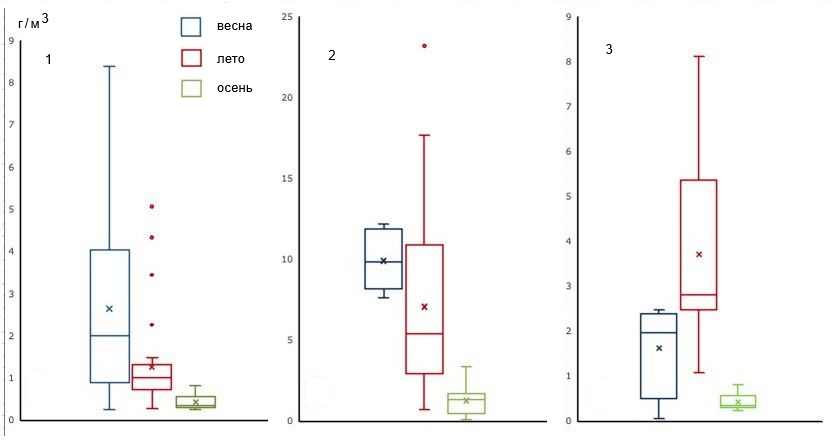
Рис. 3. Сезонные изменения средних биомасс фитопланктона Выборгского залива: 1 – нижняя часть, станция государственного мониторинга (2000–2024 гг.); 2 – средняя часть (2010–2023 гг.); 3 – верхняя часть (2001–2022 гг.)
Fig. 3. Seasonal changes in average phytoplankton biomass of the Vyborg Bay: 1 – lower part, state monitoring station (2000–2024); 2 – middle part (2010–2023); 3 – upper part (2001–2022)
Влияние значительной изрезанности береговой линии и наличия большого количества островов на формирование разнообразных условий вегетации фитопланктона наглядно демонстрирует его распределение в средней части залива в районе порта Высоцк в августе и октябре 2019 г. (рис. 4), когда на станциях, прилегающих к открытой части залива, основу биомассы составляли цианобактерии, а в бухте Ключевской, при максимальных величинах биомассы, фитопланктон формировали диатомовые, зеленые, цианобактерии, криптофитовые, золотистые, в бухте Малой Пихтовой, при минимальных величинах биомассы, основу ее составляли криптомонады. Значительно отличался от открытой части залива фитопланктон этих бухт и осенью – как величинами биомассы, так и значительной долей золотистых водорослей (см. рис. 4).
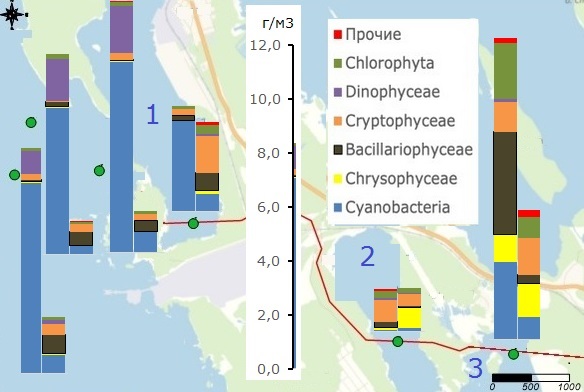
Рис. 4. Биомасса фитопланктона в средней части Выборгского залива (район порта Высоцк) в августе и октябре 2019 г.: 1 – бухта Большая Пихтовая; 2 – бухта Малая Пихтовая; 3 – бухта Ключевская
Fig. 4. Phytoplankton biomass in the middle part of Vyborg Bay (Vysotsk port area) in August and October 2019: 1 – Bolshaya Pikhtovaya Bay; 2 – Malaya Pikhtovaya Bay; 3 – Klyuchevskaya Bay
Межгодовые изменения величины биомассы летнего фитопланктона в нижней части залива, по данным станции государственного мониторинга, показали, что максимальная и минимальная величины биомасс здесь различались на порядок, наименьшие были характерны для периода 2010–2020 гг., а наибольшие отмечены в 2022–2023 гг. Состав доминирующего комплекса во все годы наблюдений был сходен и формировался преимущественно цианобактериями, диатомовыми, криптофитовыми, в отдельные годы – зелеными водорослями (рис. 5.1).
В районе порта Высоцк в средней части залива в 2008–2024 гг. состав доминирующего комплекса летнего фитопланктона был аналогичен отмеченному в нижней части при значительно превышающих его величинах биомассы (рис. 5.2). Различие минимальных и максимальных средних для исследованных акваторий величин было трехкратным.
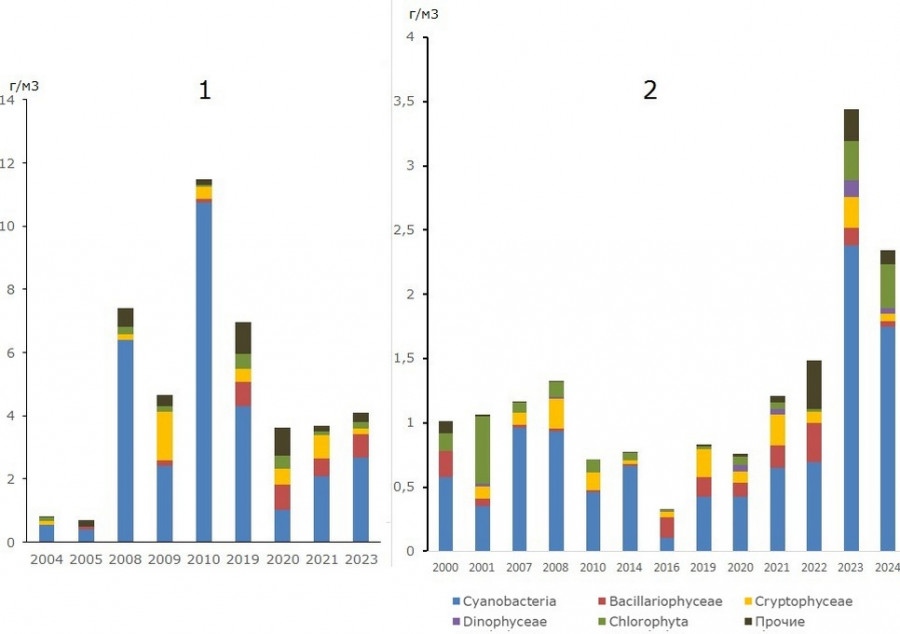
Рис. 5. Межгодовые изменения биомассы летнего фитопланктона: 1 – средней части Выборгского залива (район порта Высоцк); 2 – нижней части залива (станция государственного мониторинга)
Fig. 5. Interannual changes in summer phytoplankton biomass: 1 – middle part of Vyborg Bay (Vysotsk port area); 2 – lower part of the bay (state monitoring station)
Обсуждение
По результам исследований можно отметить, что состав доминирующего комплекса фитопланктона Выборгского залива в XXI в. не претерпел существенных изменений и характеризуется преобладанием цианопрокариот, диатомовых и криптофитовых со сходным с отмеченным в конце XX в. составом доминантов. В качестве отличий можно отметить доминирование весной – в начале лета в фитопланктоне нижней части залива, находящейся в области влияния вод глубоководной части Финского залива, эвгленовой Eutreptiella gymnastica и фотосинтезирующей (за счет клептопластии) инфузории Myrionecta rubra – компонента доминирующих комплексов фитопланктона в западных акваториях Финского залива (The Gulf of Finland…, 2016).
По средним за безледный период величинам биомассы (1.24 г/м3 в нижнем и 2.87 г/м3 – в верхнем) обе акватории характеризуются как мезотрофные (Трифонова, 1990), однако характер ее динамики наглядно демонстрирует изменения сообщества фитопланктона по мере повышения трофности акватории. В нижнем районе она близка к характерной для олиготрофно-мезотрофных – весенний максимум и значительное снижение биомассы летом, а в верхнем – значительное превышение летней биомассы над весенней, что характерно для мезотрофных водоемов, причем разница между летними и весенними величинами биомассы у средних величин, характерных для мезотрофного водоема, демонстрирует степень его олиготрофно-эвтрофной сукцессии. Более корректно такие различия в трофности акваторий отражены в классификации, по которой нижний участок характеризуется как α-мезотрофный, а верхний – как β-мезотрофный (Китаев, 2007).
Средняя за безледный период величина биомассы фитопланктона средней части залива составила 6.83 г/м3 и характеризует его акваторию как эвтрофную. Средний район выделялся наибольшими величинами биомассы, поскольку с юга испытывал меньшее по сравнению с нижней частью влияние вод Финского залива, а с севера – меньшее по сравнению с верхней частью воздействие поступления вод Сайменского канала.
По результатам исследований летнего фитопланктона нижней части Сайменского канала (от шлюза Пялли до шлюза Брусничное), проведенных в 2021 г., его биомасса при доминировании диатомовых (T. fenestrsta, Fragilaria crotonensis Kitt., Cyclotella sp.) в среднем составила 0.842 г/м3. Это подтверждает, что именно поступление водных масс канала снижает продуктивность фитопланктона верхней части залива, несмотря на обширные мелководья и другие благоприятные для развития фитопланктона условия.
Учитывая большую гетерогенность условий в акваториях верхней и средней частей залива из-за сложности морфологии его берегов, можно предположить, что обобщенная характеристика трофности этих акваторий является в значительной степени условной и зависит от местоположения станций отбора проб.
Сравнение результатов исследования фитопланктона Выборгского залива с данными 90-х гг. XX в. (Терешенкова, 2006) показало, что, несмотря на масштабные гидротехнические работы, проводившиеся на различных частях акватории Выборгского залива, в XXI в. не произошло существенных изменений как структуры доминирующего комплекса фитопланктона, так и количественных показателей его развития, что вполне объяснимо значительным восстановительным потенциалом этого сообщества, слагаемого организмами, при благоприятных условиях быстро наращивающими численность своих популяций. Выборгский залив в XXI в., как и ранее (Макарова, 1997; Лаврентьева и др., 1999; Терешенкова, 2006; Экосистема…, 2008), остается наиболее продуктивной акваторией восточной части Финского залива.
Заключение
Значительная вариабельность структуры и количественного развития фитопланктона в различных частях его акватории обусловлена морфологией берегов и гидрологическими особенностями залива, определяющими гетерогенность условий формирования фитопланктона.
Наибольшие величины биомассы фитопланктона отмечаются преимущественно в средней части, ее ориентировочно можно охарактеризовать как эвтрофную. Верхнюю часть залива, принимающую воды Сайменского канала, и нижнюю часть, прилегающую к глубоководному району восточной части Выборгского залива, – как мезотрофные.
Ядро планктонных альгоценозов залива сходно с таковым остальной акватории восточной части Финского залива и в XXI в. не претерпело значительных изменений.
Количественные показатели развития фитопланктона залива и его структура в XXI в. сходны с отмечавшимися в конце XX в. Проводившиеся в XXI в. крупномасштабные гидротехнические работы не привели к изменениям как структуры фитопланктона, так и количественных показателей его развития.
Библиография
Дроздов В. В., Коробков А. В. Влияние процесса дноуглубительных работ на экологическое состояние акваторий Выборгского залива // Ученые записки РГГМУ. 2010. № 12. С. 80–96.
Китаев С. П. Основы лимнологии для гидробиологов и ихтиологов . Петрозаводск: Карельский научный центр РАН, 2007. 395 с.
Лаврентьева Г. М., Мещерякова С. В., Мицкевич О. И., Огородникова В. А., Суслопарова О. Н., Терешенкова Т. В. Гидробиологическая характеристика Выборгского залива, пролива Бъеркезунд, бухты Батарейной и Лужской губы (восточная часть Финского залива) // Финский залив в условиях антропогенного воздействия. СПб., 1999. С. 211–256.
Методика изучения биогеоценозов внутренних водоемов . М.: Наука, 1975. 241 с.
Методические рекомендации по сбору и обработке материалов при гидробиологических исследованиях на пресноводных водоемах. Фитопланктон и его продукция . Л., 1981. 32 с.
Макарова С. В. Видовой состав и количественные характеристики фитопланктона // Экосистемные модели. Оценка современного состояния Финского залива. СПб.: Гидрометеоиздат, 1997. С. 354–365.
Терешенкова Т. В. Сравнительная характеристика летнего фитопланктона Выборгского залива, Лужской и Копорской губ, пролива Бъеркезунд и Мелководного района восточной части Финского залива // Сборник научных трудов ГосНИОРХ. 2006. Вып. 331. Т. 1. С. 37–85.
Трифонова И. С. Экология и сукцессия озерного фитопланктона . Л.: Наука, 1990. 184 с.
Экосистема эстуария реки Невы: биологическое разнообразие и экологические проблемы . М.: Товарищество научных изданий КМК, 2008. 477 с.
The Gulf of Finland assessment // Reports of the Finnish Environment institute. 2016. Vol. 27. 363 p.
Благодарности
С благодарностью за годы совместной работы и доброй памятью преждевременно ушедшей от нас в 2017 г. Терешенковой Татьяне Викторовне, которой получена основная часть использованных в этой работе архивных данных лаборатории гидробиологии СПб филиала ВНИРО (ГосНИОРХ им. Л. С. Берга).

 © 2011 - 2026
© 2011 - 2026