



Введение
Статья посвящена пространственной неоднородности населения земноводных Западной Сибири – обширного природно-экономического региона, к которому относят Западно-Сибирскую равнину и прилежащую с юго-востока часть Алтае-Саянской горной страны с Кузнецко-Салаирской горной областью и российской частью Алтайской горной области (Западная Сибирь, 1963). Рассмотрена изменчивость плотности и видового богатства населения земноводных этой территории, а также дана оценка численности и охарактеризовано распределение всех видов, отмеченных во время проведения учетов.
Приведенная информация имеет не только познавательное, теоретическое, но и прикладное значение, что обусловлено возможностью ее использования при планировании и выполнении мероприятий по сохранению биоразнообразия и мониторинге популяций земноводных, а также при проведении экологической экспертизы проектов, связанных с использованием природных ресурсов региона. Как известно, решение подобных задач подразумевает необходимость отслеживания изменений численности животных в пространстве и во времени, что требует очень больших трудозатрат из-за высокого разнообразия местообитаний и огромных площадей, на которых такие работы следует проводить. Представленные сведения могут быть взяты за основу при проведении подобных работ, что значительно снизит их трудоемкость.
Исследования, результаты которых взяты за основу для нашей статьи, выполнены в рамках направления, получившего название «Факторная зоогеография» (Равкин, Ливанов, 2008). Основная специфика этого направления обусловлена переходом из реального хорологического пространства в типологическое многомерное факторное пространство (в его математическом понимании) при описании и статистическом анализе данных. Главная цель исследования заключалась в выявлении основных трендов в пространственной изменчивости населения земноводных и коррелятивных связей с ведущими факторами среды. Информация о численности земноводных служила основанием для достижения этой цели.
Материалы
Все сведения по обилию земноводных взяты из банка данных Лаборатории зоологического мониторинга ИСиЭЖ СО РАН. Они собраны в результате совместных усилий 60 специалистов в период с 1970 по 1973 и с 1975 по 2022 г., т. е. за 52 года. За это время с помощью канавок и/или заборчиков с ловчими цилиндрами, открытыми, как правило, весь сезон с середины июля до конца августа, выполнен количественный учет земноводных. Суммарно отработано около 33700 цилиндро-суток (ц/с), при этом обследовано 1623 местообитания (биотопа), по своему масштабу и рангу примерно соответствующих таким широко используемым в геоботанике и ландшафтоведении классификационным единицам, как «растительная формация» и «ландшафтное урочище». В полученной выборке преобладают однолетние данные по обилию земноводных, поскольку бóльшая часть биотопов обследована в каждом месте однократно. Все показатели обилия, выраженные количеством особей на 100 ц/с, пересчитаны и на 1 км2 (Равкин, Ливанов, 2008). Бо́льшая часть и предварительный анализ результатов учета земноводных опубликованы (Равкин, Лукьянова, 1976; Борисович и др., 2002; Равкин и др., 2003а, б, в, 2005, 2008, 2020; Цыбулин и др., 2007, 2013; Эпова и др., 2013; Куранова, Ярцев, 2020; часть ссылок см. в перечисленных публикациях). В данной статье впервые представлены итоговые результаты обобщения на региональном уровне – по населению земноводных всей территории Западной Сибири, включая не только равнинную, но и горную часть этого региона. Для этого привлечены все собранные к настоящему времени материалы (рис. 1).
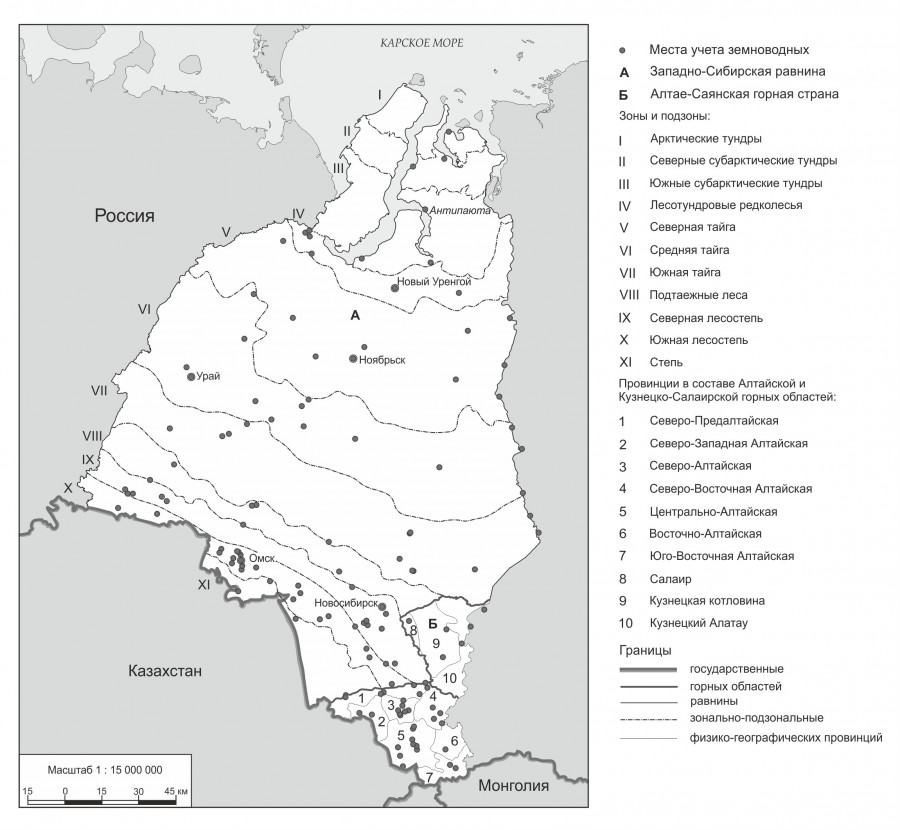
Рис. 1. Места проведения учетов земноводных на территории Западной Сибири
Fig. 1. Amphibian accounting sites in Western Siberia
Методы
Анализ материалов включал поэтапную кластеризацию всей совокупности обследованных сообществ с использованием программы факторной классификации – качественного аналога метода главных компонент (Трофимов, 1976; Равкин, Ливанов, 2008). При этом объединение классифицируемых объектов (пространственных вариантов населения) в группы (типы и подтипы населения) осуществлялось вне зависимости от их местоположения, исключительно по степени сходства-различия между собой, оцениваемого коэффициентом Жаккара (Jaccard, 1902) для количественных признаков (Наумов, 1964). Помимо факторной классификации для кластеризации дополнительно использовали метод К-средних (Миркин, 2011). При этом из состава населения были исключены сеголетки, поскольку их обилие сильно зависит от близости ловчих канавок и заборчиков к локальным водоемам выплода, не отображаемым в масштабе использованных нами карт.
Информативность классификаций, структурных графов и коррелятивных связей между изменчивостью населения земноводных и факторами внешней среды оценивалась с помощью качественной линейной аппроксимации матриц связи (Куперштох и др., 1978) – качественного аналога регрессионной модели. Показателем информативности служила доля исходной дисперсии (квадрат отклонения от среднего по всей матрице) коэффициентов сходства, учтенная отдельно каждым таксоном классификации и всей их совокупностью.
В процессе анализа возникли затруднения при интерпретации получаемых разбиений, что во многом связано со значительно возросшим объемом используемых материалов. Такая проблема неоднократно встречалась нам и ранее при работе с большими выборками. В классической статистике считается, что с накоплением анализируемых данных надежность результатов расчета возрастает. Мы же столкнулись с противоположным явлением, когда с увеличением объема материала увеличивалась ошибка выборочности (Равкин и др., 2022; 2023а, б). Это обусловлено тем, что по мере увеличения объема данных в выборке постепенно накапливается количество промежуточных сообществ, иллюстрирующих континуальность изменений. Это, в свою очередь, осложняет, а иногда и вовсе препятствует предметному объяснению состава групп, выделенных при кластерном анализе. Вторая причина затруднений при обобщении большого объема данных заключается в несовершенстве алгоритма используемых программ, в частности из-за наличия программных ограничений на количество анализируемых проб. Помимо этого, в выборке, несомненно, накапливаются погрешности и отличия, связанные с особенностями методики учета и описания местообитаний при использовании материалов, собранных нашими предшественниками.
Следует также упомянуть о проблеме с интерпретацией так называемых нулевых проб, т. е. местообитаний, где земноводных во время учета не обнаружили. Наличие таких «пустых» проб может быть обусловлено либо очень малой вероятностью встречи земноводных при низкой численности, либо с их реальным отсутствием во время проведения учета из-за экстремальности условий среды обитания. При этом зачастую неясно, каким следует считать сходство местообитаний, в которых земноводные во время учета не обнаружены: стопроцентным или же нулевым. При нулевом варианте сходства эти пробы образуют веер абсолютно не сходных между собой одиночных кластеров, хотя, руководствуясь экспертным мнением, часть из них следует объединять в одну группу местообитаний, заведомо непригодных для обитания исследуемых животных.
Наличие нулевых показателей обилия не только не позволяет применять стандартные методы оценки достоверности различий между средними вариантами населения, но и может исказить сами различия. Это обусловлено дополнительным сходством, возникающим между выборками, содержащими большое количество нулевых значений обилия. Чтобы избежать подобных искажений, варианты населения с нулевыми значениями обилия, как правило, исключают из расчетов. Однако необходимо учитывать, что в случае заведомой непригодности условий среды для обитания земноводных нулевые значения обилия это только подтверждают, и результаты учета в таких местах необходимо использовать в общем анализе материалов. Поэтому нами нули были исключены из расчетов с соответствующим уменьшением объема выборки только при оценке достоверности различий в распределении подобных видов и кластеризации сообществ. Во всех прочих расчетах к каждому варианту населения добавляли «фиктивный вид» с обилием 0.0001. Это делалось для того, чтобы все нулевые пробы стали на 100 % похожими и объединились в один кластер, в соответствии с умозрительными представлениями о причинах сходства этих вариантов населения.
Перечисленные выше недостатки иногда удается преодолеть, усреднив исходные показатели обилия по группам растительных формаций, выделенным в монографии (Ильина и др., 1985), с пояснительным текстом к карте (Растительность Западно-Сибирской равнины, 1976) в масштабе 1:1500000. В качестве примера можно привести группу формаций «Среднетаежные елово-кедровые леса и производные сообщества на их месте», объединяющую 7 выделов карты растительности под номерами 72–79. Именно такой подход мы использовали при анализе собранных материалов, однако ни одно из кластерных разбиений не удалось удовлетворительно интерпретировать. Поэтому был применен новый подход – с «экспертной кластеризацией», выполненной абсолютно умозрительно, но с учетом наличия хорошо интерпретируемых кластеров, полученных во всех разбиениях на предыдущих этапах анализа.
При анализе пространственно-временной динамики населения использованные материалы разделены сначала на два равных временных отрезка по 26 лет, с количеством проб 925 и 698, а затем таким образом, чтобы число проб в подвыборках различалось как можно меньше. В результате чего получилось 798 и 825 проб (соответственно с 1970 по 1989 г. и с 1990 по 2022 г.). Оценка достоверности различий выполнена по t-критерию Стьюдента с поправками Бонферрони и Бенджамини – Хохберга на множественность сравнений (Наркевич и др., 2020). По критерию Стьюдента различия достоверны, если вероятность случайного получения оценки по этому показателю не превышает 5 % (≤0.05). Поправку Бонферрони считают при учете множественности сравнений наиболее жесткой, а Бенджамини – Хохберга – наименее консервативной. Для каждой поправки различия достоверны, если скорректированная оценка достоверности ≤0.05.
Доверительные интервалы приведены по Е. С. Равкину и Н. Г. Челинцеву (1990) в несимметричном варианте. Вычисления произведены в основном с помощью пакетов PAST4 (Hammer et al., 2001) и Jacobi4 (Polunin et al., 2019). Относительная ошибка приведена как показатель надежности рассчитанных значений, а доверительные интервалы указаны только при описании временной динамики численности, поскольку по распределению достоверность различий не обсуждается.
Для оценки численности земноводных показатели их обилия умножены на площади местообитаний, рассчитанные по упоминавшейся выше карте «Растительность Западно-Сибирской равнины» и электронному варианту ландшафтной карты в масштабе 1:1 000 000 (Геоинформационная система Алтае-Саянского экорегиона, 2001).
Балльные оценки обилия приведены в границах, рекомендованных А. П. Кузякиным (1962), с добавлением верхних и нижних градаций (табл. 1).
Таблица 1. Балльные оценки обилия видов земноводных
| Градация обилия | Число особей на единицу пересчета |
| Чрезвычайно многочисленные | 1000 и более |
| Весьма многочисленные | 100–999 |
| Многочисленные | 10–99 |
| Обычные | 1–9 |
| Редкие | 0.1–0.9 |
| Очень редкие | 0.01–0.09 |
| Чрезвычайно редкие | менее 0.01 |
По литературным данным, в Западной Сибири обитает 11 видов земноводных (Kuranova et al., 2016), включая выделенные на основании сравнительно недавних молекулярно-генетических исследований три вида зеленых жаб и два вида озерных лягушек (Simonov et al., 2022). В данной статье эти виды рассматриваются как комплексы генетически близких форм – комплекс Bufotes viridis (Laurenti, 1768) и комплекс Pelophylax ridibundus (Pallas, 1771). Поскольку за время сбора материалов представления об объеме видов существенно изменились, на основании собранных данных невозможно судить о распределении и численности отдельных видов, входящих в состав каждого комплекса. Кроме того, за 52 года проведения учетных работ на обследованной территории нами ни разу не встречена травяная лягушка Rana temporaria Linnaeus, 1758. Таким образом, в статье представлена информация о двух видовых комплексах и 6 видах земноводных, видовые и родовые названия которых даны по Е. А. Дунаеву и В. Ф. Орловой (2017).
Результаты
Распределение
Сибирский углозуб Salamandrella keyserlingii Dybowski in Dybowski et Godlewski, 1870 на равнине предпочитает пойменные ландшафты долин крупных рек и их притоков (в среднем за лето 721 особь/км2). В полтора раза меньшие показатели его обилия отмечены на низинных и переходных (эвтрофных и мезотрофных) болотах вне пойм, и почти втрое меньшие – во внепойменных естественных и сельскохозяйственных суходольных ландшафтах. На верховых (олиготрофных) болотах углозуба в 14 раз меньше, чем на низинных и переходных болотах. По сравнению с верховыми болотами даже в поселках и застроенных садах средний уровень его обилия несколько выше (табл. 2).
Таким образом, по принятой шкале обилия углозуб на равнине характеризуется как весьма многочисленный вид в поймах, на переходных и низинных болотах, а также на суходолах, исключая селитебные территории. На верховых болотах, в поселках и застроенных садах он многочислен, а в городах ни разу не встречен. В горной части региона углозуб многочислен в среднем по предгорьям, включая как собственно предгорные ландшафты, так и мелкосопочные, горно-долинные, а также межгорно-котловинные, расположенные среди предгорий, на том же уровне абсолютных высот. В каких-либо местообитаниях за пределами этого высотного уровня его не встречали.
Таблица 2. Среднее обилие земноводных по группам равнинных и горных ландшафтов Западной Сибири, особей/км2
| Группа ландшафтов | Сибир-ский углозуб | Обыкно-венный тритон | Чесноч-ница | Серая жаба | Зеленая жаба | Озерная лягушка | Сибирская лягушка | Остромордая лягушка | Всего |
| Внепойменные суходолы (кроме застроенных) | 208 | 32 | 89 | 1452 | 10 | 1.8 | 32 | 4203 | 6028 |
| Поймы | 721 | 45 | 1579 | 504 | 0 | 195 | 1156 | 26520 | 30720 |
| Болота внепойменные низинные и переходные | 488 | 286 | 79 | 4438 | 4 | 0 | 712 | 27243 | 33251 |
| Болота верховые | 35 | 4 | 0 | 7985 | 0 | 0 | 5 | 5926 | 13956 |
| Поселки и застроенные сады | 47 | 2 | 1006 | 1185 | 0 | 0 | 82 | 3970 | 6291 |
| Города | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1192 | 1192 |
| В среднем по равнинной части региона |
272 | 54 | 344 | 1964 | 6 | 25 | 241 | 9293 | 12200 |
| Доля от общего обилия в равнинной части региона, % | 2 | 0.4 | 2.8 | 16 | 0.1 | 0.2 | 2 | 76 | 100 |
| Высокогорья | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Среднегорья | 0 | 0 | 0 | 50 | 0 | 0 | 0 | 202 | 253 |
| Низкогорья | 0 | 10 | 0 | 2866 | 0.3 | 3 | 0 | 1751 | 4629 |
| Предгорья | 12 | 0 | 0 | 526 | 0 | 124 | 0 | 8120 | 8782 |
| В среднем по горной части региона |
2 | 4 | 0 | 1215 | 0.1 | 26 | 0 | 2341 | 3588 |
| Доля от общего обилия в горной части региона, % | 0.06 | 0.1 | 0 | 34 | 0.003 | 0.7 | 0 | 65 | 100 |
Обыкновенный тритон Lissotriton vulgaris (Linnaeus, 1758) многочислен на суходолах, в поймах, на внепойменных низинных и переходных болотах. На верховых болотах, в поселках и застроенных садах он обычен, а в городах его не встречали. В горной части региона тритон отмечен только в низкогорьях, т. е. в условной группе ландшафтов (низкогорных, горно-долинных, мелкосопочных и межгорно-котловинных), занимающих абсолютные высоты до 1000 м. В среднем по группе ландшафтов низкогорного яруса он характеризуется как многочисленный вид.
Обыкновенная чесночница Pelobates fuscus (Laurenti, 1768) найдена только в западной приуральской части равнины – в Тоболо-Ишимской лесостепи и степи. Наибольший уровень ее обилия зарегистрирован в поймах, где она чрезвычайно многочисленна даже в среднем, в пересчете на всю площадь пойменных ландшафтов в пределах Западно-Сибирской равнины. Как весьма многочисленный вид со средним обилием всего лишь в 1.6 раза меньшим, чем в поймах, она характерна для поселков и застроенных садов. На внепойменных суходолах, а также на низинных и переходных болотах чесночница многочисленна, а на очень редко встречающихся в Тоболо-Ишимской лесостепи и незначительных по площади участках реликтовых верховых болот не обнаружена, так же как и в городах.
Обыкновенная серая жаба Bufo bufo (Linnaeus, 1758) чрезвычайно многочисленна в большинстве рассматриваемых групп равнинных ландшафтов, многочисленна в поймах и не встречена в городах. При этом ее обилие неуклонно сокращается по направлению от верховых болот к внепойменным низинным и переходным (почти вдвое) и далее – к суходолам (примерно втрое), поселкам и застроенным садам (в 1.2 раза), а затем – в поймах (в 2.3 раза). В горах серая жаба чрезвычайно многочисленна в группе ландшафтов низкогорного высотного яруса. В предгорьях средний уровень ее обилия почти вчетверо ниже, здесь она весьма многочисленна. В среднегорьях, на абсолютных высотах от 1000 до 2000 м, серая жаба многочисленна, несмотря на резкое сокращение ее обилия в 57 раз, а в высокогорьях (более 2000 м над уровнем моря) при проведении учета она не обнаружена.
Зеленые жабы комплекса Bufotes viridis (Laurenti, 1768) в равнинной части региона встречаются в Тоболо-Ишимской (Курганская область) и Приобской лесостепи (Новосибирск и его окрестности). При этом во время учетов в Курганской области в 1981–1985 гг. зеленые жабы не были обнаружены, несмотря на широкий охват территории (материалы В. П. Старикова, а также В. Н. и Т. К. Блиновых в Банке данных ИСиЭЖ СО РАН, далее – БД). Тем не менее в литературе имеются сведения о находках не менее 100 особей зеленой жабы во многих районах Курганской области, на основании чего ее относят к категории редких видов на северной границе ареала (Стариков, 1986, 2002; Ищенко, 2012; Simonov et al., 2022). По-видимому, это свидетельствует об активном расселении в северном направлении зеленых жаб, обитающих в степной зоне, в пределах примыкающей с юга Кустанайской области Казахстана. Так, по результатам учетов, проведенных В. Н. и Т. К. Блиновыми в 1986 г. на трех степных участках в Кустанайской области (материалы БД), в подзоне северной степи, в 50 км к югу от границы с Курганской областью (с. Аксуат), зеленые жабы не обнаружены. На втором ключевом участке в 160 км южнее Аксуата (с. Новонежинка), в подзоне средней степи, они найдены в большом количестве на тростниково-осоковых болотах (120 особей/км2), в степных местообитаниях (210–420) и крупных поселках (1500 особей/км2). На третьем участке, расположенном в 100 км к югу от второго, в подзоне южной степи (Наурзумский заповедник), зеленые жабы встречались почти повсеместно, за исключением песчано-ковыльных целинных степей, с очень высоким обилием в пределах от 2000 особей/км2 в сухих сосновых островных лесах до 94000 особей/км на тростниково-рогозовых болотах (займищах).
Локальная новосибирская популяция, обнаруженная в 1984 г. в северной лесостепи Приобья (Золотаренко, 1985), существует до настоящего времени и, по-видимому, процветает, поскольку значительно расширилась в своем распространении. По данным учетов 2003–2004 гг., показатели обилия зеленых жаб достигали на внепойменных суходолах 7029 тыс. особей/км2, а на внепойменных болотах – 437 особей/км2.
В горной части региона зеленая жаба зарегистрирована нами как редкий вид, обнаруженный только в пределах низкогорий. Но, судя по литературным данным, этих жаб неоднократно находили и в среднегорьях (Яковлев, 1999; Litvinchuk et al., 2010).
Озерные лягушки комплекса Pelophylax ridibundus (Pallas, 1771) в равниной части региона широко распространены по долинам Оби, Иртыша и крупных притоков этих рек. К настоящему времени по долине Оби они расселились от Алтая до Новосибирска и далее, по-видимому, до Томска (Kuranova et al., 2016). По долине Иртыша обстоятельно прослежено их расселение от Усть-Каменогорска до Павлодара (Каптенкина и др., 2022) и далее до границы с Россией. В 2002 г. эти лягушки были отмечены в пойме Иртыша в Омске (личное сообщение С. А. Соловьёва). По результатам проведенных учетов они обычны на внепойменных суходолах и весьма многочисленны в пойменных ландшафтах. Даже в целом по равнинной части региона средний уровень их обилия достигает 25 особей/км2. В горной части региона они приурочены преимущественно к пойменным долинам, при этом обычны в пределах низкогорий и многочисленны в предгорьях. Средний уровень их обилия на горной территории примерно такой же, как и на равнине, – 26 особей/км2.
Сибирская лягушка Rana amurensis Boulenger, 1886 чрезвычайно многочисленна только в поймах равнинных рек и весьма многочисленна на внепойменных низинных и переходных болотах. На суходолах, в поселках и застроенных садах она многочисленна, а на верховых болотах обычна. Не обнаружена R. amurensis лишь в городах на равнине, а также в горной части региона.
Остромордая лягушка Rana arvalis Nilsson, 1842 чрезвычайно многочисленна как в среднем по территории Западно-Сибирской равнины, так и во всех рассматриваемых группах ландшафтов. При этом наиболее велики показатели ее обилия на внепойменных низинных и переходных болотах, а также в поймах рек. Далее прослеживается сокращение среднего уровня ее обилия примерно в 5–6 раз на верховых болотах и на суходолах, включая поселки и застроенные сады, а затем еще почти вчетверо – в городах. В горной части региона остромордая лягушка чрезвычайно многочисленна в среднем по группам ландшафтов от предгорного до низкогорного уровня и многочисленна в среднегорьях. При этом с подъемом в низкогорья ее обилие сокращается вчетверо, при переходе к среднегорьям – еще примерно в 9 раз.
Таким образом, на равнине в пределах от степной зоны вплоть до подзональной полосы южных субарктических тундр максимальные показатели суммарного обилия земноводных характерны для внепойменных низинных и переходных болот, а также пойменных долин крупных рек, где средняя плотность населения превышает 30 тысяч особей/км2. Кроме того, чрезвычайно многочисленны земноводные на верховых болотах, хотя средний уровень их суммарного обилия здесь вдвое ниже, чем на прочих заболоченных территориях. По сравнению с этими, очевидно, оптимальными для земноводных ландшафтами, в 2–6 раз менее плотно заселены внепойменные суходолы, поселки, застроенные сады и особенно города (в 12–28 раз). В целом на Западно-Сибирской равнине больше всего остромордой лягушки. По суммарному обилию всех земноводных на ее долю приходится 76 %. На втором месте по этому показателю – серая жаба (16 %). Второстепенны по долевому участию в населении – чесночница (3 %), сибирский углозуб и сибирская лягушка (по 2 %). На последних местах – зеленые жабы, озерные лягушки и обыкновенный тритон (0.1–0.4 %).
В горной части региона, как и на равнине, больше всего остромордой лягушки и серой жабы (в среднем 2341 и 1215 особей/км2), составляющих по обилию соответственно 65 и 34 % населения земноводных, так что на долю всех остальных вместе взятых видов земноводных приходится менее 1 %. Остромордая лягушка явно предпочитает ландшафты предгорного высотного уровня (8120 особи/км2), где чаще всего встречаются, особенно в межгорных котловинах, низинные и переходные болота, а также заболоченные участки пойменных долин. В низкогорьях средний уровень ее обилия уменьшается почти в 5 раз, а в среднегорьях отмечено более резкое сокращение – еще в 9 раз. Серой жабы больше всего в низкогорьях (2866 особей/км2), в 5 раз меньше в предгорьях (526) и сравнительно мало в среднегорьях (50 особей/км2).
Пространственная неоднородность населения
По итогам анализа всех имеющихся материалов учета составлена классификация сообществ земноводных Западной Сибири, которая включает три типа населения. При этом первые два типа выделяются однозначно по результатам кластеризации, а вот формирование третьего типа населения и его разбиение на подтипы выполнено экспертно, поскольку результаты кластерного разбиения были неоднозначны и плохо интерпретировались.
Первый тип населения – полярно-высокогорный – объединяет те местообитания, где земноводные не встречены из-за экстремальности условий существования. Это все высокогорные ландшафты, полярные пустыни и все равнинные тундры, за исключением южных субарктических.
Второй – лесотундровый – тип населения включает сообщества южных субарктических тундр, лесотундры и темнохвойных лесов в северной и средней тайге, а также в среднегорьях. Во всех перечисленных местообитаниях складываются пессимальные условия для обитания земноводных, что отражено невысокими показателями суммарного обилия и видового богатства. Здесь встречено всего 3 вида – остромордая лягушка, серая жаба и сибирский углозуб, а плотность населения земноводных не превышает 4 особей/100 ц/с.
Третий – температный – тип населения представлен сообществами земноводных, населяющих местообитания с субоптимальными и оптимальными условиями существования. К нему отнесена группировка из всех прочих сообществ, не вошедших в первые два типа, дополнительно разделенная на 4 подтипа в соответствии со степенью сходства включенных в их состав вариантов населения.
Информативность классификации, рассчитанная по исходным данным, почти нулевая (0.06 %), а по усредненным показателям обилия с использованием и нулевых значений – 2 % (коэффициент множественной корреляции 0.12). Информативность структурных представлений по средним показателям обилия на 1 км2 равна 8 % дисперсии на матрице коэффициентов сходства. Из выявленных факторов среды, коррелирующих с неоднородностью сообществ земноводных, наиболее значимо расстояние до водоемов выплода, учитывающее 18 % дисперсии сходства. Это связано с обилием сеголеток, которые рассредоточиваются после выхода на сушу, и обилие их уменьшается по мере удаления от мест выплода. Влияние теплообеспеченности, по сравнению с удаленностью от водоемов выплода, в 1.2 раза меньше. Еще меньше степень корреляции с увлажнением (в 2.6 раза). Связь с типом растительности, макрорельефом (равнина – горы) и составом лесообразующих пород невелика (1–5 % дисперсии) и минимальна с застроенностью территории (0.4 %). Последнее связано в основном с незначительной площадью и количеством селитебных местообитаний в выборке. Всего этими факторами можно объяснить 29 % сходства (множественный коэффициент корреляции 0.54). Суммарная коррелятивная связь выявленных факторов и структурных представлений с пространственной неоднородностью населения равна соответственно 31 % и 0.56.
Составленная классификация использована для построения графа, отображающего (на уровне тенденций) связь пространственной изменчивости населения земноводных с ведущими (структурообразующими) факторами среды (рис. 2).
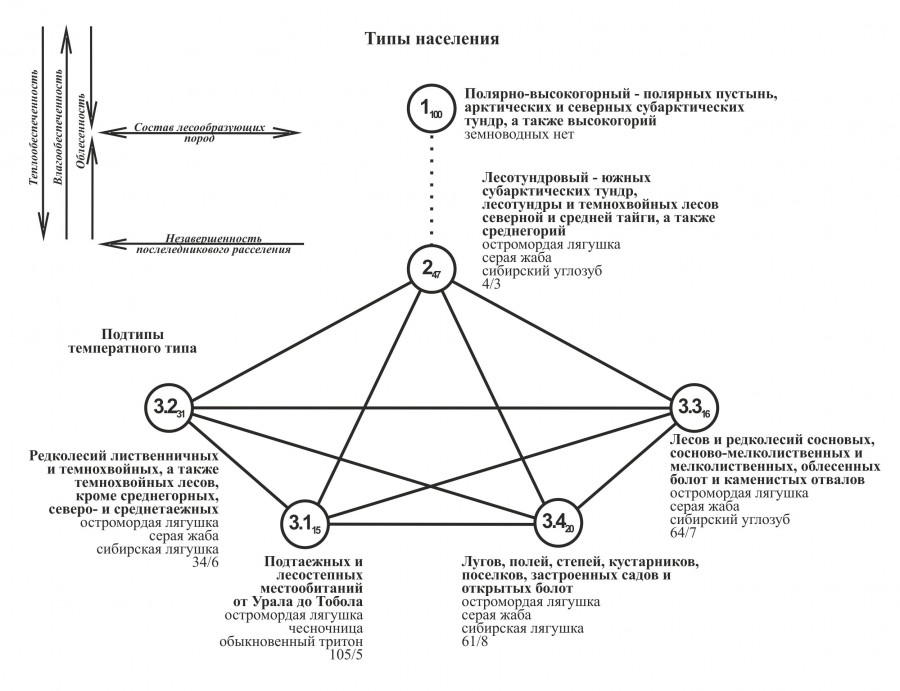
Рис. 2. Граф сходства населения земноводных Западной Сибири (3.2–3.4 – без подтаежных и лесостепных местообитаний от Урала до Тобола)
Fig. 2. Graph of the similarity of the amphibian population of Western Siberia (3.2–3.4 – excluding subtaiga and forest-steppe habitats from the Urals to Tobol)
Крупные цифры внутри кружков соответствуют номерам типов и подтипов населения земноводных. Мелкими цифрами в виде подстрочного знака показана средняя величина внутригруппового сходства (в процентах) вошедших в таксон проб. Рядом с кружками приведено название таксона классификации (см. текст), перечислены вошедшие в него группы сообществ, лидирующие по обилию виды и указаны суммарная плотность населения земноводных в количестве особей на 100 ц/с, а также число встреченных видов. Сплошными линиями, соединяющими кружки, показаны все значимые связи (>16 % сходства) между типами / подтипами населения. Нулевое сходство обозначено пунктирной линией. Стрелками в левом верхнем углу рисунка показаны факторы среды, коррелирующие с векторами изменения населения земноводных.
В целом, структура территориальных различий в населении земноводных, на наш взгляд, в значительной степени связана с изменением гидротермического режима (на усредненном уровне: зонально-подзональном для равнины и высотно-поясном для гор), определяющего степень благоприятности условий среды. Для обобщения и визуализации в реальном географическом пространстве представления о пространственной дифференциации населения земноводных, сложившегося в результате анализа данных в многомерном факторном пространстве, составлена соответствующая карта (рис. 3). В качестве легенды к ней также использована изложенная выше классификация населения земноводных.
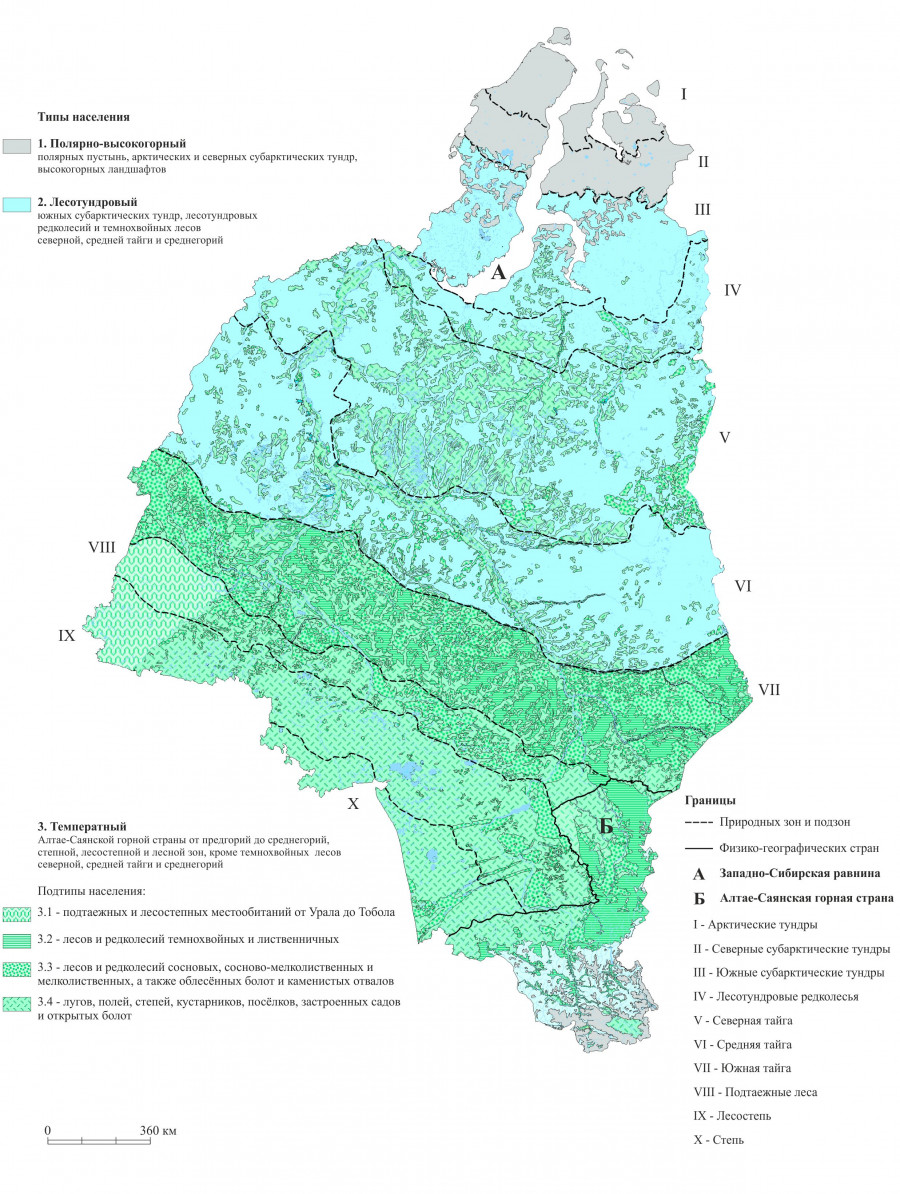 Рис. 3. Население земноводных равнинных и горных территорий Западной Сибири в пределах России (характеристики населения приведены на рис. 2)
Рис. 3. Население земноводных равнинных и горных территорий Западной Сибири в пределах России (характеристики населения приведены на рис. 2)
Fig. 3. Amphibian population of lowland and mountainous territories of Western Siberia within Russia (population characteristics are shown in Fig. 2)
На карте видны три широтные полосы, занятые вышеописанными типами населения земноводных. Самая обширная первая полоса с температным типом населения объединяет все равнинные территории с наиболее благоприятными для земноводных условиями среды: степную и лесостепную зоны, а также подзону южной тайги. По открытым болотам и поймам крупных рек этот тип населения проникает вплоть до южных субарктических тундр. Вторая полоса с лесотундровым типом населения охватывает южные субарктические тундры, лесотундровые редколесья, а также темнохвойные леса северной и средней тайги, где складываются пессимальные условия для обитания земноводных. Самая северная третья полоса включает полярные пустыни, арктические и северные субарктические тундры, где постоянное население земноводных отсутствует из-за экстремальности условий обитания.
В горах, где наблюдается высотно-ярусная дифференциация населения земноводных, температный тип занимает предгорно-низкогорный ярус с проникновением в среднегорья преимущественно по речным долинам. Распространение лесотундрового типа населения ограничено среднегорьями, тогда как в высокогорьях постоянное население земноводных отсутствует.
Численность и динамика плотности населения земноводных
Суммарная численность земноводных Западной Сибири составляет, по нашим расчетам, порядка 24 млрд особей. Подавляющее большинство земноводных обитает на равнине (97 %), при этом почти половина из них (44 %) занимает территорию средней тайги (табл. 3).
Мы сопоставили обилие земноводных в южной тайге Прииртышья и Приобья по трем временным отрезкам: 1 – 1970, 1971 гг.; 2 – 1990, 1991; 3 – 2005, 2006, 2012 и 2013 гг. Судя по данным метеослужбы г. Томска, расположенного в южной тайге, во второй и третий периоды температура воздуха в летнее время была выше, чем в первый отрезок, на 1 оС (14 оС в первый и по 15 оС во второй и третий периоды). Количество осадков было ниже соответственно на 11 и 9 мм (72; 56 и 63 мм), т. е. уменьшилось в 1.2 раза. Под Новосибирском, в Шлюзовском бору на границе лесостепи и подтаежных лесов, в 2016–2018 гг. при сходной теплообеспеченности и при увеличении увлажнения (в среднем на 13 мм) обилие земноводных в целом за два последних года возросло (Равкин и др., 2020).
Если допустить, что снижение количества осадков меняет обилие земноводных в той же степени, то плотность их населения в южной тайге по аналогии должна была уменьшиться в 1.4 и 1.3 раза. То есть увеличение летних осадков приводит к возрастанию обилия остромордой лягушки и углозуба, а снижение количества осадков – к уменьшению их численности, в основном за счет сеголеток. По-видимому, это связано с сокращением площади водоемов выплода, а иногда и с пересыханием части из них.
Таблица 3. Среднее обилие и численность земноводных в Западной Сибири
| Зона, подзона (на равнине), высотный уровень группы ландшафтов (в горах) | Особей/км2 | Всего, млн особей | |
| Полярные пустыни, арктические и северные субарктические тундры | 0 | 0 | |
| Южные субарктические тундры | 13 | 2 | |
| Лесотундра | 227 | 31 | |
| Северная тайга | 2111 | 1133 | |
| Средняя тайга | 1838 | 10507 | |
| Южная тайга | 15348 | 5994 | |
| Подтаежные леса | 21032 | 3239 | |
| Лесостепь | 7679 | 1984 | |
| Степная зона | 5920 | 455 | |
| Высокогорья | 0 | 0 | |
| Среднегорья | 253 | 13 | |
| Низкогорья | 4629 | 366 | |
| Предгорья | 8782 | 362 | |
| В среднем по равнинной части региона | 10092 | 23345 | |
| В среднем по горной части региона | 3588 | 741 | |
| В среднем по Западной Сибири | 9530 | 24086 |
В среднем по Западной Сибири снижение плотности населения земноводных во втором из трех вариантов счета снизилось на 38 %, а в третьем варианте возросло на 9 %. Такое несоответствие, по-видимому, связано с неупорядоченным по годам, природно-географическим зонам и подзонам сбором данных, поскольку материал целенаправленно собирали в первую очередь для решения зоогеографических задач. Попытка избавиться от этого недостатка для увеличения сравнимости не привела к улучшению результата. Надеясь на это, мы сначала рассчитали средневзвешенные показатели с учетом числа проб по годам (табл. 4). Не привел к лучшей сравнимости и переход от количественной к балльной оценке объема летних осадков как основного показателя влагообеспеченности. Для этого существенно меньшими считали значения, вошедшие в последнюю третью или их четвертую часть. Результаты оценки влияния количества осадков были по-прежнему противоречивыми. Например, в южной тайге, при абсолютной сравнимости данных за три периода, с уменьшением количества осадков отмечено достоверное монотонное сокращение обилия общего количества земноводных и остромордой лягушки (Равкин и др., 2020). По всей Западной Сибири снижение обилия тоже было статистически достоверным (табл. 5–7), но аналогичных изменений количества осадков при этом не отмечено. Тем не менее, на наш взгляд, дело все-таки в динамике количества летних осадков.
Таблица 4. Количество проб и среднее обилие земноводных по годам в Западной Сибири (без полярных островов, тундр, лесотундры и высокогорий)
| Год | Проб | Особей/км2 | |||||||
| Сибирский углозуб | Обыкновенный тритон | Обыкновенная чесночница | Зеленая жаба | Серая жаба | Озерная лягушка | Остромордая лягушка | Сибирская лягушка | ||
| 1970 | 27 | 84 | 0 | 0 | 0 | 59 | 0 | 36611 | 1183 |
| 1971 | 12 | 2697 | 0 | 0 | 0 | 17278 | 0 | 25356 | 304 |
| 1972 | 14 | 261 | 0 | 0 | 0 | 2116 | 0 | 17454 | 2373 |
| 1973 | 10 | 420 | 0 | 0 | 0 | 0 | 0 | 11018 | 0 |
| 1975 | 6 | 0 | 0 | 0 | 0 | 13936 | 0 | 10254 | 0 |
| 1976 | 29 | 0.1 | 0 | 0 | 0 | 0 | 0 | 6319 | 0 |
| 1977 | 92 | 54 | 0 | 0 | 0 | 648 | 0 | 6866 | 0 |
| 1978 | 41 | 22 | 36 | 0 | 0 | 696 | 0 | 11465 | 0 |
| 1979 | 19 | 1970 | 0 | 0 | 0 | 698 | 0 | 9706 | 789 |
| 1980 | 42 | 472 | 0 | 0 | 0 | 425 | 0 | 5267 | 0 |
| 1981 | 66 | 1016 | 0 | 201 | 0 | 6346 | 6 | 21967 | 351 |
| 1982 | 42 | 898 | 1056 | 1714 | 0 | 570 | 0 | 12652 | 0 |
| 1983 | 48 | 198 | 21 | 442 | 0 | 33 | 0 | 10082 | 185 |
| 1984 | 67 | 6 | 0 | 3560 | 0 | 2124 | 0 | 18076 | 1046 |
| 1985 | 35 | 138 | 0 | 1653 | 0 | 7716 | 0 | 5764 | 0 |
| 1986 | 33 | 105 | 27 | 0 | 0 | 3303 | 0 | 16905 | 60 |
| 1987 | 31 | 0 | 0 | 0 | 0 | 2510 | 0 | 4909 | 0 |
| 1988 | 35 | 5 | 7 | 0 | 1.5 | 2096 | 0 | 2228 | 0 |
| 1989 | 40 | 0 | 0 | 0 | 0 | 0 | 0 | 797 | 14 |
| 1990 | 23 | 741 | 0 | 0 | 0 | 3879 | 0 | 6919 | 2113 |
| 1991 | 14 | 839 | 24 | 0 | 0 | 5381 | 0 | 11587 | 1429 |
| 1992 | 22 | 791 | 481 | 0 | 0 | 3802 | 0 | 9805 | 14 |
| 1993 | 26 | 378 | 0 | 0 | 0 | 311 | 0 | 8082 | 0 |
| 1994 | 21 | 187 | 0 | 0 | 0 | 1334 | 0 | 2426 | 0 |
| 1995 | 15 | 0 | 0 | 0 | 0 | 173 | 0 | 5157 | 0 |
| 1996 | 25 | 19 | 0 | 0 | 0 | 318 | 0 | 2360 | 0 |
| 1997 | 21 | 0 | 0 | 0 | 0 | 449 | 0 | 222 | 0 |
| 1998 | 45 | 0 | 0 | 0 | 0 | 682 | 0 | 2053 | 0 |
| 1999 | 29 | 0 | 0 | 0 | 0 | 286 | 0 | 505 | 0 |
| 2000 | 38 | 0 | 0 | 0 | 0 | 425 | 769 | 2529 | 0 |
| 2001 | 28 | 0 | 0 | 0 | 0 | 563 | 0 | 818 | 0 |
| 2002 | 8 | 0 | 0 | 0 | 0 | 803 | 0 | 1928 | 0 |
| 2003 | 17 | 199 | 0 | 0 | 26 | 474 | 0 | 6083 | 0 |
| 2004 | 24 | 61 | 0 | 0 | 293 | 733 | 0 | 1494 | 6 |
| 2005 | 56 | 139 | 0 | 0 | 0 | 354 | 0 | 3363 | 0 |
| 2006 | 32 | 94 | 172 | 0 | 0 | 790 | 0 | 7903 | 547 |
| 2007 | 33 | 20 | 11 | 0 | 0 | 900 | 0 | 8389 | 191 |
| 2008 | 33 | 0 | 0 | 0 | 0 | 461 | 0 | 1802 | 0 |
| 2009 | 40 | 0 | 0 | 0 | 0 | 502 | 0 | 1620 | 0 |
| 2010 | 37 | 122 | 0 | 0 | 0 | 251 | 0 | 4460 | 0 |
| 2011 | 19 | 0 | 0 | 8 | 0 | 808 | 0 | 1286 | 0 |
| 2012 | 32 | 175 | 0 | 0 | 0 | 5412 | 0 | 6981 | 14 |
| 2013 | 30 | 82 | 0 | 0 | 0 | 9785 | 0 | 11154 | 0 |
| 2014 | 31 | 0 | 0 | 0 | 0 | 3715 | 0 | 893 | 0 |
| 2015 | 26 | 0 | 0 | 0 | 0 | 362 | 187 | 5804 | 0 |
| 2016 | 20 | 0 | 14 | 0 | 0 | 499 | 0 | 2231 | 0 |
| 2017 | 7 | 0 | 0 | 0 | 0 | 682 | 0 | 3777 | 0 |
| 2018 | 10 | 0 | 26 | 0 | 0 | 3232 | 0 | 12848 | 0 |
| 2019 | 9 | 0 | 0 | 0 | 0 | 423 | 0 | 2197 | 0 |
| 2020 | 9 | 0 | 0 | 0 | 0 | 832 | 0 | 1063 | 0 |
| 2021 | 16 | 31 | 0 | 0 | 0 | 393 | 0 | 3250 | 0 |
| 2022 | 13 | 10 | 0 | 0 | 0 | 417 | 0 | 589 | 0 |
Таблица 5. Среднелетнее обилие земноводных (особей/км2) в Западной Сибири по временным отрезкам за 1970–1989 и 1990–2022 гг. (лесная, лесостепная, степная зоны Западно-Сибирской равнины и соответствующие высотные пояса Алтае-Саянской горной страны)
| 1 вариант | 2 вариант | |||
| Вид | 1970–1996 | 1997–2022 | 1970–1989 | 1990–2022 |
| Сибирский углозуб | 300 | 42 | 287 | 104 |
| Обыкновенный тритон | 61 | 9 | 60 | 20 |
| Обыкновенная чесночница | 717 | 0.2 | 868 | 0.2 |
| Зеленая жаба | 201 | 11 | 243 | 9 |
| Серая жаба | 1918 | 1305 | 1953 | 1392 |
| Озерная лягушка | 0.6 | 49 | 1 | 39 |
| Остромордая лягушка | 9642 | 3526 | 10500 | 3921 |
| Сибирская лягушка | 270 | 35 | 241 | 108 |
| Всего | 13109 | 4977 | 14152 | 5593 |
| Количество проб | 966 | 698 | 798 | 866 |
Таблица 6. Достоверность различий по среднелетнему обилию земноводных в Западной Сибири в 1970–1989 и 1990–2022 гг. (по формальным и предметным критериям)
| Критерий | ||||
| Вид | ttest | при допущении нормальности распределения | Бонферрони | Бенджамини – Хохберга |
| Сибирский углозуб | 2.52 | 0.01 | 0.11 | 0.03 |
| Обыкновенный тритон | 1.00 | 0.32 | 2.86 | 0.4 |
| Обыкновенная чесночница | 4.72 | 0.000003 | 0.00002 | 0.000008 |
| Зеленая жаба | 1.85 | 0.06 | 0.57 | 0.1 |
| Серая жаба | 0.98 | 0.33 | 2.95 | 0.3 |
| Озерная лягушка | 1.83 | 0.07 | 0.61 | 0.1 |
| Остромордая лягушка | 6.27 | 0.0000000005 | 0.000000004 | 0.000000002 |
| Сибирская лягушка | 1.49 | 0.14 | 1.22 | 0.2 |
| Всего | 6.51 | 0.0000000001 | 0.0000000009 | 0.0000000009 |
Примечание. В табл. 6 и 7 жирным шрифтом показаны достоверные отличия.
Таблица 7. Достоверность различий обилия земноводных в Западной Сибири в 1970–1996 и 1997–2022 гг. (в среднем за лето)
| Критерий | ||||
| Вид | ttest | при допущении нормальности распределения | Бонферрони | Бенджамини – Хохберга |
| Сибирский углозуб | 3.509045 | 0.00046171 | 0.00369368 | 0.00092342 |
| Обыкновенный тритон | 1.275195 | 0.20241858 | 1.61934866 | 0.20241858 |
| Обыкновенная чесночница | 3.844877 | 0.00012516 | 0.00100131 | 0.00033377 |
| Зеленая жаба | 1.484607 | 0.13783767 | 1.10270137 | 0.15752877 |
| Серая жаба | 1.05902 | 0.28974452 | 2.31795618 | 0.25755069 |
| Озерная лягушка | 2.255846 | 0.02420992 | 0.19367939 | 0.0322799 |
| Остромордая лягушка | 5.746196 | 0.00000001 | 0.00000009 | 0.00000004 |
| Сибирская лягушка | 2.608553 | 0.00917416 | 0.07339329 | 0.01467866 |
| Всего | 6.096177 | 0.000000001 | 0.00000001 | 0.00000001 |
Для выявления многолетней динамики населения земноводных Западной Сибири все материалы сначала были разделены на два временных отрезка примерно поровну по числу лет: с 1970 по 1996 г. и с 1997 по 2022 г. Затем тоже на два периода, но таким образом, чтобы число анализируемых проб в каждом из отрезков было примерно одинаковым – с 1970 по 1989 г. и с 1990 по 2022 г. Данные по обилию были взяты только по территории, где земноводные обнаружены в сравнительно большом количестве, т. е. исключая полярные острова, тундровую и лесотундровую зоны, а также высокогорья. В итоге остались только пробы по лесной, лесостепной и степной зонам, а также аналогичным высотным поясам в горах. В двух случаях формально достоверное снижение обилия при допущении нормальности распределения, а также по Бонферрони и Бенджамини Беннджамини – Хохберга Хохбергу отмечено у сибирского углозуба, остромордой и сибирской лягушек. В третьем варианте счета формально были достоверны отличия по тем же видам и озерной лягушке. Показатели обилия остромордой лягушки во второй временной отрезок были в 2.7 раза меньше, чем в первый и третий отрезки. Эти отличия достоверны по всем критериям при расчетах как с сеголетками, так и без них. Материалы по чесночнице, сибирскому углозубу и сибирской лягушке по предметным соображениям следует считать «дефектными». По чесночнице – поскольку она встречается только на западе региона, до Ишима, а эта территория была обследована лишь в первый отрезок (с 1981 по 1986 г.). Численность озерных лягушек, впервые отмеченных нами при учете земноводных в лугово-степных низкогорьях Северного Алтая в 1981 г., постепенно нарастала по мере их продвижения по долине Оби в северном направлении, поэтому неудивительно, что средний уровень их обилия во второй период оказался более высоким. Сибирская лягушка встречается только в поймах крупных рек. К северу ее становится меньше. В этом же направлении по годам поочередно нами были проведены учеты земноводных, что, возможно, и увеличило степень корреляции ее обилия с зональной теплообеспеченностью. В Западной Сибири от северной тайги до Ледовитого океана сибирская лягушка не встречена. Поэтому данные, собранные нами по этому виду, видимо, непригодны для расчета динамики численности по отдельным годам. То же самое можно сказать об обилии сибирского углозуба, которое уменьшается к северу. Кроме того, очень высокое обилие этого вида прослежено только один раз в южнотаежной пойме Иртыша, где ловчая канавка была расположена около водоема выплода.
Итак, и по формальным, и по предметным соображениям можно говорить о достоверности различий только для остромордой лягушки. Уменьшение ее численности, по-видимому, связано со снижением количества осадков в летний период. Статистически недостоверное, но существенное снижение обилия отмечено, кроме того, у серой жабы – достаточно многочисленного вида.
Заключение
Максимальное обилие земноводных на Западно-Сибирской равнине свойственно внепойменным низинным и переходным болотам. В поймах крупных рек оно немного ниже из-за отрицательного влияния половодий. Кроме того, чрезвычайно многочисленны земноводные и на верховых болотах, хотя средний уровень их обилия здесь как минимум вдвое ниже, чем на прочих заболоченных территориях в связи с обеднением минерального питания фитоценозов и кормности. В 5–6 раз меньше земноводных на суходолах, в поселках, застроенных садах и в 28 раз меньше в городах.
Больше всего на Западно-Сибирской равнине остромордой лягушки. На ее долю приходится 65 % численности всех земноводных. На втором месте – серая жаба (34 %). В горах численно преобладают эти же два вида. Остромордая лягушка предпочитает здесь ландшафты предгорного уровня высот, особенно пойменные речные долины, где сравнительно много заболоченных участков. В низкогорьях обилие этой лягушки уменьшается пятикратно, а в среднегорьях – еще в 8 раз, сокращаясь до минимального уровня (202 особи/км2). Описанный тренд вполне соответствует давно сложившемуся представлению об уменьшении обилия земноводных и большинства других животных по мере подъема в горы из-за закономерного уменьшения температуры воздуха, тем самым без всяких расчетов наглядно подтверждая наличие коррелятивной связи между плотностью населения и теплообеспеченностью территории. Серой жабы больше всего в низкогорьях, в 5 раз меньше в предгорьях и в 57 раз меньше в среднегорьях. Таким образом, связь с теплообеспеченностью в зависимости от высоты местности у серой жабы менее выражена по сравнению с остромордой лягушкой. Суммарная связь выявленных факторов и структурных представлений по земноводным с пространственной неоднородностью их населения объясняет 31 % дисперсии коэффициентов сходства, коэффициент множественной корреляции – 0.56.
Суммарная численность земноводных в Западной Сибири достигает, по нашим расчетам, примерно 24 млрд особей, при этом 97 % их населения занимает равнинную часть региона. Больше всего амфибий обитает в лесной зоне, в подзоне средней тайги, где сосредоточена почти половина (44 %) населения земноводных Западной Сибири.
По формальным (полученным статистически) и предметным (умозрительным) соображениям можно говорить о достоверности межгодовых отличий в обилии по выделенным периодам только у остромордой лягушки. Уменьшение ее численности связано со снижением количества осадков в летний период. Существенное, хотя и статистически недостоверное снижение обилия отмечено, кроме того, у серой жабы – достаточно многочисленного вида.
Библиография
Борисович О. Б., Цыбулин С. М., Торопов К. В., Фомин Б. Н., Граждан К. В., Богомолова И. Н. Земноводные и пресмыкающиеся равнинных и предгорно-низкогорных ландшафтов Верхнего Приобья // Сибирский экологический журнал. 2002. Т. 9, № 4. С. 425–440.
Геоинформационная система Алтае-Саянского экорегиона. М.: ИГЕМ РАН – WWFRussia, 2001.
Дунаев Е. А., Орлова В. Ф. Земноводные и пресмыкающиеся России: Атлас-определитель. М.: Фитон XXI, 2017. 328 с.
Западная Сибирь. М.: Изд-во АН СССР, 1963. 488 с.
Золотаренко Г. С. О нахождении зеленой жабы в окрестностях Новосибирска // Вопросы герпетологии: Тезисы докладов VI Всесоюзной конференции. Л.: Наука, 1985. С. 80–81.
Ильина И. С., Лапшина Е. И., Лавренко Н. Н., Мельцер Л. И., Романова Е. А., Богоявленский Б. А., Махно В. Д. Растительный покров Западно-Сибирской равнины. Новосибирск: Наука, 1985. 251 с.
Ищенко В. Г. Зеленая жаба // Красная книга Курганской области. Изд. 2-е. Курган: Изд-во Курганского гос. ун-та, 2012. С. 93.
Каптёнкина А. Г., Дуйсебаева Т. Н., Ахмеденов К. М., Хромов В. А., Крайнюк В. Н., Саржанов Ф., Стариков С. В., Тарасовская Н. Е., Тимошенко А. Ю., Титов С. В. Ареал озерных лягушек (комплекс Pelophylax ridibundus, Amphibia, Ranidae) // Труды Зоологического института РАН. 2022. Т. 326, № 3. С. 211–238.
Кузякин А. П. Зоогеография СССР // Ученые записки Московского областного педагогического института им. Н. К. Крупской. 1962. Т. 59. С. 3–182. (Биогеография. Вып. 1).
Куперштох В. Л., Трофимов В. А., Равкин Ю. С. Пространственная организация населения птиц // Равкин Ю. С. Птицы лесной зоны Приобья. Новосибирск: Наука. Сиб. отд-ние, 1978. С. 253–269.
Куранова В. Н., Ярцев В. В. Биоразнообразие Томского Приобья. Земноводные и пресмыкающиеся. Томск: Издательский дом Томского гос. ун-та, 2020. 148 с.
Миркин Б. Г. Методы кластер-анализа для поддержки принятия решений: Обзор WP/7/2011/03. М.: Изд. дом НИУ ВШЭ, 2011. 88 с.
Наумов Р. Л. Птицы в очагах клещевого энцефалита Красноярского края: : Автореф. дис. ... канд. биол. наук. М., 1964. 19 с.
Наркевич А. Н., Виноградов К. А., Гржибовский А. М. Многочисленные сравнения в биомедицинских исследованиях: проблема и способы решения // Экология человека. 2020. № 10. С. 54–64.
Равкин Е. С., Челинцев Н. Г. Методические рекомендации по комплексному маршрутному учету птиц. М.: ВНИИприрода, 1990. 33 с.
Равкин Ю. С., Вартапетов Л. Г., Юдкин В. А., Покровская И. В., Богомолова И. Н., Цыбулин С. М., Блинов В. Н., Жуков В. С., Добротворский А. К., Блинова Т. К., Стариков В. П., Ануфриев В. М., Торопов К. В., Соловьёв С. А., Тертицкий Г. М., Шор Е. Л. Пространственно-типологическая структура населения земноводных Западно-Сибирской равнины // Сибирский экологический журнал. 2003а. № 5. С. 603–610.
Равкин Ю. С., Ефимов В. М., Кокорина И. П., Чеснокова С. В., Лялина М. И. Пространственно-временная неоднородность населения пресмыкающихся Западной Сибири // Зоологический журнал. 2023б. Т. 102, № 9. С. 1046–1058.
Равкин Ю. С., Куранова В. Н., Цыбулин С. М., Богомолова И. Н., Юдкин В. А., Торопов К. В., Булахова Н. А., Борисович О. Б., Панов В. В., Добротворский А. К. Численность, распределение и пространственно-типологическая неоднородность населения земноводных и пресмыкающихся в Томской и Новосибирской областях // Амфибии и рептилии в Западной Сибири (сохранение биоразнообразия проблемы экологической этики и экологического образования). Новосибирск: ООО «Ревик-К», 2003б. С. 20–32.
Равкин Ю. С., Ливанов С. Г. Факторная зоогеография. Новосибирск: Наука, 2008. 205 с.
Равкин Ю. С., Лукьянова И. В. География позвоночных южной тайги Западной Сибири. Новосибирск: Наука, 1976. 450 с.
Равкин Ю. С., Одинцев О. А., Богомолова И. Н. Особенности изучения пространственно-типологической изменчивости населения отдельных групп видов птиц и их распределения (на примере врановых) // Сибирский экологический журнал. 2023а. № 1. С. 25-37.
Равкин Ю. С., Цыбулин С. М., Ананин А. А., Ивлиев В. Г., Железнова Т. К., Вартапетов Л. Г., Юдкин В. А., Жуков В. С., Преображенская Е. С., Стишов М. С., Торопов К. В., Равкин Е. С., Богомолова И. Н., Чеснокова С. В., Лялина М. И. Эколого-географическая организация и структура летнего населения птиц Северной Евразии // Журнал общей биологии. 2022. Т. 83, № 4. С. 302–320.
Равкин Ю. С., Цыбулин С. М., Ливанов С. Г., Малков П. Ю., Торопов К. В., Малков Н. П., Грабовский М. А., Швецов Ю. Г., Дубатолов В. В., Малков Ю. П., Бондаренко А. В., Вартапетов Л. Г., Митрофанов О. Б., Вознийчук О. П., Борисович О. Б. Особенности биоразнообразия Российского Алтая на примере модельных групп животных // Успехи современной биологии. 2003в. Т. 123, № 4. С. 409–420.
Равкин Ю. С., Цыбулин С. М., Ливанов С. Г., Торопов К. В., Куранова В. Н., Стариков В. П., Чеснокова С. В. Картографический анализ населения земноводных, пресмыкающихся и птиц Западно-Сибирской равнины и Алтая // Сибирский экологический журнал. 2008. Т. 15, № 5. С. 745–750.
Равкин Ю. С., Цыбулин С. М., Лялина М. И. К оценке качества экстраполяционного прогноза численности наземных позвоночных по ландшафтным аналогам (на примере Шлюзовского лесоболотного комплекса «Сказочный») // Сибирский экологический журнал. 2005. Т. 12, № 3. С. 427–433.
Растительность Западно-Сибирской равнины: Карты / Спец. содерж. разраб. Ин-т географии Сибири и Дальнего Востока СО АН СССР по общ. руковод. акад. В. Б. Сочавы; Сост. и подгот. к печати ф-кой № 4 ГУГК в 1975 г.; Ред. В. Я. Михайленко. 1:1 500 000. М.: ГУГК при СМ СССР, 1976.
Стариков В. П. Кадастровая сводка по земноводным Курганской области // Всесоюзное совещание по проблеме кадастра и учета животного мира. Ч. 2. М., 1986. С. 435–436.
Стариков В. П. Зеленая жаба // Красная книга Курганской области. Курган: Зауралье, 2002. С. 113–114.
Трофимов В. А. Модели и методы качественного факторного анализа матрицы связи // Проблемы анализа дискретной информации. Ч. 2. Новосибирск, 1976. С. 24–36.
Цыбулин С. М., Равкин Ю. С., Жимулёв И. Ф., Панов В. В., Вартапетов Л. Г., Жуков В. С., Богомолова И. Н., Николаева О. Н. Динамика численности и пространственная неоднородность сообществ. Позвоночные // Динамика экосистем Новосибирского Академгородка. Новосибирск: Изд-во СО РАН, 2013. С. 309–318.
Цыбулин С. М., Равкин Ю. С., Панов В. В., Бабуева Р. В. Животный мир. Позвоночные // Природа Академгородка: 50 лет спустя. Новосибирск: Изд-во СО РАН, 2007. С. 166–177.
Эпова Л. А., Куранова В. Н., Бабина С. Г. Видовое разнообразие, биотопическое распределение земноводных и пресмыкающихся заповедника «Кузнецкий Алатау» в градиенте высотной поясности (Юго-восток Западной Сибири) // Вестник Томского государственного университета. Биология. 2013. № 4 (24). С. 77–97.
Яковлев В. А. Кадастр земноводных и пресмыкающихся Республики Алтай // Животный мир Алтае-Саянской горной страны. Горно-Алтайск: РИО «Универ-Принт» ГАГУ, 1999. С. 175–214.
Hammer Ø., Harper D. A. T., Ryan P. D. PAST: Paleontological Statistics Software Package for Education and Data Analysis // Palaeontologia Electronica. 2001. Vol. 4, № 1. P. 1–9.
Jaccard P. Lois de distribution florale dans la zone alpine // Bulletin de la Société Vaudoise des Sciences Naturelles. 1902. Vol. 38, № 144. P. 69–130.
Kuranova V. N., Yakovlev V. A., Simonov E. P., Ischenko V.G., Yartsev V. V., Bogomolova I. N. Diversity, distribution and nature conservation status of Western Siberia Amphibia // Principy ekologii. 2016. Vol. 5, № 3. P. 77.
Litvinchuk S. N., Kazakov V. I., Pasynkova R. A., Borkin L. J., Kuranova V. N., Rosanov J. M. Tetraploid green toad species (Bufo pewzowi) from the Altay Mountains: the first record for Russia // Russian Journal of Herpetology. 2010. Vol. 17, № 4. P. 290–298.
Polunin D., Shtaiger I., Efimov V. JACOBI4 software for multivariate analysis of biological data // bioRxiv. 2019. С. 803684.
Simonov E., Kuranova V. N., Lisachov A., Yartsev V. V., Bogomolova I. N. Database of Amphibia distribution in West Siberia (Russia) // Biodiversity Data Journal. 2022. 10: e82436. DOI: 10.3897/BDJ.10.e82436
Благодарности
Исследования, послужившие основой для написания данной статьи, проведены в основном по программе ФНИ государственных академий на 2021–2025 гг. (проект FWGS-2021-0002).

 © 2011 - 2026
© 2011 - 2026