



Введение
Настоящее сообщение отличает популяционно-экологический подход к анализу состояния, динамики численности, многовидовой организации и структуры населения мелких млекопитающих Северо-Восточного Приладожья. Ландшафты этой территории, относящиеся в основном к зоне таежных лесов и представляющие для многих из изученных видов мелких млекопитающих северную периферию ареала, постоянно испытывают мощнейший антропогенный пресс, причем со стороны не только напряженной лесоэксплуатации, но и других форм интенсивных антропогенных воздействий. Это делает данный регион весьма показательной и удобной экологической моделью сильно нарушенной природной среды, уникальным полигоном для научного анализа, прогнозирования и разработки мер компенсации и вместе с тем доступным тест-объектом, достаточно точным и адекватным индикатором разнообразных экологических нарушений.
Благодаря достаточно высокой численности, видовому разнообразию и экологической лабильности мышевидные грызуны и землеройки, проявляя здесь наиболее рельефные популяционные адаптации в ответ на разнообразные формы внешних воздействий, могут рассматриваться в качестве ценнейшей биологической модели для построения общей теории популяционной организации видов, включая и сформулированную для Фенноскандии (Levins, 1969; Hanski, 1998, 1999; Hanski, Simberloff, 1997), но весьма актуальную и для Карелии, концепцию метапопуляций, как и выявления здесь основных принципов приспособления к жизни у границ ареалов, в т. ч. и разработки методов биоиндикации при оценке направлений и масштабов антропогенных влияний на популяции животных. Вместе с тем изучение экологических особенностей этих видов представляет самостоятельный прикладной интерес, особенно в связи с их эпидемиологическим и лесохозяйственным значением, а также разнообразной биоценотической ролью.
Известно, что популяция как основная экологическая и биохорологическая единица вида служит и конкретной формой его существования, и элементарной единицей эволюции. В связи с этим она приобрела целый комплекс механизмов, обеспечивающих возможность адаптивного ответа популяции как единого целого на любые внешние воздействия. У мелких млекопитающих как животных с относительно низкой индивидуальной стойкостью формируется особенно хорошо выраженная популяционная организация (эффективная групповая стойкость). Это делает их и незаменимым объектом для соответствующих популяционно-экологических изысканий.
Основным содержанием и целью данной работы является всесторонний анализ популяционной экологии изученных видов и их группировок и расшифровка на этой основе специфических и общих экологических механизмов их приспособлений к экстремальным условиям таежного Севера. Популяционный подход к изучению групповых адаптаций и адаптивных комплексов одинаково важен как для обоснования ведущего их значения в формообразовании, так и для решения многих практических вопросов, таких как прогнозирование и регулирование численности животных, участвующих в формировании природных очагов инфекций, повышение биологической продуктивности лесных угодий или охрана природных ландшафтов.
Материалы
Сообщение базируется на стационарных и экспедиционных исследованиях, проводившихся автором и под его руководством на протяжении 60 с лишним лет (1957–2020 гг.) на территории северо-восточного побережья Ладожского озера.
Методы
В основе полученных материалов – результаты массовых количественных учетов мелких зверьков (табл. 1, 2), выполнявшихся двумя стандартными методами – ловушко-линиями с расстановкой каждой из 25–50 давилок Геро и 30-метровыми ловчими канавками с вкопанными в них вровень с дном канавок тремя конусной формы металлическими цилиндрами. Соответственно, за показатель численности принимали число зверьков, попавших за сутки работы в 100 ловушек (на 100 ловушко-суток) и отловленных в конусы за 10 суток работы одной канавки (на 10 канавко-суток), а за индекс доминирования – относительное количество представителей данного вида, выраженное в процентах ко всем отловленным данным способом зверькам.
Таблица 1. Сводные данные о численности и соотношении видов мелких млекопитающих в Приладожье (1966–2014 гг.)
| Вид | Отлов ловушками Геро (309272 ловушко-суток) | Отлов канавками (9177 канавко-суток) | ||||
| абс. число | на 100 ловушко-суток | % | абс. число | на 10 канавко-суток | % | |
| Обыкновенная бурозубка | 5645 | 1.83 | 46.8 | 4708 | 5.13 | 47.06 |
| Средняя бурозубка | 217 | 0.07 | 1.79 | 1210 | 1.32 | 12.1 |
| Малая бурозубка | 324 | 0.10 | 2.56 | 1890 | 2.06 | 18.9 |
| Равнозубая бурозубка | 74 | 0.02 | 0.51 | 87 | 0.09 | 0.8 |
| Крошечная бурозубка | 4 | 0.001 | 0.03 | 60 | 0.07 | 0.63 |
| Водяная кутора | 60 | 0.02 | 0.51 | 115 | 0.13 | 1.12 |
| Лесная мышовка | 110 | 0.004 | 0.10 | 657 | 0.72 | 6.6 |
| Полевая мышь | 18 | 0.0006 | 0.02 | 3 | 0.003 | 0.03 |
| Мышь-малютка | 5 | 0.0002 | 0.005 | 19 | 0.02 | 0.17 |
| Лесной лемминг | 6 | 0.0002 | 0.005 | 176 | 0.19 | 1.74 |
| Рыжая полевка | 4992 | 1.61 | 41.2 | 939 | 1.02 | 9.34 |
| Красная полевка | 220 | 0.07 | 1.79 | 17 | 0.02 | 0.18 |
| Обыкновенная полевка | 3 | 0.001 | 0.03 | 43 | 0.05 | 0.45 |
| Темная полевка | 502 | 0.16 | 4.1 | 327 | 0.036 | 0.33 |
| Полевка-экономка | 53 | 0.02 | 0.51 | 45 | 0.05 | 0.46 |
| Всего | 12233 | 3.907 | 100 | 10296 | 10.909 | 100 |
Таблица 2. Численность и видовое разнообразие мелких млекопитающих в основных типах биотопов
| Биотоп | Учеты ловушками | Учеты канавками | ||||||
| экз. на 100 ловушко-суток | число видов | индекс разнообразия | индекс выровненности | экз. на 10 канавко-суток | число видов | индекс разнообразия | индекс выровненности | |
| Сосняки лишайниковые | 0.51 | 4 | 0.47 | 0.98 | Учеты не проводились | |||
| Сосняки-зеленомошники | 2.68 | 7 | 0.57 | 0.94 | 11.1 | 13 | 0.73 | 0.73 |
| Ельники-зеленомошники | 3.51 | 15 | 0.55 | 0.57 | 21 | 12 | 0.66 | 0.66 |
| Смешанные и лиственные леса | 4 | 12 | 0.56 | 0.62 | Учеты не проводились | |||
| Лиственное мелколесье | 4.89 | 10 | 0.57 | 0.81 | 11.54 | 13 | 0.67 | 0.7 |
| Зарастающая вырубка | 6.3 | 9 | 0.37 | 0.78 | Учеты не проводились | |||
| Луга и другие с/х угодья | 4.61 | 9 | 0.53 | 0.76 | Учеты не проводились | |||
Для оценки степени предпочтения, оказываемого видом тому или иному местообитанию (и его преферентного статуса в сообществе), вычислялся также особый показатель – коэффициент (или индекс) верности биотопу, предложенный сибирскими зоологами (Глотов и др., 1978). Кроме того, для оценки видового разнообразия населения мелких зверьков каждого биотопа определялись показатели разнообразия (Н) и выравненности (е) (Одум, 1975).
Всего за период исследований отработано свыше 400 тыс. ловушко-суток и 10 тыс. канавко-суток. Общее число добытых зверьков превысило 30 тысяч. Из них на долю насекомоядных приходится 63.7 %, грызунов – 36.3 % (см. табл. 1). Еще заметнее эта диспропорция в уловах канавками, где землеройки составляют 79 %, тогда как в добыче ловушками – только 50 %.
Результаты
Согласно полученным данным, группа мелких млекопитающих насчитывает в пределах рассматриваемой территории 21 вид, которые по степени доминирования в суммарных уловах располагаются следующим образом: обыкновенная бурозубка (Sorex araneus L.), рыжая полевка (Myodes (Clethrionomys) glareolus Schreb.), темная полевка (Microtus agrestis L.), малая бурозубка (Sorex minutus L.), средняя бурозубка (Sorex caecutiens Laxm.), лесная мышовка (Sicista betulina Pall.), водяная полевка (Arvicola terrestris L.), красная полевка (Myodes (СL.) rutilus Pall.), лесной лемминг (Myopus schisticolor Pall.), водяная кутора (Neomys fodiens Penn.), равнозубая бурозубка (Sorex isodon Turov), полевка-экономка (М. oeconomus Pall.), мышь-малютка (Micromys minutus Pall.), крошечная бурозубка (S. minutissimus Zimm.), полевая мышь (Apodemus agrarius Pall.), обыкновенная полевка (M. arvalis Pall.), красно-серая полевка (Myodes (Clethrionomys) rufocanus Sund.), серая крыса (Ruttus norvegicus Berk.), домовая мышь (Mus musculus L.), лесная и желтогорлая мыши (Apodemus sylvaticus L. и A. Flavicolus Melch).
При этом наиболее многочисленными видами, как в целом по региону, так и в большинстве обследованных биотопов, оказались обыкновенная бурозубка и рыжая полевка, составившие в наших сборах мелких млекопитающих соответственно 45.3 и 26.3 %, т. е. в сумме почти 72 %. Далее по убыванию численности следуют три вида-содоминанта – малая и средняя бурозубки и темная полевка (9.5, 6.2 и 5.7 %) и три второстепенных вида – лесная мышовка, лесной лемминг и красная полевка (3.2, 0.8 и 0.8 %). На остальные виды приходится всего 2.2 %, и их следует отнести к группе крайне малочисленных.
Такое соотношение видов в общих чертах сохраняется в различные годы, но, как уже говорилось, зависит от способа отлова (табл. 1). В сборах канавками значительно выше как общая доля насекомоядных, так и численность доминирующего и второстепенных видов землероек и грызунов. Например, по сравнению с добычей ловушками удельный вес таких содоминантов, как средняя и малая бурозубки и лесная мышовка, увеличивается в уловах канавками в 5–6 раз, а такие редкие виды, как крошечная бурозубка, кутора и лесной лемминг, и вовсе попадаются только в канавки. Напротив, в сборах давилками преобладают немногие виды мышевидных грызунов и занимающая второе место обыкновенная бурозубка, а целый ряд второстепенных и редких видов вообще не представлен.
Общий показатель учета давилками по региону в целом колеблется по годам от 0.34 до 18.5 на 100 ловушко-суток и составляет в среднем 7.8, канавками – от 2.7 до 25.4, в среднем 14.0 на 10 канавко-суток. Сопоставление этих данных с результатами отлова в других регионах Европейского Севера (Ивантер, 1975) позволяет оценить общую численность мелких млекопитающих в Восточной Фенноскандии как среднюю. В то же время по сравнению с более южными областями она выглядит невысокой (особенно низка численность мышевидных грызунов), что объясняется не только крайней неустойчивостью среды и однообразием кормовой базы, характерными вообще для всего таежного Севера (Шварц, 1963), но и ландшафтными особенностями охваченной учетами территории, в частности господством бедных в экологическом отношении частоствольных ельников, чистых лишайниковых сосняков, моховых болот и хвойно-лиственных молодняков жерднякового типа.
Невысокий средний многолетний показатель учета мелких млекопитающих обусловлен также особым характером многолетней динамики численности как отдельных видов, так и всего населения землероек и грызунов рассматриваемого региона. Как показывают данные специальных учетов, для всех изученных популяций мелких зверьков типичны значительные по размаху, но небольшой частоты и обычно неправильного «рваного» ритма флуктуации, отличительной чертой которых являются длительность и глубина депрессий, редкость и кратковременность подъемов и общая растянутость цикла. Вместе с тем отсутствие четкой синхронности в колебаниях численности различных видов и групп мелких млекопитающих приводит к тому, что в отдельные годы в уловах преобладают то землеройки (1969, 1971, 1975–1977, 1980–1982, 1984–1986, 1989, 1991–1993, 1998–1999, 2002, 2006, 2009, 2011 гг.), то мышевидные грызуны (1964, 1966–1968, 1970, 1973–1974, 1978–1979, 1983, 1990, 1994, 2003, 2007, 2010, 2014 гг.), и аспект фауны меняется в зависимости от особенностей года с «землеройко-полевкового» на «полевко-землеройковый» (рис. 1).
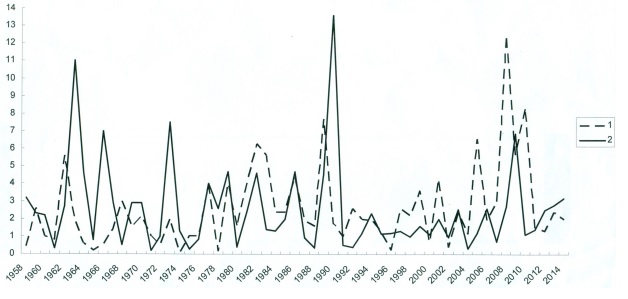
Рис. 1. Суммарная численность мышевидных грызунов (1) и землероек-бурозубок (2) в Карельском Приладожье. По оси абсцисс – годы, по оси ординат – экземпляры на 100 ловушко-суток
Fig. 1. Total number of mouse-like rodents (1) and shrews (2) in the Karelian Ladoga region
Along the abscissa axis – years, along the ordinate axis – specimens per 100 trap-days
Не всегда согласованно колеблется и численность отдельных представителей каждой из этих групп. Однако последнее обстоятельство почти не сказывается на масштабах и характере изменений общей численности зверьков, поскольку население их образовано в основном популяциями лишь двух видов – обыкновенной бурозубки и рыжей полевки. Эти доминантные виды определяют уровень и многолетние колебания численности как всего населения мелких млекопитающих в целом, так и их группировок в отдельных местообитаниях, за исключением лиственных молодняков, сельскохозяйственных угодий и других элементов культурного ландшафта, где кроме обыкновенной бурозубки и рыжей полевки господствует темная полевка, составляющая до 30 % и более всего улова. Таким образом, в отношении видовой структуры население мелких млекопитающих нашего региона представляет довольно стабильную «двухдоминантную» систему (группировку) видовых популяций. Такая структура, как и асинхронность колебаний численности двух господствующих видов, безусловно, целесообразна при ограниченной емкости и однообразии угодий. Она придает населению зверьков необходимую стойкость к воздействию неблагоприятных условий и может рассматриваться как одна из форм адаптаций животных на популяционно-биоценотическом уровне.
Ядро фауны мелких млекопитающих региона составляют широко распространенные по всей лесной полосе виды, такие как обыкновенная бурозубка, рыжая и темная полевки. Эти виды отличаются наиболее высокой и стабильной численностью и занимают доминирующее положение в большинстве природных комплексов. Северо-восточный («таежный» или «сибирский») элемент представляют средняя и крошечная бурозубки, красная и красно-серая полевки и лесной лемминг. Они селятся преимущественно в таежных лесах северного типа, но и здесь не достигают высокой плотности. Хорошо представлены в местной фауне млекопитающие южного происхождения: лесные западноевропейские виды, выходцы из широколиственных лесов и лесостепных пространств. Это малая бурозубка, кутора, лесная мышовка, мышь-малютка, лесная и желтогорлая мыши и обыкновенная полевка. Распространение их ограничено преимущественно Южной Карелией, где они встречаются спорадически, в основном на участках лиственного леса, сенокосных и пахотных угодьях и других элементах культурного ландшафта. Существенное значение имеют виды-убиквисты («космополиты»), распространение которых охватывает несколько ландшафтных зон. К этой группе относятся водяная полевка, полевка-экономка и равнозубая бурозубка. Завершают список три синантропных вида: серая и черная крысы и домовая мышь. Они живут в черте поселков, городах и в естественных угодьях встречаются редко.
В последние столетия формирование таежной териофауны проходит под постоянным воздействием интенсивной хозяйственной деятельности человека. Развитие земледелия, прогрессирующее сокращение лесных площадей и изменение возрастного и породного состава древостоев приводят к трансформации естественных ландшафтов, которые приобретают все более южный облик. Этот процесс стимулирует дальнейшее расселение видов, связанных в своем существовании преимущественно с открытыми стациями и лиственными насаждениями. Все дальше на север проникают лесная мышовка, желтогорлая мышь и мышь-малютка. В то же время обыкновенная полевка и полевая мышь, которые три-четыре десятилетия назад также продвигались к северу, в последние годы вновь отступают на юг. Это вызвано, в частности, резким сокращением посевных площадей под зерновые.
Как известно, приспособление наземных животных к сложно дифференцированной среде происходит не только на организменном и популяционном, но и более высоком интегрированном экосистемном (биоценотическом) уровне. Применительно к мелким млекопитающим это наиболее четко проявляется в приуроченности их населения к определенным биотопам и формировании соответствующих биоценотических группировок. На основе анализа ландшафтных особенностей изучаемого региона и закономерностей пространственного распределения землероек и мышевидных грызунов мы выделили на этой территории семь подобных группировок. Ниже приводится их краткая характеристика, основанная на многолетних количественных учетах и изучении структурных особенностей и динамики смешанного животного населения этих формирований.
Подводя итоги проведенных исследований, необходимо остановиться на некоторых общих закономерностях биотопического и пространственного распределения популяционных группировок и всего населения мелких млекопитающих Приладожья в связи с изменениями, вызванными антропогенным фактором. С этой целью нами проведен кластерный анализ многовидовых биоценотических группировок мелких млекопитающих этого региона (Ивантер, Коросов, 1992), в результате которого были выделены группы биотопов, наиболее близкие по условиям обитания и видовому составу зверьков (рис. 2). Наиболее богаты видами оказались еловые и смешанные леса. Обедненными териокомплексами, представленными в основном политопными видами, характеризуются сосняки-зеленомошники, вырубки и лиственное мелколесье (см. табл. 2). Выявляется близость населения зверьков из двух типов сосняков, но в лишайниковых борах видов еще меньше. Наконец, особое положение занимает биотопический комплекс экотона – границы между лугом и лесом, включающий представителей смежных биотопов. При этом выявленный характер отношений между биоценотическими комплексами млекопитающих сохраняется вне зависимости от метода количественных учетов: и по видовому составу, и по численности (показатель учета) население зверьков сосняков-зеленомошников все же ближе к мелколесью, чем к зеленомошным ельникам. Как показали учеты, наиболее плотно населены мелкими млекопитающими спелые лиственные леса с умеренной хвойной примесью, зарастающие вырубки 3–10-летнего возраста, участки культурного ландшафта, а также травяно-зеленомошные ельники с участием лиственных пород, хорошим подлеском и развитым травостоем. Эти же биотопы отличаются и наиболее богатым видовым спектром мелких зверьков. В лишайниковых борах и монотонных таежных ельниках без лиственной примеси и подлеска численность землероек и грызунов минимальная. Остальные биотопы занимают в этом отношении промежуточное положение. Таким образом, в целом коренные типы лесных угодий существенно уступают производным лесам, элементам культурного ландшафта, вырубкам и другим типам трансформированных человеком площадей как по разнообразию видовой структуры соответствующих биоценотических группировок животных, так и по их численности. Это говорит о том, что антропогенный фактор оказывает на распределение и численность мелких млекопитающих в общем положительное влияние.

Рис. 2. Биоценотические группировки мелких млекопитающих Карельского Приладожья: I – сосняки лишайниковые; II – сосняки-зеленомошники; III – ельники-зеленомошники; IV – ельники заболоченные; V – смешанные хвойно-лиственные леса; VI – спелые лиственные леса; VII – лиственное мелколесье; VIII – вырубки; IX – луга и другие сельскохозяйственные угодья
Fig. 2. Biocenotic groups of small mammals of the Karelian Lake Ladoga region: I – lichen pine forests; II – green moss pine forests; III – green moss spruce forests; IV – swampy spruce forests; V – mixed coniferous-deciduous forests; VI – mature deciduous forests; VII – deciduous half-grown forests; VIII – clearings; IX – meadows and other agricultural lands
Изучение биоценотических группировок мелких млекопитающих Приладожья, их структуры и динамики во времени и пространстве позволяет констатировать своеобразную и неоднозначную роль различных биотопов, в частности коренных и антропогенных ландшафтов, в жизни популяций и всего смешанного населения землероек и грызунов (см. табл. 2). Так, если зеленомошные и травяные леса (сосняки и особенно ельники), как уже отмечалось выше, служат для большинства видов основными местообитаниями круглый год и населены стабильными по составу, хотя и немногочисленными группировками мелких млекопитающих, то производные биотопы, и прежде всего зарастающие вырубки и участки мелколесья, представляют для них временные, хотя и весьма важные местообитания, значение которых повышается лишь в годы подъемов численности. С этим связан выявленный нами специфический характер движения численности видов в коренных и антропогенных биотопах: при общей синхронности многолетних колебаний в антропогенном ландшафте они гораздо более резкие, чем в коренных лесах, где численность популяций более стабильна, хотя и держится на менее высоком уровне. Та же закономерность прослеживается и при анализе сезонных изменений численности зверьков. В коренных древостоях нарастание численности от весны к осени идет обычно более умеренными темпами и равномернее, чем в антропогенном ландшафте, куда при интенсивном размножении популяции в массе выселяются зверьки из соседних лесных биотопов. В результате численность землероек и грызунов возрастает здесь быстрее и резче, причем в тем большей мере, чем активнее протекает репродукция популяции в целом. Напротив, в годы низкого уровня размножения численность зверьков в элементах антропогенного ландшафта увеличивается в основном за счет местного, обычно немногочисленного поголовья, и нарастание ее к концу сезона нередко даже менее выражено, чем в коренных биотопах. Таким образом, в коренных местообитаниях численность мелких млекопитающих более устойчива и колебания ее менее резкие, сглаженные, а в трансформированных, наоборот, население землероек и грызунов крайне динамично, нестабильно и испытывает резкие флуктуации численности по годам и особенно сезонам.
Сезонные изменения численности землероек-бурозубок несколько различаются по срокам и темпам в разные годы и в разных местообитаниях в зависимости от численности и состава населения, состояния, интенсивности размножения и уровня смертности популяции. Вместе с тем общая схема процесса сохраняется: численность зверьков резко нарастает от мая к июлю, достигает максимума в августе – сентябре, резко снижается к ноябрю – декабрю, стабилизируется зимой и вновь сокращается в апреле – мае (годовой минимум).
Уровень видового разнообразия мелких млекопитающих зависит и от плотности их населения (табл. 3, 4). В годы высокой численности группировки более выровненные, а в периоды депрессий отличаются неоднородностью.
Таблица 3. Численность (экз. на 100 ловушко-суток), индекс доминирования (доля в уловах), разнообразие и выровненность в годы с разным уровнем численности мелких млекопитающих по данным учетов ловушками
| Вид | Высокая численность | Средняя численность | Низкая численность | |||
| экз. на 100 ловушко-суток | индекс доминирования | экз. на 100 ловушко-суток | индекс доминирования | экз. на 100 ловушко-суток | индекс доминирования | |
| Обыкновенная бурозубка | 3.76 | 0.395 | 1 | 0.338 | 0.1 | 0.453 |
| Средняя бурозубка | 0.06 | 0.006 | 0.08 | 0.026 | 0 | 0.016 |
| Малая бурозубка | 0.02 | 0.002 | 0.07 | 0.022 | 0.01 | 0.031 |
| Равнозубая бурозубка | - | - | 0.02 | 0.005 | 0 | 0.16 |
| Водяная кутора | 0.03 | 0.003 | 0 | 0.001 | 0 | 0 |
| Лесная мышовка | 0.02 | 0.002 | 0.05 | 0.016 | 0.01 | 0.025 |
| Полевая мышь | 0.02 | 0.002 | - | - | 0 | 0.008 |
| Мышь-малютка | 0.01 | 0.001 | 0 | - | - | - |
| Рыжая полевка | 4.72 | 0.496 | 1.33 | 0.448 | 0.09 | 0.409 |
| Красная полевка | 0.2 | 0.021 | 0.16 | 0.052 | 0.01 | 0.031 |
| Темная полевка | 0.67 | 0.07 | 0.24 | 0.08 | 0.4 | 0.1081 |
| Полевка-экономка | 0.01 | 0.001 | 0.03 | 0.01 | 0 | 0.008 |
| Всего | 9.51 | 1 | 2.96 | 1 | 0.22 | 1 |
| Число видов | 11 | 11 | 10 | |||
| Разнообразие (Н) | 0.47 | 0.61 | 0.67 | |||
| Выровненность (е) | 0.451 | 0.588 | 0.672 | |||
Таблица 4. Численность (экз. на 10 канавко-суток), индекс доминирования (доля в уловах), разнообразие и выровненность в годы с разным уровнем численности мелких млекопитающих по данным учетов канавками
| Вид | Высокая численность | Средняя численность | Низкая численность | |||
| экз. на 10 канавко-суток | индекс доминирования | экз. на 10 канавко-суток | индекс доминирования | экз. на 10 канавко-суток | индекс доминирования | |
| Обыкновенная бурозубка | 6.68 | 0.542 | 3.63 | 0.37 | 3.65 | 0.453 |
| Средняя бурозубка | 1 | 0.81 | 1.18 | 0.12 | 1.88 | 0.016 |
| Малая бурозубка | 1.8 | 0.146 | 1.28 | 0.13 | 1.6 | 0.031 |
| Равнозубая бурозубка | 0.03 | 0.003 | 0.05 | 0.005 | 0.03 | 0.016 |
| Крошечная бурозубка | 0.14 | 0.011 | 0.01 | 0.001 | 0.03 | 0 |
| Водяная кутора | 0.12 | 0.01 | 0.19 | 0.02 | 0.04 | 0.025 |
| Лесная мышовка | 0.48 | 0.039 | 0.55 | 0.056 | 0.87 | 0.008 |
| Мышь-малютка | - | - | 0.05 | 0.005 | - | - |
| Лесной лемминг | 0.11 | 0.009 | 0.42 | 0.043 | - | 0.409 |
| Рыжая полевка | 1.4 | 0.114 | 0.9 | 0.004 | 0.01 | 0.031 |
| Темная полевка | 0.54 | 0.044 | 0.038 | 0.038 | 0.4 | 0.181 |
| Полевка-экономка | - | - | 0.14 | 0.015 | - | 0.008 |
| Число видов | 12 | 14 | 11 | |||
| Разнообразие (Н) | 0.35 | 0.8 | 0.72 | |||
| Выровненность (е) | 0.6 | 0.7 | 0.69 | |||
В пределах одного и того же биотопа размещение землероек и грызунов носит мозаичный характер и сильно зависит от особенностей рельефа, состава и сомкнутости древостоя, развития подлеска и травянистой растительности, степени захламленности, количества и качества корма. В сомкнутых древостоях они тяготеют к рединам и опушкам, а в редкостойных лесах и на окраинах населенных пунктов – к тенистым захламленным участкам с богатым подлеском и группами кустарника. Увеличение стенотопности и выход в стации, затронутые деятельностью человека (вырубки, элементы культурного ландшафта и др.) – черты, характерные для всех видов мелких млекопитающих, живущих в тайге у северных границ распространения (лесная мышовка, темная и обыкновенная полевки, малая бурозубка и др.).
В известной мере такая же тенденция проявляется у рыжей полевки и обыкновенной бурозубки, что позволяет рассматривать ее в качестве имеющей приспособительный смысл географической особенности биотопического размещения всего населения мелких млекопитающих изучаемого региона. Об увеличении стенотопности и «синантропизма» южных видов зверьков при расселении их к северу сообщают Н. П. Наумов (1948), А. Н. Формозов (1948), Н. В. Башенина (1972, 1977) и Дж. Таст (Tast, 1968), отмечали это и мы (Ивантер, 1975, 2018).
Территориальное распределение отдельных видов и всего населения мелких млекопитающих в нашем регионе в целом отличается значительной динамичностью и закономерно трансформируется по сезонам и годам в связи с изменением численности популяций. Весной и в начале лета плотность населения зверьков минимальная и они концентрируются в оптимальных местообитаниях, чаще всего в богатых травяно-зеленомошных ельниках, в смешанных насаждениях по берегам ручьев и рек, на травяных опушках и в пределах культурного ландшафта (захламленные и поросшие кустарником каменные гряды среди полей, обочины дорог, межи и прочие «неудобные земли» на полях и сенокосах). Летом по мере размножения и расселения молодняка отдельные «поселения» постепенно сливаются, и зверьки более или менее равномерно заселяют все подходящие биотопы. Осенью эта дисперсия особенно хорошо выражена, но к зиме животные вновь концентрируются в немногих «поселениях», и распределение их приобретает очаговый характер.
Еще отчетливее годовые изменения в характере биотопического размещения. В годы подъема численности население землероек и грызунов широко растекается по всей территории, они встречаются в самых разнообразных биотопах. В большей степени это характерно для молодых расселяющихся животных, тогда как зимовавшие более консервативны и придерживаются лишь наиболее благоприятных мест обитания. Совершенно иная картина наблюдается при депрессии численности зверьков. В этом случае популяция состоит из отдельных поселений (парцеллярных группировок), приуроченных к немногим лучшим участкам местности, и размещается неравномерно, мозаично. Такие оптимальные участки, получившие название «стации переживания» или «резерваты» вида (Наумов, 1948), обеспечивают возможность нормального существования немногочисленного сохранившегося поголовья даже в условиях пессимума и служат очагами восстановления численности при наступлении благоприятных условий. В монотонных ландшафтах таежной зоны их очень немного, поэтому перепады численности по годам достигают значительной величины, и прежде всего за счет длительных и глубоких депрессий.
Динамическая смена аспектов пространственного распределения с диффузного, равномерного при высокой численности зверьков, на мозаичный, лоскутный, в периоды депрессий имеет важное приспособительное значение и способствует процветанию и устойчивости вида. Этим путем достигается необходимый для репродукции контакт особей, удовлетворение их кормовых и защитных потребностей в условиях резкого сокращения численности, равномерное, без перенаселения и конкуренции, освоение среды обитания и заселение новых территорий при увеличении интенсивности размножения. Таким образом, территориальность является эффективным средством регулирования плотности населения и механизмом гомеостаза популяции.
Высокая лабильность пространственной структуры популяции и закономерная годовая и сезонная смена типов поселения в связи с динамикой численности в равной мере свойственны и всему населению мелких млекопитающих изучаемого региона, и каждому из рассматриваемых видов. Вместе с тем конкретный тип пространственной структуры популяции и процесс ее трансформации во времени имеют у различных видов свои особенности. Например, для популяций куторы, лесной мышовки, полевки-экономки, полевой мыши, мыши-малютки и домовой мыши более характерен инсулярный тип пространственной структуры (по классификации В. Е. Флинта, 1972, 1977), отличающийся стабильно мозаичным распределением поселений; для красной полевки – калейдоскопический (термин Ю. В. Ковалевского и Э. И. Коренберга, 1974), главная особенность которого заключается в регулярном перемещении участков повышенного обилия зверьков; для темной и рыжей полевок и всех видов бурозубок – пульсирующий. При годовой и сезонной перестройке типа поселения с мозаичного на диффузный и обратно сохраняется сеть постоянных элементарных поселений в стациях переживания. Все это отражает смешанный характер фауны, разнообразие ландшафтов и динамичность природных условий Карелии. Но поскольку пульсирующий тип пространственной структуры популяции присущ большинству таежных видов мелких млекопитающих, и притом видам наиболее многочисленным и широко распространенным, можно, следовательно, говорить о специфичности его для всего населения землероек и грызунов лесной зоны.
Прогрессирующее антропогенное воздействие на таежные природные комплексы Восточной Фенноскандии привело к характерным изменениям в составе животного населения этого своеобразного региона, в т. ч. и такого важного компонента биоценоза, как мелкие млекопитающие. Наряду с приобретением фауной более южного облика в результате появления новых западноевропейских видов (полевая мышь, мышь-малютка, обыкновенная полевка и др.) и увеличения численности и расширения ареалов южных форм, заселивших Карелию в прошлом (рыжая полевка, малая бурозубка, лесная мышовка), сокращаются местообитания, пригодные для существования типичных северотаежных видов. При современных темпах хозяйственного освоения региона многие из этих животных не успевают приспосабливаться к меняющейся экологической обстановке, что влечет за собой сокращение численности и области распространения и в итоге – постепенное исчезновение ряда представителей сибирского таежного комплекса: лесного лемминга, красной и красно-серой полевок, средней бурозубки и др. Все это приводит к перестройке видовой структуры современной фауны мелких млекопитающих региона и к становлению нового фаунистического комплекса, характеризующегося более южным колоритом (в частности, преобладанием в его составе фаунистических элементов широколиственных лесов), качественной бедностью (за счет выпадения ряда северотаежных видов) и несущего отчетливые черты разнообразных и весьма существенных антропогенных воздействий (рис. 3).
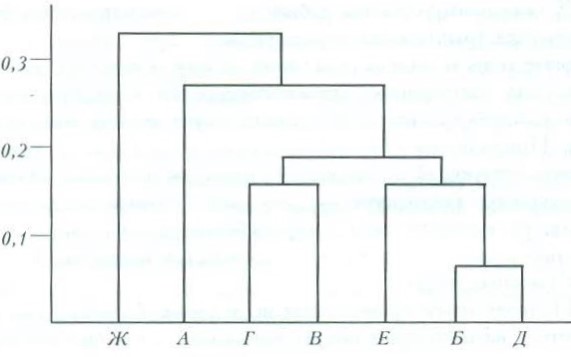
Рис. 3. Дендрограмма сходства биотопов по видовому составу мелких млекопитающих. По оси абсцисс – биотопы: А – лишайниковые сосняки, Б – сосняки зеленомошные, В – ельники, Г – спелые лиственные и смешанные леса, Д – лиственное мелколесье, Е – молодые зарастающие вырубки, Ж – граница сеяного луга с ольшаником. По оси ординат: мера различия между биотопами (коэффициент сходства Съеренсена)
Fig. 3. Dendrogram of similarity of biotopes by species composition of small mammals. Along the abscissa axis – biotopes: А – lichen pine forests, Б – green moss pine forests, В – spruce forests, Г – mature deciduous and mixed forests, Д – deciduous undergrowth, Е – young overgrown clearcuts, Ж – boundary of sown meadow with alder forest. Along the ordinate axis: measure of difference between biotopes (Sorensen's similarity coefficient)
Заключение
Подводя итоги выполненным исследованиям, следует прежде всего подчеркнуть главный полученный нами вывод: выявленные нами пути приспособления мелких млекопитающих к среде обитания отличаются большой сложностью и многообразием и в каждом конкретном случае определяются как спецификой окружающей обстановки, так и популяционно-экологическими особенностями самих животных. Процесс адаптации затрагивает все уровни их организации – от молекулярного до биоценотического и имеет определенные различия в разных условиях существования у разных видов и популяций. Мелкие млекопитающие представляют особую жизненную форму, характеризующуюся сравнительно коротким жизненным циклом, высоким уровнем обмена веществ, несовершенством терморегуляции и повышенной чувствительностью к внешним воздействиям. Низкую индивидуальную стойкость и повышенную в связи с этим смертность от абиотических факторов они вынуждены компенсировать интенсификацией размножения, адаптивной гибкостью и в итоге увеличением групповой стойкости. Последнее достижимо только на пути приобретения совершенной популяционной организации, вот почему основным содержанием приспособительной эволюции мелких видов млекопитающих служит выработка групповых приспособительных реакций и разнообразных популяционных адаптаций, позволяющих им, несмотря на значительные потери вследствие слабой индивидуальной резистентности, успешно осваивать экстремальную среду обитания периферических зон ареала.
Предпринятый популяционно-экологический анализ многовидового населения мелких млекопитающих Северо-Восточного Приладожья показал высокую адаптированность этих животных к местным условиям и выявил у них целый комплекс специфических групповых приспособлений, многие из которых хотя и не являются исключительной прерогативой таежных популяций, но проявляются здесь наиболее ярко и своеобразно. Условия северной периферии ареала с полным основанием можно отнести к экстремальным, и несмотря на что освоение их подчас проходит у отдельных видов мелких млекопитающих разными путями, главные направления адаптаций имеют общие особенности, присущие всем или большинству представителей этой группы животных. Так, для многих грызунов и землероек характерно увеличение стенотопности и выход в стации, затронутые активной деятельностью человека. Экологическая структура северных популяций отличается динамичностью и закономерно перестраивается по сезонам и годам в связи с динамикой численности. При этом адаптивная стратегия периферических популяций постоянно находится в состоянии становления, и то обстоятельство, что полной приспособленности так и не достигается, определяет постоянную готовность видов к микроэволюционным перестройкам в ответ на изменения среды.
Библиография
Башенина Н. В. Основные пути адаптации мышевидных грызунов (Myomorpha, Rodentia) : Автореф. дис. ... д-ра биол. наук. Свердловск, 1972. 66 с.
Башенина Н. В. Пути адаптаций мышевидных грызунов . М.: Наука, 1977. 294 с.
Глотов И. Н., Ердаков Л. Н., Кузякин В. А. и др. Сообщества мелких млекопитающих Барабы . Новосибирск: Наука, 1978. 231 с.
Ивантер Э. В. Очерки популяционной экологии мелких млекопитающих на северной периферии ареала . М.: Товарищество научных изданий КМК, 2018. 770 с.
Ивантер Э. В. Популяционная экология мелких млекопитающих таежного Северо-Запада СССР . Л.: Наука, 1975. 244 с.
Ивантер Э. В., Коросов А. В. Основы биометрии: введение в статистический анализ биологических явлений и процессов : Учеб. пособие. Петрозаводск: Изд-во ПетрГУ, 1992. 168 с.
Ковалевский Ю. В., Коренберг Э. И. Пространственная структура популяций лесных мышевидных грызунов в природных очагах болезней человека // Природноочаговые инфекции и инвазии. Вильнюс, 1974. С. 112–117.
Наумов Н. П. Очерки сравнительной экологии мышевидных грызунов . М.; Л.: Изд-во АН СССР, 1948. 293 с.
Одум Ю. Основы экологии . М.: Мир, 1975. 740 с.
Флинт В. Е. Пространственная структура популяций мелких млекопитающих . М.: Наука, 1977. 182 с.
Флинт В. Е. Типы пространственной структуры популяций у мелких млекопитающих // Популяционная структура вида у млекопитающих. М.: Изд-во МГУ, 1972. С. 161–163.
Формозов А. Н. Мелкие грызуны и насекомоядные Шарьинского района Костромской области в период 1930–1940 гг. // Материалы по грызунам. М.: Изд-во МОИП, 1948. Вып. 3. С. 3–110.
Шварц С. С. Пути приспособления наземных позвоночных животных в условиях существования в Субарктике. Т. 1. Млекопитающие . Свердловск, 1963. Вып. 33. 133 с. (Труды Института биологии Уральского филиала АН СССР).
Hanski I. Metapopulation dynamics // Nature. 1998. Vol. 396 (6706). P. 41–49.
Hanski I. Metapopulation ecology. Oxford; New York: Oxford University Press, 1999. 313 p.
Hanski I., Sinberloff D. The metapopulationapproach, its history, conceptual domain, and appliea-tion to conservation // Metapopulation Biology: Ecology, Genetics and Evolution. Academic Press, 1997. P. 5–26.
Levins R. Some demographic and genetic consequences of environmental heterogeneity for biological control // Bulletin of the Entomological Society of America. 1969. Vol. 15 (3). P. 237–240.
Tast J. Influence of the root vole (Microtus oeconomus Pallas) upon the habitat selection of the field vole (Microtus agrestis L.) in Northern Finland // Suomalais tiedekat. 1968. Vol. A4, No 136. P. 1–23.

 © 2011 - 2026
© 2011 - 2026