



Введение
В умеренных районах Северного полушария один из наиболее распространенных видов рода мятлик (Poa L.) – Poa pratensis L. (Poaceae), мятлик луговой – многолетний корневищный дерновинный злак. Засухоустойчив, зимостоек, теневынослив. Почвы предпочитает рыхлые, плодородные, умеренно увлажненные (Медведев, Сметанникова, 1981). Факультативный апомикт (Кайбелева и др., 2016). Ценный компонент луговых и лугово-степных фитоценозов, увеличение площади которых важно не только в связи с сохранением биоразнообразия и восстановлением естественной растительности нарушенных территорий (Clewell, Aronson, 2006; Funk et al., 2008; Świerszcz et al., 2024), но и для оптимизации агроландшафтов с целью уменьшения экологической напряженности (Беленков и др., 2023). Благодаря устойчивости и долголетию – мятлик сохраняется в травостое более 10 лет (Тимошкин, Тимошкина, 2023) – P. pratensis L. не только одно из наиболее востребованных пастбищных растений, но и широко применяется в составе травосмесей для различных агрофитоценозов (Вахрушев и др., 2023). Прогнозирование и регуляция развития мятликовых сообществ имеет прямое хозяйственное значение.
Наиболее частые ценозообразующие виды травянистых растений на деградированных лесных почвах Кемеровской области (Загурская, Уфимцев, 2022):
- Filipendula ulmaria (L.) Maxim. (лабазник вязолистный) – доминант климаксовых сообществ;
- Chamaenerion angustifolium (L.) Scop. (иван-чай узколистный) – доминант послепожарных суходольных сообществ;
- Cirsium arvense integrifolium & Grab. = Cirsium setosum (Willd.) Besser ex M. Bieb. (бодяк щетинистый) – доминант раннесукцессионных залежных сообществ;
- Poa pratensis (мятлик обыкновенный) – один из основных доминант луговых сообществ.
Направление развития сообществ может нарушаться при внедрении инвазионных видов, например Solidago canadensis L., обладающего огромным конкурентным потенциалом (Загурская, 2022; Świerszcz et al., 2024).
При изучении процессов восстановления растительности на нарушенных почвах ключевыми вопросами являются механизмы взаимодействия потенциальных доминант и субдоминант травянистых сообществ на различных стадиях сукцессии при самовозобновлении растительности нарушенных экотопов (Загурская, Уфимцев, 2022). Отмечено большее ингибирующее влияние деревьев Pinus sylvestris L. по сравнению с Acer negundo L. на растения P. pratensis в подкроновом пространстве этих пород (Цандекова, Колмогорова, 2022). Показан положительный эффект стимуляторов роста и гуминовых удобрений на биометрические показатели мятлика (Филимонова, Тазина, 2022).
Генеративный потенциал вида – наиболее важное обстоятельство на этапе расселения вида и захвата территории. На семенную продуктивность особей влияют не только эдафические, но и биотические факторы, в частности наличие конкуренции с растениями других видов.
Цель работы – изучить влияние фитомассы растений S. canadensis и аборигенных доминант основных травянистых сообществ деградированных лесных почв на развитие генеративных побегов и формирование соплодий (метелок) у растений P. pratensis.
Материалы
Семена P. pratensis 18 мая 2022 г. сеяли в ящики размером 40 × 50 см, заполненные гомогенизированной серой лесной почвой, которые размещали на экспериментальном участке под открытым небом (один ящик = вариант эксперимента, итого 5 ящиков). Посев осуществляли поперечными рядами (длина 40 см) по 4 на ящик, около 1 г (3–4 мл) семян на ряд. Агротехника: полив в начале вегетации и прополка по мере необходимости, поверхностное рыхление почвы.
После формирования у опытных растений четвертого настоящего листа по всей поверхности почвы в ящике вносили свежую, измельченную до 1–2 см2 надземную массу растений S. canadensis (SC), F. ulmaria (FU), C. setosum (CS), Ch. angustifolium (CA) без жестких стеблей, плодов, больных или поврежденных частей. Первая обработка – 13 июля, периодичность 10–14 дней. При внесении растительного материала по мере необходимости удаляли грубые частицы и спрессованные остатки старой мульчи.
В конце августа (через неделю после последнего внесения) отмечены признаки завершения вегетации, 31 августа 2022 г. отобраны образцы для исследования методом высечек из каждого ряда размером 5 × 10 см на глубину почвенного слоя около 10 см. Влияние фитомассы оценивали в сравнении с контролем (без внесения материала чужеродных растений).
Между растениями повторностей (рядов) в пределах вариантов (ящиков) статистически значимых различий не обнаружено. Это предоставляет возможность последующего анализа объединенных выборок.
Стадия развития генеративных побегов существенно варьировала. Определение конкретных стадий для отдельных экземпляров не оправдано методически (при данном количестве образцов корректная статистическая обработка множества групп невозможна), поэтому одновременно приводится название обеих стадий развития: соплодия (соцветия) и т. п.
Методы
Для оценки растений использовали количественный, линейный и гравиметрический методы.
Для сравнения отобраны следующие параметры:
- количество генеративных растений,
- количество генеративных побегов,
- суммарная высота ГП на растении,
- средняя высота ГП,
- масса генеративной части побега,
- масса генеративной части растения.
Статистическую обработку полученных результатов проводили в программе JASP (© The JASP Team) с использованием методов анализа для выборок с различными типами распределения данных (Boxplot, однофакторный дисперсионный анализ, Letter-Based Grouping, критерий Краскела – Уоллиса). Во всех случаях критический уровень значимости принимался равным 0.05.
Результаты
По данным ряда авторов, P. pratensis достигает генеративной стадии (полного развития растений) на 2–4-й год вегетации, в первый сезон образуя обильные вегетативные побеги (Медведев, Сметанникова, 1981; Луганская, Лукиных, 2019). Это хорошо согласуется с результатами, полученными нами в 2021 г. (рис. 1.4): за 110 дней вегетации всего пять экземпляров в трех вариантах эксперимента достигли генеративной стадии, причем все соцветия были недоразвиты (развернулась метелка только на одном экземпляре). Однако в эксперименте 2022 г. на 111-й день плодоносящие растения первого года жизни были обнаружены во всех исследуемых группах (рис. 1, 2, таблица). Как известно, на репродуктивные процессы P. pratensis оказывает влияние продолжительность светового дня (Кайбелева и др., 2016), интенсивность инсоляции, гидротермический режим, а также другие факторы.

Рис. 1. Фактура эксперимента и морфологическое разнообразие растений полученных растений P. pratensis в 2022 г. Здесь и далее: Solidago canadensis (SC), Filipendula ulmaria (FU), Cirsium setosum (CS), Chamaenerion angustifolium (CA); 1 – примеры растений P. pratensis; 2 – ящики с растениями после внесения чужеродной фитомассы; 3 – развитие растений (на примере контроля): 3.1 – до начала опыта, 3.2 – перед сбором образцов; 4 – растения P. pratensis предыдущего сезона 6 сентября 2021 г. (завершение вегетации)
Fig. 1. The texture of the experiment and the morphological diversity of the obtained plants of P. pratensis in 2022. Hereafter: Solidago canadensis (SC), Filipendula ulmaria (FU), Cirsium setosum (CS), Chamaenerion angustifolium (CA); 1 – examples of P. pratensis plants; 2 – boxes with plants after the introduction of foreign phytomasses; 3 – plant development (using the example of control): 3.1 – before the start of the experiment, 3.2 – before collecting samples; 4 – P. pratensis plants of the previous season on September 6, 2021 (end of the growing season)
Развитие растений P. pratensis до внесения чужеродной фитомассы характеризовалось высокой внутренней изменчивостью, но было сходно во всех вариантах. После разделения отобранных для изучения куртин получено от 46 до 36 растений в каждом варианте эксперимента. Из числа образцов исключали слишком мелкие (неразвитые) и поврежденные экземпляры: в итоговых выборках оказалось по 33 растения на каждый вариант.
Не все экспериментальные растения к моменту сбора достигли генеративной стадии (таблица, рис. 2). При этом наличие и степень сформированности генеративных побегов однозначно не соотносились с размерами растений или особенностями развития их вегетативных частей.
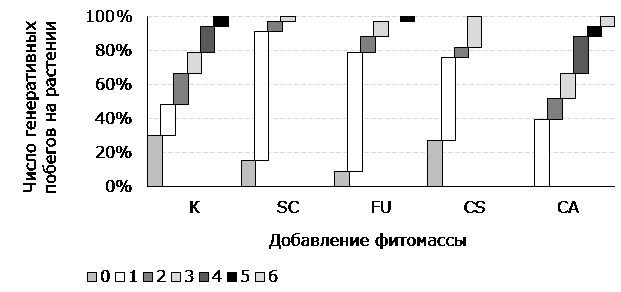
Рис. 2. Соотношение растений с различным количеством генеративных побегов при внесении фитомассы растений-конкурентов (%). Здесь и далее: контроль (К). Цветовое кодирование количества генеративных побегов на растении отражено в легенде
Fig. 2. The ratio of plants with a different number of generative shoots when applying phytomass of competing plants (%). Here and further: control (K). The color coding of the number of generative shoots on a plant is reflected in the legend
Только при обработке Ch. angustifolium плодоносили все модельные растения. Они же отличались максимальным числом генеративных побегов на растение и наибольшей массой зрелых метелок.
Минимальная масса соплодий, приведенная в таблице, может свидетельствовать только о степени зрелости наименьшего из образцов и не характеризует данную выборку без учета дополнительных показателей, в частности количества генеративных побегов на одно растение.
Показатели развития генеративной части P. pratensis при поверхностном внесении фитомассы конкурентных растений в 2022 г.
| Показатель | Тип данных | К | SC | FU | CS | CA |
| Количество генеративных растений, шт. (%) | Число (проценты) | 23 (70) | 28 (85) | 30 (91) | 24 (73) | 33 (100) |
| Количество генеративных побегов (ГП), шт. (%) | Число (проценты) | 70 (80) | 37 (43) | 46 (53) | 47 (54) | 87 (100) |
| Суммарная высота ГП на растении, мм | Mean ± SD
Med(Min–Max) |
290 ± 257 278(0–821) | 255 ± 160 285(0–644) | 329 ± 253 305(0–1238) | 269 ± 257 263(0–847) | 534 ± 382 447(75–1345) |
| Средняя высота ГП, мм | Mean ± SD
Med(Min–Max) |
115 ± 85 138(0–273) | 226 ± 124 238(0–412) | 233 ± 109 250(0–385) | 168 ± 121 212(0–324) | 185 ± 72 188(20–332) |
| Масса генеративной части побега, мг | Mean ± SD
Med(Min–Max) |
14.1 ± 12.8 14.9(0–50.8) | 30.1 ± 18.2 33.6(0–63.0) | 29.7 ± 15.4 28.2(0–64.6) | 18.6 ± 15.6 21.2(0–55.0) | 29.9 ± 17.9 25.7(6.2–86.6) |
| Масса генеративной части растения, мг | Mean ± SD
Med(Min–Max) |
32.8 ± 27.9 31.4(0–88.1) | 33.0 ± 19.8 35.5(0–63.0) | 38.6 ± 21.7 36.5(0–79.5) | 28.4 ± 27.3 22.4(0–95.20) | 74.6 ± 53.8 64.2(6.2–191.4) |
Наименьшее число генеративных растений и масса плодов отмечены в контроле, однако для этих растений характерна значительная вариабельность количества генеративных побегов на растение и относительная выровненность зрелости плодов. Для остальных проанализированных образцов отмечается снижение количества генеративных побегов на одно растение по сравнению с необработанными растениями, однако общее количество плодоносящих особей и большинство весовых показателей превышают аналогичные в контроле.
При измерении растений также было отмечено, что у изученных растений различались размеры и масса зрелого соплодия (метелки светло-желтого или светло-бежевого цвета без элементов цветка, зерновки относительно крупные, твердые, не осыпавшиеся, не поврежденные). В контроле и при добавлении надземной массы C. setosum оказалось минимальное количество таких образцов (11 и 10 соответственно). На рис. 3 представлены результаты по десяти максимальным значениям в выборках.
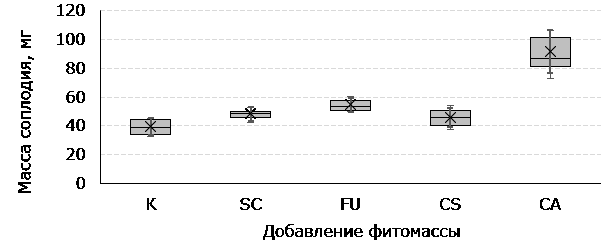
Рис. 3. Вариабельность массы зрелых соплодий P. pratensis под влиянием фитомассы растений-конкурентов: коробка – 1 и 3 квартиль, перекладина – медиана, планки погрешностей – минимальное и максимальное значения, точка – среднее арифметическое, планки погрешностей (тонкая линия) – стандартное отклонение
Fig. 3. Variability of the mass of mature P. pratensis coplodia under the influence of phytomass of competing plants: box – 1 and 3 quartiles, crossbar – median, error bars – minimum and maximum values, point – arithmetic mean, error bars (thin line) – standard deviation
Обсуждение
По изученным показателям отмечены статистически значимые различия для растений P. pratensis, выращенных при внесении фитомассы C. angustifolium: количество генеративных растений и генеративных побегов на растениях, суммарная высота генеративных побегов, суммарная масса генеративных частей на растении. Вес зрелых метелок мятлика, развивавшихся при поверхностном внесении надземных частей C. angustifolium, существенно превосходит остальные. У растений в других вариантах различия не так значительны, следует отметить только то, что контрольные растения обладали наименьшей массой соплодий.
При изучении морфологических показателей для отдельных побегов, а не суммарно по растениям, значимые расхождения отмечены только для контрольных растений (минимальные значения).
Согласно имеющимся данным, распределение приоритетов между вегетативными и генеративными процессами у P. pratensis может определяться наличием ресурсов, в том числе количеством доступного азота в окружающей среде (Li et al., 2024), а также их ограничением и другими стрессовыми факторами (Malyshev, Henry, 2012; Pertierra et al., 2013). То есть при внесении фитомассы C. angustifolium создаются наиболее благоприятные условия для ускоренного развития и плодоношения P. pratensis.
Контрольные экземпляры к моменту сбора характеризовались наименьшей степенью созревания семян (масса соплодий), при этом по количеству генеративных побегов (как суммарно, так и в среднем на одно растение) уступают только развивающимся под воздействием C. angustifolium. Это свидетельствует о вероятном замедлении плодообразования у данных растений (на момент измерения зрелости достигли менее 7 % побегов), однако их репродуктивный потенциал достаточно велик по сравнению с большинством остальных вариантов.
К сожалению, постановка нашего эксперимента не предусматривала обязательный контроль почвенных показателей в ходе эксперимента, в связи с чем затруднительно оценить влияние внесения растительных компонентов на изменение качества почвы. Косвенными свидетельствами протекания этих процессов является уплотнение (для Filipendula ulmaria и Cirsium setosum) или разрыхление субстрата (Chamaenerion angustifolium) относительно контроля в конце сезона. Полученные нами результаты совпадают с литературными данными для контрольных растений, поскольку практически все онтогенетические исследования получены в условиях полевого эксперимента с обязательными агрохимическими процедурами, в т. ч. прополкой. В естественных условиях, скорее всего, смена фаз развития и успешность генеративных процессов будут зависеть от совокупности действия различных факторов, в первую очередь от плодородности почв и видового состава растительных сообществ.
Заключение
Поверхностное внесение в посевы P. pratensis фитомассы других растений не только существенно ускоряет созревание семян, но также может существенно увеличивать генеративный потенциал мятлика (например, под влиянием C. angustifolium). Вероятно, данные эффекты объясняются изменением структуры и свойств почвы, аналогично действию растений-сидератов.
Библиография
Беленков А. И., Киричкова И. В., Габуншина А. А. Агроэколого-хозяйственный баланс и оптимизация природопользования на территории Камышинского района Волгоградской области // Экологический вестник Северного Кавказа. 2023. Т. 19, № 1. С. 60–67.
Вахрушева В. В., Прядильщикова Е. Н., Симонов Г. А. Продуктивность пастбищного фитоциноза в неблагоприятных погодных условиях // Эффективное животноводство. 2023. № 2 (184). С. 64–68. DOI: 10.24412/cl-33489-2023-2-64-68
Загурская Ю. В. Основные аспекты изучения инвазивных видов рода Solidago // Трансформация экосистем. 2022. Т. 5, № 2. С. 1–14. DOI: 10.23859/estr-211029
Загурская Ю. В., Уфимцев В. И. Влияние травяных настоев на прорастание семян Solidago canadensis и доминантных растений деградированных лесных почв // Экосистемы. 2022. Вып. 31. C. 95–101.
Кайбелева Э. И., Куренная Т. Е., Юдакова О. И. Влияние продолжительности фотопериода на проявление апомиксиса у Poa pratensis L. // Бюллетень Ботанического сада Саратовского государственного университета. 2016. Т. 14, вып. 2. С. 71–76.
Луганская С. Н., Лукиных Г. Л. Морфобиологическая характеристика многолетних злаковых трав, используемых для создания газонов в условиях Среднего Урала: Метод. указания . Екатеринбург: УГЛТУ, 2019. 35 с.
Медведев П. Ф., Сметанникова А. И. Кормовые растения европейской части СССР: Справочник . Л.: Колос. Ленингр. отд-ние, 1981. 336 с.
Тимошкин О. А., Тимошкина О. Ю. Многолетние травы для создания газонов в условиях лесостепи Среднего Поволжья // Кормопроизводство. 2023. № 1. С. 16–21. DOI: 10.25685/KRM.2023.47.76.001
Филимонова М. А., Тазина С. В. Исследование эффективности применения гуминовых удобрений в сочетании со стимуляторами роста на мятлике луговом // Вестник ландшафтной архитектуры. 2022. № 29. С. 73–76.
Цандекова О. Л., Колмогорова Е. Ю. Роль антиоксидантов в механизмах адаптации Poa pratensis L. К влиянию древесных растений // Бюллетень Государственного Никитского ботанического сада. 2022. № 144. С. 132–138. DOI: 10.36305/0513-1634-2022-144-132-138
Clewell A. F., Aronson J. Motivations for the Restoration of Ecosystems // Conservation Biology. 2006. Vol. 20, issue 2. P. 420–428. DOI: 10.1111/j.1523-1739.2006.00340.x
Funk J. L., Cleland E. E., Suding K. N., Zavaleta E. S. Restoration through reassembly: Plant traits and invasion resistance // Trends in Ecology & Evolution. 2008. Vol. 23, issue 12. P. 695–703. DOI: 10.1016/j.tree.2008.07.013
Li D., Wang J., Chen R., Chen J., Zong J., Li L., Hao D., Guo H. Nitrogen acquisition, assimilation, and seasonal cycling in perennial grasses: review // Plant Science. 2024. Vol. 342. P. 112054. DOI: 10.1016/j.plantsci.2024.112054
Malyshev A. V., Henry H. A. L. Frost damage and winter nitrogen uptake by the grass Poa pratensis L.: consequences for vegetative versus reproductive growth // Plant Ecology. 2012. Vol. 213. P. 1739–1747. DOI: 10.1007/s11258-012-0127-0
Pertierra L. R., Lara F., Benayas J., Hughes K. A. Poa pratensis L., current status of the longest-established non-native vascular plant in the Antarctic // Polar Biology. 2013. Vol. 36. P. 1473–1481. DOI: 10.1007/s00300-013-1367-8
Świerszcz S., Czarniecka-Wiera M., Szymura T. H., Szymura M. From invasive species stand to species-rich grassland: Long-term changes in plant species composition during Solidago invaded site restoration // Journal of Environmental Management, 2024. Vol. 353. P. 120216. DOI: 10.1016/j.jenvman.2024.120216
Благодарности
Работа выполнена в рамках проекта гос. задания отдела Кузбасский ботанический сад ФИЦ УУХ СО РАН № АААА-А21-121011590010-5.

 © 2011 - 2026
© 2011 - 2026