



Введение
Активное хозяйственное освоение водных экологических систем в XX в. привело к становлению проблемы биологических инвазий, т.е. миграции и вселению гидробионтов за пределы своих «материнских» ареалов (Биологические инвазии…, 2004; Стерлигова, Ильмаст, 2009). К антропогенно-обусловленным причинам расселения видов относятся преобразование среды обитания или изменение ее экологических характеристик, целенаправленная интродукция и реинтродукции видов, а также случайное вселение (Биологические инвазии..., 2004).
Для водных экологических систем данная проблема наиболее актуальна, поскольку при их трансформации устраняются географические барьеры для распространения обитающих в них гидробионтов. В результате данного процесса возможно изменение состава и структуры рассматриваемых сообществ, что приводит к экологическим и экономическим последствиям (Lodge, 1993).
Одним из интересных аспектов исследования инвазионного процесса является изучение фенотипического разнообразия чужеродных видов. Известно, что внутрипопуляционная изменчивость количественных признаков является результатом сложной системы взаимодействия генофонда группы и условий конкретной ландшафтно-географической зоны (Кирпичников, 1987; Павлов, 2007). Сильно различающаяся численность стартовых группировок (от нескольких единиц до сотен тысяч) при разных путях инвазии, а также воздействие естественного отбора в конечном итоге формируют определенное фенотипическое разнообразие популяций чужеродных видов, которое представляет основу для их возможной дифференциации.
В бассейне реки Вычегда, являющейся одной из крупнейших рек Республики Коми, в XX–XXI вв. произошла натурализация белоглазки Ballerus sapa (Pallas, 1814). Основными источниками вселения здесь явились гидротехнические сооружения, среди которых наибольшую роль в процессе расселения сыграли каналы, соединяющие бассейны крупнейших речных систем (Новоселов, 2000).
Иной коридор инвазий представлен различными искусственными водоемами, среди которых, помимо водохранилищ, есть даже незначительные по площади пожарные водоемы и карьеры (Захаров, Бознак, 2011; Рафиков 2016). Так, например, в пожарных водоемах дачных обществ в нескольких километрах от г. Сыктывкара обнаружен ротан-головешка Perccottus glenii Dybowski, 1877, вселенный туда аквариумистами или рыбаками-любителями в 90-х годах прошлого столетия (Бознак, 2004).
Неоднородность условий формирования популяций рассматриваемых видов и разная численность их стартовых группировок позволяют нам предположить, что показатели морфологического разнообразия и стабильность развития особей в популяциях ротана-головешки и белоглазки могут отличаться.
Наблюдения за процессами инвазии дают возможность выявить происходящие изменения рыбной части сообщества крупных речных систем европейского северо-востока России, что необходимо для прогнозирования возможных экологических и экономических последствий. Изучение биологических особенностей популяций чужеродных видов, характеризующихся разными механизмами (способами) инвазии, позволит оценить процесс адаптации вида к новым условиям обитания, внести вклад в понимание причин и направленности микроэволюционного процесса при расселении видов.
Цель данной работы – описать фенотипическое разнообразие популяций белоглазки и ротана-головешки в пределах бассейна р. Вычегда.
Материалы
Материал, использованный для данной работы, собирался крючковыми и сетными орудиями лова в нескольких малых прудах (район расположения – N 61°40'56.8" E 50°45'48.0") на территории дачных массивов с 2012 по 2014 г., а также русловой части р. Вычегда в районе г. Сыктывкара с 2022 по 2024 г. Объем обработанного материала составил: две выборки по 30 экз. ротана-головешки и 31 экз. белоглазки. Также для сравнительного анализа использована выборка леща в количестве 34 экз. из того же района отлова (рис. 1).
Площадь малых прудов, где обитает ротан, составляет от 3 до 6 га, грунт песчанный с наилком. Вдоль берегов произрастают лиственные и хвойные породы деревьев, а также отмечены заросли рогоза и различных видов осок. Вода имеет коричневатую окраску с прозрачностью около 0.9 м по диску Секки. По результатам гидрохимического анализа, проведенного на базе лаборатории «Экоаналит» Института биологии Коми НЦ УрО РАН, вода относится к гидрокарбонатно-кальциевому классу 1-го типа с повышенным содержанием ионов натрия и калия. Минерализация составила до 300 мг/л, что в два раза выше, чем в пойменных водоемах бассейна среднего течения р. Вычегда. В летний период отмечено большое количество нитчатых водорослей, покрывающих собой дно водоема, а также зарастание поверхности водного зеркала исследованных прудов ряской малой Lemna minor L.
Более высокая минерализация, содержание азота и фосфора, а также интенсивное развитие водорослей свидетельствуют о происходящем антропогенном эвтрофировании указанных водных объектов (Рафиков, 2018).
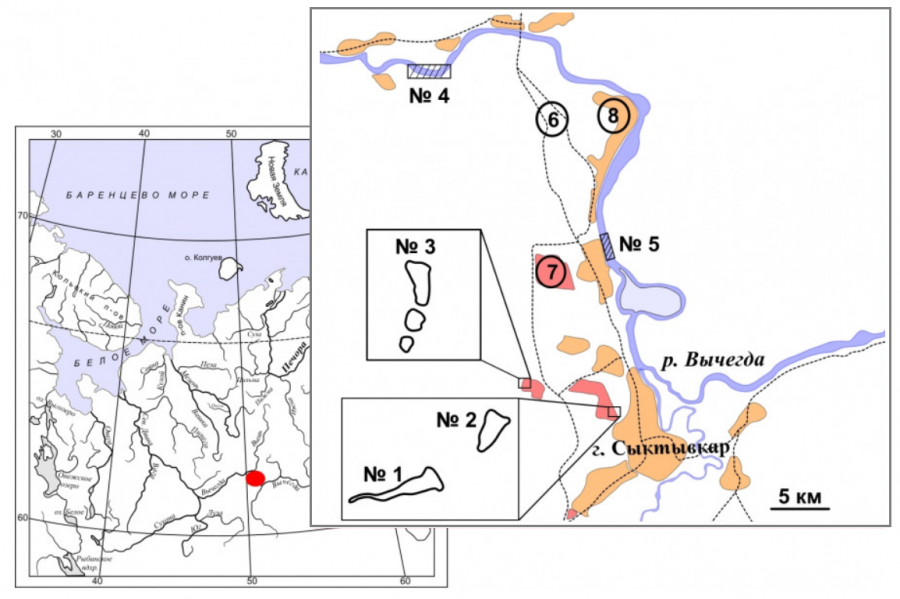
Рис. 1. Карта-схема района исследования в бассейне р. Вычегда. Условные обозначения: № 1 и 2 – места отлова ротана в дачном массиве «Дырнос», № 3 – место поимки ротана в дачном массиве «Находка», № 4–5 – места отлова белоглазки и леща в русле р. Вычегда, 6 – автодороги, 7 – дачи, 8 – населенные пункты
Fig. 1. Scematic map of the research area in the Vychegda River basin. Symbols: 1 and 2 – ponds, places of catching Chinese sleeper in the dacha community "Dyrnos", 3 – pond, place of catching Chinese sleeper in the dacha community “Nakhodka”, 4–5 – places of catching white-eye bream and eastern bream in the Vychegda River, 6 – highways, 7 – dachas, 8 – settlements
Методы
Обработку материала проводили в лабораторных условиях по стандартным методикам (Правдин, 1966), использовав для подготовки данной работы лишь группу стабильных в онтогенезе меристических признаков: ll – число прободенных чешуй в боковой линии (для ротана squma – число поперечных рядов чешуй в боку тела), D или D2, A, V, P – число ветвистых лучей соответственно в спинном (для ротана во втором спинном), анальном, брюшном и грудном плавнике, sp. br. – число тычинок на 1-й жаберной дуге, vert. – общее число позвонков. Для более точного определения количества лучей в плавниках и тычинок на первой жаберной дуге указанные структуры окрашивали ализариновым красным (Якубовский, 1970). Значения парных морфологических признаков представлены по левой стороне тела. При наличии сросшихся позвонков их количество было определено по остаткам сочленовых поверхностей и остистым отросткам, произведен перерасчет их общего количества.
Статистическая обработка данных проведена с использованием программного пакета PAST 3.25. Вследствие отсутствия полового диморфизма по всем исследованным меристическим признакам выборки представлены смешанным по половому составу материалом. Значение признаков номинативных форм представлено из монографии «Атлас пресноводных рыб России» (Атлас…, 2003).
Для оценки стабильности развития чужеродных видов проведено определение частоты асимметричного проявления на признак (Захаров и др., 2000) в билатеральных морфологических признаках (squma или ll, P, V, sp. br.), определен уровень морфологического разнообразия с применением коэффициента вариации, показателя (µ) Животовского и доли (h) редких морф (Животовский, 1980) на основании семи признаков, указанных в табл. 1 и 3.
Результаты
На фоне достаточно интенсивного рыболовства, сосредоточенного в рассматриваемом водотоке, хозяйственно ценных (сиг обыкновенный, хариус европейский) и крупных частиковых (щука, лещ) видов рыб, увеличивается промысловое значение менее ценных представителей ихтиофауны, одним из которых является расселяющаяся белоглазка. В бассейне р. Северная Двина она стала отмечаться в 1960-х гг., а в р. Вычегда – в начале 1970-х гг. (Соловкина, 1975; Новоселов, 2000). На сегодняшний день данный вид освоил русловые биотопы достаточно обширной территории бассейна р. Северная Двина, в т.ч. нижнее и среднее течения (до района с. Усть-Кулом) самого крупного притока – р. Вычегда.
Нам неизвестно, как именно (посредством аквариумистов или рыбаков-любителей) ротан-головешка был акклиматизирован в пруды дачного общества возле г. Сыктывкара, однако необходимо отметить, что по прошествии двух десятилетий спектр водоемов, где он был обнаружен, увеличился. Помимо озер № 1 и 2, где отобран материал, обнаружены особи ротана в других пожарных водоемах (№ 3) на удалении 10 км от первого места обнаружения (см. рис. 1). В иных водных объектах бассейна р. Вычегда данный вид пока не зафиксирован.
Величина и диапазон изменчивости меристических признаков белоглазки и ротана-головешки из бассейна р. Вычегда (см. табл. 1) соответствуют их видовым стандартам по большей части показателей (Атлас..., 2003). Однако необходимо отметить, что белоглазка характеризуется большим количеством лучей в анальном плавнике и тычинок на жаберной дуге. Ротан, в свою очередь, имеет большее число лучей в грудном и спинном плавниках. При анализе опубликованных данных обнаружены разнонаправленные значимые отличия с указанными выборками. Например, ротан из водоемов Республики Коми значимо отличается (Tst = 4 при p < 0.05) меньшим количеством ветвистых лучей во втором спинном и анальном (Tst = 7.5 при p < 0.05) плавниках по сравнению с нативной популяцией из района г. Хабаровска. Однако какая-либо географическая обусловленность (зависимость от широты местности) этих изменений не прослеживается.
Таблица 1. Значения меристических признаков ротана-головешки по нашим и опубликованным данным
| Район поимки, широта | Значение морфологического признака | Источник | ||||||
| squma | D2 | A | V | P | sp. br. | vert. | ||
| Номинативная форма | 36–43 | 9–11 | 7–10 | 5 | 10–14 | 11–14 | - | Атлас..., 2003 |
| Респ. Коми, пруды (61°) | 36–43 | 8–12 | 6–10 | 5 | 14–18 | 10–14 | 29–31 | Наши данные (объединенная выборка, n = 60) |
| 38.5 ± 0.2 | 9.9 ± 0.1 | 8.2 ± 0.1 | 5 | 15.5 ± 0.1 | 11.9 ± 0.1 | 29.8 ± 0.1 | ||
| Архангельская обл., озеро (62°) | - | 8–11 | 7–11 | 29–31 | Касьянов, Горошкова, 2012 | |||
| - | 10.1 0.2 | 9.1 ± 0.2 | 30.0 ± 0.1 | |||||
| г. Хабаровск, старица (48°) | - | 9–12 | 9–12 | - | - | - | 29–31 | |
| - | 10.8 ± 0.2 | 9.9 ± 0.2 | - | - | - | 30.2 ± 0.1 | ||
| г. Саранск, озеро (54°) | - | 10–12 | 8–12 | - | - | - | 29–31 | |
| - | 10.8 ± 0.2 | 9.3 ± 0.2 | - | - | - | 29.9 ± 0.1 | ||
| Подмосковье, карьер (55°) | - | 6–12 | 6–11 | 4–5 | 8–15 | - | - | Щербакова и др., 2017 |
| - | 10 | 8 | 5 | 11 | - | - | ||
| Томская обл., пруд (56°) | - | 10–12 | 8–10 | 5 | 10–14 | 10–13 | - | Сусляев и др., 2016 |
| - | 11.3 ± 0.1 | 9.0 ± 0.2 | 5 | 13.1 ± 0.2 | 10.9 ± 0.1 | |||
| Самарская обл., озеро (53°) | - | 9–12 | 8–11 | 5 | 15–18 | - | - | Шемонаев, Кириленко, 2013 |
| - | 10.9 ± 0.1 | 9.7 ± 0.1 | 5 | 16.7 ± 0.1 | - | - | ||
| Иркутское вод-ще (52°) | 38–44 | 12–14 | 10–13 | 5 | 15–17 | - | - | Богданов, 2023 |
| 41 | 13 | 11.5 | 5 | 16.2 | - | - | ||
| Верхнее течение р. Обь, пойма (53°) | 36–42 | 10–12 | 8–10 | - | 11–14 | 11–12 | 28–30 | Журавлев, 2012 |
| 37.7 ± 0.3 | 11.0 ± 0.1 | 9.1 ± 0.1 | - | 12.3 ± 0.1 | 11.1 ± 0.1 | 28.4 ± 0.1 | ||
| Сербия, пойма р. Дунай (44°) | - | 9–13 | 7–11 | 3–6 | 12–14 | - | - | Nikolić et al., 2021 |
| - | 10.4 ± 0.1 | 9.7 ± 0.1 | 3.3 ± 0.1 | 13.0 ± 0.1 | - | - | ||
Примечание. Над чертой – пределы варьирования, под чертой – среднее значение признака ± его ошибка (при наличии).
Результаты, полученные на основании морфологических описаний одних и тех же признаков (указаны в табл. 1 и 3), свидетельствуют о том, что популяционное разнообразие белоглазки (µ = 3.61 ± 0.11 и h = 0.14 ± 0.02) и ротана (µ = 3.44 ± 0.12 и 3.45 ± 0.09; h = 0.15 ± 0.03 и 0.09 ± 0.02) достоверно не различаются. В то же время показатель частоты асимметричного проявления на признак и его балльная оценка для ротана-головешки указывают на существенные отклонения стабильности индивидуального развития его выборок от нормы (табл. 2).
Таблица 2. Значения показателей морфологического разнообразия и флуктуирующей асимметрии исследованных выборок белоглазки, леща и ротана-головешки
| Вид | Число особей | ЧАП/П | Балл | Показатель Животовского (µ) | Доля редких морф (h) |
| Белоглазка | 31 | 0.26 ± 0.03 | 1 | 3.61 ± 0.11 | 0.14 ± 0.02 |
| Лещ | 34 | 0.30 ± 0.04 | 2 | 3.93 ± 0.11 | 0.13 ± 0.02 |
| Ротан озеро № 1 | 30 | 0.36 ± 0.04 | 3 | 3.45 ± 0.09 | 0.09 ± 0.02 |
| Ротан озеро № 2 | 30 | 0.40 ± 0.04 | 4 | 3.44 ± 0.12 | 0.15 ± 0.03 |
Примечание. ЧАП/П – частота асимметричного проявления на признак, балл – балльная оценка показателя асимметрии.
Одним из факторов успешной натурализации вселенцев является их филогенетическое сходство с аборигенными видами рыб (Xu et al., 2024). Так, например, для белоглазки, активно расселяющейся по магистральному руслу р. Вычегда, можно указать близкородственный вид – лещ Abramis brama (Linnaeus, 1758). Это довольно широко распространенный и массовый вид в бассейне изучаемой реки. Морфологическое разнообразие его выборки оказалось выше (3.93 ± 0.11, Tst = 2.1 при p < 0.05), чем у белоглазки, при отсутствии значимых различий в уровне асимметрии.
Наибольший вклад в показатель общей морфологической изменчивости леща и белоглазки внесли одни и те же морфологические структуры, среди которых: число прободенных чешуй в боку тела, ветвистых лучей в анальном плавнике и количество жаберных тычинок (см. табл. 3).
Таблица 3. Основные показатели изменчивости меристических признаков белоглазки и леща из бассейна р. Вычегда
| Признак | Белоглазка (n = 31) | Лещ (n = 34) | ||||||||
| min–max | X ± xm | СV | m | µ | min–max | X ± xm | СV | m | µ | |
| ll | 49–54 | 51.8 ± 0.2 | 2.3 | 6 | 5.3 ± 0.4 | 52–60 | 55.4 ± 0.4 | 3.1 | 9 | 8.2 ± 0.5 |
| D | 8 | 8 | 0 | 1 | 1.0 ± 0.0 | 9–10 | 9.1 ± 0.1 | 3.2 | 2 | 1.6 ± 0.1 |
| A | 36–43 | 40.0 ± 0.3 | 4.5 | 8 | 7.3 ± 0.4 | 23–28 | 25.6 ± 0.2 | 5.3 | 6 | 5.7 ± 0.2 |
| V | 7–8 | 8.0 ± 0.1 | 2.3 | 2 | 1.4 ± 0.2 | 8–9 | 8.0 ± 0.1 | 2.2 | 2 | 1.3 ± 0.2 |
| P | 16–18 | 16.7 ± 0.1 | 3.4 | 3 | 2.6 ± 0.2 | 16–17 | 16.5 ± 0.1 | 3.1 | 2 | 2.0 ± 0.0 |
| sp. br. | 20–25 | 22.2 ± 0.2 | 5.9 | 6 | 5.4 ± 0.3 | 21–27 | 24.0 ± 0.3 | 6.2 | 7 | 6.1 ± 0.4 |
| vert. | 45–47 | 46.4 ± 0.1 | 1.2 | 3 | 2.5 ± 0.2 | 43–45 | 44.1 ± 0.1 | 1.3 | 3 | 2.7 ± 0.2 |
| Уровень морфологического разнообразия, рассчитанный по выборке (µ) | 3.6 ± 0.1 | 3.9 ± 0.1 | ||||||||
Примечание. min–max – пределы варьирования, x ± xm – среднее значение признака ± его ошибка, СV – коэффициент вариации, m – количество вариантов (морф), µ – показатель Животовского.
Обсуждение
Рассматриваемые виды рыб очень отличаются по своим предпочтениям к главным экологическим характеристикам, что подтверждается принадлежностью к разным фаунистическим комплексам рыб (Никольский, 1980). Так, например, белоглазка относится к понтокаспийскому ихтиокомплексу, а ротан-головешка – к китайскому равнинному. Однако, несмотря на то, что основная черта представителей понтокаспиского ихтиокомплекса – это теплолюбивость, близкое филогенетическое сходство белоглазки с широко распространенным в данном бассейне лещом может свидетельствовать о ее предрасположенности к расселению в условиях Севера (Xu et al., 2024). На сегодняшний день белоглазка расселилась по руслу Северной Двины и вплоть до верхнего течения одного из крупных притоков – р. Вычегда.
Представители китайского равнинного комплекса имеют высокий инвазивный потенциал, что позволило им расселиться по всей России, сопредельным странам и не только (Решетникав, 2009; Интересова, 2016; Кудерский, 2015; Лукина, 2011). В частности, ротан характеризуется такими особенностями, как короткий жизненный цикл, широкий спектр питания и значительный уровень толерантности к различным экологическим факторам (Касьянов, Горошкова, 2012). Данные виды можно рассматривать как пример разных механизмов вселения – саморасселения и целенаправленного или случайного вселения малой группой.
Проведенное исследование показало, что размах изменчивости выборок белоглазки и ротана-головешки на приобретенной части ареала по некоторым морфологическим признакам выходит за пределы ранее описанной номинативной формы изучаемых видов (Атлас…, 2003).
Известно, что число элементов в некоторых скелетных структурах рыб может не только отражать наследственную норму реакции, сформировавшуюся в процессе адаптации вида к условиям конкретной локальной ландшафтно-географической зоны, но также быть связано с температурой окружающей среды, при которой проходит ранний онтогенез (Кожара и др., 1996; Ершов, 2003). Нативные популяции ротана-головешки практически не обитали севернее 54° с.ш., а белоглазки севернее 59–60° с.ш. Известно, что значительный уровень изменчивости может наблюдаться в группировках, обитающих в неоптимальных условиях, например на границе занимаемого ареала (Яковлев, Изюмов, 1982). Более высокие показатели изменчивости отмечались для инвазивных популяций ротана и другими авторами (Журавлев, 2012; Касьянов, Горошкова, 2012; Шемонаев, Кириленко, 2013). Количественные изменения в морфотипе исследованных группировок свидетельствуют об увеличении изменчивости особей и возможных адаптивных перестройках к условиям приобретенной части ареала.
Показано отсутствие значимых различий в показателях популяционного разнообразия (индекс Животовского и доля редких морф) между выборками белоглазки и ротана-головешки. В то же время при сравнении белоглазки и близкородственного вида – леща обнаружены значимые различия. С одной стороны, это может объясняться индивидуальным характером изменчивости разных видов рыб, с другой – отражать общую тенденцию снижения популяционного разнообразия при вселении на новую территорию.
Популяция, формирующаяся из небольшого числа особей в водоеме малой площади, испытывает эффекты «основателя» и «горлышка бутылки», приводящие к обеднению генофонда. Некоторые авторы связывали с пониженной величиной гетерозиготности популяции (которая также наблюдается и при инвазивном процессе) повышенный уровень флуктуирующей асимметрии, т.е. ухудшение процессов стабильности развития особей (Handford, 1980; Møller, 1997). В таких условиях ранний онтогенез наиболее подвержен воздействию разных экологических факторов, что затрудняет поддержание гомеостаза развития особей. Данное предположение может объяснять высокие значения асимметрии в выборках ротана-головешки (4 балла по 5-балльной шкале – значительный уровень отклонений от нормы). Ранее были описаны различные фенодивиации (аномалии позвоночного столба и лучей в плавниках) у исследованных выборок ротана. Так, например, у особей из пруда № 2 до 57 % (17 особей из 30 экз.) от выборки имели компрессии позвонков, включающее срастание позвонков в количестве от 1 до 3 случаев на 1 особь. Количество аномалий позвоночника в выборке из пруда № 1 составила до 20 %, или 6 особей из 30 экз. (Рафиков, 2018). Для чужеродных популяций ротана уже описаны случаи большого числа аномалий (до 40 % выборки) в строении плавников (Шемонаев, Кириленко, 2013). Показан высокий уровень флуктуирующей асимметрии (0.35–0.42, что соответствует 3–4 баллам), рассчитанной по единой методике с применением тех же признаков для популяций ротана из некоторых водоемов Тюменской области (Бойченко, 2020).
В качестве альтернативной причины качественных и количественных изменений фенотипических характеристик указанных видов можно предположить неравнозначный характер отбора в популяциях видов-вселенцев, находящихся в разных условиях. При расселении в естественном водном объекте чужеродный вид встраивается в систему отношений в сообществе, как и аборигенные виды, в частности отношения по типу «хищник – жертва». В изолированных водных объектах может не быть естественных хищников, которые бы нивелировали особей с дефектами развития.
После накопления морфологических описаний других чужеродных видов рыб, обитающих на территории Республики Коми, высказанная гипотеза будет проверена на более широком перечне видов из разных местообитаний. В дополнение к изучению фенотипических признаков актуально применение дополнительных молекулярно-генетических маркеров для изучения изменений генетического разнообразия расселяющихся видов.
Заключение
Проведенная работа показала, что в бассейне р. Вычегда расширяется ареал чужеродных видов рыб. С момента начала инвазии белоглазка распространилась по русловым местообитаниям нижнего и среднего течений рассматриваемой реки. Количество водных объектов, где был обнаружен ротан-головешка, также увеличилось до трех, однако все они являются пожарными водоемами дачных обществ. В придаточных (пойменные озера, курьи) водоемах бассейна р. Вычегда данный вид пока не отмечался.
В результате исследования было установлено, что размах изменчивости по некоторым морфологическим признакам выходит за пределы ранее описанной номинативной формы изучаемых видов, что можно расценивать как происходящую адаптацию исследованных чужеродных видов к обитанию в условиях приобретенной части ареала.
Показано, что популяция, формирующаяся из нескольких особей в водоеме малой площади, характеризуется низкой стабильностью индивидуального развития ее особей и наличием большого числа фенодивиаций. В качестве основной гипотезы необходимо указать, что подобные популяции могут подвергаться большему воздействию экологических факторов в период раннего онтогенеза по сравнению с самостоятельно расселяющимися группировками других видов в русле крупных рек.
Библиография
Атлас пресноводных рыб России: В 2 т. Т. 1 / Под ред. Ю. С. Решетникова. М.: Наука, 2003. 379 с.
Биологические инвазии в водных и наземных экосистемах / А. Ф. Алимов, Н. Г. Богуцкая, М. И. Орлова . М.: Товарищество научных изданий КМК, 2004. 436 с.
Богданов Б. Э. Ротан-головешка Perccottus glenii в системе Иркутского водохранилища: распространение, морфология, экология // Байкальский зоологический журнал. 2023. № 2 (34). С. 21–25.
Бознак Э. И. Головешка-ротан Percсottus glenii (Eleotridae) из бассейна реки Вычегды // Вопросы ихтиологии. 2004. Т. 44, № 5. С. 712–713.
Бойченко К. А. Флуктуирующая асимметрия ротана Perccottus glenni в водоемах Тюменской области // Фундаментальные основы инновационного развития науки и образования: Сборник статей VII Международной научно-практической конференции, Пенза, 23 апреля 2020 года. Пенза: Наука и Просвещение, 2020. С. 24–26.
Ершов Н. П. О географической изменчивости меристических признаков у бельдюги Zoarces viviparus (L.) и керчака Myoxocephalus scorpius (L.) // Вестник Санкт-Петербургского университета. Серия 3: Биология. 2003. № 4. С. 64–72.
Животовский Л. А. Показатель внутрипопуляционного разнообразия // Журнал общей биологии. 1980. Т. 41, № 6. С. 828–836.
Журавлев В. Б. Морфоэкологическая характеристика ротана пойменных водоемов верхнего течения Оби // Вестник НГАУ (Новосибирский государственный аграрный университет). 2012. № 2-2 (23). С. 28–31.
Захаров А. Б., Бознак Э. И. Современные изменения рыбного населения крупных рек Европейского Северо-Востока России // Российский журнал биологических инвазий. 2011. № 1. С. 23–33.
Захаров В. М., Баранов А. С., Борисов В. И., Валецкий А. В., Кряжева Н. Г., Чистякова Е. К., Чубинишвили А. Т. Здоровье среды: методика оценки . М.: Центр экологической политики России, 2000. 68 с.
Интересова Е. А. Чужеродные виды рыб в бассейне Оби // Российский журнал биологических инвазий. 2016. Т. 9, № 1. С. 83–100.
Касьянов А. Н., Горошкова Т. В. Изучение морфологических признаков у ротана Perccottus glenii (Perciformes, Eleotrididae), интродуцированного в водоемы европейской части России // Сибирский экологический журнал. 2012. № 1. С. 81–86.
Кирпичников В. С. Генетика и селекция рыб . Л.: Наука, 1987. 520 с.
Кожара А. В., Изюмов Ю. Г., Касьянов А. Н. Общая и географическая изменчивость числа позвонков у некоторых пресноводных рыб // Вопросы ихтиологии. 1996. Т. 36, вып. 2. С. 179–194.
Кудерский Л. А. Избранные труды. Исследования по ихтиологии, рыбному хозяйству и смежным дисциплинам. Акклиматизация рыб в водоемах России . (Сборник научных трудов ФГБНУ ГосНИОРХ. Т. 4, вып. 343.) СПб.: Товарищество научных изданий КМК, 2015. 290 с.
Лукина И. И. Распространение ротана (Perccottus Glenii Dybowski, 1877) в Беларуси // Российский журнал биологических инвазий. 2011. № 2. С. 114–119.
Никольский Г. В. Структура вида и закономерности изменчивости рыб . М.: Пищевая промышленность, 1980. 183 с.
Новоселов А. П. Современное состояние рыбной части сообществ в водоемах Европейского Северо-Востока России : Автореф. дис. … д-ра биол. наук. М., 2000. 50 с.
Павлов Д. А. Морфологическая изменчивость в раннем онтогенезе костистых рыб . М.: ГЕОС, 2007. 264 с.
Правдин И. Ф. Руководство по изучению рыб . М.: Пищепромиздат, 1966. 376 с.
Решетников А. Н. Современный ареал ротана Perccottus glenii Dybowski, 1877 (Odontobutidae, Pisces) в Евразии // Российский журнал биологических инвазий. 2009. № 3. С. 22–35.
Решетников А. Н, Голубцов А. С., Журавлев В. Б., Ломакин С. Л., Резвый А. С. Расширение ареалов ротана Perccottus glenii, верховки Leucaspius delineatus и уклейки Alburnus alburnus в бассейне р. Обь // Сибирский экологический журнал. 2017. № 6. С. 696–707.
Рафиков Р. Р. Морфологическая характеристика и особенности питания головешки-ротана (Perccottus glenii Dybowski, 1877) в северо-восточной части приобретенного ареала (территория Республики Коми) // Известия Самарского научного центра РАН. 2018. Т. 20, № 5. С. 43–49.
Рафиков Р. Р. Формирование рыбного населения искусственных водоемов на территории Республики Коми : Автореф. дис. … канд. биол. наук. Сыктывкар, 2016. 20 с.
Соловкина Л. Н. Рыбные ресурсы Коми АССР . Сыктывкар: Коми книжное изд-во, 1975. 168 с.
Стерлигова О. П., Ильмаст Н. В. Виды-вселенцы в водных экосистемах Карелии // Вопросы ихтиологии. 2009. Т. 49, № 3. С. 372–370.
Сусляев В. В., Решетникова С. Н., Интересова Е. А. Биология ротана Perccottus glenii Dybowski, 1877 в водоемах южно-таежной зоны Западной Сибири // Вестник НГАУ (Новосибирский государственный аграрный университет). 2016. № 1 (38). С. 78–85.
Татарко К. И. Влияние температуры на меристические признаки рыб // Вопросы ихтиологии. 1968. Т. 8, № 3. С. 425–439.
Шемонаев Е. В., Кириленко Е. В. Аномалии плавников ротана-головешки Perccottus glenii Dybowski, 1877 из озер Мордовинской поймы // Экологический сборник 4: Труды молодых ученых Поволжья. Всероссийская научная конференция с международным участием / Под ред. проф. С. В. Саксонова. Тольятти: ИЭВБ РАН: Кассандра, 2013. XXII + 211 с.
Щербакова В. Д., Сайнчук А. Д., Самойлов К. Ю., Бурменский В. А., Павлов С. Д., Пивоваров Е. А., Сенчукова А. Л. Ротан-головешка (Perccottus glenii Dybowski, 1877) из озера-карьера Сима (Одинцовский район, Московская область): первые данные о новой популяции вида // Бюллетень Московского общества испытателей природы. Отдел биологический. 2017. Т. 122, № 6. С. 14–24.
Яковлев В. Н., Изюмов Ю. Г. Морфологическая изменчивость и внутривидовая структура волжского леща // Экология водных организмов верхневолжских водохранилищ. Л.: Наука, 1982. С. 171–193.
Якубовский М. Методы выявления и окраски системы каналов в боковой линии и костных образований у рыб in toto // Зоологический журнал. 1970. Т. 49, № 9. С. 1398–1402.
Handford P. Heterozygosity at enzyme loci and morphological variation // Nature. 1980. Vol. 286. P. 261–262. DOI: 10.1038/286261a0
Lodge D. M. Biological invasions: lessons for ecology // Trends in Ecology and Evolution. 1993. Vol. 8. P. 133–137. DOI: 10.1016/0169-5347(93)90025-K
Møller A. P. Developmental stability and fitness: A review // The American Naturalist. 1997. Vol. 149, № 5. P. 916–932.
Nikolić D., Skorić S., Cvijanović G., Jaćimović M., Đikanović V., Mićković B. Morphometric and meristic characteristics of the Amur sleeper (Perccottus glenii) from the Danube River drainage channel // Archives of Biological Sciences. 2021. Vol. 73. P. 381–388.
Xu M., Li Sp., Liu C., Todesco P., Dick J., Fang M., Wei H., Yu F., Shu L., Wang X., Gu D., Mu X. Global freshwater fish invasion linked to the presence of closely related species // Nature Communications. 2024. Vol. 15:1411. P. 1–11. DOI: 10.1038/s41467-024-45736-8
Благодарности
Работа выполнена в рамках государственного задания «Разнообразие фауны и пространственно-экологическая структура животного населения европейского северо-востока России и сопредельных территорий в условиях изменения окружающей среды и хозяйственного освоения» № гос. регистрации 122040600025-2 (2022-2024 гг.).

 © 2011 - 2026
© 2011 - 2026