



Введение
Проблема разделения симпатрическими видами ресурсов окружающей среды – одна из значимых в экологии. Смягчению или устранению конкуренции способствуют видовые различия в использовании пространства, во времени его использования и в пищевой специфике (Ананьева, 1981). Первый из названных аспектов – пространственное, или биотопическое, разделение – имеет, как правило, превалирующее значение (Schoener, 1989). В условиях нарастающих изменений климата и масштабных антропогенных воздействий, влекущих стремительную трансформацию ландшафтов, именно биотопические предпочтения нередко определяют перспективы выживания видов в меняющейся природной среде.
На рубеже ХХ и XXI вв. динамичные сукцессионные процессы в направлении остепнения затронули Терский песчаный массив в Восточном Предкавказье (Биткаева, Николаев, 2001). Одним из результатов зарастания песков стало сокращение в регионе области обитания большинства представителей туранского герпетофаунистического комплекса (Лотиев, Батхиев, 2019).
Наблюдения последних лет подтверждают обоснованность внесения всех достоверно существующих в Чеченской Республике рептилий туранского генезиса – степной агамы, Trapelus sanguinolentus (Pallas, 1814); ушастой круглоголовки, Phrynocephalus mystaceus (Pallas, 1776); круглоголовки-вертихвостки, Phrynocephalus guttatus (Gmelin, 1789); быстрой ящурки, Eremias velox (Pallas, 1771); разноцветной ящурки, Eremias arguta (Pallas, 1773); песчаного удавчика, Eryx miliaris (Pallas, 1773) – в региональную Красную книгу (Красная книга Чеченской Республики, 2020). Подвидовая принадлежность региональных популяций вышеперечисленных видов, часто дискуссионная, не является предметом данной статьи.
Целью работы является описание особенностей биотопического размещения трех фоновых видов ящериц Терских песков в период относительно стабильной экологической обстановки и оценка их современного охранного статуса.
Материалы
Терский песчаный массив (= Терские пески) находится в административных границах Чеченской Республики (преобладающая часть), Республики Дагестан и Ставропольского края, простирается с запада на восток приблизительно на 180 км, с севера на юг – на 70–80 км, между точками с координатами 43°20'–44°10' с. ш. и 45°–46°20' в. д. (рис. 1). Его площадь более 6 тыс. км2 (Биткаева, Николаев, 2001).

Рис. 1. Терские пески на карте-схеме северо-западного Прикаспия (по: Биткаева, Николаев, 2001, с изменениями); район исследований отмечен красной точкой
Fig. 1. Terek sands on the schematic map of the northwestern Caspian region (according to Bitkaeva, Nikolaev, 2001, with modifications); the research area is marked with a red dot
Изучение биотопических предпочтений трех симпатрических видов ящериц (Phr. guttatus, E. arguta, E. velox) проводилось в весенне-летний период 1983–1984 гг. на юго-восточной периферии Терских песков, близ станицы Старогладковской (Чеченская Республика).
Стационарный замкнутый маршрут (объект «Артезиан») протяженностью 1 км был заложен в 5 км к западу от оз. Будары (ныне гидрологический памятник природы «Степная жемчужина»). Он проходил по участкам с песками различной степени закрепленности и зарослями жостера Палласа (Rhamnus pallasii Fich. et C. A. Mey) (рис. 2). Координаты локалитета: 43°37.0′ с. ш.; 46°18.5′ в. д. Длины отрезков, лежащих в пределах одной стации (термины «биотоп» и «стация» трактуются авторами как синонимы), характеризуют следующие цифры: незакрепленные пески (проективное покрытие меньше 15 %) – 242 м; слабо и умеренно закрепленные пески (проективное покрытие 15–50 %) – 348 м; заросшие пески (проективное покрытие больше 50 %) – 340 м; заросли кустарника – 70 м (проективное покрытие от 15 до 90 %).
В последующие годы (1985–1993; 2007–2023) наблюдения в этом районе были продолжены. В общей сложности на маршруте «Артезиан» было выполнено 84 результативных учета (32 – в 1983–1984 гг.), зафиксировано 996 встреч рептилий 9 видов.
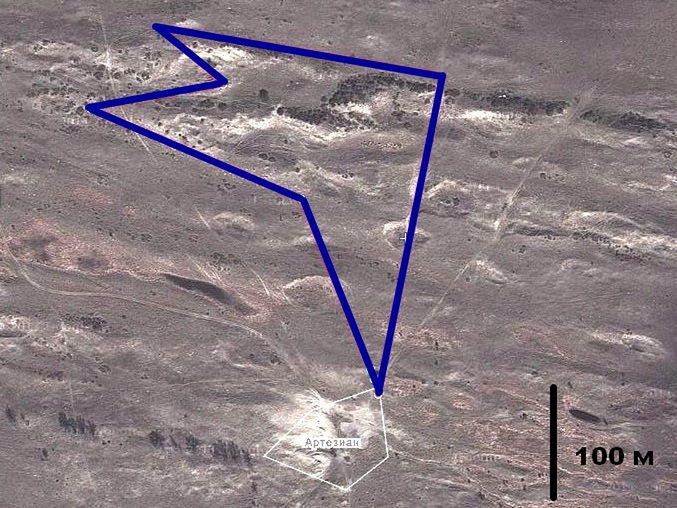
Рис. 2. Карта-схема стационарного учетного маршрута «Артезиан» в 5 км к западу от озера Будары (на основе https://wikimapia.org/)
Fig. 2. Schematic map of the stationary accounting route “Artesian” 5 km west of Lake Budary (based on https://wikimapia.org/)
Методы
Изучение активности рептилий на маршруте «Артезиан» проводилось трансектным методом (Динесман, Калецкая, 1952): ширина учетной полосы 2 м, длина 1000 м. Фиксировалось обнаружение особей рептилий в пределах каждого из четырех выделенных биотопов. Затем по результатам весенних (апрель – 4 учета) и летних (июнь – июль – 28 учетов) наблюдений определяли среднее арифметическое число встреч ящериц по каждой стации в отдельности.
Для получения сопоставимых результатов все полученные данные были пересчитаны на 1 км условного маршрута – в пределах одного биотопа – по формуле:
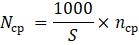 ,
,
где Nср – среднее число особей вида на 1 км маршрута, проходящего в пределах одного биотопа; S – фактическая длина маршрутного отрезка, лежащего в пределах одного биотопа (в метрах); nср – среднее число особей вида, отмеченных на отрезке S за один учет.
Единовременность проведения учета во всех стациях одного локалитета смягчала проблему сопоставимости данных разных учетов, каждый из которых фиксировал биотопическое распределение животных в короткий временной отрезок (время прохождения маршрута составляло не более 1 часа). Погодные условия, суточные перемещения ящериц, поло-возрастной состав их популяций в данном случае в расчет не принимались.
Доля (соотношение) встречаемости (в %) вида в биотопе определялась по формуле:
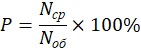 ,
,
где P – доля встречаемости вида в биотопе; Nср – среднее число особей вида на 1 км маршрута, проходящего в пределах одного биотопа; Nоб – среднее общее число особей всех трех видов ящериц на 1 км маршрута, проходящего в пределах одного биотопа.
Результаты
Полученные учетные данные позволили определить средние показатели встречаемости круглоголовки-вертихвостки, разноцветной и быстрой ящурок на 1 км маршрута (табл. 1, рис. 3–4), а также долю (соотношение) их встреч (в %) в пределах каждого из четырех выделенных биотопов (табл. 2).
Таблица 1. Встречаемость трех симпатрических видов ящериц по стациям (среднее число особей на 1 км маршрута в пределах биотопа): объект «Артезиан», 1983–1984 гг.
| Время учета | Биотоп | Phrynocephalus guttatus | Eremias arguta | Eremias velox |
| Апрель (4 учета) | незакрепленные пески | 11.8 | 0 | 1.8 |
| слабо закрепленные пески | 17 | 8.3 | 5.8 | |
| закрепленные пески | 0 | 3.8 | 0 | |
| заросли кустарника | 0 | 0 | 0 | |
| Июнь – июль (28 учетов) | незакрепленные пески | 12.8 | 0.1 | 1.9 |
| слабо закрепленные пески | 3.5 | 2.8 | 3.5 | |
| закрепленные пески | 0 | 3.6 | 0.9 | |
| заросли кустарника | 0.7 | 1.1 | 7.8 |
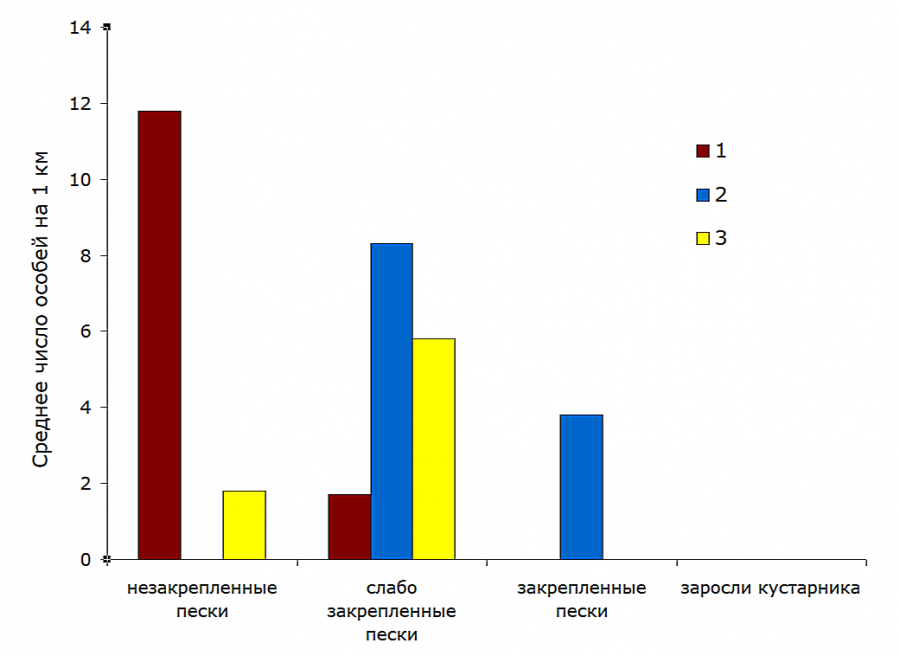
Рис. 3. Распределение трех симпатрических видов ящериц по биотопам (объект «Артезиан», апрель 1983–1984 гг.): 1 – Phr. guttatus; 2 – E. arguta; 3 – E. velox
Fig. 3. Distribution of three sympatric species of lizards by biotopes (Artesian object, April 1983–1984): 1 – Phr. guttatus; 2 – E. arguta; 3 – E. velox
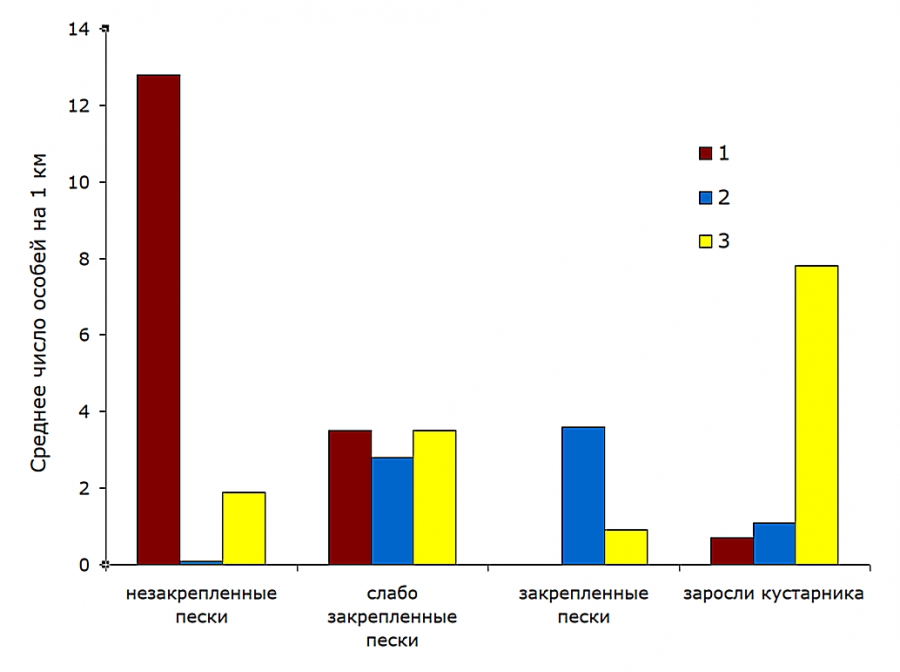
Рис. 4. Распределение трех симпатрических видов ящериц по биотопам (объект «Артезиан», июнь – июль 1983–1984 гг.): 1 – Phr. guttatus; 2 – E. arguta; 3 – E. velox
Fig. 4. Distribution of three sympatric species of lizards by biotopes (object "Artesian", June – July 1983–1984): 1 – Phr. guttatus; 2 – E. arguta; 3 – E. velox
Таблица 2. Соотношения (доли) встречаемости трех симпатрических видов ящериц в выделенных биотопах (округлено до целых %): объект «Артезиан», 1983–1984 гг.
| Время учета | Биотоп | Phrynocephalus guttatus | Eremias arguta | Eremias velox |
| Апрель (4 учета) | незакрепленные пески | 87 | 0 | 13 |
| слабо закрепленные пески | 10 | 53 | 37 | |
| закрепленные пески | 0 | 100 | 0 | |
| заросли кустарника | 0 | 0 | 0 | |
| Июнь – июль (28 учетов) | незакрепленные пески | 86 | 1 | 13 |
| слабо закрепленные пески | 36 | 28 | 36 | |
| закрепленные пески | 0 | 80 | 20 | |
| заросли кустарника | 7 | 12 | 81 |
Приведенные данные наглядно показывают пространственно-биотопическое разделение симпатрических видов.
Незакрепленные пески – предпочитаемая стация круглоголовки-вертихвостки. Средняя встречаемость этих ящериц была здесь в 3.7 (июнь – июль) – 7 (апрель) раз выше, чем на смежных участках слабо закрепленных песков. Они же абсолютно преобладали в биотопе. Доля их встреч составляла 86 (июнь – июль) – 87 % (апрель). Значительно представительство быстрой ящурки – 13 %, а единичные встречи разноцветной ящурки носили случайный характер.
Слабо закрепленные пески населяли все три вида ящериц. В апреле это предпочитаемый биотоп для быстрой и разноцветной ящурок при численном доминировании (53 % встреч) E. arguta. Плотность первой здесь в 3.2 раза выше, чем на незакрепленных песках, второй – в 2.2 раза выше, чем на закрепленных песках. Летом картина существенно меняется: зоны активности всех трех видов ящериц смещаются, частично или преимущественно, в более закрытые, затененные биотопы. Круглоголовки-вертихвостки – в слабо закрепленные пески (доля их участия здесь достигает 36 %), быстрой ящурки – в заросли кустарника (где они абсолютно доминируют – 81 % встреч), разноцветной ящурки – в закрепленные пески (абсолютный доминант – 80 % встреч). Такие перемещения требуют от ящериц миграций лишь на несколько десятков метров. Заметим, что круглоголовки-вертихвостки во все сезоны придерживались участков выдела «слабо закрепленные пески» с наименьшим проективным покрытием, а быстрые ящурки встречались лишь среди разреженных кустарников по кромке зарослей.
Закрепленные пески населены исключительно (весной) или в основном (летом – 80 % встреч) разноцветной ящуркой. Они же являются предпочитаемой стацией для вида в летнее время: плотность E. arguta превышает здесь таковую на слабо закрепленных песках в 1.3 раза. Присутствие быстрой ящурки отмечено только в июне – июле (20 % встреч).
Кустарниковые заросли весной не привлекают ящериц, но летом плотно заселены, в первую очередь быстрой ящуркой, где вид встречался в 2.2 раза чаще, чем на участках разреженной растительности, и абсолютно доминировал (81 % встреч). На долю разноцветной ящурки приходилось 12 % встреч. Круглоголовка-вертихвостка отмечена только по границам выдела и незакрепленных песков – 7 % встреч. Помимо вышеназванных видов, в 1983–1984 гг. исключительно в кустарниковых зарослях встречали полосатую ящерицу, Lacerta strigata Eichwald, 1831, в силу своей относительной редкости в тот период (в среднем 0.5 особи на 1 результативный учет) не включенную в анализ.
Таким образом, каждый из трех видов ящериц в районе наших наблюдений имел свои, причем различные, предпочитаемые стации (см. рис. 4): Phr. guttatus – незакрепленные пески, E. arguta – закрепленные пески, E. velox – кустарниковые заросли. Только в апреле, видимо, непродолжительное время после выхода с мест зимовок оба вида ящурок имели максимальную численность в пределах одного биотопа (см. рис. 3). Слабо закрепленные пески, поросшие разреженной ксерофитной травянистой и кустарниковой растительностью, используются ящурками и круглоголовками в сопоставимой степени.
Как правило, в том биотопе, где частота встреч вида достигает максимума, он является доминирующим. Исключение составляет быстрая ящурка: в апреле она отдает предпочтение слабо закрепленным пескам, где доминирует в этот короткий период ящурка разноцветная (см. рис. 3). Летом на долю вида-доминанта в соответствующей стации приходится от 80 до 86 % общего числа учтенных ящериц.
Обсуждение
В последующие годы (1985–1993, 2007–2023) наблюдения и учеты в районе объекта «Артезиан» были продолжены. Их основные результаты опубликованы ранее (Лотиев, Батхиев, 2018). С 1989 г. на маршруте начала встречаться T. sanguinolentus, которая стала здесь обычным немногочисленным видом. В 2010 г. ее численность достигла максимума (до 5 ос./км), в последующие годы вид исчез. Были отмечены единичные встречи желтопузика, Pseudopus apodus (Pallas, 1775). Значительно увеличилась численность полосатой ящерицы. Доминирующим видом стала разноцветная ящурка, в то время как круглоголовка-вертихвостка исчезла около 2000 г., а быстрая ящурка после 2008 г.
Таким образом, в настоящее время из трех фоновых видов ящериц в 1983–1984 гг. два – круглоголовка-вертихвостка и быстрая ящурка – полностью исчезли, а разноцветная ящурка, напротив, пока сохраняет численность и расширила область обитания.
Столь стремительные и глубокие изменения в составе заурофауны объясняются быстрым остепнением территории в районе исследований: к началу XXI в. пятна голых песков, связанные преимущественно с котловинами выдувания, в разной степени закрепились. Из-за этого исчезли круглоголовки-вертихвостки, их прежние местообитания заняли степные агамы. Дальнейшее зарастание слабо закрепленных песков, увеличение их проективного покрытия, особенно за счет повсеместного распространения мшаников, привело к резкому сокращению территории распространения и численности таких полупустынных видов, как степная агама и быстрая ящурка. Подходящие биотопические условия сохранились лишь для разноцветной ящурки; для полосатой ящерицы природная среда стала более благоприятной.
Заключение
Фоновыми симпатрическими видами заурофауны юго-восточной перефирии Терских песков в начале 1980-х гг. были круглоголовка-вертихвостка, быстрая и разноцветная ящурки, весьма различные в своих экологических предпочтениях и использовании пространства. Наиболее стенобионтные из них Phr. guttatus были жестко приурочены к пятнам незакрепленных песков, E. velox тяготели к слабо закрепленным пескам с кустарниковой растительностью, а E. arguta избегали открытых песков и предпочитали пески закрепленные.
Последующие сукцессионные процессы, господствующий в последние десятилетия тренд на остепнение территории Терских песков привели к полному исчезновению в районе исследований двух из трех наиболее многочисленных видов – круглоголовки-вертихвостки и быстрой ящурки. Лишь разноцветная ящурка сохраняет пока свою численность и расширила область обитания.
При сохранении существующих тенденций можно ожидать скорого и полного исчезновения в Терских песках таких облигатных псаммофилов, как ушастая круглоголовка и круглоголовка-вертихвостка. За ними могут последовать быстрая ящурка и степная агама. Разноцветной ящурки вымирание пока не угрожает, но дальнейшее зарастание песков ухудшит и ее условия существования. Складывающиеся ландшафтно-биотопические условия однозначно благоприятны лишь для полосатой ящерицы, уже расселившейся по всему Терскому песчаному массиву.
Описанная ситуация с состоянием заурофауны в Терских песках коррелирует с общим положением пустынно-полупустынной биоты туранского генезиса в Затеречье. Существует реальная угроза утраты целого ряда видов не только во флоре и фауне Чеченской Республики, но и России в целом. Так, степная агама известна в нашей стране исключительно в Терских песках, преобладающая часть которых лежит в границах Чеченской Республики.
Библиография
Ананьева Н. Б. К изучению симпатрических видов (на примере рептилий) // Проблемы новейшей истории эволюционного учения. Л.: Наука, 1981. С. 15–26.
Биткаева Л. М., Николаев В. А. Ландшафты и антропогенное опустынивание Терских песков . М.: МГУ, 2001. 172 с.
Динесман Л. Г., Калецкая М. Л. Методы количественного учета амфибий и рептилий // Методы учета численности и географического распределения наземных позвоночных. М.: Изд-во АН СССР, 1952. С. 329–340.
Красная книга Чеченской Республики . Ростов-н/Д: ООО «Южный издательский дом», 2020. 480 с.
Лотиев К. Ю., Батхиев А. М. О деградации Туранского герпетофаунистического комплекса в Терском песчаном массиве (Восточное Предкавказье) // Известия высших учебных заведений. Поволжский регион. Естественные науки. 2019. № 2 (26). С. 115–128.
Schoener T. W. The ecological niche // Ecological Concepts / J. M. Cherret (Ed.). Oxford, 1989. P. 79–113.
Благодарности
Важную консультативную и/или организационную помощь нашей работе оказали Н. Б. Ананьева, Б. С. Туниев, Т. Ю. Точиев. Отправной точкой исследований на протяжении более 40 лет была база при гидрологическом памятнике природы Чеченской Республики «Степная жемчужина», существующая заботой охотоведа Е. Е. Сумачева. Успеху полевых наблюдений всемерно способствовали С. В. Буров, Р. Х. Гайрабеков, И. И. Гизатулин, С. Р. Наурханов, Р. С. Эржапова. Всем указанным лицам, а также уважаемым рецензентам авторы выражают искреннюю и глубокую признательность.

 © 2011 - 2026
© 2011 - 2026