



Введение
Городские зеленые насаждения и зеленые территории вне границ города улучшают экологическую обстановку населенного пункта и выполняют разнообразные функции, в т. ч. рекреационную. Однако в результате рекреационной нагрузки на территории происходят изменения во всех природных комплексах. Самые распространенные виды рекреационного воздействия – механическое (вытаптывание, обламывание ветвей, ожог почвы от костров и др.); вынос посетителями ягод, грибов, цветов и др. Именно вытаптывание растительного покрова и уплотнение почвы является постоянным фактором воздействия. Это приводит к обеднению видового состава, снижению проективного покрытия, уменьшению фитомассы растений, изменению структуры напочвенного покрова и подстилки. На рекреационной территории происходит рудерализация фитоценозов, что может привести в течение очень короткого по экологическим меркам времени (20–30 лет) к формированию сетевой структуры леса – чередованию нарушенных и ненарушенных участков. При этом не только нарушаются, но могут и прекратиться процессы естественного возобновления древесной и травянистой растительности. Значительные изменения происходят в зооценозах: меняется соотношение видов, снижается их численность, происходит трансформация микробоценозов (Скрипальщикова, 2008).
Безусловно, рекреационные нагрузки зависят от видов отдыха (массовый повседневный отдых или экскурсии, туризм), т. е. определяются не только количеством отдыхающих, но и концентрацией, и временем пребывания отдыхающих на единице площади. Изменение видового состава растений на территории и площадь вытоптанной поверхности являются одним из важных признаков рекреационной нарушенности территории (Грешилова, Скрипальщикова, 2009).
Целью нашей работы является оценка рекреационной нарушенности территории по результатам геоботанических исследований и доле площадей, занятых тропиночными сетями.
Материалы
Исследования выполнялись на территории экопарка «Гремячая грива», который расположен на территории городских округов г. Красноярск, г. Дивногорск и Емельяновского муниципального района Красноярского края. «Гремячая грива» – цепь сопок, протянувшихся от города Красноярска до Собакиной речки. На территории парка имеется развитая рекреационная инфраструктура: тропы для семейных прогулок, велосипедные дорожки, спортивные комплексы, игровые площадки, пути для любителей интенсивных пеших путешествий по пересеченной местности, экологические тропы, места стоянок, видовые площадки на вершинах и склонах сопок (Экопарк…, 2018; МКЭО Гремячая грива, 2022).
Основная часть исследований проводилась летом 2021 г. В изучаемых лесных сообществах были заложены пробные площади размером 20 × 20 м и сделаны геоботанические описания по традиционным методикам (Методы изучения…, 2002). Размещение пробных площадей на территории парка «Гремячая грива» представлено на рис. 1. Координаты пробных площадей даны в табл. 1.

Рис. 1. Размещение пробных площадей на территории парка «Гремячая грива»
Fig. 1. Placement of sample plots on the territory of the Gremyachaya Griva park
Помимо этого, были проведены съемки с использованием беспилотных летательных аппаратов (БПЛА) в сосняках северной части экопарка (географические координаты 54° с. ш., 091° в. д.); в сосновых, березово-сосновых и березовых лесах в районе «Визит-центр-2» (пробные площади № 7, 23); в сосняках на территории пробных площадей № 18, 20 и 21 («Третья сопка экопарка»).
Методы
Описание живого напочвенного покрова проводилось на учетных площадках размером 100 м2 на каждой пробной площади, для оценки обилия видов была использована комбинированная шкала обилия-покрытия Ж. Браун-Бланке: r – единично; + – менее 1 %; 1 – 1–5 %; 2 – 6–25 %; 3 – 26–50 %; 4 – 51–75 %; 5 – 76–100 %.
Номенклатура сосудистых растений и мхов дана согласно (The Plant List…, 2013). Для выявления принадлежности видов к основным экологическим и эколого-ценотическим группам использовалась классификация С. В. Дегтевой и А. Б. Новаковского и база данных «Ценофонд лесов Европейской России» (Дегтева, 2011; Ценофонд лесов…, 2012) с учетом классификации Т. Н. Буториной (1963).
Интегральная оценка различий видового разнообразия исследуемых сообществ пробных площадей проводилась с помощью ранговых распределений или «кривых значимости видов». Для построения этих распределений виды ранжируются в порядке убывания обилия. Для описания ранговых распределений видов в сообществах используется уравнение Ципфа – Парето (McGill et al., 2007; Matthews, Whittaker, 2015). Ранг вида – его порядковый номер i в ряду распределения обилия s(i) в порядке убывания обилия. В двойных логарифмических координатах график распределения «ранг – обилие» вида – прямая линия.
На территории экопарка «Гремячая грива» производилась съемка с БПЛА dji p4 multispectral ряда для количественной оценки доли тропиночной сети. Изображения были получены с высоты 300 м, разрешение 4000 х 3000 точек. В одном случае (пробные площади №18, 20 и 21) использовался БПЛА с другим разрешением камеры (1600 х 1300 точек). Вследствие этого удобнее в дальнейшем говорить не об абсолютных площадях территорий, а о доле покрытия тропинок от общей площади (в пикселях) лесного покрытия. Далее в графическом редакторе были вырезаны с изображений технические сооружения, водоемы – все объекты, которые не участвовали в дальнейшем расчете. Были прочерчены маршруты тропинок с учетом их ширины. Для дальнейшего расчета площадей тропинки и лесное покрытие были контрастированы.
Результаты
По традиционной эколого-фитоценотической классификации исследуемые сообщества можно отнести к ассоциациям разнотравной и разнотравно-осочковой (березняки и сосняки разнотравные, осочково-разнотравные) и орляковой серий типов леса. Пробные площади № 1–10, 23 закладывались в наиболее посещаемой части экопарка, пробные площади № 18, 20, 21 («Третья сопка экопарка») – на условно фоновой территории (см. рис. 1, табл. 1).
Сообщества разнотравной серии типов леса – одни из наиболее ценных в рекреационном отношении, поскольку в них создается оптимальный режим освещения, а хорошо развитый травянистый покров весьма декоративен. Однако в результате высокой рекреационной нагрузки привлекательность лесных сообществ снижается и запускается процесс рекреационной дигрессии (Казанская и др., 1977).
Таблица 1. Характеристики пробных площадей
| № пробных площадей | Географические координаты | Тип леса / Основная порода (H – высота, м; D – диаметр, см) | Доминанты подлеска / живого напочвенного покрова | Стадия дигрессии* |
| 1, 3 | 55°00.13´ с. ш. 92°45.80´ в. д
55°59.91´с. ш. 92°45.01´ в. д. |
Березняк разнотравный /
Betula pendula H = 18–22; D = 30–40 |
Padus avium, Rosa acicularis / Carex macroura, Agrimonia pilosa, Taraxacum officinale, Cimicifuga foetida | 3 |
| 2 | 55°59.88´с. ш. 92°45.09´ в. д. | Березняк разнотравный /
B. pendula H = 20–25; D = 30–40 |
Caragana arborescens / C. macroura, A. pilosa | 2 |
| 4, 5 | 56°00.00´с. ш. 92°45.59´ в. д
55°59.76´с. ш. 92°44.59´ в. д. |
Березняк орляковый /
B. pendula H = 20–25; D = 30–40 |
P. avium / Pteridium aquilinum, C. macroura, Anthriscus sylvestris | 2 |
| 6 | 55°59.77´с. ш. 92°44.88´ в. д. | Сосняк разнотравный / Pinus sylvestris
H = 17–20; D = 20–25 |
P. avium / C. macroura, C. foetida, A. sylvestris | 2 |
| 7 | 55°59.74´с. ш. 92°44.18´ в. д. | Сосняк разнотравный / P. sylvestris H = 20–25; D = 35–40 |
P. avium / C. macroura, A. sylvestris, Trifolium repens, A. pilosa | 3 |
| 9 | 55°59.64´ с. ш. 92°44.60´ в. д. | Березняк осочково-разнотравный / B. pendula H = 15–18; D = 35–40 |
Padus avium, Acer negundo / C. macroura, Cirsium setosum | 2 |
| 10 | 55° 59.74´с. ш. 92° 44.21´ в. д. | Сосняк разнотравный / P. sylvestris
H = 20–25; D = 25–30 |
P. avium, Crataegus sanguinea, Spiraea media, Cotoneaster melanocarpus, Sorbus sibirica / C. macroura, C. foetida | 2 |
| 23 | 55°59.76´с. ш. 92°44.18´ в. д. | Сосняк разнотравно-мертвопокровный / P. sylvestris
H = 20–25; D = 30–35 |
- / T. repens; A. pilosa;Plantago media | 3 |
| 18 | 56°00.69´ с. ш. 092°43.48´ в. д. | Березняк осочково-разнотравный / B. pendula H = 20–25; D = 20–25 |
C. melanocarpus, S. sibirica, Swida alba, Salix caprea / C. macroura; Rubus saxatilis; Lathyrus gmelinii | 1 |
| 20 | 56°00.60´ с. ш. 092°42.68´ в. д. | Сосняк орляковый / P. sylvestris H = 25–30; D = 35–50 |
S. sibirica, P. avium, R. acicularis / P. aquilinum, L. gmelinii, Brachypodium pinnatum | 1 |
| 21 | 56°00.59´ с. ш. 092°42.68´ в. д. | Березняк орляковый / B. pendula H = 22–25; D = 25–30 |
S. caprea, P. avium, / P. aquilinum, L. gmelinii, Milium effusum | 1 |
Примечание. * по шкале В. П. Чижовой (2011).
В древесном ярусе обследованных березняков преобладающей породой является Betula pendula Roth. Большинство деревьев являются перестойными, что может представлять опасность для отдыхающих из-за риска их падения во время сильного ветра. Поваленные деревья отмечаются на территории довольно часто.
Древесный ярус сосняков представлен Pinus sylvestris L. как естественного, так и искусственного происхождения. Возраст деревьев основного полога варьирует от 50 до 80 лет. На фоновой территории встречаются деревья более старшего возраста.
Подрост во всех сообществах в основном представлен сосной высотой до 5 м небольшого диаметра. Большую долю в подросте занимает береза вегетативного происхождения, единично встречаются особи темнохвойных пород. Сосновый подрост, как правило, угнетен (пожелтевшая хвоя, однобокость), что объясняется прежде всего недостатком света под пологом. В целом количество молодых деревьев невелико, поэтому на отдельных участках заложены лесные культуры из ели сибирской и сосны сибирской. В настоящий момент их высота достигает 4–5 м, жизненное состояние хорошее.
Аборигенные виды кустарников, наиболее часто встречающиеся в подлеске, – черемуха (Padus avium Mill.), шиповник (Rosa acicularis Lindl.), калина (Viburnum opulus L.), рябина (Sorbus sibirica Hedl.), кизильник (Cotoneaster melanocarpus Fisch. ex Blytt), карагана древовидная (Caragana arborescens Lam.), как правило, весьма декоративны. Черемуха привлекает посетителей своим пышным цветением в период с мая по июнь, а шиповник – с июня по июль. В конце лета – начале осени привлекательность для отдыхающих создают плодоносящие растения рябины, шиповника, калины и кизильника.
Сообщества орляковой серии типов леса описаны как в рекреационной части экопарка, так и на условно ненарушенной территории. В целом сообщества с орляком менее декоративны, поскольку его густой полог создает монотонность, а также сильно затеняет поверхность почвы и препятствует развитию красочных видов лесного разнотравья.
Березняки и сосняки разнотравные и орляковые относятся к производным типам леса и представляют собой ряды антропогенной трансформации зеленомошных типов леса (Горчаковский, Иванова, 2008). В число доминантов живого напочвенного покрова таких лесов входят Pteridium aquilinum и Carex macroura. Орляк обыкновенный (Pteridium aquilinum) относится к высококонкурентным видам и при создании благоприятных условий достаточно быстро разрастается. Его процветанию способствуют, с одной стороны, биологические особенности, с другой – различные антропогенные факторы (вырубки, пожары, умеренный сбор молодых вай, используемых в пищу, и др.) (Ершова, 2012). Осока большехвостая (Carex macroura) – вид с исключительно широким эколого-ценотическим ареалом, толерантный к недостатку света и влаги, но довольно требовательный к богатству почвы, благодаря чему растет как под пологом темнохвойных пород, так и в светлых лесах. Легко выдерживает конкуренцию с крупнотравьем, но чувствительна к воздействию зеленых мхов (Буторина, 1963). Как вид, образующий плотную дернину, осока, видимо, достаточно устойчива к антропогенной нагрузке, тем более что в этих условиях конкуренция со стороны мхов отсутствует.
В рекреационной части экопарка часто встречаются участки с механически нарушенным почвенным покровом из-за строительства объектов благоустройства. На таких участках отмечены заросли рудеральных видов растений Cirsium setosum (Willd.) Bess., Arctium tomentosum Mill., Urtica dioica L. и др. – бурьян. Заросли бурьяна визуально неприятны для отдыхающих.
В целом видовое флористическое богатство живого напочвенного покрова исследуемых сообществ на пробных площадях варьирует от 22 до 38 видов на 100 м2.
На рис. 2 представлена эколого-ценотическая структура березняков разнотравных (пробные площади № 1, 2, 3), на рис. 3 – эколого-ценотическая структура березняков орляковых (пробные площади № 4, 5, 20, 21) различных стадий дигрессии.

Рис. 2. Эколого-ценотическая структура березняков разнотравных различных стадий дигрессии: 1 – луговая и лугово-опушечная (Md), 2 – неморальная (Nm), 3 – бореальная (мелкотравье) (Br_m), 4 – бореальная (кустарнички и вечнозеленые травы) (Br_k), 5 – высокотравная (H), 6 – рудеральная (Ru)
Fig. 2. Ecological and cenotic structure of forb birch forests of different stages of digression: 1 - meadow and meadow-marginal (Md), 2 - nemoral (Nm), 3 - boreal (small grass) (Br_m), 4 - boreal (shrubs and evergreen grasses) (Br_k), 5 - tall grass (H), 6 – ruderal (Ru)
В эколого-ценотической структуре сообществ (рис. 2, 3) преобладают виды лугово-опушечной ЭЦГ (Md), на 2-й и 3-й стадиях дигрессии появляются рудеральные виды (Ru) при одновременном снижении количества мелкотравья и неморальной ЭЦГ. Отмечены такие рудеральные виды, как Taraxacum officinale (L.) Weber ex F. H. Wigg., Chenopodium album L., Cirsium setosum (Willd.) Besser ex M. Bieb., Arctium tomentosum Mill., Carduus crispus Guirão ex Nyman, Plantago lanceolata L., Stellaria graminea L. и др. В сообществах 3-й стадии дигрессии исчезает высокотравье (Hh).
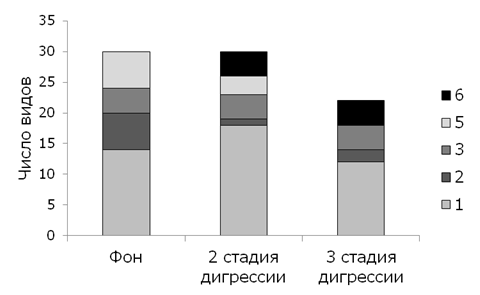
Рис. 3. Эколого-ценотическая структура березняков орляковых различных стадий дигрессии: 1 – луговая и лугово-опушечная (Md), 2 – неморальная (Nm), 3 – бореальная (мелкотравье) (Br_m), 4 – бореальная (кустарнички и вечнозеленые травы) (Br_k), 5 – высокотравная (H), 6 – рудеральная (Ru)
Fig. 3. Ecological and coenotic structure of bracken birch forests at different stages of digression: 1 - meadow and meadow-marginal (Md), 2 - nemoral (Nm), 3 - boreal (small grass) (Br_m), 4 - boreal (shrubs and evergreen grasses) (Br_k), 5 - tall grass (H), 6 – ruderal (Ru)
На рис. 4 (А, Б, В, Г) представлены ранговые распределения относительного обилия видов травяно-кустарничкового яруса для ряда пробных площадей. Ранговое распределение характеризуется следующим уравнением:
lns (i) = a – b lni (1),
где параметр а – логарифм обилия вида ранга 1, а параметр b характеризует конкуренцию между видами в сообществе. Точность приближения данных учетов с помощью уравнения (1) характеризуется коэффициентом детерминации R2. Чем ближе R2 к 1, тем корректнее представление данных с помощью уравнения (1).

Рис. 4. Ранговое распределение относительного обилия видов травяно-кустарничкового яруса: 1 – обильные виды, 2 – редкие и чрезвычайно редкие виды в следующих сообществах: А) березняк разнотравный (пробная площадь № 2, стадия дигрессии 2); Б) березняк разнотравный (пробные площади № 1 и 3, стадия дигрессии 3); В) березняк орляковый (пробные площади № 4 и 5, стадия дигрессии 2); Г) сосняк разнотравный (пробные площади № 10 и 23, стадии дигрессии 2 и 3 соответственно)
Fig. 4. Rank distribution of the relative abundance of grass and shrub tier species: 1 - abundant species, 2 - rare and extremely rare species in the following communities: А) forb birch forest (plot No. 2, digression stage 2); Б) forb birch forest (plots Nos. 1 and 3, stage of digression 3); В) bracken birch (plots No. 4 and 5, stage of digression 2); Г) forb pine forest (plots No. 10 and 23, stages of digression 2 and 3, respectively)
Как видно из рис. 4 (А–Г), все варианты ранговых распределений обилия для четырех сообществ показывают довольно равномерное убывание обилия (модель «разломанного стержня»). Известно, что, чем круче идет прямая, тем меньше общее разнообразие и сильнее доминирование одного или нескольких видов. В нашем случае прямые всех вариантов сообществ полого снижаются и коэффициенты b уравнения (1) невелики, что говорит о том, что разнообразие довольно высокое, но при этом имеет место доминирование нескольких видов. Вместе с тем в любом конкретном сообществе такое распределение обязательно имеет «хвост», образованный редкими видами (по шкале Браун – Бланке – редкие и чрезвычайно редкие). В целом для каждой прямой рангового распределения весь набор видов можно разделить на две части: малое число обильных видов (черным цветом) и большое количество редких видов (хвост). Так, например, на пробной площади №2 в березняке разнотравном (см. рис. 4 А) выявлено 29 видов, из которых 20 – виды редкие и чрезвычайно редкие, при этом первые два ранга в ряду распределения занимают Carex macroura с проективным покрытием в живом напочвенном покрове 65 % и Agrimonia pilosa – 30 %.
Для сравнения возьмем ранговое распределение видов на пробных площадях в березняке разнотравном (см. рис. 4 Б). Еще более пологая прямая, больше видов-доминантов: C. macroura с проективным покрытием 65 %, A. pilosa с проективным покрытием 30 %, Taraxacum officinale с проективным покрытием 25 %, Cimicifuga foetida с проективным покрытием 30 %, Fragaria vesca с проективным покрытием 25 %. «Хвост» из редких по обилию видов состоит из 14 видов на пробных площадях № 1 и 16 видов на пробной площади № 3.
Параметры регрессионных уравнений для рангового распределения на разных пробных площадях приведены в табл. 2.
Таблица 2. Параметры регрессионных уравнений для рангового распределения на разных пробных площадях
| Тип леса (на пробной площади №) | Общее количество видов, N |
Количество видов чрезвычайно редких и редких, N0 |
Статистические показатели |
Стадия дигрессии, d |
||||
| доля чрезвычайно редких и редких видов, p = N0 / N | σ(p) – стандартное отклонение р | логарифм обилия вида ранга 1, а | коэффициент, характеризующий конкуренцию между видами, b | коэффициент детерминации, R2 | ||||
| Березняк разнотравный (№ 2) | 29 | 20 | 0. 690 | 0.056 | 1.346 | 0.340 | 0.87 | 2 |
| Березняк орляковый (№ 4 и 5) | 41 | 32 | 0.780 | 0.065 | 1.313 | 0.490 | 0.86 | 2 |
| Сосняк разнотравный (№ 10) Сосняк разнотравно-мертвопокровный (№ 23) | 49 | 42 | 0.857 | 0.050 | 0.844 | 0.490 | 0.91 | 2.5 |
| Березняк разнотравный (№ 1 и 3) | 34 | 27 | 0.794 | 0.069 | 1.090 | 0.230 | 0.90 | 3 |
| Березняк орляковый (№ 21) | 36 | 30 | 0.830 | 0.062 | 1.348 | 0.458 | 0.66 | 1 |
| Сосняк орляковый (№ 20) | 35 | 29 | 0.829 | 0.064 | 1.348 | 0.458 | 0.66 | 1 |
| Березняк осочково-разнотравный (№ 18) | 30 | 27 | 0.900 | 0.055 | 1.325 | 0.670 | 0.87 | 1 |
Анализ табл. 2 показывает, что существует линейная связь между стадией d дигрессии и коэффициентом b рангового уравнения (1) (рис. 5).
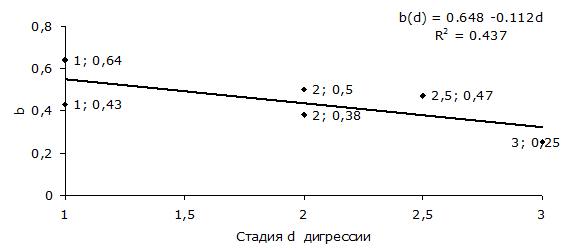
Рис. 5. Связь между стадией d дигрессии и коэффициентом b рангового уравнения (1)
Fig. 5. The relationship between the digression stage d and the coefficient b of the rank equation (1)
Из рис. 5 следует, что, чем сильнее антропогенное воздействие на сообщество, тем меньше коэффициент конкуренции между видами на территории. Чувствительность Χ = ∂b/∂d ≈ -0.11, т. е. с увеличением стадии дигрессии на 1 значение коэффициента b уменьшается на 0.11.
Итак, исследования видового разнообразия живого напочвенного покрова в растительных сообществах на территории экопарка показали, что происходит антропогенная трансформация естественных территорий. Наименее выражено антропогенное воздействие в сообществах на пробных площадях №18, 20, 21 (условно фоновые в табл. 1). Эти сообщества характеризуются высоким видовым богатством (32–37–38 видов на 100 м2 соответственно), что типично для данных типов леса. Данные пробные площади отнесли к I стадии дигрессии. Наибольшее число обследованных территорий (пробные площади № 2, 4, 5, 6, 9, 10) относятся ко второй стадии дигрессии. В сообществах этой стадии дигрессии количество видов может быть 25 (пробная площадь № 4), 29 (пробная площадь № 10) или даже немного больше (например, 38 видов на пробной площади № 5), в т. ч. за счет внедрения рудеральных видов. И наконец, пробные площади №1, 3, 7, 23 могут быть отнесены к третьей стадии дигрессии. Они сильно подвержены антропогенному влиянию. В этих сообществах отмечается снижение видовой насыщенности. В экологической структуре нарушенных сообществ повышена роль гелиофитов (Phlomoides tuberosa L., Centaurea scabiosa L., Geranium pseudosibiricum J. Mayer и др.).
Геоботанический анализ позволяет достаточно детально оценить состояние рекреационных территорий, однако проведение таких исследований связано со значительными затратами времени и требует от исследователей высокого профессионального уровня. Вряд ли такой метод можно рекомендовать для постоянного мониторинга. Альтернативой методам геоботанических исследований могут стать дистанционные наблюдения с использованием беспилотных летательных аппаратов (БПЛА). На рис. 6 приведены снимки БПЛА для разных территорий экопарка.

Рис. 6. Контрастированное изображение тропиночной сети ряда пробных площадей экопарка «Гремячая грива»
Fig. 6. Contrasted image of the pathway network on a number of the sample plots of the Gremyachaya griva ecopark
В программе Wing (авторская разработка А. В. Ковалева) было рассчитано количество пикселей для тропинок и лесных массивов. Результаты приведены в табл. 3.
Таблица 3. Расчеты числа пикселей в зоне тропинок на изученных пробных площадях
| Пробная площадь | Стадия дигрессии | Всего пикселей в расчете | Число пикселей в зоне тропинок | Доля пикселей в зоне тропинок |
| А) Сосняки северной части экопарка | 1 | 12000000 | 90633 | 0.008 |
| Б) Сосновые, березово-сосновые и березовые леса на территории пробных площадей № 7, 23 | 2–3 | 9341221 | 561484 | 0.060 |
| В) Сосняки у границы экопарка, на территории пробных площадей № 18, 20, 21 | 1 | 2080000 | 34470 | 0.017 |
Как следует из табл. 3, обследованные пробные площади сильно различались по доле площади, подвергшейся рекреационному воздействию, – от примерно 1 % площади территорий, слабо посещаемых населением, до 6 % площади, специально предназначенной для рекреации.
Заключение
В традиционных методах оценки нагрузки на экосистему используются либо учеты посещаемости территории, либо геоботанический анализ. Однако для надежной оценки посещаемости необходимы длительные наблюдения, т. к. посещаемость сильно колеблется на коротких интервалах времени. Поэтому такой метод требует большого числа наблюдателей, работающих в течение длительного времени. Следовательно, он крайне затратен и трудоемок. Геоботанический анализ, оценки соотношения видов травянистых растений, выделение доли редких и очень редких видов на пробных площадях позволяют в ходе единственного учета достаточно точно определить стадию дигрессии. Однако, если для проведения учетов посещаемости необходим персонал с минимальным профессиональным уровнем, то применение методов геоботанического анализа требует привлечения специалистов очень высокого уровня и больших трудозатрат на проведение исследований. Так, в нашей работе среднее время, необходимое для полевых учетов и последующей обработки данных для одной пробной площади, составило около 5 часов. Альтернативный вариант оценки стадии дигрессии по доле тропиночной сети на пробной площади не требует значительных затрат времени и знания ботанической номенклатуры, дистанционная оценка пробной площади с помощью БПЛА производилась в течение примерно 10 минут. Таким образом, меньшая точность дистанционного метода мониторинга компенсируется возможностями охвата больших территорий и большей повторности измерений. В связи с этим его можно рекомендовать для проведения регулярного мониторинга состояния растительных сообществ, подвергающихся антропогенному воздействию в сочетании с периодическими геоботаническими учетами.
Библиография
Буторина Т. Н. Эколого-ценотический анализ кустарничково-травяного яруса лесных ассоциаций // Типы лесов Сибири. М.: Изд-во АН СССР, 1963. С. 30–51.
Горчаковский П. Л., Иванова Л. А. Оценка состояния и прогноз антропогенной динамики растительного покрова особо охраняемых природных территорий на основе экологического картографирования // Экология. 2008. № 4. С. 243–252.
Грешилова Н. В., Скрипальщикова Л. Н. Оценка рекреационных нагрузок на пригородные сосновые и березовые насаждения г. Красноярска // Актуальные проблемы лесного комплекса. 2009. № 22. С. 81–84.
Дегтева С. В., Новаковский А. Б. Эколого-ценотические группы сосудистых растений в фитоценозах ландшафтов бассейна верхней и средней Печоры . Екатеринбург: УрО РАН, 2011. 196 с.
Ершова Э. А. Антропогенная трансформация в пригородных сообществах с участием в травостое папоротника орляка (Pteridium pinetorum) // Растительный мир Азиатской России. 2012. № 2 (10). С. 132–138.
Казанская Н. С., Ланина В. В., Марфенин Н. Н. Рекреационные леса (состояние, охрана, перспективы использования) . М.: Лесная промышленность, 1977. 95 с.
Методы изучения лесных сообществ / Е. Н. Андреева и др.; Отв. ред. В. Т. Ярмишко, И. В. Лянгузова. СПб.: НИИХимии СПбГУ, 2002. 240 с.
МКЭО «Гремячая грива» . СПб., 2022. 182 с.
Скрипальщикова Л. Н. Экологические проблемы пригородных лесов // География и природные ресурсы. 2008. № 1. С. 50–54.
Ценофонд лесов Европейской России: База данных. 2012. URL: http://cepl.rssi.ru/bio/flora/main.htm (дата обращения: 20.02.2023).
Чижова В. П. Рекреационные ландшафты: устойчивость, нормирование, управление (Recreational landscapes: stability, regulation, management)]. Смоленск: Ойкумена, 2011. 176 с.
Экопарк «Гремячая грива»: Официальный сайт. 2018. URL: https://visitsiberia.info/eko-park-gremyachaya-griva.html (дата обращения: 20.02.2023).
Matthews T. J., Whittaker R. J. On the species abundance distribution in applied ecology and biodiversity management // Journal of Applied Ecology. 2015. Vol. 52. P. 443–454.
McGill B. J., Etienne R. S., Gray J. S., Alonso D., Anderson M. J., Benecha H. K. & White E. P. Species abundance distributions: moving beyond single prediction theories to integration within an ecological framework // Ecology letters. 2007. Vol. 1 (10). P. 995–1015.
The Plant List. 2013. URL: www.plantlist.org.
Благодарности
Работа поддержана грантом Красноярского краевого фонда науки № КФН-771, проектом FSRZ-2023-0007.

 © 2011 - 2026
© 2011 - 2026