



Введение
Несмотря на неослабевающий интерес к изучению биологии средней бурозубки, в т. ч. и в связи с ее относительной малочисленностью и своеобразием существования в европейской части ареала, экология и особенно размножение и динамика численности вида в этих условиях по-прежнему остаются исследованными совершенно недостаточно. Настоящее сообщение, основанное на многолетних (1958–2018 гг.) стационарных и экспедиционных исследованиях, охвативших территорию Карелии и прилежащих регионов Восточной Фенноскандии, представляет собой попытку восполнить этот существенный пробел. Обитая здесь на северо-западной периферии ареала, средняя бурозубка вырабатывает весьма эффективные структурно-популяционные адаптации и благодаря этому успешно преодолевает экстремальные экологические условия, сохраняя достаточно высокие показатели репродукции и стабильное положение в формирующемся населении мелких млекопитающих региона.
Материалы
Материалом для настоящего сообщения послужили многолетние сборы зверьков изучаемого вида, составившие в сумме 1879 добытых и исследованных средних бурозубок.
Методы
Для изучения размножения всех отловленных зверьков разделили на две возрастные группы. При этом в качестве основных возрастных критериев использовали форму (конфигурацию и состояние швов) черепа и степень стертости зубов, а в качестве второстепенных (вспомогательных) – особенности снашивания волосяного покрова на хвосте, лапах и ушных раковинах. Кроме того, в сомнительных случаях привлекали и некоторые другие показатели, в частности размеры тимуса, который, как известно, полностью инволюирует к осени первого года жизни и у перезимовавших зверьков практически не выражен.
Особи первой возрастной группы (сеголетки) характеризуются высокой сжатой с боков черепной коробкой (череп «вздутый») и отсутствием гребней. Швы между костями хорошо заметны. Зубы имеют острые вершины и гребни. Эмаль коронок не стерта. Половая система, как правило, недоразвита. Тимус хорошо развит, его абсолютный вес достигает 30–170 мг. Хвост равномерно покрыт волосами («пушистый»). Волосяной покров на лапках и ушных раковинах хорошо развит и почти не имеет следов снашивания. В подавляющем большинстве случаев это неполовозрелые зверьки (subadultus).
Зверьки второй возрастной группы (перезимовавшие) отличаются утолщенным черепом с хорошо развитыми гребнями и слабо заметными швами, последние имеют зигзагообразный рисунок. Вершины промежуточных зубов в результате снашивания округленные, коренные зубы уплощены, с тупыми конусами и гребнями. Пигментация на зубах почти не заметна. Волосяной покров хвоста, лап и ушных раковин сильно сношен, а местами совсем отсутствует («лысины»). Это половозрелые зверьки (adultus).
Различаются возрастные группы и по строению половой системы. У неполовозрелых самцов семенники очень малы и, как правило, имеют округлую форму (диаметр около 1 мм, вес не более 5–6 мг). Придатки семенника и семенные пузырьки недоразвиты. У половозрелых, перезимовавших бурозубок семенники значительно крупнее, имеют продолговатую форму (длина 5–16 мм, ширина 3–8 мм) и хорошо заметны даже у невскрытых зверьков (в виде двух вздутий на нижней стороне тела). Придатки семенника, простатическая и куперова железы и семенные пузырьки очень хорошо выражены, последние наполнены спермой и сильно вздуты. У самцов, участвующих в размножении, кроме того, увеличены боковые кожные железы.
У молодых неполовозрелых самок матка полупрозрачная, тонкая и короткая, общая длина ее рогов составляет в среднем 8.6 (3.2–16.1) мм, тела 10.9 (1.5–24.3) мм. У взрослых зверьков тело матки составляет в среднем 59 % от длины рогов, а у молодых – 76 %.
У кормящих или ранее кормивших самок соски обсосаны и волосяной покров вокруг них сильно вытерт (особенно если самка кормит второй выводок), но у первых млечные железы лактируют, а у вторых – нет. У беременных на ранних стадиях в яичниках хорошо заметны розоватые желтые тела, позднее – эмбрионы на рогах матки, а сама матка сильно увеличена в длину и ширину. Плацентарные пятна образуются, но очень быстро (уже на 2–3-й день после родов) исчезают. Иногда удается определить и недавно оплодотворенных самок, признаки беременности у которых еще не выражены. В матке у таких самок бывает заметна мутная беловатая жидкость – сперма.
Сроки и интенсивность размножения зверьков определялись на основании предпринятого нами специального гистологического исследования полового цикла самцов и самок (Ивантер, 1975). Остальные исследования проводились по общепринятым методикам, неоднократно описанным в наших предыдущих публикациях (Ивантер, 1975, 2018, 2019, 2020а, б; Ивантер, Макаров, 2001), а также в методической сводке Е. В. Карасевой с соавторами (2008).
Кроме того, при анализе разовой плодовитости и других показателей репродукции средней бурозубки и расчетов их достоверности, а также при оценке влияния экзо- и эндогенных факторов на ход и интенсивность размножения и динамику численности популяции нами применялись статистические приемы, включающие в себя проверку выборок на нормальность распределения, вычисление средних и их статистических ошибок (методы Стьюдента и Фишера) и проведение регрессионного и дисперсионного анализов, а также определение симметричности и асимметричности распределений, в т. ч. для выявления репродуктивных тенденций в микроэволюции (Ивантер, Коросов, 1992, 2011).
Изучение линьки волосяного покрова средней бурозубки, как и других мелких млекопитающих (Ивантер, 2021), проводилось обычными, использовавшимися в аналогичных исследованиях методами (Кузнецов, 1932; Бигман и др., 1948; Депарма, 1951; Когтева, 1963; Соколов и др., 1986). При этом особенно широко применялось «картирование» – зарисовка на коллекционных карточках с трафаретами контура шкурки изменений окраски мездры и расположения линных пятен. Кроме того, в световом микроскопе определяли функциональное состояние наружных покровов, дифференцировали зрелые волосы от растущих, различали пигментированные и непигментированные участки закладки волоса у изученных образцов кожи и волос. Необходимость световой микроскопии была в данном случае вызвана тем, то только с ее помощью можно, например, безошибочно отличить растущий зимний волос от окончательно выросшего летнего той же длины. Для этого достаточно рассмотреть под бинокулярным микроскопом строение волосяной луковицы: у зрелых волос оно закрытого (колбовидного) типа, а у растущих – открытого (сосочкового). Кроме того, закладка волос не везде сопровождается потемнением мездры. Поскольку волосы землероек имеют зонарную окраску и их концевые сегменты на разных участках шкурки неодинаковы по цвету (на спине темно-коричневые, на боках – бурые, а на брюхе – белые), то на боках и спине линные пятна бывают видны сразу, а на брюхе – лишь после того, как пробьются через кожу былые верхушки волос. Сегменты же стержня, лежащие ниже вершины, окрашены в черный цвет. Поэтому при микроскопии кожи с брюшка без видимых признаков линьки легко обнаруживаются характерные изменения (в частности, функционирующие волосяные фолликулы), свидетельствующие о сезонной смене волос при так называемой скрытой пигментации.
Трудности в изучении линьки землероек связаны также с разновозрастным составом популяции. Продолжительность линьки одного зверька, как известно, невелика (Borowski, 1958). Но у всей популяции этот процесс растянут, и в один и тот же период в ее составе встречаются особи, находящиеся на разных стадиях линьки. Отсюда неизбежные сложности в определении последовательности и топографии сезонной смены шерсти.
Определение теплоизоляционных свойств шкурок (по удельной теплопроводности) выполняли с помощью специального прибора ИТ-3, изготовленного по нашему заказу для измерения теплопроводности небольших по размеру неметаллических объектов в Киевском институте технической теплофизики АН УССР. Существенное преимущество этого прибора заключается в большой чувствительности и точности, а также в том, что все необходимые для расчета теплопроводности величины получают при его использовании непосредственным измерением.
Результаты
Размножение
При выделении возрастных групп у средней бурозубки мы руководствовались теми же признаками, что и в отношении других представителей рода Sorex: выраженностью черепных гребней и швов между костями, степенью стертости зубов, опушенности хвоста, лап и ушей. Учитывались также развитие тимуса (у зимовавших он не развит) и состояние половой системы (табл. 1). Вместе с тем средняя бурозубка имеет некоторые специфические особенности, отличающие ее от других видов землероек. Прежде всего это касается размеров гонад, по которым средняя бурозубка уступает не только обыкновенной, но и малой бурозубке. Однако в этих рамках возрастные отличия выражены у нее достаточно четко, особенно в отношении длины и веса семенников. Что же касается размеров матки, то по этому признаку средних бурозубок разного возраста отличить трудно (крайние значения перекрываются, а средние довольно близки) – во всяком случае труднее, чем у других видов бурозубок.
Таблица 1. Характеристика половой системы средней бурозубки
| Показатель | Зимовавшие | Прибылые |
| Самцы | ||
| Число исследованных зверьков | 28 | 205 |
| Длина семенника, мм | 6.4 (4.3–12.7) | 1.1 (0.6–3.1) |
| Длина семенных пузырьков, мм | 5.4 (4.5–6.5) | – |
| Вес двух семенников, мг | 123 (71–144) | 2.4 (1.0–5.2) |
| Самки | ||
| Число исследованных зверьков | 17 | 147 |
| Суммарная длина рогов матки, мм | 10.5 (6.5–16.2) | 7.0 (2.3–11.9 ) |
| Длина влагалища (тела матки), мм | 4.4 (2.3–6.7) | 3.6 (1.0–6.4) |
Можно отметить и видовую специфику в строении матки. У средних бурозубок индекс матки (отношение длины влагалища к суммарной длине рогов) составляет у молодых 50, а у старых 40 %. Это говорит, во-первых, о том, что половое созревание сказывается преимущественно только на удлинении рогов (длина влагалища почти не меняется с возрастом), во-вторых, об ином строении матки у рассматриваемого вида. В сравнении с маткой обыкновенной землеройки матка средней бурозубки имеет более короткое тело и длинные рога, а в сопоставлении с малой, наоборот, более длинное тело и короткие рога. Напомним, что индекс матки у обыкновенной бурозубки составляет 65–75, у малой – 35–46 %.
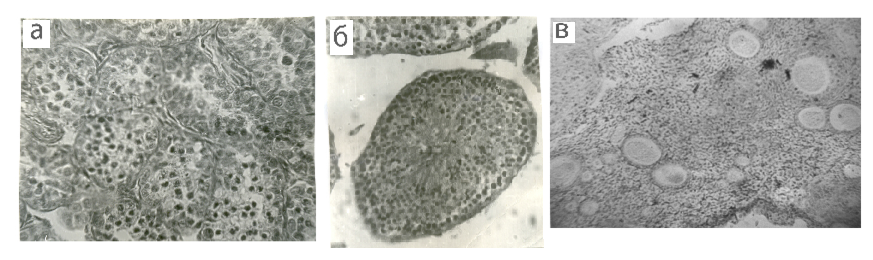
Рис. 1. Поперечные срезы извитых канальцев семенника в состоянии покоя (а), активного сперматогенеза (б) и яичника самки в стадии фолликулярной активности (в) у средней бурозубки. Микрофото, х400
Fig. 1. Transverse sections of the convoluted tubules of the testis at rest (a), active spermatogenesis (б) and the female ovary at the stage of follicular activity (в) in the common shrew. Microphoto, x400
Мы не располагаем достаточным материалом, позволяющим точно установить фенологические сроки начала размножения средних бурозубок в Карелии. Это объясняется тем, что по большинству лет наблюдений отловленные перезимовавшие зверьки оказывались уже вступившими в размножение. Однако в те годы, когда работа на стационарах начиналась еще весной, отловы зверьков этого вида свидетельствовали об отсутствии у них признаков размножения (гона и течки) и в марте, и в первой половине апреля. Лишь в середине апреля часть зверьков уже готова к размножению. В эти сроки у самцов размеры семенников увеличиваются до 7.5–8.1 мм, а их вес – до 160–180 мг. К этому времени достигают половой зрелости и самки. Суммарная длина рогов составляет у них 8.2–8.9 мм. Так что гон приходится в наших условиях, как, впрочем, и на большей части видового ареала, на конец апреля – начало мая. Своеобразие хода гаметогенеза у самцов и самок средней бурозубки подтверждают и гистологические исследования (рис. 1).
Первые беременные самки были добыты в Карелии 26.06.1959, 17.06.1986 и 19.06.2004 г. Однако сезон размножения у средних бурозубок начинается здесь, очевидно, значительно раньше, поскольку в год массовых весенне-летних отловов в заповеднике «Кивач» (1959) молодых зверьков мы отлавливали, например, начиная с 11 июня. Судя по этой дате, спаривание средних бурозубок происходит в наших условиях в конце апреля, т. е. несколько раньше, чем у обыкновенной и малой. Самцы же этого вида с уже увеличенными семенниками, свидетельствующими об активном сперматогенезе, отлавливались нами с 22 апреля.
Таким образом, наиболее вероятные сроки начала репродуктивного цикла средней бурозубки в исследуемом регионе приходятся на последнюю декаду апреля – начало мая.
Отмеченные фенологические сроки начала репродуктивного периода подтверждаются и сопоставлением сроков первого появления в уловах молодых зверьков с известными периодами беременности (20 дней; Dehnel, 1952) и выкармливания молодых в гнезде (20–21 день; Dehnel, 1950; Abe, 1968). Так, в Карелии первые молодые особи этого вида начинают попадаться в ловушки и цилиндры в первой половине июня (8.06 и 11.06.1959, 7.06.1967, 7.06.2003 г.), в массе же это наблюдается в период с 10 по 21 июня, что соответствует началу периода спаривания средних бурозубок в конце апреля – начале мая. В 1959 г. последние беременные самки этого вида были отловлены 18.10 и 12.11.1959 г., правда, в другие годы их поимки приходились на гораздо более ранние сроки: обычно середину – конец сентября. Таким образом, весь период размножения у средней бурозубки продолжается в наших условиях около 5 месяцев, но наиболее интенсивно протекает в июле – августе.
Из добытых нами во все время исследований взрослых самок 6 оказалось только что оплодотворенными, 25 – беременными, 9 – одновременно беременными и подсосными (лактирующими), 13 – лактирующими и 8 – со следами недавнего размножения. Лишь у двух самок признаков размножения не было обнаружено вообще. Это говорит о достаточно большой интенсивности репродукции данного вида.
Взрослые самцы готовы к спариванию уже в апреле. У всех добытых в этот месяц зимовавших самцов семенники были достаточно упругие, превышали в длину 7 мм и содержали в придатках массу зрелых сперматозоидов. С этого момента они сохраняют сексуальную активность не только весь летний сезон, но и осенью. Во всяком случае, признаки затухания половой функции (потеря упругости семенников и др.) появляются у самцов чуть ли не на месяц позднее, чем у самок. В целом же степень сперматогенной активности семенников, если о ней можно судить по размерам и внешнему виду этого органа, у самцов средней бурозубки в течение всего лета и в начале осени остается на одном уровне.
Вплоть до сентября размножение продолжается весьма интенсивно, но в середине этого месяца, по-видимому, и заканчивается. Что же касается упомянутых выше случаев обнаружения беременных зверьков в октябре, тем более в ноябре (1959 г.), то их, скорее всего, следует отнести к артефактам.
Эмбрионов у одной самки было от 5 до 11, в среднем (n = 21) 7.7. Это несколько больше, чем у других видов землероек, и выше, чем у того же вида в других частях ареала (табл. 2). По рогам матки эмбрионы распределялись довольно равномерно: в правом обнаружено в среднем 7.4, в левом – 7.3. Рассасывания зародышей не отмечено.
Число генераций в течение года не менее двух, что доказывается поимкой кормящих самок, имеющих новую беременность. Кроме того, и в отловах молодых землероек намечаются два довольно заметных пика: 20–30 июня и 26 июля – 19 августа. Возможно, что часть самок имеет три выводка в течение лета, но прямых доказательств этому пока нет. Случаев размножения сеголеток мы не отмечали, и сомнительно, чтобы в наших условия они вообще могли иметь место. В других регионах средней тайги Европейского Севера и Зауралья такое хотя и случается, но достаточно редко: доля сеголеток, участвующих в размножении, не превышает там 1.2–1.8 % (Рамазанова, 1983; Бобрецов, 2004). В то же время в Южной Сибири, Северо-Восточной Азии и Пермской области созревание и участие в размножении прибылых средних землероек – более или менее нормальное явление, охватывающее от 4–8 до 15–17 % и даже больше зверьков этой возрастной группы (Реймерс, 1966; Долгов и др., 1968; Ревин, 1989; Докучаев, 1990; Куприянова, 1994; Бобрецов, 2004). По данным Н. Е. Докучаева, сеголетки у средних бурозубок вступают в размножение, как правило, при низкой численности популяции. Но первым на обратную зависимость между числом зверьков, созревающих в год рождения, и общей численностью популяции обратил внимание Г. Штайн (Stein, 1961). Участие в размножении самок-сеголеток, таким образом, может являться у этого вида важным популяционным регуляторным механизмом, компенсирующим низкую численность бурозубок в отдельные годы.
Таблица 2. Плодовитость средней бурозубки в разных частях ареала
| Район исследований | n пределы | Число эмбрионов на самку | Среднее | Источник |
| Карелия | 21 | 5–1 | 7.7 | наши данные |
| Башкирия | 7.1 | Снигиревская, 1947 | ||
| Волжско-Камский край | 3 | 6–8 | 7.0 | Попов, 1960 |
| Западная Сибирь | 25 | 2–11 | 7.0 | Юдин, 1962 |
| в т. ч. | ||||
| Томская область | 19 | 5–9 | 6.1 | Юдин, 1962 |
| Там же | 5 | 4–8 | 6.7 | Глотов и др., 1978 |
| Барабинская низменность | 4 | 5–8 | 6.3 | Глотов и др., 1978 |
| Приобье (Алтайский край) | 5 | 7–11 | 7.0 | Юдин, 1962 |
| Горный Алтай | 3 | 5–6 | 5.3 | Юдин, Барсова, 1967 |
| Юг Средней Сибири | 6.0 | Реймерс, 1966 | ||
| Верхняя Лена | 16 | 1–11 | 6.0 | Реймерс, Воронов, 1963 |
| Якутия | 10 | 2–9 | 6.6 | Тавровский и др., 1971 |
| Северный Сахалин | 2 | 6.0 | Воронов, 1993 | |
| Север Дальнего Востока | 8 | 4–8 | 7.1 | Юдин и др., 1976 |
| Бассейн Омолона | 54 | 3–9 | 8.3 | Докучаев, 1990 |
| Восточное Прибайкалье | 22 | 5–9 | 7.6 | Матурова, 1982 |
| Бассейн Чемиджа | 44 | 2–11 | 7.5 | Докучаев, 1990 |
| Южная Якутия | 22 | 1–8 | 5.7 | Ревин, 1989 |
| Алтай | 20 | 4–8 | 5.4 | Юдин и др., 1979 |
| Урал | 11 | 4–9 | 7.5 | Большаков и др., 1996 |
| Предверхоянье | 11 | 5–11 | 8.3 | Ревин и др., 1988 |
| Коми | 26 | 4–10 | 6.6 | Куприянова, 1994 |
| Печоро-Илычский заповедник | 36 | 2–10 | 7.1 | Бобрецов, 2004 |
| Алтае-Саянская горная страна | 21 | 4–14 | 9.5 | Юдин и др., 1979 |
| о. Хоккайдо | 8 | 4–8 | 7.1 | Abe, 1968 |
Экологическая структура популяции
О темпах воспроизводства популяции средней бурозубки в Карелии говорят и особенности динамики ее возрастного и полового состава (табл. 3). Так, уже в середине июня прибылые составляют в среднем 72.4 % от общего числа отлавливаемых зверьков. К осени доля сеголеток еще более возрастает, а в августе – сентябре варьирует в пределах 89–93 %, после чего к началу октября полностью вымирает. Это свидетельствует о достаточно высокой интенсивности размножения рассматриваемого вида в нашем регионе и общем высоком уровне воспроизводства популяции. По годам возрастной состав популяции также меняется, коррелируя с интенсивностью размножения и общей численностью популяции. Чем выше уровень размножения, тем больше доля молодых в отловах (1961, 1969, 1974, 1989, 2001 гг.). И наоборот, в годы с низким темпом репродукции, вследствие малочисленности молодых зверьков, возрастает удельный вес взрослых (зимовавших) особей (1976–1978, 1981–1982, 1996–1997, 2011 гг.).
Соотношение полов зависит от возраста, сезона года и состояния популяции. Среди перезимовавших зверьков преобладают самцы, особенно в первую половину лета, когда они отличаются повышенной активностью, а самки заняты выкармливанием молодняка, отсиживаются в гнездах и редко выходят на поверхность. У прибылых зверьков также отмечается несколько большая численность самцов, но причина здесь иная. Возможно, как и у обыкновенной бурозубки, это связано с преобладанием особей мужского пола среди эмбрионов.
Таблица 3. Возрастной и половой состав популяции средней бурозубки в Карелии
| Место и сроки исследования | Число прибылых | Число зимовавших | Общее число зверьков | |||||
| абс. | из них самцы, % | χ2 | абс. | из них самцы, % | χ2 | абс. | из них зимовавшие, % | |
| Заповедник «Кивач» (1959–1961) | 86 | 53.5 | 6.0 | 34 | 50.0 | 0.1 | 120 | 28.3 |
| Пудожский район (1960–1962) | 84 | 42.8 | 0.1 | 3 | – | – | 87 | 3.5 |
| Питкярантский район (1967–2014) | 843 | 55.0 | 1.8 | 112 | 76.7 | 1.0 | 959 | 11.7 |
| Пряжинский район (2004–2016) | 281 | 55.9 | 1.7 | 45 | 64.4 | 0.2 | 332 | 13.6 |
| Всего | 1294 | 54.5 | 1.6 | 194 | 68.0 | 0.1 | 1498 | 13.0 |
| По месяцам: июнь | 155 | 58.1 | 5.8 | 82 | 82.9 | 4.5 | 258 | 28.7 |
| июль | 421 | 52.5 | 5.0 | 54 | 63.0 | 0.1 | 447 | 10.1 |
| август | 544 | 52.8 | 0.2 | 52 | 36.5 | 0.9 | 647 | 6.6 |
| сентябрь | 113 | 49.6 | 2.8 | 6 | – | – | 114 | 3.5 |
| октябрь | 52 | 69.2 | 2.1 | – | – | – | 26 | – |
| ноябрь – январь | 9 | 66.7 | 5.1 | – | – | – | 6 | – |
Тем не менее уже в июле соотношение полов у взрослых особей выравнивается, а к концу репродуктивного сезона наблюдается численное преобладание самок. Снижение же интенсивности размножения к осени приводит к тому, что все большее количество самок, закончивших кормление выводков, становятся активнее и подвижнее, выходят на поверхность и в результате в большем количестве попадаются в давилки и цилиндры. Что же касается самцов, то с окончанием репродуктивного периода их активность снижается. Они почти на 1–2 месяца раньше самок стареют и вымирают (Dehnel, 1950).
Линька
Первая линька у прибылых бурозубок происходит осенью. Согласно наблюдениям Н. Е. Докучаева (1983), в дальневосточном регионе она начинается у зверьков всегда на дорсальной стороне тела, обычно на задней части спины в районе крестца (рис. 2А), но может начинаться на голове или на холке (рис. 2Б). В связи с этим несколько различается и дальнейший порядок смены меха, что можно наблюдать на вышеуказанном рисунке. Иногда закладка пигмента происходит на этих трех участках одновременно. Далее пигмент полосой захватывает всю спинную часть тела и верх головы. На следующем этапе линька заходит на бока и охватывает всю голову. Зимний мех вначале появляется на голове и спине, причем нередко это происходит до того, как появится пигмент на нижней стороне тела. Реже пигментированной бывает вся поверхность шкуры. В любом случае, пигмент вначале исчезает на голове. В последнюю очередь линяет мех у передних ног, на животе и в паху. У средней бурозубки дольше всего пигмент сохраняется в паховой области в отличие от обыкновенной бурозубки, у которой, по данным С. Боровского (Borowski, 1968), осенняя линька заканчивается на горле.

Рис. 2. Изменения в пигментации мездры при осенней линьке у сеголеток средней бурозубки (по: Докучаев, 1983). А – линька начинается на огузке; Б – линька начинается на холке или на голове.
Fig. 2. Changes in the pigmentation of the mezra during autumn molting in this year brood of the common shrew (according to: Dokuchaev, 1983). A - molting begins on the rump; Б - molting begins at the withers or on the head
Все исследователи при описании осенней линьки у бурозубок указывали, что линька осенью начинается на крестце и распространяется вперед по спине. Наши материалы 1976 и 1977 гг. по Карелии согласуются с этим положением (табл. 4). Однако в 1975 г. осенняя линька началась на холке и на голове одновременно. Этот год особенно выделялся тем, что у многих бурозубок (особенно у самцов) мездра на голове и в передней части спины очистилась от пигмента еще до того, как на брюшной стороне тела начался процесс линьки.
Волновой тип линьки в осенний период, наблюдавшийся у отдельных экземпляров обыкновенной и малой (Sorcx minutus L.) бурозубок (Borowski, 1964, 1973), нами не отмечен. Все экземпляры средней бурозубки в нашем материале имели полную осеннюю линьку.
Нами не было обнаружено половых различий в ходе осенней линьки у сеголеток, однако имелись четкие различия в сроках и ходе их осенней линьки в разные годы. Данные о соотношении линяющих зверьков в июле – сентябре за три года исследований (см. табл. 4) показывают, что если в 1976 и 1977 гг. в июле зверьки не линяли, то в этом же месяце 1975 г. с линькой было отмечено 8.2 % отловленных бурозубок. Первые зверьки с пигментацией на мездре отловлены 20 июля 1975 г. В 1976 и 1977 гг. первые линяющие средние бурозубки были отмечены 7 августа. В августе 1975 г. из 92 отловленных зверьков 6 (6.5 %) уже полностью сменили летний мех или имели остатки пигмента у передних ног и в паху. В августе 1976 г. бурозубки в зимнем меху не отлавливались. Зимний мех в августе 1977 г. имели только два зверька из 179 (1.1 %).
Анализ метеорологических данных за эти годы показал, что сроки начала осенней линьки не коррелируют с температурой. Так, 1976 г. характеризовался более холодным летом; средняя температура августа в этом году была ниже, чем в 1975 и 1977 гг., соответственно на 2.1 и 0.5°.
Тем не менее в августе 1976 г. линяло наименьшее число зверьков. С. Боровский (Borowski, 1964) при изучении влияния температуры на сроки осенней линьки у обыкновенной бурозубки при содержании в неволе также пришел к заключению, что разница температур в 2–3° между опытными группами не повлияла на сроки начала линьки.
Таблица 4. Интенсивность осенней линьки сеголеток средней бурозубки в разные годы исследований в Карелии (по: Ивантер и др., 1985) (данные по самцам и самкам объединены)
| 1975 г. | 1976 г. | 1977 г. | |||||||
| n | из них с линькой | % | n | из них с линькой | % | n | из них с линькой | % | |
| Июль | 147 | 12 | 8.2 | 30 | – | – | 27 | – | – |
| Август | 92 | 25 | 27.2 | 121 | 6 | 5 | 179 | 31 | 17.3 |
| Сентябрь | 30 | 9 | 30 | 25 | 20 | 80 | 21 | 12 | 57.1 |
Причина изменений сроков начала осенней линьки заключается в разном возрастном составе сеголеток в указанные годы. В более ранних исследованиях уже отмечалось (Шварц, 1955), что у Sorex araneus первыми начинают линьку зверьки весенних пометов, т. е. более старшие по возрасту. Это подтверждается и нашими исследованиями по средней бурозубке. В 1975 г. при высокой весенней численности бурозубок в размножении участвовали только зимовавшие самки. В течение лета они принесли три помета. Линька у сеголеток началась в июле, и количество линяющих зверьков равномерно нарастало до сентября. Растянутый период осенней линьки в 1975 г. связан с тем, что прибылые бурозубки вступали в линьку в зависимости от своего календарного возраста как бы поэтапно. В 1976 и 1977 гг. весенняя численность бурозубок была низкая. В эти годы в размножении принимали участие самки-сеголетки первого помета, которые и дали основной прирост популяции. Кроме того, в 1976 г. из-за холодной затяжной весны часть первых пометов, очевидно, погибла. Так, сеголетки начали ловиться лишь в июле, более чем на 20 дней позже обычного. В результате этого осенью 1976 и 1977 гг. популяции оказались более однородными по возрастному составу. В основном это были зверьки второго поколения. Линька их проходила в одно время, поэтому процент линяющих зверьков в эти годы резко возрастал от августа к сентябрю. Особенно ярко это было выражено в 1976 г. (см. табл. 4).
К зиме сеголетки средних бурозубок имеют более длинный зимний мех. Высота его увеличивается от 4.0 до 7.3 мм (табл. 5). Значительно изменяется и густота меха. Измерения, проведенные на обыкновенной бурозубке, показали, что густота зимнего меха на 31 % выше летнего (Borowski, 1958), а его теплоизолирующие свойства – примерно на 18 % (Gebczynski, Olszewski, 1963).
Таблица 5. Сезонная изменчивость высоты меха у различных половозрастных групп средней бурозубки Карелии*
| Зимний мех | Весенний мех | Летний мех | ||||
| n | М ± m | n | М ± m | n | М ± m | |
| Зимовавшие самцы | 32 | 7.3 ± 0.09 | 68 | 0.1 ± 0.04 | 36 | 3.7 ± 0.05 |
| Самки | 80 | 7.3 ± 0.05 | – | – | 53 | 4.2 ± 0.03 |
| Сеголетки | 14 | 7.3 ± 0.08 | – | – | 40 | 4.0 ± 0.03 |
Примечание. * – высота зимнего меха измерялась у сеголеток осенью, а у зимовавших – в начале года.
Весенняя линька у самцов. Первые небольшие пигментные пятна у особей мужского пола появляются на шкурке в конце февраля. В течение второй половины марта у самцов происходит линька меха на голове и на боковых железах. Именно в это время происходит половое созревание и идет интенсивный рост генеративных органов. Имеется четкая зависимость между сроками начала линьки и половым развитием животных. У двух самцов, отловленных 13 и 14 марта 1974 г., линькой была занята почти вся поверхность кожи, а один самец от 14 марта того же года уже перелинял и имел остатки пигмента только на огузке. Вес их семенников равнялся соответственно 62, 80 и 102 мг, в то время как средний вес семенников в марте у других самцов был равен 27.2 мг (=22). С другой стороны, в первой декаде апреля того же года было отловлено четыре самца, у которых сперматогенез еще не начался. Средний вес семенников у них был равен 48 мг, тогда как все остальные самцы в апреле имели активный сперматогенез, и средний вес семенников в популяции равнялся 117.8 мг. Линька у этих 4 экземпляров шла вокруг боковых желез и на голове, и только у одного зверька начал линять живот, в то время как у большинства зверьков в это время зимний мех уже сменился на весенний.
Линька на боковых железах начинается в центре и, постепенно расширяясь, захватывает всю их поверхность (рис. 3А). Линькой в этот период занята незначительная площадь шкуры. С 25 марта по 15 апреля (только 3 экз., отловленные 13 и 14 марта, линяли раньше) происходит смена меха на всей шкуре (весенняя линька). От паховой области вдоль живота она идет вперед по телу, заходит на бока и на голову и распространяется вдоль спинной части тела. На боковых железах, а у части зверьков и на голове, пигмента к этому времени уже нет. Очищение мездры происходит в той же последовательности, как начиналась линька, т. е. освобождается от пигмента вначале вся нижняя сторона тела и затем голова. Дольше всего пигмент сохраняется на спинной стороне тела.

Рис. 3. Изменения в пигментации мездры при весенних и старческой линьках у средней бурозубки (по: Докучаев, 1983). А – весенняя линька I у самцов; Б – весенняя линька II у самцов; В – весенняя линька I у самок; Г – весенняя линька II у самок; Д – старческая линька (самцы и самки вместе)
Fig. 3. Changes in the pigmentation of the mezra during spring and senile molting in the common shrew (according to: Dokuchaev, 1983). A – spring molt I in males; Б – spring molt II in males; В – spring molt I in females; Г – spring molt II in females; D – senile molting (males and females together)
У многих зверьков на огузке зимний мех не сменяется в течение первой весенней линьки и остается до второй весенней линьки (рис. 3Б, нижняя ветвь). Зимний мех тогда выделяется хорошо заметным островком более длинного волоса. Отсутствие пигмента в таких случаях указывает на то, что мы имеем дело с прерванной линькой. У 5 экз. отмечен волновой тип линьки.
Весенняя линька II начинается у самцов во второй половине мая и имеет обратное направление, т. е. начинается на спине и заканчивается на голове и вентральной стороне тела (см. рис. 3Б). Она начинается обычно на огузке, где еще сохранялся зимний волос, и захватывает всю спину. Следовательно, зимний мех на части спинной стороны тела сменяется непосредственно на летний. Смена зимнего меха сразу на летний также известна для обыкновенной бурозубки (Skaren, 1973). С. Боровский (Borowski, 1968), однако, замечает, что у Sorеx araneus зимний мех после перерыва всегда сменяется весенним до начала второй весенней линьки. 1 экз., выделенный им в первой декаде мая, имел весенний мех на боках и животе (при отсутствии пигмента на этих участках тела). На спинной стороне у этого экземпляра кожа была пигментирована, но сохранялся зимний волос. Возможно, он как раз менял зимний мех сразу на летний. Отмечая довольно поздние сроки, С. Боровский отнес, тем не менее, этот экземпляр к группе с возобновившейся первой весенней линькой.
Во время весенней линьки II мездра очищается в первую очередь на холке. Когда спинная сторона перелиняет, начинают линять голова, бока и нижняя сторона тела. В конце мая – начале июня часто можно видеть самцов, у которых на спине короткий летний мех резко отделяется от более длинного весеннего меха на остальных частях тела.
У средней бурозубки смена меха на боковых железах не нарушает общего хода второй весенней линьки. Для обыкновенной бурозубки С. Боровский (Borowski, 1968) указывает, что при второй весенней линьке пигмент возле боковых желез, так же как и в паховой области, исчезает в последнюю очередь.
Весенняя линька II заканчивается в первой декаде июня. Изредка в июне встречаются также зверьки с прерванной второй весенней линькой. Один зимовавший самец с прерванной линькой, у которого весенний мех сохранялся на крестце и между ушей, был отловлен крайне поздно – 15 июля.
В процессе весенних линек у самцов зимний мех, высота которого в среднем равна 7.3 мм, сначала сменяется более коротким весенним мехом высотой 6.1 мм, а затем летним высотой 3.7 мм (см. табл. 5). Густота летнего меха по сравнению с густотой зимнего также значительно уменьшается.
Весенняя линька у самок. У самок смена шерстного покрова от зимы к лету имеет некоторые особенности. Первая самка с признаками линьки была отловлена 14 марта 1974 г. Процесс начинается на лбу и, постепенно расширяясь, захватывает всю голову (рис. 3В). Линька в основном идет по волновому типу. Довольно часто она отмечается на горле и у передних ног с наружной стороны. В результате весенней линьки I мех меняется у самок только на передней части тела, и весенний мех к моменту второй весенней линьки занимает не более 30 % площади шкуры. На остальной части тела остается зимний мех. Полная смена меха происходит во время весенней линьки II (рис. 3Г). Начинается линька на спине и затем сразу охватывает всю площадь шкуры. Проходит она в очень сжатые сроки. В годы исследований весенняя линька II проходила в период от 6 по 14 мая – в течение одной недели. С 15 мая все зимовавшие самки имели летний мех, чистую мездру, и лишь у отдельных зверьков наблюдались остатки черного пигмента на спинной стороне тела, возможно, из-за более длительного роста волоса в этих местах.
Таким образом, самки имеют неполную весеннюю линьку, и зимний мех у них на большей части тела непосредственно сменяется летним. Высота зимнего меха у самок равна 7.3 мм, а летнего – 4.2 мм.
Весенняя линька у самок начинается позже, чем у самцов. Как известно, половое созревание самок весной отстает от такового самцов почти на месяц. Это также подтверждает наличие связи между линькой и генеративным состоянием этих животных. Сроки весенней линьки II у самок совпадают с начальными стадиями беременности. Летний мех у них образуется значительно раньше, чем у самцов. Указание на подобный ход смены шерстного покрова от зимы к лету у самок известно и для обыкновенной бурозубки (Crowcroft, 1957; Borowski, 1968; Skaren, 1973).
Такие особенности в смене меха весной у самок имеют, очевидно, определенное адаптивное значение, т. к. способствуют более экономному расходу энергии. В летний период, кроме того, зимовавшие самки имеют более длинный мех (см. табл. 5).
Феномен наличия двух весенних линек у землероек остается пока необъясненным. Очевидно, только тщательные исследования по энергетике землероек могут дать ответ на вопрос о том, какой выигрыш получает организм животного вследствие такой смены меха.
Старческая линька. С. Боровский (Borowski, 1963) установил, что перезимовавшие землеройки, сменившие зимний наряд на летний, не имеют нормальной осенней линьки. В летний период у них наблюдается местная линька, ограниченная отдельными участками и без определенного направления (рис. 3Д). Она состоит в росте новых летних коротких волос. В нашем материале имеются зверьки с таким типом линьки в течение всего летнего периода как среди самцов, так и среди самок. К осени число зверьков, имеющих старческую линьку, увеличивается, особенно это заметно со второй половины августа.
Старческая линька проходит, как правило, на дорсальной стороне тела и часто вокруг боковых желез (у самцов и у самок). В двух случаях наблюдалась волновая линька.
У некоторых зверьков мех был вытерт до лысин, однако линька не наблюдалась. У части индивидуумов старческая линька может появляться тотчас после завершения весенней (Borowski, 1963). Следовательно, старческая линька лишь отчасти может быть названа компенсирующей, которая способствует сохранению целостности шерстного покрова. Например, у бурозубки, имеющей на боку затянувшуюся рану, на месте раны и вокруг нее на мездре было широкое поле пигмента и наблюдался рост нового волоса.
У одной самки, отловленной 18 сентября 1977 г., закладка пигмента в шкуре наблюдалась по всей спинной поверхности и была сходна с нормальной осенней линькой сеголеток. Более поздние отловы мы не проводили, поэтому неизвестно, появляется ли у зимовавших зверьков после такой линьки зимний мех. При содержании обыкновенных бурозубок в неволе С. Боровский (Borowski, 1964) имел два случая, когда после осенней линьки у зимовавших зверьков вновь вырос длинный зимний мех. Однако он считает, что этому могли способствовать хорошие кормовые условия неволи. В естественных условиях зимовавшие бурозубки осенью не получали вновь зимнего меха и даже в феврале имели короткий летний волос (Borowski, 1963).
Динамика численности
В условиях изучаемого региона численность средней бурозубки заметно изменяется по годам (с амплитудой от 0 до 1.3 на 100 лов.-сут. и от 0 до 8.1 на 10 кан.-сут.), причем без сколько-нибудь четкой периодичности и в целом не синхронно с таковой фонового вида (рис. 4). Если же отвлечься от частностей, то за 60 лет наблюдений вырисовывается всего четыре довольно нечетких, размытых периодов подъема. Первый – с 1959 по 1962, второй – с 1969 по 1975 гг., третий, чуть более низкий, – с 1985 по 1992, и четвертый – с 1999 по 2007 г.
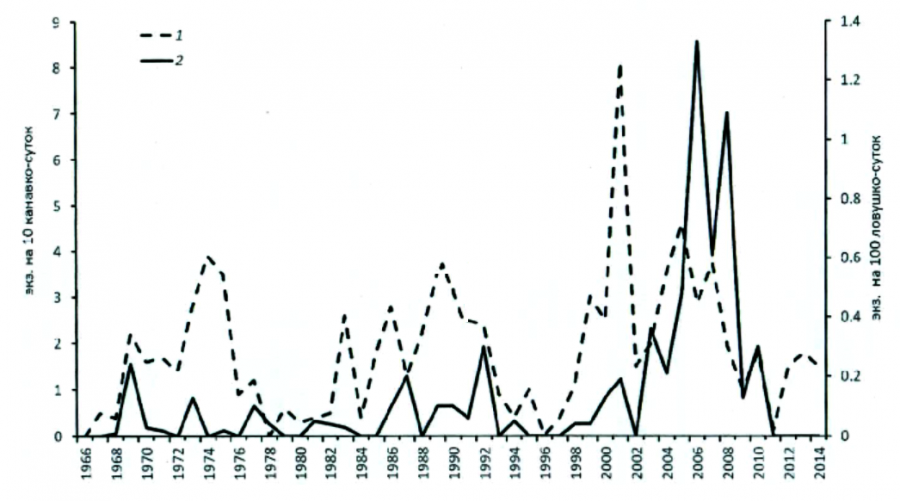
Рис. 4. Многолетняя динамика численности средней бурозубки по учетам в Карельском Приладожье: 1 – ловушки; 2 – канавки
Fig. 4. Long-term dynamics of the abundance of the common shrew according to censuses in the Karelian Ladoga region: 1 – traps; 2 – grooves
Соответственно, под категорию депрессионных попадают 1960–1968, 1976–1984 и 1993–1998, 2009–2014 гг. При этом никакой достоверной цикличности в смене подъемов и спадов численности ни визуально (см. рис. 1), ни с помощью статистического анализа данных учетов (Ивантер и др., 2008) выявить не удается. Следует помнить, что в Карелии, как и в целом в европейской части ареала, средняя бурозубка является видом подчиненным, тогда как в Сибири она – повсеместно господствующий вид. Это относится и к рис. 5, построенному по данным учетов в «Киваче», Каскеснаволоке и Костомукше.
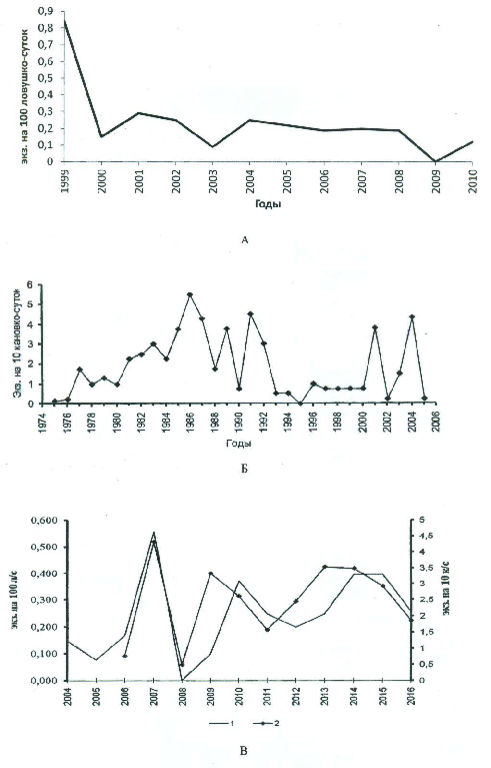
Рис. 5. Изменения численности средней бурозубки по данным учетов ловушко-линиями в Костомукшском заповеднике (А), заповеднике «Кивач» (Б), стационаре Каскеснаволок (В; 1 – линии, 2 – канавки)
Fig. 5. Changes in the abundance of the common shrew according to trap-line surveys in the Kostomuksha Reserve (A), the Kivach Reserve (Б), and the Kaskesnavolok station (В; 1 – lines, 2 – grooves)
Еще сложнее обстоит дело с выявлением причин многолетних изменений численности. Что касается Сибири, где, как уже говорилось, средняя бурозубка – бессмененый доминант (как, например, обыкновенная в Европе), то здесь ее численность, отражая действие двух противоположных процессов, размножения и смертности, изменяется под влиянием сложного сочетания целого ряда экзогенных и эндогенных факторов: зимние экологические условия, обильные осадки в бесснежный период, фенология, ход и сроки наступления весны, обеспеченность зверьков кормами, конкурентные отношения с господствующим видом и, наконец, численность популяции в предшествующий период времени (Докучаев, 1990). В Европе же, где средняя бурозубка – второстепенный вид, на первый план должны были бы выйти его конкурентные отношения с господствующим видом землероек. Однако, по крайней мере в нашем регионе, мы, как ни странно, вместо ожидаемой обратной корреляции между численностью, наблюдаем вполне значимую прямую зависимость (Ивантер и др., 2008). Отсюда вывод, что причиной многолетних колебаний численности данного вида на таежном Северо-Западе России, как, впрочем, и в Сибири, скорее всего, служат климатические (и прежде всего метеоролого-фенологические) факторы.
Что же касается сезонных изменений численности средней бурозубки, то, как и у других видов землероек-бурозубок, они несколько различаются по срокам и темпам в разные годы и в разных местообитаниях в зависимости от численности и состава населения, состояния, интенсивности размножения и уровня смертности популяции (Ивантер, Макаров, 2001). Вместе с тем общая схема процесса сохраняется: численность зверьков заметно нарастает от мая к июлю, достигает максимума в августе – сентябре, резко снижается к ноябрю – декабрю, стабилизируется зимой и вновь сокращается в апреле – мае (годовой минимум).
Заключение
Как показали многолетние экспедиционные и стационарные исследования, выявленные выше экологические особенности средней бурозубки в специфических условиях европейской, представляющей периферийную северо-западную часть ее ареала, территории отличаются крайней динамичностью и в значительной степени связаны как с плотностью популяции, так и с внешними, экологическими факторами среды. Весьма эффективные структурно-популяционные адаптации, затрагивающие все сферы жизнедеятельности вида, обеспечивают относительную устойчивость и оптимальное состояние его популяций в экстремальных для них приграничных условиях. Наряду с более частными специальными приспособлениями индивидуального ранга в периферических его населениях успешно реализуются широкие структурно-популяционные адаптивные комплексы, отличающиеся значительной лабильностью и высокой скоростью компенсаторной перестройки. В характере биотопического и пространственного размещения это означает переход к отчетливому синантропизму, а в репродукции – к удлинению периода размножения и достижению максимально эффективного воспроизводства за счет более полного и интенсивного участия в нем взрослых зверьков. Для процессов линьки характерна ее крайняя растянутость и максимальная мобилизация механизмов физической терморегуляции, проявляющаяся в крайнем усложнении структуры волосяного покрова. Вызванная же ужесточением отбора и специфической перестройкой возрастного и полового состава популяции активизация микроэволюционных процессов и проявляющиеся в этих условиях характерные адаптивные особенности динамики численности обеспечивают виду успешное продвижение на северо-запад и освоение им широких, далеко простирающихся от центра ареала новых территорий.
Библиография
Бигман В. В., Кирсанов И. К., Церевитинов Б. Ф. Товароведение животного сырья . М.: Центросоюз, 1948. 538 с.
Бобрецов А. В. Насекомояные // Бобрецов А. В. и др. Млекопитающие Печоро-Илычского заповедника . Сыктывкар: Коми кн. изд-во, 2004. С. 206–301.
Большаков В. Н., Васильев А. Г., Шарова Л. П. Фауна и популяционная экология землероек Урала (Mammalia, Sorcidae) . Екатеринбург, 1996. 268 с.
Воронов Г. А. География мелких млекопитающих южной тайги Приуралья, Средней Сибири и Дальнего Востока (антропогенная динамика фауны и населения) . Пермь: Изд-во Пермск. ун-та, 1993. 233 с.
Глотов И. Н., Ердаков Л. Н., Кузякин В. А. и др. Сообщества мелких млекопитающих Барабы . Новосибирск: Наука, 1978. 231 с.
Депарма Н. К. Линька кротов, ее последовательность и сроки // Труды Всесоюзного научно-исследовательского института охотничьего хозяйства. М., 1951. Вып. 10. С. 93–127.
Докучаев Н. Е. Механизмы, обеспечивающие восстановление численности бурозубок Северо-Восточной Сибири // Биологические проблемы Севера: Тезисы Х Всесоюзн. семинара. Магадан, 1983. Ч. II. C. 20–21.
Докучаев Н. Е. Экология бурозубок Северо-Восточной Азии . М.: Наука, 1990. 158 с.
Долгов В. А., Чабовский В. И., Шилова С. А. и др. Некоторые вопросы экологии бурозубок и их значение в очагах клещевого энцефалита // Бюллетень Московского общества испытателей природы. Отд. биол. 1968. Т. 73, вып. 6. С. 17–28.
Ивантер Э. В. К изучению динамики численности обыкновенной бурозубки (опыт аналитического обзора состояния проблемы) // Зоологический журнал. 2019. Т. 98, № 8. С. 949–959.
Ивантер Э. В. К изучению линьки волосяного покрова мелких лесных млекопитающих (опыт аналитического обзора состояния проблемы) // Успехи современной биологии. 2021. Т. 141, № 4. С. 399–416.
Ивантер Э. В. К изучению размножения обыкновенной бурозубки (Sorex araneus L.) // Зоологический журнал. 2020а. Т. 99, № 2. C. 189–202.
Ивантер Э. В. К экологии размножения малой бурозубки (Sorex minutus L.) на северной периферии ареала // Экология. 2020б. № 2. С. 141–148.
Ивантер Э. В. Очерки популяционной экологии мелких млекопитающих на северной периферии ареала . М.: Товарищество научных изданий КМК, 2018. 770 с.
Ивантер Э. В. Популяционная экология мелких млекопитающих таежного Северо-Запада СССР . Л.: Наука, 1975. 246 с.
Ивантер Э. В., Коросов А. В. Основы биометрии. Введение в статистический анализ биологических явлений и процессов . Петрозаводск: Изд-во ПетрГУ, 1992. 164 с.
Ивантер Э. В., Коросов А. В. Введение в количественную биологию . Петрозаводск: Изд-во ПетрГУ, 2011. 304 с.
Ивантер Э. В., Макаров А. М. Территориальная экология землероек-бурозубок (Sorex, Insectivora) . Петрозаводск: Изд-во ПетрГУ, 2001. 272 с.
Ивантер Э. В., Ивантер Т. В., Туманов И. Л. Адаптивные особенности мелких млекопитающих: Эколого-морфологические и физиологические аспекты . Л.: Наука, 1985. 318 с.
Ивантер Э. В., Коросов А. В., Якимова А. Е. Статистический анализ многолетних изменений численности мелких млекопитающих Северо-Восточного Приладожья // Труды Петрозаводского государственного университета. Серия: Биология: Вопросы популяционной экологии. Петрозаводск: Изд-во ПетрГУ, 2008. С. 5–24.
Карасева Е. В., Телицына А. Ю., Жигальский О. А. Методы изучения грызунов в полевых условиях . М.: Изд-во ЛКИ, 2008. 418 с.
Когтева Е. З. Сезонная изменчивость и возрастные особенности строения кожи и волосяного покрова крота, зайца-беляка и енотовидной собаки // Промысловая фауна и охотничье хозяйство Северо-Запада РСФСР. Л., 1963. С. 213–371.
Кузнецов Б. А. Товароведение пушномехового сырья СССР . М.: Сов. Азия, 1932. 464 с.
Куприянова И. Ф. Сем. Soricidae – землеройковые // Фауна Европейского северо-востока России. СПб.: Наука, 1994. Т. 2, ч. 1. С. 11–67.
Матурова Р. Т. Мелкие млекопитающие хребта Улан-Бургасы (Восточное Забайкалье) . Новосибирск: Наука. Сиб. отд-ние, 1982. 101 с.
Попов В. А. Млекопитающие Волжско-Камского края . Казань, 1960. 466 с.
Рамазанова Ф. Р. Размножение средней бурозубки в тайге заповедника «Малая Сосьва» // Исследования актуальных проблем териологии. Свердловск, 1983. С. 72–74.
Ревин Ю. В. Млекопитающие Южной Якутии . Новосибирск: Наука. Сиб. отд-ние, 1989. 321 с.
Ревин Ю. В., Сафронов В. М., Вольперт Я. Л. и др. Экология и динамика численности млекопитающих Предверхоянья . Новосибирск: Наука. Сиб. отд-ние, 1988. 200 с.
Реймерс Н. Ф. Птицы и млекопитающие южной тайги Средней Сибири . М.; Л.: Наука, 1966. 419 с.
Реймерс Н. Ф., Воронов Г. А. Насекомоядные и грызуны Верхней Лены . Иркутск, 1963. 190 с.
Снигиревская Е. М. Материалы по биологии размножения и колебаниям численности землероек в Башкирском заповеднике // Труды Башкирского государственного заповедника. 1947. Вып. 1. С. 12–18.
Соколов В. Е. и др. Руководство по изучению кожного покрова млекопитающих . М.: Наука, 1986. 279 с.
Тавровский В. А., Егоров О. В., Кривошеев В. Г. и др. Млекопитающие Якутии . М.: Наука, 1971. 660 с.
Церевитинов Б. Ф. Топографические особенности волосяного покрова пушных зверей // Труды ВНИИЖП. 1958. Вып. 17. С. 256–397.
Шварц С. С. Биология землероек лесостепного Зауралья // Зоологический журнал. 1955. Т. 34, вып. 4. С. 915–927.
Юдин Б. С. Экология бурозубок (род Sorex) Западной Сибири // Вопросы экологии, зоогеографии и систематики животных (Тр. Биол. ин-та СО АН СССР). Новосибирск, 1962. Вып. 8. С. 33–134.
Юдин Б. С., Барсова Л. И. Землеройки кедровых лесов Прителецкого очага клещевого энцефалита // Природа очагов клещевого энцефалита на Алтае. Новосибирск: Наука. Сиб. отд-ние, 1967. С. 51–59.
Юдин Б. С., Галкина Л. И., Потапкина А. Ф. Млекопитающие Алтае-Саянской горной страны . Новосибирск: Наука. Сиб. отд-ние, 1979. 293 с.
Юдин Б. С., Кривошеев В. Г., Беляев В. Г. Мелкие млекопитающие севера Дальнего Востока . Новосибирск: Наука. Сиб. отд-ние, 1976. 288 с.
Borowski S. Variation in density of coat during the cycle of araneus araneus L. // Acta theriol. 1958. Vol. 2, № 14. P. 286–289.
Borowski S. Old-age moult in the common shrew Sorex araneus Linnscus, 1758 // Acta theriol. 1963. Vol. 7, № 19. P. 374–385.
Borowski S. Moult of shrews (Sorex L.) under laboratory condition // Acta theriol. 1964. Vol. 8, № 8. P. 125–135.
Borowski S. Variations in coat and the common shrew // Acta theriol. 1968. Vol. 13, № 30. P. 483–498.
Borowski S. Variations in coat and colour in representatives of the genera Sorex L and Neomys Kaup // Acta theriol. 1973. Vol. 18, № 14. P. 247–279.
Crowcroft P. The life of the shrew. London, 1957. 166 p.
Dehnel A. Badania nad rodzgiem Sorex L. // Ann. Univer M. Curie-Sklodowska. Lublin, 1950. Sect. C. Vol. 5, № 2. S. 1–63.
Dehnel A. The biology of breeding of the Common Shrew Sorex araneus L. in laboratory condition // Ann. Univer M. Curie-Sklodowska. Lublin, 1952. Sect. C. Vol. 6, № 11. S. 359–376.
Gebczynski M., Olsewski J. Katathermometric measurements of insulating properities of the fur in small mammals // Acta theriol. 1963. Vol. 7, № 19. P. 369–371.
Stein G. H. Beziehungcn zwischen Bestandsdichte und Vermehrung bei der Waldspitzmaus Sorex araneus und weiteren Rotzahnspitzmauzen // Z. Saugetierkunde. 1961. Vol. 26, № 1. S. 143‒156.
Благодарности
Работа проводилась в плане выполнения Государственного задания Федерального исследовательского центра «Карельский научный центр Российской академии наук».

 © 2011 - 2026
© 2011 - 2026