



Введение
Обширная область исследований посвящена изучению морфофизиологических адаптаций черепах к существованию на границе водной, наземной и воздушной среды (Wyneken et al., 2007). В дельту Волги заходит восточная часть ареала вида с таким образом жизни – болотной черепахи Emys orbicularis (Linnaeus, 1758), представленной здесь популяциями гаплолинии I номинативного подвида E. o. orbicularis (Lenk et al., 1999; Дуйсебаева и др., 2019). В связи с нарастающими отклонениями сезонных температурных показателей и прессом антропогенной нагрузки особое внимание привлекает изучение эколого-физиологических механизмов адаптации черепах к новым формирующимся условиям, которые вызваны деградацией и загрязнением окружающей среды, болезнями или интродукцией инвазивных видов. Гематологические показатели являются индикаторами детерминантов окружающей среды (Alleman et al., 1992; Соколина и др., 1997; Вершинин, 2004; Павлов, 2019), а также ценными диагностическими инструментами при оценке состояния организма животных и изучении экофизиологического статуса организмов и популяций in situ (Davis et al., 2008; Perpinan, Sanchez, 2009). При этом весьма плодотворным методологическим подходом считается определение лейкоцитарной формулы крови с расчетом связанных с ней интегральных индексов. Ранее проведенными исследованиями в эритроцитах черепах E. orbicularis из природных популяций речных бассейнов Урала (Оренбургская область) и Волги (Астраханская область) обнаружены и идентифицированы гемопаразиты Haemogregarina spp. (Романова и др., 2021). В цикле работ, посвященных распространению и интенсивности инвазии водных черепах, выделяется два современных направления: во-первых, молекулярная идентификация (Ozvegy et al., 2015; Arizza et al., 2016) и морфология (Mihalca et al., 2002; Soares et al., 2014) гемопаразитов; во-вторых, иммуногематологический подход, позволяющий оценивать видоспецифические адаптивные реакции и состояние иммунной системы организма системы крови черепах при инвазии с учетом гендерных, возрастных и биотопических различий. Следует отметить, что в рамках этого направления имеющиеся сведения о характере количественных изменений в крови черепах при заражении гемогрегаринами носят фрагментарный характер и зачастую противоречивы (Mihalca et al., 2002; Salakij et al., 2002; Ozvegy et al., 2015; Arizza et al., 2016). Кроме того, полностью отсутствуют динамические исследования по сравнительному анализу популяционных иммуногематологических характеристик водных черепах, обеспечивающих важнейшие гомеостатические функции организма. В этой связи задача отслеживания изменений направленности адаптивных реакций системы крови болотной черепахи в условиях естественных биотопов при инвазии является актуальной и необходимой как для теории, так и для решения практических природоохранных вопросов в области экофизиологии рептилий.
С учетом вышеизложенного целью работы являлся сравнительный анализ зараженности гемогрегаринами и лейкоцитарного профиля крови болотной черепахи E. orbicularis из Астраханской области в период двухлетних наблюдений (2020–2021 гг.).
Материалы
Объект исследования – особи E. orbicularis, добытые в окрестностях пос. Комсомольский Красноярского района Астраханской области, в слабопроточном озере на правобережье р. Ахтуба. За два года было выловлено 108 особей: 48 особей (28 самок и 20 самцов) 27–29 августа 2020 г.; 60 особей (20 самок, 20 самцов и 20 молодых) 25–26 августа 2021 г. Проанализировано 204 гематологических препарата.
Работы проводили в соответствии с «Международными руководящим принципами для биомедицинских исследований на животных» (International Guiding…, 2012). Кровь у черепах брали путем прокола хвостовой вены. После взятия крови всех особей возвратили в места отлова.
Методы
Мазки крови (по два от каждой особи) готовили с помощью гематологического штапеля, высушивали на воздухе, затем фиксировали в спиртоэфирной смеси (1:1). Зафиксированные мазки хранили до окрашивания при комнатной температуре. Мазки окрашивали в течение 20 минут красителем Гимза азур-эозин по Романовскому (10–12-кратный раствор, фирма «Биолот», Россия), приготовленном на фосфатном буфере (рН 6.8–7.2). Готовые мазки просматривали на микроскопе «Meiji Techno», Япония, серия МТ 4000 с иммерсией, при увеличении ×1500, с дифференцированным подсчетом лейкоцитов (Alleman et al., 1992; Соколина и др., 1997). По морфологическим особенностям определяли пять типов лейкоцитарных клеток (в %): гранулоциты (гетерофилы, базофилы, эозинофилы) и агранулоциты (моноциты, лимфоциты). Цифровые изображения препаратов выполняли камерой «Vision САМ» для тринокулярного микроскопа «Meiju Techno» (увеличение ×1500).
На основании лейкоцитарной формулы крови рассчитали интегральные лейкоцитарные индексы в относительных единицах:
1) индекс сдвига лейкоцитов, ИСЛ = ∑ гранулоцитов / ∑ агранулоцитов;
2) индекс соотношения лимфоцитов и эозинофилов, ИСЛЭ = Л / Э;
3) индекс соотношения гетерофилов и эозинофилов, ИСГЭ = Г / Э;
4) лимфоцитарно-гранулоцитарный индекс, ИЛГ = Л × 10 / (Э + Г + Б);
5) индекс соотношения гетерофилов и лимфоцитов, ИСГЛ = Г / Л,
где Л – лимфоциты; Э – эозинофилы; Г – гетерофилы; Б – базофилы.
Пол взрослых черепах определяли по внешним морфологическим признакам, у молодых особей половую принадлежность не устанавливали. Измеряли длину карапакса (в мм) черепах по его прямой средней линии от переднего края загривкового щитка до заднего конца шва между надхвостовыми щитками штангенциркулем.
Учет гемопаразитов проводили на 500 эритроцитов у каждой особи. Экстенсивность инвазии (Е – количество особей вида, зараженных гемопаразитами, по отношению ко всему числу исследованных особей) рассчитывали по формуле: E = (n / N) × 100 %. Среднюю интенсивность инвазии (I – количество паразитов, приходящихся в среднем на одну черепаху) рассчитывали по формуле: I = m / n. Индекс обилия (ИО – количество гемопаразитов, приходящихся на одну исследованную особь) рассчитывали по формуле: ИО = m / N, где n – количество зараженных особей; m – количество обнаруженных гемопаразитов; N – количество исследованных особей хозяев.
Окуляр-микрометром проводили измерение диаметра лейкоцитарных клеток (2r), большой (2a) и малой (2b) осей здоровых и инфицированных эритроцитов. Площадь (мкм2) клеток рассчитывали по формулам круга S = π × r2 или эллипса: S = π × a × b, где r – диаметр круга; а – большая полуось эллипса; b – малая полуось эллипса; π = 3.14.
Полученные первичные данные проверяли на соответствие нормальному распределению по специальным критериям Шапиро – Уилка, Лиллиефорса и Андерсона – Дарлинга. Ввиду несоответствия первичных данных нормальному распределению, центральные тенденции и рассеяние показателей описывали медианой (Me) и интерквартильным размахом (IQR). Многофакторным дисперсионным анализом с использованием лямбды Уилкса (λWilks) в качестве критериальной статистики оценивали влияние факторов (пола, возраста, зараженности, года взятия) на лейкоцитарные показатели; значимость отличий (p-значения) рассчитывали путем рандомизации с использованием перестановочной процедуры (Шитиков, Розенберг, 2014; Якимов, 2019). Данные сравнивали методами непараметрической статистики с расчетом критериев: Краскела – Уоллиса (H) (при множественном сравнении независимых групп по одному признаку), Данна (D) (множественный критерий при попарном сравнении групп), Манна – Уитни (u) при сравнении двух групп, Вилкоксона (w) при сравнении зависимых выборок, критерием z – при сравнении долей с поправкой Холма, при α = 0.007, ранговым коэффициентом корреляции Спирмена (ρ) в пакете прикладных программ «Rstudio». За величину статистической значимости принимали α = 0.05.
Результаты
Средняя длина карапакса половозрелых самок болотной черепахи составляла 137.25 ± 5.31 мм, самцов 155.00 ± 1.92 мм. Половых различий по этому показателю у взрослых болотных черепах не выявлено (D = 2.07, p = 0.07). У сеголеток он равен 90.50 ± 0.90 мм, что значимо меньше по сравнению с самками (D = 6.10, p < 0.001) и самцами (D = 8.18, p < 0.001). Самцы и самки болотной черепахи, добытые в разные годы, по средней длине карапакса не различались (самки: w = 0.27, p = 0.77; самцы w = 0.57, p = 0.56).
При дифференциации округлых лейкоцитарных клеток болотной черепахи с учетом расположения, наличия зернистости в цитоплазме и формы ядра окуляр-микрометром измерили диаметр и рассчитали площадь клеток. Лейкоцитарные клетки самцов, за исключением эозинофилов, были крупнее аналогичных клеток самок и молодых особей. Самыми крупными лейкоцитами у всех особей являлись гетерофилы, самыми мелкими – лимфоциты (табл. 1).
Таблица 1. Морфометрия клеток лейкоцитарного ряда крови Emys orbicularis изученной популяции в 2021 г.
| Показатели лейкограммы | 1. Самцы половозрелые, n = 20 | 2. Самки половозрелые, n = 20 |
3. Сеголетки, n = 20 |
Статистические показатели | ||||
| Me | IQR | Me | IQR | Me | IQR | |||
| Гетерофилы | d | 16 | 2 | 15 | 3 | 14 | 2 | Н = 42.298, р < 0.001 D1-2 = 4.725, р < 0.001 D1-3 = 6.233, р < 0.001 |
| S | 201.06 | 50.27 | 176.71 | 68.33 | 153.94 | 43.98 | ||
| Эозинофилы | d | 14 | 3 | 13 | 2.25 | 13 | 3 | Н = 3.970, р = 0.137 |
| S | 153.94 | 63.62 | 132.73 | 46.53 | 132.73 | 63.62 | ||
| Базофилы | d | 13 | 2 | 12 | 2 | 12 | 1 | Н = 25.890, p < 0.001 D1-2 = 3.404, p = 0.001 D1-3 = 4.977, p < 0.001 |
| S | 132.73 | 40.84 | 113.1 | 37.7 | 113.1 | 18.06 | ||
| Моноциты | d | 13 | 3 | 12 | 2 | 12 | 2 | Н = 37.811, p < 0.001 D1-2 = 5.477, p < 0.001 D1-3 = 5.160, p < 0.001 |
| S | 132.73 | 63.62 | 113.1 | 37.7 | 113.1 | 37.7 | ||
| Лимфоциты | d | 10 | 1 | 9 | 3 | 9 | 1 | Н = 25.332, p < 0.001 D1-2 = 3.723, p < 0.001 D1-3 = 4.795, p < 0.001 |
| S | 78.54 | 14.92 | 63.62 | 31.22 | 63.62 | 13.35 | ||
Примечание. d – диаметр клетки, мкм; S – площадь клетки, мкм2; H – критерий Краскела – Уоллиса; D – критерий Данна; p – достигнутый уровень значимости.
Значимо не различались параметры базофилов и моноцитов у самцов (D = 0.62, p = 0.78), самок (D = 1.76, p = 0.13) и молодых (D = 0.89, p = 0.55), а также диаметры и площади эозинофилов и моноцитов (D = 0.83, p = 0.59) в выборке самок.
В период начальных наблюдений (2020 г.) в лейкоцитарном профиле крови половозрелых болотных черепах значимые половые различия обнаружены по содержанию моноцитов, доля которых была выше в крови самцов (Романова и др., 2021). Неполовозрелых особей в этот период не анализировали.
Анализ лейкоцитарного состава крови в 2021 г. показал значимые половые различия по содержанию гранулоцитов: эозинофилов (H = 28.58, p < 0.001) и базофилов (H = 8.30, p = 0.01). Доля эозинофилов в крови самцов была значимо выше по сравнению с самками (D = 2.85, p = 0.01) и молодыми (D = 5.32, p < 0.001), доля базофилов меньше по сравнению с молодыми (D = 2.75, p = 0.01). Различие в количественном содержании эозинофилов в крови особей болотной черепахи разного пола и возраста отразилось на интегральных лейкоцитарных показателях. Так, у неполовозрелых особей с низкой долей эозинофилов в крови индекс соотношения гетерофилов к эозинофилам (ИСГЭ) был выше по сравнению с самками (D = 2.92, p = 0.01) и самцами (D = 5.17, p < 0.001), а индекс соотношения лимфоцитов к эозинофилам (ИСЛЭ) выше по сравнению с самцами (D = 5.10, p < 0.001).
В динамике двухлетних наблюдений в лейкоцитарных профилях самцов болотной черепахи отмечено повышение относительного содержания гетерофилов и снижение доли моноцитов (табл. 2).
Таблица 2. Изменение лейкоцитарного профиля самцов Emys orbicularis в течение 2020–2021 гг.
| Показатели лейкограммы | Самцы 2020, n = 20 | Самцы 2021, n = 20 | Статистические показатели | |||
| Me | IQR | Me | IQR | Критерий Вилкоксона (w) | Уровень значимости (р) | |
| Гетерофилы, % | 11.50 | 9.50 | 15.00 | 13.00 | 2.56 | 0.01 |
| Эозинофилы, % | 12.00 | 7.50 | 13.00 | 11.50 | 1.48 | 0.13 |
| Базофилы, % | 21.50 | 13.50 | 20.50 | 7.50 | 0.63 | 0.52 |
| Моноциты, % | 10.00 | 8.00 | 6.00 | 3.00 | 4.70 | 0.000003 |
| Лимфоциты, % | 42.00 | 8.50 | 41.00 | 7.00 | 0.64 | 0.52 |
| ИСЛ, отн. ед. | 0.86 | 0.33 | 1.10 | 0.41 | 3.09 | 0.001 |
| ИСЛЭ, отн. ед. | 3.41 | 3.45 | 3.11 | 3.06 | 0.81 | 0.41 |
| ИСГЭ, отн. ед. | 1.15 | 0.91 | 1.08 | 1.38 | 0.90 | 0.36 |
| ИЛГ, отн. ед. | 9.29 | 3.38 | 6.54 | 3.03 | 2.28 | 0.02 |
| ИСГЛ, отн. ед. | 0.26 | 0.23 | 0.22 | 0.45 | 2.50 | 0.01 |
Примечание. Жирным выделены статистически значимые показатели.
В анализируемый период времени в крови самок обнаружено пониженное содержание моноцитов (табл. 3).
Таблица 3. Изменение лейкоцитарного профиля самок Emys orbicularis в течение 2020–2021 гг.
| Показатели лейкограммы | Самки 2020, n = 28 | Самки 2021, n = 20 | Статистические показатели | |||
| Me | IQR | Me | IQR | Критерий Вилкоксона (w) | Уровень значимости (р) | |
| Гетерофилы, % | 12.00 | 10.00 | 17.00 | 7.50 | 1.89 | 0.05 |
| Эозинофилы, % | 10.50 | 10.00 | 8.00 | 9.50 | 0.73 | 0.46 |
| Базофилы, % | 23.50 | 10.00 | 24.00 | 10.00 | 1.86 | 0.06 |
| Моноциты, % | 7.50 | 3.00 | 4.00 | 3.00 | 3.24 | 0.001 |
| Лимфоциты, % | 45.00 | 11.00 | 38.00 | 9.50 | 1.08 | 0.27 |
| ИСЛ, отн. ед. | 0.88 | 0.37 | 1.11 | 0.31 | 2.27 | 0.02 |
| ИСЛЭ, отн. ед. | 4.15 | 5.40 | 4.90 | 6.24 | 0.04 | 0.96 |
| ИСГЭ, отн. ед. | 1.27 | 1.50 | 1.70 | 2.22 | 1.12 | 0.25 |
| ИЛГ, отн. ед. | 9.36 | 4.53 | 8.03 | 2.81 | 1.81 | 0.06 |
| ИСГЛ, отн. ед. | 0.24 | 0.25 | 0.38 | 0.27 | 1.47 | 0.13 |
Примечание. Жирным выделены статистически значимые показатели.
Интегральные индексы реактивности, дополняя друг друга, свидетельствовали о значимых изменениях в состоянии организма болотных черепах в наблюдаемый период времени, обусловленных комплексом внешних и внутренних экологических факторов, среди которых особая роль отводится гемопаразитам. Хорошо известно, что гемопротозойные одноклеточные рода Haemogregarina (Coccidia: Adeleorina: Haemogregarinidae) паразитируют в основном в организме эктотермных позвоночных, в т. ч. и черепах (Данилевский, 1891; Финкельштейн, 1908; Мкртчян, 1966; Danilewsky, 1885; Segade et al., 2006; Zelmer, Platt, 2008; Adl et al., 2012; Dvorakova et al., 2014, 2015).
Проблема систематики гемогрегарин не является предметом настоящей работы, но следует отметить, что морфология, внешний вид и форма гемогрегариновых паразитов (микрогаметоцитов, макрогаметоцитов и трофозоитов, расположенных внутри эритроцитов зараженных особей) позволила отнести их к роду Haemogregarina из семейства Haemogregarinidae (Mihalca et al., 2002; Telford, 2009; Soares et al., 2014; Arizza et al., 2016). Расположенные в эритроцитах черепахи микрогаметоциты имели бобовидную форму и большое центральное ядро (рисунок, а), макрогаметоциты имели овальную форму с зернистыми ядрами и располагались вблизи ядра эритроцита (рисунок, б). Зрелые трофозоиты U-образной формы смещали ядро эритроцитов в полярное положение и превышали его размеры (рисунок, в).
а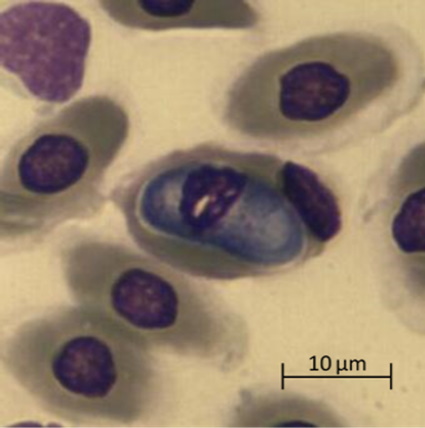
б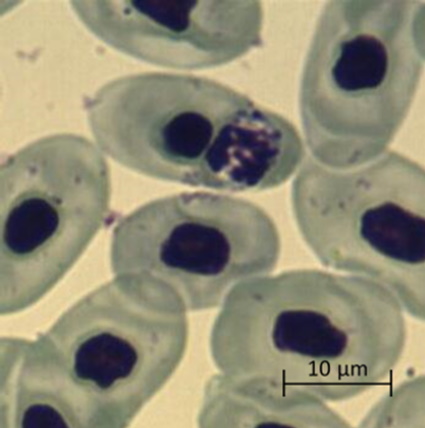
в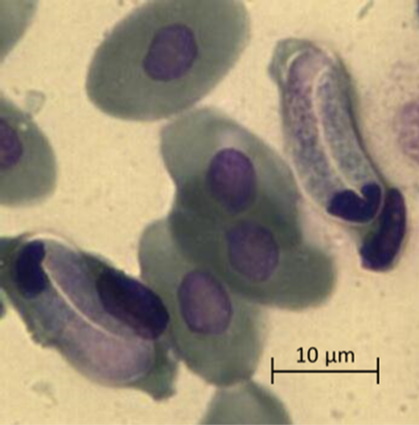
Микрофотографии паразитарных форм Haemogregarina spp., выполненные камерой Vision CAM для тринокулярного микроскопа Meiji Techno (×1500): а – макрогаметоциты; б – микрогаметоциты; в – трофозоиты
Micrographs of parasitic forms of Haemogregarina spp. made by the Vision CAM camera for the Meiji Techno trinocular microscope (×1500): a – macrogametocytes; б – microgametocytes; в – trophozoites
Многомерный дисперсионный анализ лейкоцитарного состава крови здоровых и инфицированных половозрелых самцов и самок, добытых в Астраханской области за два года (табл. 4), выявил значимое влияние таких факторов, как: пол (λWilks = 0.879, p = 0.005), год добычи (λWilks = 0.685, p = 0.002), и их взаимодействия (λWilks <0.001, p = 0.001). Эффект фактора инфицированности гемопаразитами оказался не значимым (λWilks = 0.981, p = 0.731).
Таблица 4. Лейкоцитарный состав крови здоровых и инфицированных половозрелых самцов и самок болотной черепахи разных лет взятия (2020–2021 гг.)
| Показатели лейкограммы | Самцы | Самки | ||||||
| Здоровые | Инфицированные | Здоровые | Инфицированные | |||||
| Me | IQR | Me | IQR | Me | IQR | Me | IQR | |
| 2020 год | ||||||||
| Гетерофилы, % | 6.00 | 7.00 | 12.00 | 8.00 | 9.00 | 4.00 | 13.50 | 11.00 |
| Эозинофилы, % | 12.00 | 6.00 | 12.00 | 9.00 | 12.00 | 10.00 | 12.00 | 7.50 |
| Базофилы, % | 28.00 | 12.00 | 21.00 | 12.00 | 25.00 | 8.00 | 20.00 | 10.00 |
| Моноциты, % | 12.00 | 7.00 | 10.00 | 7.00 | 7.00 | 2.00 | 9.70 | 4.50 |
| Лимфоциты, % | 44.00 | 4.00 | 42.00 | 9.00 | 47.00 | 8.00 | 42.00 | 11.50 |
| Гранулоциты, % от лейкоцитов | 49.00 | 11.00 | 46.00 | 9.00 | 47.00 | 10.00 | 48.50 | 11.50 |
| Агранулоциты, % от лейкоцитов | 51.00 | 11.00 | 54.00 | 9.00 | 53.00 | 10.00 | 51.50 | 11.00 |
| 2021 год | ||||||||
| Гетерофилы, % | 22.00 | 8.00 | 15.00 | 13.00 | 22.50 | 4.50 | 16.50 | 7.00 |
| Эозинофилы, % | 11.00 | 4.00 | 13.50 | 13.00 | 17.00 | 22.00 | 8.00 | 8.50 |
| Базофилы, % | 27.00 | 8.00 | 20.00 | 7.00 | 20.00 | 12.50 | 24.00 | 9.50 |
| Моноциты, % | 2.50 | 3.00 | 6.00 | 3.00 | 4.50 | 4.00 | 6.00 | 3.00 |
| Лимфоциты, % | 37.50 | 1.00 | 41.50 | 8.00 | 34.00 | 2.00 | 44.50 | 9.00 |
| Гранулоциты, % от лейкоцитов | 60.00 | 4.00 | 52.00 | 8.00 | 60.00 | 5.00 | 52.00 | 6.50 |
| Агранулоциты, % от лейкоцитов | 40.00 | 4.00 | 48.00 | 8.00 | 40.00 | 5.00 | 48.50 | 6.50 |
Результаты многомерного дисперсионного анализа лейкоцитарного состава крови половозрелых и молодых особей болотных черепах, добытых в 2021 г. (табл. 5), показали значимое влияние фактора возраста (λWilks = 0.855, p = 0.001), а также значимое взаимодействие факторов возраст – зараженность паразитами (λWilks < 0.001, p = 0.001).
Таблица 5. Лейкоцитарный состав периферической крови молодых и взрослых особей Emys orbicularis из Астраханской области, 2021 г.
| Показатель лейкограммы | Emys orbicularis | |||
| Половозрелые | Сеголетки | |||
| Me | IQR | Me | IQR | |
| Здоровые | ||||
| Гетерофилы, % | 22.50 | 6.00 | 18.50 | 9.50 |
| Эозинофилы, % | 11.00 | 20.00 | 5.00 | 3.50 |
| Базофилы, % | 23.00 | 14.00 | 25.00 | 4.50 |
| Моноциты, % | 4.00 | 3.00 | 6.00 | 5.00 |
| Лимфоциты, % | 35.50 | 3.00 | 45.00 | 10.50 |
| Гранулоциты, % от лейкоцитов | 60.33 | 4.00 | 49.50 | 10.00 |
| Агранулоциты, % от лейкоцитов | 40.00 | 4.00 | 50.50 | 10.00 |
| Инфицированные | ||||
| Гетерофилы, % | 15.00 | 10.00 | 20.50 | 6.00 |
| Эозинофилы, % | 10.50 | 11.00 | 6.50 | 5.50 |
| Базофилы, % | 22.00 | 10.00 | 23.50 | 10.50 |
| Моноциты, % | 6.00 | 3.00 | 5.50 | 4.50 |
| Лимфоциты, % | 42.00 | 10.00 | 42.00 | 6.50 |
| Гранулоциты, % от лейкоцитов | 52.00 | 8.00 | 51.50 | 7.00 |
| Агранулоциты, % от лейкоцитов | 48.00 | 8.00 | 48.50 | 7.00 |
В литературе имеются сведения о гипертрофии и изменении формы инфицированных гемогрегаринами эритроцитов черепах (Mihalca et al., 2002; Ozvegy et al., 2015; Arizza et al., 2016). Проведенный сравнительный морфометрический анализ выявил половые различия в параметрах инфицированных эритроцитов. Так, площадь эритроцитов без паразитов самцов статистически значимо превышала аналогичный показатель самок (z = 2.74, p = 0.003), но не изменялась по сравнению со здоровыми клетками (z = 1.42, p = 0.92). Пораженные эритроциты самок болотной черепахи имели большую площадь (z = 2.11, p = 0.02) и были слегка расширены (z = 4.15, p = 0.0001), в отличие от здоровых эритроцитов (табл. 6).
Таблица 6. Морфометрические параметры эритроцитов Emys orbicularis (Me / IQR)
| Пол черепах | Эритроциты крови | a, мкм | b, мкм | a / b | S, мкм2 |
| Самки | 1. Без паразитов, n = 100 | 20.00 / 2.00 | 12.00 / 1.00 | 1.73 / 0.32 | 188.50 / 33.97 |
| 2. С паразитами, n = 100 | 20.00 / 2.50 | 13.00 / 3.00 | 1.53 / 0.50 | 197.92 / 54.98 | |
| Самцы | 3. Без паразитов, n = 100 | 20.00 / 2.00 | 12.00 / 2.00 | 1.73 / 0.37 | 197.92 / 44.77 |
| 4. С паразитами, n = 100 | 20.00 / 2.00 | 12.00 / 2.00 | 1.70 / 0.51 | 193.99 / 36.91 | |
| Статистические показатели | H = 13.15, p < 0.001; D2-3 = 3.56, p = 0.003 |
H = 19.58, p < 0.001; D1-2 = 4.39, p < 0.001 |
H = 20.42, p < 0.001; D1-2 = 4.05, p < 0.001; D2-3 = 3.60, p = 0.001; D2-4 = 3.23, p < 0.001 |
H = 8.31, p = 0.04; D1-2 = 2.11, p = 0.02; D1-3 = 2.74, p = 0.003 |
|
| В целом по выборке | Без паразитов, n = 200 | 20.00 / 1.00 | 12.00 / 2.00 | 1.73 / 0.32 | 188.50 / 34.56 |
| С паразитами, n = 200 | 20.00 / 3.00 | 12.00 / 3.00 | 1.66 / 0.49 | 193.99 / 43.20 | |
| Статистические показатели: | u = 22283, p = 0.04 |
u = 16923, p = 0.007 |
u = 23610, p = 0.002 |
u = 19025, p = 0.40 |
|
Примечание. a – длина большой оси; b – длина малой оси; S – площадь клетки; n – количество просмотренных клеток; H – критерий Краскела – Уоллиса; D – критерий Данна; u – критерий Манна – Уитни.
Отметим, что в эритроцитах отдельной особи (как самцов, так и самок) встречались гемопаразиты на разных стадиях развития (например, макрогаметы и трофозоиты). Можно полагать, что морфометрические различия в параметрах инфицированных эритроцитов могут быть обусловлены не только половой принадлежностью, но и стадией развития гемогрегариновых паразитов или их количеством в клетке. Однако это предположение требует специального дополнительного объемного паразитологического анализа.
Показатели заражения болотных черепах Haemogregarina spp. в 2021 г. статистически значимо отличались от значений 2020 г. Межгодовые различия по суммарному популяционному показателю заражения гемогрегаринами свидетельствовали о повышении доли инфицированных эритроцитов (z = 9.84, р < 0.001) в крови болотных черепах. При этом существенно выросла доля зараженных клеток в крови самок (z = 9.11, р < 0.001) в отличие от самцов, у которых интенсивность инвазии и доля пораженных эритроцитов (z = 1.94, р = 0.1) оставались на прежнем уровне (табл. 7).
Таблица 7. Зараженность болотных черепах из Астраханской области гемогрегаринами
| Выборки | Число зараженных особей | Число паразитов на 500 эритроцитов | Доля клеток с паразитами | E, % | I, отн. ед. | ИО, отн. ед. |
| 2020 г. | ||||||
| Самки (n = 23) | 9 | 32 | 0.064 | 39.13 | 3.55 | 1.39 |
| Самцы (n = 19) | 14 | 82 | 0.164 | 73.68 | 5.85 | 4.31 |
| В целом по выборке (n = 42) | 23 | 114 | 0.228 | 54.76 | 4.95 | 2.71 |
| 2021 г. | ||||||
| Самки (n = 20) | 18 | 141 | 0.282 | 90 | 7.42 | 7.05 |
| Самцы (n = 20) | 19 | 106 | 0.212 | 95 | 5.88 | 5.3 |
| Молодые (n = 20) | 8 | 18 | 0.036 | 40 | 2.25 | 0.9 |
| В целом по выборке (n = 60) | 45 | 265 | 0.530 | 75 | 5.88 | 4.41 |
Более мелкие молодые особи отличались от самцов и самок более низкой зараженностью гемопаразитами. Проведенный анализ выявил значимое различие в количестве эритроцитов с паразитами у молодых по сравнению со взрослыми самками (z = 10.63, р < 0.001) и самцами (z = 8.44, р < 0.001). Отметим, что экстенсивность инвазии и доли эритроцитов с гемогрегаринами в крови самок и самцов в 2021 г. не различались (z = 2.56, р > 0.007).
Для определения направленности адаптивных реакций организма при инвазии был проведен сравнительный анализ индексов реактивности здоровых особей и инфицированных гемогрегаринами. У самок болотных черепах при инвазии гемопаразитами выявлено возрастание индексов ИСГЭ (D = 2.40, p = 0.04) и ИСГЛ (D = 2.58, p = 0.02) за счет изменения соотношения гранулоцитарных лейкоцитарных компонентов, что иллюстрировало существенные различия в адаптационных механизмах организма на уровне системы крови. У инвазионных самцов в динамике наблюдаемого периода наблюдалось возрастание ИСЛ (D = 2.78, p = 0.01), что свидетельствовало о преобладающей роли гранулоцитов в формировании адаптивного иммунного ответа организма (табл. 8).
Таблица 8. Значение лейкоцитарных индексов здоровых болотных черепах и при инвазии гемопаразитами
| Лейкоцитарный индекс, отн. ед. | 1. Без инвазии | При инвазии | Статистические показатели | ||||
| 2. 2020 г. | 3. 2021 г. | ||||||
| Me | IQR | Me | IQR | Me | IQR | ||
| Самки | |||||||
| ИСЛ | 0.88 | 0.34 | 0.94 | 0.45 | 1.08 | 0.28 | Н = 2.06,
р = 0.35 |
| ИСЛЭ | 4.41 | 6.40 | 4.19 | 3.92 | 5.14 | 6.02 | Н = 2.30,
р = 0.31 |
| ИСГЭ | 0.83 | 1.50 | 1.47 | 1.60 | 1.70 | 1.69 | Н = 6.04,
р = 0.04 D1-3 = 2.40, p = 0.04 |
| ИЛГ | 9.56 | 3.51 | 9.03 | 4.18 | 8.33 | 2.85 | Н = 1.56,
р = 0.45 |
| ИСГЛ | 0.21 | 0.26 | 0.45 | 0.30 | 0.35 | 0.21 | Н = 7.40,
р = 0.02 D1-3 = 2.58, p = 0.02 |
| Самцы | |||||||
| ИСЛ | 1.00 | 0.68 | 0.85 | 0.31 | 1.08 | 0.34 | Н = 8.99,
р = 0.01 D2-3 =2 .98, p = 0.008 |
| ИСЛЭ | 3.66 | 3.62 | 3.33 | 3.18 | 3.11 | 3.10 | Н = 1.62,
р = 0.44 |
| ИСГЭ | 1.20 | 1.58 | 1.22 | 1.08 | 1.05 | 1.33 | Н = 1.16,
р = 0.92 |
| ИЛГ | 8.57 | 5.06 | 9.50 | 3.27 | 8.00 | 3.25 | Н = 2.69,
р = 0.25 |
| ИСГЛ | 0.21 | 0.48 | 0.28 | 0.22 | 0.34 | 0.43 | Н = 3.61,
р = 0.16 |
| В целом по выборке | |||||||
| ИСЛ | 0.94 | 0.43 | 0.88 | 0.42 | 1.08 | 0.34 | Н = 8.87,
р = 0.01 D2-3 = 2.78, p = 0.01 |
| ИСЛЭ | 4.02 | 5.55 | 3.50 | 3.63 | 4.36 | 4.53 | Н = 0.70,
р = 0.70 |
| ИСГЭ | 0.95 | 1.35 | 1.25 | 1.50 | 1.40 | 1.75 | Н = 2.88,
р = 0.23 |
| ИЛГ | 9.19 | 4.58 | 9.09 | 3.84 | 9.07 | 2.94 | Н = 2.90,
р = 0.23 |
| ИСГЛ | 0.21 | 0.35 | 0.30 | 0.20 | 0.34 | 0.29 | Н = 9.69,
р = 0.007 D1-3 = 3.10, p = 0.005 |
Примечание. Жирным выделены статистически значимые показатели.
Значимые значения (р < 0.05) корреляционных коэффициентов для самок черепах иллюстрировали снижение суммарного содержания гранулоцитов (ρ = -0.46) за счет вклада гетерофилов (ρ = -0.32), повышение содержания агранулоцитов (ρ = 0.39) за счет вклада лимфоцитов (ρ = 0.35) (табл. 9). Для самцов и молоди болотной черепахи в отличие от самок корреляционной взаимосвязи гематологических показателей и наличия гемопаразитов не обнаружено.
Таблица 9. Анализ взаимосвязи интенсивности инвазии с лейкоцитарным профилем крови болотной черепахи коэффициентом ранговой корреляции Спирмена (ρ)
| Показатель | Самцы | Самки | Сеголетки | Вся выборка |
| Гетерофилы, % | 0.15 | -0.32 | 0.11 | -0.13 |
| Эозинофилы,% | 0.04 | -0.20 | 0.10 | 0.21 |
| Базофилы,% | -0.08 | 0.12 | -0.04 | -0.13 |
| Моноциты, % | 0.14 | 0.15 | 0.17 | 0.13 |
| Лимфоциты, % | -0.18 | 0.35 | -0.18 | -0.04 |
| ИСЛ, отн ед. | 0.14 | -0.43 | 0.12 | -0.03 |
| ИСЛЭ, отн ед. | -0.04 | 0.22 | -0.15 | -0.21 |
| ИСГЭ, отн ед. | 0.01 | 0.07 | -0.08 | -0.22 |
| ИЛГ, отн. ед. | -0.16 | 0.41 | -0.15 | -0.01 |
| ИСГЛ, отн. ед. | 0.18 | -0.33 | 0.12 | -0.09 |
Примечание. Жирным выделены статистически значимые показатели.
В целом по выборке выявлена положительная слабая взаимосвязь между показателями инвазии и содержанием в периферической крови эозинофилов (ρ = 0.21), а также отрицательная слабая взаимосвязь с интегральными лейкоцитарными индексами ИСЛЭ (ρ = -0.21) и ИСГЭ (ρ = -0.22).
Обсуждение
Сравнительный анализ данных лейкоцитарного профиля крови болотной черепахи выявил количественно-качественные изменения в иммуногематологических показателях, направленные на адаптивный ответ и обеспечивающие функционирование организма. При изучении динамики интегральных лейкоцитарных индексов реактивности организма обращает на себя внимание возрастание индекса ИСЛ у самцов и самок, что иллюстрировало снижение реагирования иммунитета в целом и доминирующей роли гранулоцитов в неспецифических адаптивных реакциях организма. Отметим, что у самцов, но не у самок, одновременно отмечено и понижение ИЛГ, что может свидетельствовать о недостаточном ресурсе реакций специфического иммунитета. При этом отсутствие возрастание индекса ИСГЛ характеризует достаточную реактивность активного ответа организма на комплекс факторов среды, в т. ч. с учетом инвазии.
Анализ динамики интегральных лейкоцитарных индексов болотных черепах исследованной выборки позволяет заключить, что при поражении гемогрегаринами в организме инфицированных особей происходит изменение соотношения лейкоцитарных клеток. По всей видимости, отставание в реагировании на внедрение гемопаразитов со стороны моноцитов (самцы и самки) приводит, с одной стороны, к запаздыванию фазы завершенного фагоцитоза, а с другой – к поздней активации лимфоцитов как эффекторного звена иммунного ответа организма хозяина, что обеспечивает более благоприятные условия для существования паразитарных форм Haemogregarina spp. Отметим, что ИЛГ находился в пределах нормальных значений у здоровых и инфицированных черепах. Этот индекс рассматривается как показатель сбалансированности реакции крови на заражение и на фоне незначительного повышения индекса ИСГЛ отражает влияние паразита, когда реактивность организма хозяина не выходит за пределы нормальных значений. Выявленная корреляционная взаимосвязь между показателями инвазии и возрастанием в крови количества эозинофилов подтверждает факт участия эозинофилов в иммунном ответе черепах против паразитарных форм Haemogregarina spp., что поддерживается и другими исследованиями (Mihalca et al., 2002).
Виды рода Haemogregarina spp. являются внутриэритроцитарными паразитами, имеют сложный жизненный цикл, который включает мерогонию и образование гаметоцитов в организме промежуточного хозяина (черепаха), а также гамогонию и спорогонию в кишечнике окончательного хозяина (пиявка, беспозвоночный переносчик) (Telford, 2009). Зараженные Haemogregarina spp. пиявки передают мерозоиты черепахам (Paperna, 1989; Siddall, Desser, 2001). Первичная мерогония происходит в легких, печени и селезенки промежуточного хозяина. Вторичная мерогония имеет место в эритроцитах, где формируются различные паразитарные стадии, в т. ч. и гамонты (Ozvegy et al., 2015).
Несмотря на существующее мнение о непатогенности гемогрегарин для черепах (Финкельштейн, 1908; Мкртчян, 1966; Mihalca et al., 2002; Perpinan, Sanchez, 2009) и их слабой связи с организмом хозяина (Данилевский, 1891; Soares et al., 2014), появляется все больше фактов, свидетельствующих о серьезных последствиях, происходящих в эритроцитах рептилий после внедрения в них паразитов, связанных с интенсификацией метаболизма и функциональной нагрузки эритроцита (Бейер, Сидоренко, 1972), изменением значения гематокрита, уменьшением числа эритроцитов и уровня гемоглобина (Thrall et al., 2004; Stacy et al., 2011; Ozvegy et al., 2015). Эритроциты являются постоянной составной частью микроокружения иммунокомпетентных клеток, следует отметить их роль в регуляции и реализации иммунного ответа. Способность эритроцитов сорбировать, концентрировать и модифицировать различные биологические активные вещества (антигены, медиаторы, гормоны) и оказывать воздействие на лимфопролиферацию, представляя их лимфоцитам, является важным звеном механизма регуляции иммуногенеза в естественных условиях, при стрессе и патологии (Васильев, 1975; Прокопенко, Сипливая, 1992; Кирдей, Дмитриева, 1995). Эритроциты рептилий вследствие их интенсивной метаболической активности служат прекрасным модельным объектом для изучения взаимоотношений паразита и хозяина на клеточном уровне (Бейер, Сидоренко, 1972). Можно полагать, что выявленные морфометрические параметры эритроцитов при инвазии, отражающие направленное изменение размеров клеток, по всей видимости, являются лишь внешней формой выражения происходящих в метаболизме клеток отклонений, направленных на обеспечение существования паразита в крови черепах до момента попадания в организм окончательного хозяина.
Заражение паразитами в популяции нарастает, о чем свидетельствуют суммарные популяционные показатели инвазии. Положительная умеренная корреляционная взаимосвязь между интенсивностью инвазии и длиной карапакса болотной черепахи (ρ = 0.36, р < 0.05) позволяет прогнозировать более высокую зараженность в процессе онтогенетического развития рептилий. На динамику передачи гемопаразитов на черепах, скорее всего, влияют сезонные колебания уровня воды в бассейне Нижней Волги. Перемещение и пребывание болотных черепах в районах затопления, по всей видимости, может увеличивать возможность заражения черепах через пиявок, чему способствует избыточное увлажнение и застойный водный режим (Sawyer, 1986).
Заключение
Проведенный сравнительный анализа адаптивных реакций системы крови и морфометрических показателей клеток крови болотной черепахи из Астраханской области показал, что пути адаптации на уровне системы крови к комплексу факторов среды, включая инвазию гемопаразитами, имеют существенные половые различия и временную динамику, иллюстрируя снижение реагирования иммунитета в целом и доминирующую роль гранулоцитов в адаптивных реакциях организма. Изучена зараженность болотных черепах гемопаразитами Haemogregarina spp. Показано возрастание суммарных популяционных показателей инвазии за двухлетний период, преимущественно за счет заражения самок. Высокая и нарастающая инвазия болотных черепах гемопаразитами свидетельствует о стабильном цикле передачи инфекции в Астраханской области. Для получения точных характеристик и особенностей жизненного цикла гемогрегарин в крови болотных черепах и ответа на вопрос о влиянии кровепаразитов на организм хозяина требуется проведение специального дополнительного паразитологического исследования и сравнительного экофизиологического анализа иммунного статуса инфицированных особей.
Библиография
Бейер Т. В., Сидоренко Н. В. Цитохимическое исследование гемогрегарин рептилий Армении. II. Сдвиги в содержании гемоглобина и общих белков в эритроцитах скальных ящериц, зараженных гемогрегаринами // Паразитология. 1972. Т. VI. Вып. 4. C. 385–390.
Васильев Н. В. Очерки о роли кроветворной ткани в антителообразовании . Томск: Изд-во Томского ун-та, 1975. 302 с.
Вершинин В. Л. Гемопоэз бесхвостых амфибий – специфика адаптациогенеза видов в современных экосистемах // Зоологический журнал. 2004. Т. 83, № 11. С. 1367–1374.
Данилевский В. Я. Исследования по сравнительной паразитологии крови. Зоопаразиты крови у пресмыкающихся . Т. 2. Харьков, 1891. 145 с.
Дуйсебаева Т. Н., Доронин И. В., Малахов Д. В., Кукушкин О. В., Бакиев А. Г. ГИС-анализ распространения и условий обитания Emys orbicularis orbicularis (Testudines, Emydidae): методические аспекты // Известия высших учебных заведений. Поволжский регион. Естественные науки. 2019. № 1. С. 28–40. DOI: 10.21685/2307-9150-2019-1-4.
Кирдей Е. Г., Дмитриева Л. А. Роль эритроцитов в регуляции и реализации иммунного ответа // Сибирский медицинский журнал. 1995. Т. 4, № 3. С. 5–8.
Мкртчян З. А. Материалы к изучению паразитов крови рептилий Армении // Биологический журнал Армении. 1966. Т. XIX, № 10. С. 92–100.
Павлов А. В. Ключевые моменты гематологии рептилий: особенности оценки лейкоцитарной части крови // Известия высших учебных заведений. Поволжский регион. Естественные науки. 2019. № 1. С. 138–152. DOI: 10.21685/2307-9150-2019-1-14.
Прокопенко Л. Г., Сипливая Л. Е. Эритроциты как модуляторы иммунологических реакций // Успехи физиологических наук. 1992. Т. 23, № 4. С. 89–106.
Романова Е. Б., Столярова И. А., Бакиев А. Г., Горелов Р. А. Сравнительный лейкоцитарный профиль Emys orbicularis (Reptilia: Emydidae) из двух популяций // Современная герпетология. 2021. Т. 21, № 1/2. С. 30–42. DOI: 10.18500/1814-6090-2021-21-1-2-30-42.
Соколина Ф. М., Павлов А. В., Юсупов Р. Х. Гематология пресмыкающихся: Методическое пособие по курсу герпетологии, большому практикуму и спецсеминарам . Казань: Казан. гос. ун-т, 1997. 31 с.
Финкельштейн Н. Я. Чужеядные крови холоднокровных Кавказа // Архив Биологических Наук. 1908. Т. 13. Вып. 2. С. 132–164.
Шитиков В. К., Розенберг Г. С. Рандомизация и бутстреп: статистический анализ в биологии и экологии с использованием R. . Тольятти: Кассандра, 2014. 314 с.
Якимов В. Н. Основы анализа биомедицинских и экологических данных в среде R. Часть 2: Учебное пособие. . Нижний Новгород: Нижегородский госуниверситет, 2019. 168 с.
Adl S. M., Simpson A. G. B., Heiss A., Lane C. E., Lukeš J., Bass D., Bowser S. S., Brown M. W., Burki F., Dunthorn M., Hampl V., Hoppenrath M., Lara E., Mitchell E. A. D., Gall L. L., Lynn D. H., Mcmanus H., Mozley-Stanridge S. E., Parfrey L. W., Pawlowski J., Rueckert S., Shadwick L., Spiegel F. W., Schoch C. L., Smirnov A. The revised classification of eukaryotes // Journal of Eukaryotic Microbiology. 2012. Vol. 59. Issue 5. P. 429–493. DOI: 10.1111/j.1550-7408.2012.00644.x.
Alleman A. R., Jacobson E. R., Raskin R. E. Morphologic and cytochemical characteristics of blood cells from the desert tortoise (Gopherus agassizii) // American Journal of Veterinary Research. 1992. Vol. 53. P. 1645–1651.
Arizza V., Sacco F., Russo D., Scardino R., Arculeo M., Vamberger M., Marrone F. The good, the bad and the ugly: Emys trinacris, Placobdella costata, and Haemogregarina stepanowi in Sicily (Testudines, Annelida, and Apicomplexa) // Folia Parasitologica. 2016. Vol. 63: 029. DOI: 10.14411/fp.2016.029.
Danilewsky B. Die Hämatozoën der Kaltblüter // Archiv für Mikroskopische Anatomie. 1885. Band 24. S. 588–598.
Davis A. K., Maney D. L., Maerz J. C. The use of leukocyte profiles to measure stress in vertebrates: a review for ecologists // Functional Ecology. 2008. Vol. 22. P. 760–767. DOI: 10.1111/j.1365-2435.2008.01467.x.
Dvořáková N., Kvičerová J., Hostovský M., Široký P. Haemogregarines of freshwater turtles from Southeast Asia with a description of Haemogregarina sacaliae sp. n. and a redescription of Haemogregarina pellegrini Laveran and Pettit, 1910 // Parasitology. 2015. Vol. 42. Issue 6. P. 816–826. DOI: 10.1017/S0031182014001930.
Dvořáková N., Kvičerová J., Papoušek I., Javanbakht H., Tiar G., Kami H., Široký P. Haemogregarines from western Palaearctic freshwater turtles (genera Emys, Mauremys) are conspecific with Haemogregarina stepanowi Danilewsky, 1885 // Parasitology. 2014. Vol. 141. Issue 4. P. 522–530. DOI: 10.1017/S0031182013001820.
International Guiding Principles for Biomedical Research Involving Animals. 2012. URL: https://grants.nih.gov/grants/olaw/guiding_principles_2012.pdf (дата обращения: 27.05.2022).
Lenk P., Fritz U., Joger U., Wink M. Mitochondrial phylogeography of the European pond turtle, Emys orbicularis (Linnaeus, 1758) // Molecular Ecology. 1999. Vol. 8. Issue 11. P. 1911–1922.
Mihalca A., Achelaritei D., Popescu P. Haemoparasites of the genus Haemogregarina in a population of European pond turtles (Emys orbicularis) from Drăgăşani, Vâlcea county, Romania // Scientia Parasitologica. 2002. Vol. 2. P. 22–27.
Özvegy J., Marinković D., Vučićević M., Gajić B., Stevanović J., Krnjaić D., Aleksić-Kovačević S. Cytological and molecular identification of Haemogregarina stepanowi in blood samples of the European pond turtle (Emys orbicularis) from quarantine at Belgrade Zoo // Acta Veterinaria-Beograd. 2015. Vol. 65, No 4. P. 443–453. DOI: 10.1515/acve-2015-0037.
Paperna I. Developmental cycle of chelonian haemogregarines in leeches with extra-intestinal multisporozoite oocysts and a note on the blood stages in the chelonian hosts // Diseases of Aquatic Organisms. 1989. Vol. 7. P. 149–153.
Perpiñán D., Sánchez C. Morphologic and Cytochemical Characteristics of Blood Cells from the European Pond Turtle (Emys orbicularis) and the Mediterranean Pond Turtle (Mauremys leprosa) // Journal of Herpetological Medicine and Surgery. 2009. Vol. 19. No 4. P. 119–127.
Salakij C., Salakij J., Suthunmapinunta P., Chanhome L. Hematology, Morphology and Ultrastructure of Blood Cells and Blood Parasites from Puff-faced Watersnakes (Homalopsis buccata) // Kasetsart Journal – Natural Science. 2002. Vol. 36. P. 35–43.
Sawyer R. T. Leech biology and behavior. Vol. 3. Oxford: Oxford University Press; Clarendon Press, 1986. 240 p.
Segade P., Crespo C., Ayres C., Cordero A., Arias M. C., García-Estévez J. M., Iglesias Blanco R. Eimeria species from the European pond turtle, Emys orbicularis (Reptilia: Testudines), in Galicia (NW Spain), with description of two new species // Journal of Parasitology. 2006. Vol. 92. Issue 1. P. 69–72. DOI: 10.1645/GE-3491.1.
Siddall M. E., Desser S. S. Transmission of Haemogregarina balli from painted turtles to snapping turtles through the leech Placobdella ornata // Journal of Parasitology. 2001. Vol. 87. Issue 5. P. 1217–1218. DOI: 10.1645/0022-3395(2001)0872.0.CO.
Soares P., de Brito E. S., Paiva F., Pavan D., Viana L.A. Haemogrerarina spp. in a wild population from Podocnemis unifilis Troschel, 1848 in the Brazilian Amazonia // Parasitology Research. 2014. Vol. 113. P. 4499–4503. DOI: 10.1007/s00436-014-4139-7.
Stacy N. I., Alleman A. R., Sayler K. A. Diagnosic hematology of reptiles // Clinics in Laboratory Medicine. 2011. Vol. 3. P. 87–108. DOI: 10.1016/j.cll.2010.10.006.
Telford Jr. S. R. Hemoparasites of the Reptilia: Color Atlas and Text. New York: CRC Press, 2009. 376 p.
Thrall M. A., Dale C., Baker E., Lassen E. D. Hematology of reptiles // Veterinary Hematology and Clinical Chemistry. Pennsylvania, USA: Lippincott Williams & Wilkins, 2004. P. 259–276.
Wyneken J., Bels V. L., Godfrey M. H. Biology of Turtles. Boca Raton: CRC Press, 2007. 408 p.
Zelmer D. A., Platt T. R. Structure and similarity of helminth communities of six species of Australian turtles // Journal of Parasitology. 2008. Vol. 94. Issue 4. P. 781–787. DOI: 10.1645/GE-1487.1.

 © 2011 - 2026
© 2011 - 2026