



Введение
При изучении экологии ушастых круглоголовок (Phrynocephalus mystaceus) на бархане Сарыкум в Республике Дагестан (РФ) авторы обнаружили, что их суточная активность не заканчивалась после того, как они вечером покидали поверхность почвы.
Исследователям, когда-либо изучавшим экологию ушастых круглоголовок, было известно, что эти ящерицы – типично дневные, активные исключительно в жаркое дневное время при высоких температурах внешней среды и тела. Поэтому совершенно невероятно даже предположение о том, что они могут быть активны вечером, после захода солнца. Тем не менее наши наблюдения показали, что на Сарыкуме эти дневные ящерицы встречались активными даже в плотных сумерках, иногда почти в полной темноте (Окштейн и др., 2021).
Задача настоящей работы – описание сумеречной активности типично дневного вида ящериц, выявление ее отличительных особенностей и постановка ряда экологических и физиологических вопросов, проявившихся в данной ситуации, ответы на которые еще только предстоит найти.
Материалы
Исследования экологии ушастых круглоголовок проводились нами на бархане Сарыкум в Республике Дагестан летом и осенью 2019 и 2021 гг. В течение этого времени было зарегистрировано более 80 эпизодов сумеречной активности ушастых круглоголовок.
Методы
При исследовании экологии и суточной активности ушастых круглоголовок мы регистрировали утреннее появление круглоголовок из ночных убежищ (из ночных нор или из поверхностного слоя песка с глубины 3–4 см), дневную активность, а затем подготовку норы для ночевки и уход в ночное убежище.
Наблюдения за поведением ушастых круглоголовок мы проводили на регулярных маршрутах, а также в процессе подробного изучения поведения этих ящериц на полигоне размером 100 х 40 м. В процессе работы мы производили индивидуальное мечение круглоголовок, нанося на их спины номера большого размера с помощью спиртового маркера черного цвета. Эти номера были легко заметны издали, позволяли идентифицировать особей и наблюдать за их индивидуальным поведением, перемещениями, причем не только на свету в дневное время, но и в глубоких сумерках, и даже в темноте в свете фонаря. Эти номера хорошо читались в течение всего сезона работы, а в отдельных случаях более или менее отчетливо были видны даже следующей весной.
Температуры тела ящериц измеряли с помощью электротермометра Mastech MS6512 фирмы Precision Mastech Enterprises. Температуры внешней среды (температуру воздуха на высоте 3 см над поверхностью почвы, температуру поверхности почвы, температуру в песке на глубине 5, 10 и 15 см) измеряли с помощью цифровых электротермометров DС-2 с выносными датчиками, размещенными на временной метеорологической площадке непосредственно в месте проведения работ. Методика термобиологических исследований описывалась нами ранее (Черлин, 2014 и др.).
В тексте часто используется термин «полная активность», который стоит определить. Полная активность – это тип активности, при котором рептилии находятся в физиологическом состоянии, в полной мере обеспечивающем эффективное протекание всех основных поведенческих актов (локомоция, терморегуляция, питание, сексуальность, территориальность, защита, коммуникации и т. п.). В этом состоянии движения рептилий активны, быстры (адекватно видовым особенностям) и точны (Черлин, 2014).
Все температуры в данной статье указываются в градусах Цельсия.
Результаты
В подавляющем большинстве географических точек, где живут ушастые круглоголовки, исследователи отмечали, что их дневная активность заканчивалась обычно примерно к 19–20 часам (Бадмаева и др., 1981; Банников и др., 1977; Богданов, 1960, 1962, 1965; Богданов, Сударев, 1989; Божанский, 2014; Джамирзоев и др., 2013; Доронин, 2013; Ждокова, 2013; Киреев, 1983; Литвинов, Югов, 2013; Лотиев, 2020; Мазанаева, 2009, 2020; Никольский, 1915; Параскив, 1956; Саид-Алиев, 1979; Тертышников, 2002; Тертышников, Горовая, 1984; Хонякина, 1962а, б, 1964; Черлин, Музыченко, 1983а; Шаммаков, 1981; Шнитников, 1928; Roitberg et al., 2000 и др.). К этому времени круглоголовки либо выкапывали ночную нору, либо тонули в песке, закапываясь в его поверхностный слой боковыми движениями тела. После этого исследователи отмечали их появление на поверхности только следующим утром.
На Сарыкуме мы отметили активность ушастых круглоголовок, которая отличалась от ожидаемой, многократно описанной другими авторами. Здесь, как и в других местах, круглоголовки вначале завершали дневную активность к 17:46–19:48, когда солнце переставало сильно греть и температуры окружающей среды заметно снижались. Не имея возможности поддерживать высокую температуру тела, ящерицы остывали, а затем закапывались в песок латеральными движениями так, что на поверхности оставалась только голова. В таком положении они пребывали примерно до 19:30–20:00 (в целом около 50 мин.). Затем, когда сгущались сумерки, некоторые ящерицы вылезали из нор или выкапывались из песка (рис. 1), и уже почти в полной темноте начинали вести вторую часть своей вечерней активности – сумеречную активность (табл. 1).
В ходе сумеречной активности круглоголовки успешно охотились на насекомых, самцы преследовали молодых особей и друг друга. В сумерках мы наблюдали три неудавшихся попытки спаривания: в двух из них самец не смог догнать убегающую самку, а в одном случае самка приняла позу отказа от спаривания. Мало того, ящерицы активно коммуницировали друг с другом, скручивая и раскручивая хвост (рис. 2) и применяя другие демонстративные поведенческие реакции (топтание на месте и т. п.).
При неожиданной встрече с человеком круглоголовки от него убегали, активно применяя различные уловки, как днем, многократно меняя направление движения в зависимости от перемещения человека, затаиваясь у основания кустов, забегая в непросматриваемую для человека зону за растительностью и прикапываясь там и др., т. е. вполне творчески и оперативно применяли весь репертуар своих защитных поведенческих реакций.

Рис. 1. Ушастые круглоголовки добровольно выкапываются из песка для начала вечерней активности. А – 17.06.2021, 19:55, фото В. А. Черлина. В – 22.06.2021, 19:20, фото с видео И. Л. Окштейна. Фотографии, сделанные с видеозаписей (рис. 1В, рис. 2) темные, недостаточно четкие и контрастные. Но технические возможности не позволяли сделать их более качественными. А если их все-таки сильно улучшить, применив Photoshop или другую подобную программу, то пропадет понимание того, что фото сделано почти ночью. Поэтому мы их оставили такими, какие они есть
Fig. 1. The toad-headed agama voluntarily dug out of the sand for the beginning of the evening activity. A – 17.06.2021, 19:55, photo by V. A. Cherlin. B – 22.06.2021, 19:20, photo from the video by I. L. Okshtein. The photographs taken from the video recordings (Fig. 1B, Fig. 2) are dark, insufficiently clear and contrasting. But the technical capabilities did not allow making them better. And if they would be improved by using Photoshop or another similar program, then the understanding that the photo was taken almost at night disappeared. Therefore, we left them as they are

Рис. 2. Активность меченых ушастых круглоголовок на поверхности в сумерках. А – 22.06.2021, 19:30. В – 22.06.2021, 19:35. Фото с видео И. Л. Окштейна
Fig. 2. Activity of the toad-headed agama on the surface at dusk. A – 22.06.2021, 7:30 pm. B – 22.06.2021, 19:35. Photo from the video by I. L. Okshtein
Таблица 1. Время протекания вечерней активности ушастых круглоголовок на бархане Сарыкум в 2021 г.
| Месяц | Время окончания дневной и вечерней активности (вместе с остыванием) | Время начала сумеречной активности | Время окончания сумеречной активности | |||
| час. | n | час. | n | час. | n | |
| Июнь | 17:34–19:48 | 8 | 19:31–20:03 | 14 | 19:58–20:23 | 7 |
| Август | 17:11–18:18 | 5 | 18:18–19:01 | 11 | 18:56–19:09 | 12 |
Перебегая с места на место на достаточно большие расстояния, ящерицы оставались приблизительно в границах своих «дневных» индивидуальных участков. Однако иногда сумеречная активность происходила далеко за границами их участков (например, у одного самца участки его дневной и сумеречной активности оказались разделены расстоянием более 50 м).
В результате можно констатировать, что в период сумеречной активности круглоголовки делали все то же самое, что и при дневной активности.
В сумерках круглоголовки были активны в среднем полчаса. Причем эта часть их активности проходила зачастую уже в плотных сумерках или даже почти в темноте (см. рис. 1, 2).
Завершалась сумеречная активность полным закапыванием в песок латеральными движениями туловища либо выкапыванием ночных нор. Последние зарегистрированные нами встречи ушастых круглоголовок во время сумеречной активности на Сарыкуме происходили примерно в 20:10–20:20.
За период наблюдений 2019 и 2021 гг. мы зарегистрировали на бархане Сарыкум более 80 эпизодов сумеречной активности ушастых круглоголовок. В июне 2021 г. мы наблюдали 43 эпизода сумеречной активности, которую проявили 22 из 44 регулярно наблюдавшихся нами особей (7 из 15 самцов, 11 из 16 самок, 4 из 13 молодых особей предположительно 2020 г. рождения).
Двухфазная вечерняя активность на Сарыкуме типично дневного и высокотемпературного вида – ушастой круглоголовки – явление в целом необычное. Несмотря на то, что оно встречается здесь регулярно, опубликованные материалы по экологии этих ящериц непосредственно на Сарыкуме (Джамирзоев и др., 2013; Мазанаева, 2009, 2020; Хонякина, 1962а, б, 1964; Roitberg et al., 2000 и др.) показали, что до нас на бархане Сарыкум это явление никто не наблюдал.
Обсуждение
Типы вечерней активности ушастых круглоголовок
Как мы уже говорили в разделе «Результаты», ситуация с окончанием дневной активности получается достаточно однородная для всех мест обитания – круглоголовки заканчивают активность примерно к 19–20 часам. Но эта одинаковость результатов, выраженных в публикациях цифрами, на деле оказывается лишь кажущейся. На самом деле у ушастых круглоголовок можно наблюдать два сильно различающихся сценария их вечернего поведения.
Первый сценарий: ушастые круглоголовки активны во второй половине дня, иногда в жаркий сезон – после дневного перерыва. Они продолжали свою вечернюю активность примерно до 19:00–20:00. К этому времени активность ящериц постепенно затухала. Это сценарий обычный, который наблюдали исследователи в разных географических точках в разные сезоны года. Он описан практически во всех публикациях, цитированных выше, за исключением упомянутых ранее наших наблюдений на бархане Сарыкум (Окштейн и др., 2021). Эту обычную, привычную ситуацию можно назвать первичной вечерней активностью.
Второй сценарий: после окончания первичной вечерней активности к вечеру ушастые круглоголовки уходили на ночевку в норы или прикапывались в верхнем слое песка. Но после этого, примерно после 19:30, они вновь выходили на поверхность и некоторое время продолжали активность уже в глубоких сумерках или даже почти в темноте примерно до 20:20, иногда – позже. Это сценарий непривычный, но реально существующий. Его впервые обнаружила группа И. Л. Окштейна на бархане Сарыкум летом 2019 г. Этот второй цикл вечерней активности можно назвать вторичной вечерней (сумеречной) активностью. Далее будет показано, что вторичная вечерняя активность отличается от первичной важными особенностями.
При обсуждении результатов наших наблюдений мы столкнулись с критикой по поводу выводов о сумеречной активности ушастых круглоголовок. По мнению оппонентов, сами исследователи могли вспугивать ящериц, которые при этом выскакивали из песка или нор, после чего бегали от наблюдателей. Но это возражение опровергается тем обстоятельством, что в процессе наблюдений за отдельными ящерицами мы регистрировали на поверхности одновременно и в тех же местах многих круглоголовок разных полов и возрастов, которые определенным образом взаимодействовали друг с другом и проявляли другие формы активности (питание, элементы регуляции температуры и т. п.). Таким образом, сумеречная активность была у них массовая, а не единичная, связанная с беспокоящей работой исследователей.
Температурные условия при вечерней активности ушастых круглоголовок
По нашим данным из Средней Азии и непосредственно с бархана Сарыкум, в период дневной активности ушастые круглоголовки имеют диапазон температур тела при полной активности («диапазон добровольных температур тела») 34–44.5º, а температуры термостабилизации («предпочитаемые») у них ~39–40º (Черлин, Музыченко, 1983а; Черлин, 2014).
Когда на Сарыкуме заканчивалась первичная вечерняя активность, т. е. когда круглоголовки, проведя вторую половину дня на поверхности, примерно в 18:30 начинали уходить в ночные убежища, температура их тела оказывалась 35–36º, т. е. была близка к нижней границе температуры полной активности для этого вида. Это мы наблюдали в Туркменистане на ст. Репетек в Каракумах (Черлин, Музыченко, 1983а; рис. 3), в Узбекистане в Кызылкумах, а также в Дагестане на Сарыкуме (рис. 4).
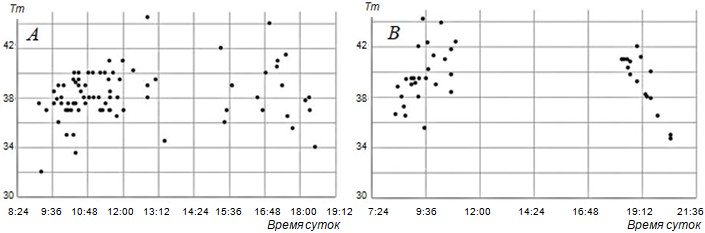
Рис. 3. Режим суточной активности и температура тела ушастых круглоголовок в состоянии полной активности по времени суток в апреле и сентябре (А) и в июне – июле (В) на ст. Репетек (Каракумы, Туркменистан). Tm – температура тела (ректальная) (Черлин, Музыченко, 1983а)
Fig. 3. Regime of daily activity and body temperature of toad-headed agamas in the state of full activity by time of day in April – September (A) and in June – July (B) at st. Repetek (Karakum, Turkmenistan). Tm – body temperature (rectal) (Cherlin, Muzychenko, 1983a)
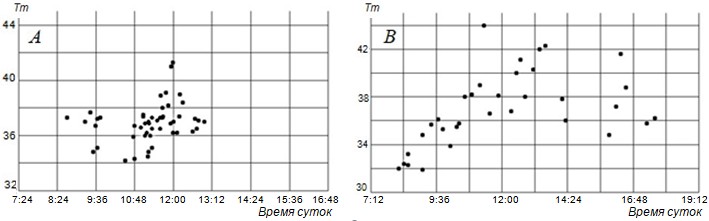
Рис. 4. Режим суточной активности и температура тела ушастых круглоголовок в состоянии полной активности по времени суток в мае (А) и июле (В) на бархане Сарыкум. Tm – температура тела (ректальная)
Fig. 4. Regime of daily activity and body temperature of toad-headed agamas in the state of full activity by time of day in May (A) and July (B) on the Sarykum dune. Tm – body temperature (rectal)
При этом в конце дневной активности интенсивность солнечного излучения резко ослабевала, сокращалась и разница между температурой тела и температурами внешней среды – воздуха (рис. 5) и особенно поверхности почвы (рис. 6).
Температуры тела ящериц в ходе вторичной вечерней (сумеречной) активности у ушастых круглоголовок на бархане Сарыкум находились в пределах 25.9–31.4°, в среднем 29.1 ± 1.7, n = 15 (табл. 2).
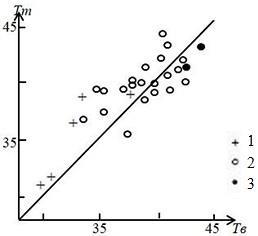
Рис. 5. Зависимость температуры тела (Tm) от температуры воздуха (Тв) у ушастых круглоголовок при утренней активности на ст. Репетек (Каракумы, Туркменистан). Условные обозначения для утренней активности: 1 – нагревание, 2 – термостабилизирующее поведение, 3 – добровольный перегрев. Прямая на графике отмечает равенство температур тела и воздуха (Черлин, Музыченко, 1983а)
Fig. 5. Dependence of body temperature (Tm) on air temperature (Тв) in toad-headed agamas during morning activity at st. Repetek (Karakum, Turkmenistan). Legend for morning activity: 1 – heating, 2 – thermostabilizing behavior, 3 – voluntary overheating. The straight line оn the graph marks the equality of body and air temperatures (Cherlin, Muzychenko, 1983a)
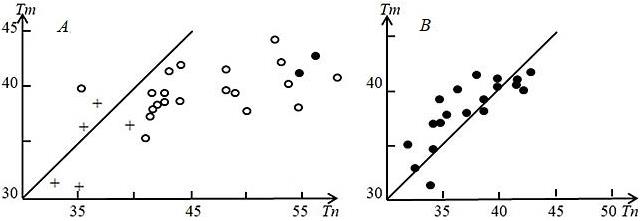
Рис. 6. Зависимость температуры тела (Tm) от температуры почвы (Тп) у ушастых круглоголовок при утренней (А, условные обозначения как на рис. 5) и вечерней (В) активности на ст. Репетек (Каракумы, Туркменистан). Прямые на графике отмечают равенство температур тела и поверхности почвы (Черлин, Музыченко, 1983а)
Fig. 6. Dependence of body temperature (Tm) on soil temperature (Тп) in toad-headed agamas during morning (A, legend as in Fig. 5) and evening (B) activity at st. Repetek (Karakum, Turkmenistan). The straight lines оn the graphs marks the equality of body and soil surface temperatures (Cherlin, Muzychenko, 1983a)
Таблица 2. Температура тела ушастых круглоголовок при вторичной вечерней активности на бархане Сарыкум
в 2021 г.
| Месяц | Ректальная температура при сумеречной активности | n |
| Июнь | 25.9–31.4 | 15 |
| Август | 28.9–29.7 | 3 |
Температурные условия внешней среды в это время приведены в табл. 3.
Таблица 3. Характерные температуры внешней среды в 20:00 в середине июня на бархане Сарыкум
| Показатель температуры среды | Температура |
| Температура воздуха на высоте 3 см | 25–31 |
| Температура поверхности песка | 28.5–29.9 |
| Температура песка на глубине 3 см | 30.0–34.3 |
| Температура песка на глубине 15 см | 32.7 |
Летом на бархане Сарыкум при перебежках в период вторичной вечерней активности круглоголовки все время держали тело приподнятым над грунтом, а при остановках либо закапывались в песок, либо продолжали держать тело приподнятым над почвой. Температура песка в это время в поверхностном слое (на глубине 3 см), куда закапывались круглоголовки, и особенно в ночных норках, куда они иногда уходили (примерно на глубине 15 см), была выше, чем на поверхности (см. табл. 3). Значит, при нахождении в грунте ящерицы, хотя и несильно, но все-таки согревались, а стоя на поверхности – охлаждались. Таким образом, очевидно, что по крайней мере часть поведения круглоголовок при сумеречной активности была терморегуляционно ориентирована. Дополнительным аргументом в пользу терморегуляционного значения нахождения в грунте в закопанном состоянии при вторичной вечерней активности служит тот факт, что в сумерках 22.06.2021, когда температура воздуха была достаточно высока (не 25–31º, как обычно, а 31.7–33.9º), все три индивидуально наблюдавшихся в этот момент круглоголовки в ходе сумеречной активности в грунт не закапывались.
Обращает на себя внимание то, что в период вторичной вечерней активности ушастые круглоголовки старались погреться на или в почве при температурах внешней среды (а следовательно, и тела) 25–31°, причем они имели возможность повысить температуру своего тела максимум до 34°. А уже при температурах среды выше ~32º они не стремились повышать температуру тела. То есть в этой ситуации уровень температуры тела в 25.9–31.4° был им достаточен и они могли реализовывать при нем полную активность, которая была характерна для них днем, только при 34–44.5º.
Данные, любезно предоставленные нам Н. А. Литвиновым, об условиях активности ушастых круглоголовок в Досанге (Астраханская область) весной и осенью 2012 и 2013 гг., говорят о том, что последние встречи ушастых круглоголовок вечером происходили там в легких сумерках (последняя встреча была в 20:17). Температура воздуха в это время была 26.9–32.6º, поверхности почвы 28.0–33.4º, а ректальная температура круглоголовок 29.3–31.5º. Эти данные соответствуют материалам, полученным на Сарыкуме при вторичной вечерней активности.
В результате ушастые круглоголовки на бархане Сарыкум и в Астраханской области (возможно, и в некоторых других местах) демонстрируют двухфазную вечернюю активность. Ящерицы были активны днем при высоких температурах внешней среды и имели диапазон температур тела при полной активности 34–44.5º. А вечером, в поздних сумерках и даже почти в темноте, в период вторичной вечерней активности они демонстрировали практически все те же, что и днем, формы активности и вполне хорошую подвижность. Но делали они это при температурах тела примерно на 10º ниже, чем днем.
По нашим представлениям, возможны по крайней мере два варианта интерпретации этой необычной ситуации. Первый вариант – у ушастых круглоголовок по каким-то причинам есть физиологическая возможность быть активными при намного более широком диапазоне температур тела, чем диапазон полной активности, проявляющийся днем. Реакция максимизации температуры тела (Черлин, 2014) заставляет их быть активными при температурах тела, близких к максимальной температуре полной активности, насколько этот уровень температур тела в данных условиях достижим. И, кроме того, у них здесь имеются очень важные мотивации, заставляющие их находиться в активном состоянии вечером, даже почти в полной темноте (например, необходимость дополнительного вечернего питания). В результате круглоголовки оказываются активными при более низких температурах тела, чем днем, хотя реакция максимизации температуры тела заставляет их нагреваться до максимально возможных в данных условиях температур, прижимаясь к более теплой, чем воздух, поверхности почты или закапываясь в еще не остывшие слои песка. Второй вариант – у ушастых круглоголовок есть два диапазона температур полной активности, разделенные хиатусом примерно в 10º. Хотя этот вариант не вписывается в представления об организации терморегуляции у этих ящериц и у рептилий в целом. Ничего подобного пока не отмечено ни у одного другого вида, ни у одной другой группы рептилий!
Световые условия при вечерней активности ушастых круглоголовок
По наблюдениям разных исследователей, в различных географических точках вечерняя активность ушастых круглоголовок оканчивалась примерно в 19–20 часов и лишь иногда продолжалась до 21 часа. Но о световых условиях при окончании вечерней активности круглоголовок никто из авторов ничего не писал. А в свете рассматриваемой нами проблемы этот материал достаточно важен.
Информацию пришлось собирать по крупицам, по полевым записям, в личных разговорах с исследователями, искать климатические данные в литературе и Интернете.
По нашим данным и материалам других авторов удалось восстановить информацию о том, что в Средней Азии, в частности в Туркмении и Узбекистане, ушастые круглоголовки действительно встречались вечером примерно до 19:30–20:20. Это была явная первичная вечерняя активность. Но важное обстоятельство заключается в том, что около 20 часов в Каракумах и Кызылкумах было еще достаточно светло, и этих ящериц встречали только пока было светло.
Нам самим приходилось много работать в Каракумах, Кызылкумах, в других среднеазиатских песках во все сезоны года, в разных биотопах, населенных ушастыми круглоголовками, и днем, и вечером, и ночью. Но никаких намеков на вечернюю активность этих ящериц после захода солнца мы никогда и нигде не отмечали. Подобное же положение дел подтвердили нам А. Ю. Целлариус и Ю. Г. Меньшиков, долгое время работавшие в Репетекском заповеднике в Каракумах (Туркменистан) и в Кызылкумском заповеднике (Узбекистан), а также Г. В. Полынова, Н. А. Литвинов, И. Ю. Барсуков и С. Б. Пугач, работавшие в разное время с этими ящерицами в окрестностях поселка Досанг (Астраханская область). В вечернее время, когда внешняя температура и уровень солнечной радиации существенно снижались, дневная активность ушастых круглоголовок заканчивалась, и примерно в 19–20 часов они уходили в ночные убежища (Литвинов, Югов, 2013). По сведениям, любезно предоставленным нам Н. А. Литвиновым, наиболее поздняя встреча ушастой круглоголовки была зарегистрирована им в 20:17, и это было только начало сумерек, т. е. еще почти в светлое время.
Получается, что летом примерно в 20 часов, когда ушастые круглоголовки заканчивают свою активность, в Каракумах и Кызылкумах еще достаточно светло, в Астраханской области и в Калмыкии начинаются сумерки, а на Сарыкуме уже плотные сумерки, т. е. почти темно. Так ли это, а если так, то почему?
Материалы табл. 4 показывают, что по чисто географическим и астрономическим причинам в Средней Азии заход солнца происходит примерно на час позже, чем в более северных широтах на юге России.
Таблица 4. Время захода солнца в разных точках обитания ушастых круглоголовок на 15 апреля, 15 июня и 1 сентября
| Географи- ческое место | Время захода солнца по датам | Источник в Интернете | ||
| 15.04 | 15.06 | 01.09 | ||
| Ашхабад | 19:42 | 20:31 | 19:38 | https://voshod-solnca.ru/sun/ашхабад |
| Элиста | 18:50 | 19:57 | 18:41 | https://ru.365.wiki/world/russia/ |
| Астрахань | 18:35 | 19:41 | 18:26 | |
| Махачкала | 18:32 | 19:30 | 18:24 | |
Что же касается сравнения районов Астраханской области, Калмыкии и Сарыкума, то здесь тоже есть некоторые географические обстоятельства. Дело в том, что и в Калмыкии, и в Досанге рельеф местности, где живут ушастые круглоголовки, более или менее равнинный и лишь слегка волнистый. Сарыкум же – бархан, на север и восток от которого тянутся равнинные степи. А вот с запада располагается хоть и не высокий, но все-таки горный Кумторкалинский хребет, с юга – более высокий хребет Нарат-Тюбе. Таким образом, вечером, на закате, прежде всего Кумторкалинский хребет загораживает Сарыкум от заходящего солнца, и это, скорее всего, способствует тому, что сумерки спускаются на бархан немного раньше, чем на другие соседние, но открытые для вечерних солнечных лучей регионы. Именно поэтому, видимо, на бархане Сарыкум темнеет немного раньше, чем в Калмыкии и Астраханской области, т. е. примерно в 20 часов на Сарыкуме уже почти темно, а в Калмыкии и в Досанге еще только начинает смеркаться. Следовательно, вполне логично, что если в опубликованных материалах, касающихся разных географических точек, написано, что круглоголовки оканчивают активность примерно в 20 часов, то на самом деле это может означать, что в Средней Азии это будет еще засветло, на юге России – начало сумерек, а на Сарыкуме – глубокие сумерки.
Единственную «пограничную» ситуацию описали В. И. Бадмаева с соавторами в Калмыкии (регион, территориально близкий к дагестанскому Сарыкуму), где весной последних круглоголовок отмечали в 19:00, в июне – в 20:00, в июле – в 21:00 (Бадмаева и др., 1981). Дело в том, что 21:00 – время, когда наверняка уже было или совсем темно, или это были плотные сумерки. Хотя и тут об условиях освещенности авторы статьи, к огромному сожалению, ничего не упомянули.
Но из всех остальных выделяются устные воспоминания Е. А. Дунаева. Он рассказал нам, что при работе в песках около пос. Досанг в Астраханской области он встречал активных ушастых круглоголовок (и самок, и самцов, и подростков) вечером, уже практически в темноте, иногда до 22 часов. Круглоголовки были вполне активны, и он некоторых из них сфотографировал (рис. 7).

Рис. 7. Ушастые круглоголовки на поверхности в плотных сумерках. Фотографии сделаны 06.08.2010 в 20:01
Е. А. Дунаевым в окрестностях пос. Досанг (Астраханская область) и любезно предоставлены нам автором
Fig. 7. The toad-headed agamas on the surface in dense twilight. Photos were taken on 06.08.2010, 20:01 by E. A. Dunaev near Dosang village (Astrakhan region) and kindly provided to us by the author
Литературные данные о ночной активности дневных видов рептилий
Некоторые авторы пытались рассматривать, анализировать и обсуждать проблему дневной / ночной активности ящериц (Vidan et al., 2017; Team, 2021). Правда, эти работы анализируют лишь внешние, статистические стороны связи активности ящериц с факторами внешней среды – среднегодовой температурой, среднегодовой ночной температурой, среднегодовым увлажнением, сезонностью параметров, аридностью и т. п. (Vidan et al., 2017). Поразительно то, что авторы посчитали, что их работа – первая, в которой предприняты попытки объяснить, зачем ящерицы активны ночью, с чем это вообще связано и что до сих пор это никому не известно. Результатом их работы явились выводы о том, что ночная активность не проявляется у ящериц, живущих в высоких широтах и высоко в горах, потому что ночью в этих регионах слишком низкие температуры. И эти очевидные выводы они посчитали «пионерными».
Однако, наверное, здесь имеет смысл вспомнить о работах, касающихся, например, термобиологии таких ночных видов, как сцинковые гекконы, Teratoscincus scincus (Черлин и др., 1983). В них подробно показано, что весь режим суточной и сезонной активности жестко связан с суточной динамикой и взаимодействием температурных условий на поверхности почвы и в ее глубине (т. е. в убежищах ящериц), что ящерицы поддерживают определенную динамику температуры своего тела, разнообразно используя условия на поверхности и в норах. Норы используются ими иногда как место, куда можно уйти от слишком высоких температур на поверхности, а иногда – как «грелки», где можно погреться, если на поверхности слишком прохладно (Черлин, Музыченко, 1983б). В других работах продемонстрированы эколого-физиологические связи динамики условий на поверхности почвы и в убежищах с режимом протекания сезонных физиологических циклов (Черлин, Целлариус, 1981; Черлин, 2014 и др.). Другими словами, сейчас уже имеется достаточно объемный материал, позволяющий понять закономерности формирования режимов дневной и ночной активности у разных групп рептилий и даже его прогнозировать (Cherlin, 1991, 2015).
Есть некоторые группы змей и ящериц, которые весной и осенью активны днем, а в летнюю жару переходят на ночную активность. Таковы множество видов гекконов (Gekkota – Крымов, 2021), среднеазиатские эфы (Echis multisquamatus – Черлин, Целлариус, 1981), гюрзы (Macrovipera lebetina – Черлин, Шепилов, 2014) и многие другие змеи и ящерицы (Богданов, 1960, 1962, 1965; Богданов, Сударев, 1989).
Для дальнейшего обсуждения нашей темы нам придется еще раз вернуться к понятию «дневные» ящерицы. Кроме температуры, оно включает характеристику дневного света, т. е. освещенности. Другими словами, дневные ящерицы – это те, которые активны в дневное, светлое время суток. Вот здесь и начинаются неопределенности и несогласованности, примеры которых демонстрируют опубликованные материалы о ночной активности ящериц, в норме являющихся дневными видами.
Имеются публикации, которые говорят о возможной ночной активности заборных игуан Sceloporus jarrovi и S. virgatis (Duncan et al., 2003). Правда, об активности этих ящериц в ночное время авторы судят исключительно по тому, что они встречаются в питании (в гнездах) мелких сов. Однако многие ящерицы в теплых регионах ночуют не в норах и других укрытиях, а на поверхности почвы или на ветках. Так часто делают, например, степные агамы (Trapelus sanguinolentus) в Средней Азии, которые летом часто проводят ночь либо на поверхности почвы, либо на ветках кустов. Теоретически мелкие совы могут охотиться на подобных ящериц и в ночное время, несмотря на то, что последние ночью спят. Но мелкие совы (как, например, сычи в Средней Азии) часто начинают активность и охоту не в полной темноте, а только при наступлении сумерек. В это время дневные ящерицы заканчивают дневную активность, вполне могут еще двигаться, устраиваясь на ночлег, и в это время с большой вероятностью могут стать их жертвой. Следовательно, наличие дневных ящериц в гнездах или в рационе мелких сов однозначно не говорит о том, что эти ящерицы активны ночью. В цитируемой нами статье нет прямых наблюдений за поимкой активных заборных игуан совами ночью. Так что, по нашему мнению, вывод о ночной активности этих ящериц в данной работе не доказан.
Другая публикация описывает ночную активность дневного гребенчатого анолиса (Anolis cristatellus) при лунном свете (Brisbane, van den Burg, 2020). Работы проводились на острове Доминика (Малые Антильские острова) в середине января 2019 г. В данном случае авторы непосредственно наблюдали, как анолисы примерно в 20 часов по местному времени питались мелкими лягушками, и даже привели в статье соответствующие фотографии. Таким образом, ночная (вечерняя) активность дневных ящериц документально подтверждена.
Однако, без всякого сомнения, объяснить ночную активность анолисов, не изучая температурные условия, практически невозможно. К сожалению, авторы не исследовали причины и условия, способствовавшие данной ситуации. Мы попытались сами восстановить недостающие материалы. В январе температуры воздуха в Доминике днем достигают 30°, а ночью опускаются примерно до 25° или немного ниже (География Доминики, 2021; Климат Доминики, 2021). Но анолисы были встречены примерно в 20 часов, т. е. в самом начале темного периода, а значит, температура явно не успевала в это время упасть ниже 27–29°.
Данные из различных литературных источников показывают, что температура тела гребенчатых анолисов при полной активности находится в очень узких пределах 29–30° (Huey, Webster, 1976; Lister, 1976). Следовательно, в начале ночи, а точнее – поздним вечером температуры воздуха позволяли гребенчатым анолисам иметь температуру тела, близкую к нижней границе диапазона температур их полной активности, или даже вообще достигать этого диапазона. То есть температурные условия давали им возможность быть активными в это время. А яркий лунный свет дополнял эффект, допускающий и активизирующий такую вечернюю активность. Таким образом, описанная ситуация с анолисами имеет вполне понятные причины и объяснимые условия. Она является типичным примером продолжения, «растягивания» дневной активности, т. е. примером первичной вечерней активности.
Эти и другие материалы позволяют заключить, что типично дневные виды рептилий могут переходить на вечерне-ночной образ жизни, но чаще всего в особых случаях, когда внешние климатические условия позволяют им поддерживать важнейшие физиологические параметры (в частности и прежде всего – температуру тела) в границах, допускающих полную активность. В природных условиях должны быть достаточно веские причины для такой смены поведения. Видимо, основная причина – это питание: в течение дня по ряду обстоятельств ящерицам не хватает питания, а ночью у лампы, где собираются насекомые, «добрать» недостающее количество корма достаточно просто, и это не требует больших энергетических затрат.
Ряд описанных в литературе вариантов необычной вечерне-ночной активности типично дневных видов рептилий касается некоторых представителей варанов. Вся их биология, экология, строение колбочек в глазах (Röll, Horn, 1999 и др.) свидетельствуют о том, что это типично дневные виды. Тем не менее в литературе есть указания на сумеречную и даже ночную активность в природе по крайней мере у 9 видов варанов – Varanus spenceri, V. panoptes, V. tristis, V. glebopalma, V. rosenbergi, V. melinus, V. glauerli, V. salvator, V. dumerilli, а в условиях неволи у V. acanthurus, V. giganteus, V. indicus, V. keithhornei, V. mertensi, V. varius (Bohme, Ziegler, 1997; Cota et al., 2008; Irwin et al., 1996a, b; Jones, 1998; Trembauth, 2000; Yong et al., 2008 и др.).
Есть предположения о том, что V. glebopalma питается в основном в первые два часа после захода солнца (Christian, 1977; Wilson, Knowles, 1988). В природе активность, охоту и питание этих ящериц отмечали в 18:30–19:00 и с 19 до 21 часа. Тем не менее радиотелеметрические исследования экологии V. glebopalma вечерней и ночной активности у них не выявили (Sweet, 1999). С. Свит посчитал, что наблюдавшаяся другими исследователями вечерняя и ночная активность варанов являлась результатом того, что они беспокоили ящериц во время работы.
Однако этих варанов наблюдали охотящимися и поедающими лягушек примерно с 20 до 23 часов при температуре воздуха 19.5º (Rhind et al., 2013). Не исключено, что наличие или отсутствие активности в разных работах отражают популяционные и / или сезонные различия. Возможно также, что варанов из мест их ночевок вытесняют какие-то хищники (Rhind et al., 2013). Кроме того, замечено, что в ночное время V. glebopalma предпочитают засадную, «поджидательную» стратегию охоты, что вообще не характерно для варанов. Именно это может способствовать тому, что они могут охотиться при низкой освещенности (Rhind et al., 2013). Мало того, по нашему мнению, возможности охоты в сумерках и ночью у варанов способствует то, что в их биологии, в ориентации во внешней среде, при социальных коммуникациях и при охоте большое значение имеет, кроме зрения, еще и осязательная информация, получаемая при помощи языка, похожего на змеиный, которым они ощупывают все вокруг. Исследуя таким образом следы на почве, они способны различать знакомых варанов, обитающих на данной территории, находить пищевые объекты и получать другую важную информацию, что мы наблюдали при работе с серыми варанами, Varanus griseus, в Кызылкумах (Целлариус и др., 1991). Осязание у V. glebopalma в значительной степени может замещать информацию зрительную и давать возможность быть активными почти без света.
В этой же связи можно предположить, что вечернюю и ночную активность можно было бы ожидать, например, у обитающих в тропических регионах, живущих в теплой лесной подстилке мелких сцинковых ящериц, хотя пока нам такие опубликованные данные не известны. Но по устным сообщениям А. П. Лисачева, в Малайзии он наблюдал ночную активность бронзовых мабуй (Eutropis macularia) примерно при 25º. Причем в основном их активность на поверхности проявлялась именно ночью, а не днем, не утром или вечером, когда они, видимо, грелись в верхнем слое подстилки при более высоких температурах. Кроме того, в тех же районах ночную активность проявляли сцинки родов Lygosoma и Sphenomorphus. В целом же среди сцинковых ящериц Евразии 20 видов могут быть признаны ночными, 165 видов – дневными и 22 вида – способными вести и дневной, и ночной образ жизни (Vidan et al., 2017).
Интересные материалы имеются по экологии хентаунской круглоголовки (Phrynocephalus rossikowi). Этот мелкий вид круглоголовок распространен в нижнем течении Амударьи, в Каракалпакии и прилежащих районах Узбекистана (Банников и др., 1977). Их находили на узкой полосе щебня, протянувшейся вдоль левого берега Амударьи, и на других щебнистых равнинах. Данных об экологии этого вида ящериц в литературе немного. Наиболее подробные исследования на эту тему проводились Д. А. Бондаренко в Дейнауском районе Чарджоуской области летом и осенью (Бондаренко, 1984). Он показал, что эти ящерицы, обитая иногда на песках, могут, как песчаные круглоголовки, закапываться в верхний слой почвы боковыми движениями тела, и так же они роют норки (Бондаренко, 1984).
По его устному сообщению, в июле, когда температуры среды были очень высокими, круглоголовки были активны в двухпиковом режиме. После короткого периода утренней активности круглоголовки уходили с поверхности и вновь начинали появляться только после 18 часов. Пик активности приходился примерно на 20 часов. Последние круглоголовки встречались уже в сумерках. Этот вид оказался очень неустойчив к высокой температуре среды.
В начале августа у хентаунских круглоголовок наблюдалось два пика активности – утренний и вечерний. Утром круглоголовки появлялись, когда температура поверхности почвы была около 25º. Максимальная активность ящериц в первой и второй половине дня – при температуре воздуха 32–34º, почвы – 35–42º. В середине дня в их активности наступал перерыв. Ящерицы уходили с поверхности при температуре воздуха 36–37º, почвы – 46–48º. При таких температурах они не могли находиться на поверхности. Их биотопы располагались на открытой, хорошо прогреваемой каменистой поверхности, которая, разогреваясь за день, длительное время сохраняла высокую температуру, неблагоприятную для хентаунских круглоголовок. Это определяло продолжительность дневного перерыва в их активности. В это время температура в норах ящериц на глубине 15 см была около 34.5º, что для них вполне комфортно. Вечером ящерицы появлялись поздно, при температуре воздуха 36–39º, почвы – 42–44º. Исчезали они с поверхности к 20:30, когда температура воздуха снижалась до 32–33º, почвы – до 27–28º. Отдельные особи сохраняли активность в сумерках и прекращали ее, затаившись на поверхности с наступлением темноты. Этим объясняется раннее начало утренней активности у хентаунских круглоголовок: ящерицы, оставшиеся на поверхности с вечера, утром быстро разогревались и рано начинали активность.
Осенью (в конце октября) цикл активности хентаунских круглоголовок был неустойчивым в связи с переменной погодой. Активность их была однопиковой. Утром активные круглоголовки появлялись на поверхности с 7:30. Они начинали активно передвигаться по территории и питаться при температурах среды не ниже 20°. Прекращалась активность с заходом солнца и наступлением темноты. В это время температура воздуха и поверхности почвы снижалась до 25° и ниже. А в пасмурную, ветреную погоду, когда температуры внешней среды не поднимались выше 12–14º, хентаунские круглоголовки уже не встречались (Бондаренко, 1979, 1982).
Хотя прямых измерений температуры тела хентаунских круглоголовок Д. А. Бондаренко не производил, но по его же косвенным данным можно сделать предварительное заключение о температурах тела при их полной активности. Скорее всего, активность хентаунских круглоголовок днем начиналась при температурах тела примерно от 30–32º, а заканчивалась – около 37–39º.
Исследования на близкородственных видах показали, что у пестрой круглоголовки Phrynocephalus versicolor максимальная зарегистрированная температура (максимальная температура полной активности) была 38.5º (Кропачев, 2013), а у круглоголовки-вертихвостки Phrynocephalus guttatus этот показатель составлял, по одним данным, 40.9º (Югов и др., 2014), по другим – 41.3° (Окштейн, 2016). К сожалению, по материалам многих авторов минимальные температуры полной активности определить не удается вследствие недостаточной определенности примененной методики исследований. Но, по данным И. Л. Окштейна, минимальная температура полной активности у круглоголовки-вертихвостки составляет примерно 35.7° (Окштейн, 2016). У тибетской круглоголовки Phrynocephalus theobaldi минимальная температура полной активности была 32º, максимальная – около 39º (Nanhoe, Ouboter, 1987). Таким образом, температурные характеристики активности хентаунских круглоголовок вполне укладываются в диапазоны близкородственных видов.
Следовательно, у хентаунских круглоголовок, так же как и у ушастых, наблюдалась вечерняя активность, вплоть до почти полной темноты. При этом, судя по материалам наблюдений, вечерняя активность этих круглоголовок происходила в температурных условиях, которые позволяли им удерживать температуру тела хотя бы на нижнем уровне диапазона температуры полной активности – 30–32º. Скорее всего, это было окончание дневной активности, т. е. первичная вечерняя активность.
Если описанные выше случаи иллюстрируют чисто природные ситуации, позволяющие увидеть вечернюю активность дневных видов, то есть и другие случаи, описывающие вечернюю активность дневных рептилий, связанную с деятельностью человека. Речь идет об агаме колонистов Agama agama (Amadi et al., 2021) и о грузинской ящерице Darevskia rudis (Afsar et al., 2018). Оба эти случая, по сути, похожи друг на друга. Дело в том, что и агама, и грузинская ящерица были активны в вечернее и ночное время под осветительными лампами накаливания, которые использовались человеком для освещения и были закреплены на стенах домов. При этом у агам были изучены и температуры тела. Охотящиеся после захода солнца под лампами ящерицы имели значительно более высокую температуру тела (в среднем 36.18º), чем неактивные ящерицы (в среднем 31.94º). При этом днем агамы имели среднюю температуру тела при активности примерно 36º (James, Porter, 1979). То есть агамы в период ночной активности с помощью ламп накаливания поддерживали у себя такую же температуру тела, что и днем при солнечном свете. Однако вариации в температуре тела были значительно выше у ящериц, добывающих пищу, чем у неактивных ящериц. Вероятно, это происходило из-за необходимости курсировать между теплом под лампами накаливания и более прохладными участками, где приходилось охотиться. Регрессионный анализ температуры тела в зависимости от времени у охотящихся и не охотящихся ящериц подтвердил предположение о том, что эти ящерицы поддерживали температуру тела путем активной терморегуляции, в то время как у неактивных и, следовательно, не греющихся ящериц в течение того же периода времени температура тела снижалась. Но данные этой работы не позволили определить, является ли основной мотивацией ночной активности использование ламп накаливания для терморегуляции или для охоты (Amadi et al., 2021).
Такая же ночная активность, связанная с использованием ночных осветительных ламп, отмечается иногда у мелких анолисов (Amadi et al., 2021).
Все описанные в литературе случаи вечерней активности дневных ящериц являются естественным окончанием, угасанием их дневного цикла активности, т. е. это первичная вечерняя активность. И закономерностью этого типа вечерней активности является то, что ящерицы заканчивают суточный цикл, постепенно остывают, переходя в малоактивное состояние, и рано или поздно уходят в ночные убежища или просто в места ночевки (например, на поверхности). Оканчивается первичная вечерняя активность у всех этих видов при температурах тела, приближающихся к минимальной температуре диапазона полной активности, из которого температура их тела не выходит.
Характерные особенности, предположительные причины и смысл вторичной вечерней активности
Обобщив все описанные выше факты о феномене вторичной вечерней активности у ушастых круглоголовок на бархане Сарыкум, можно однозначно заключить следующее. Во время вторичной вечерней активности ушастые круглоголовки проявляют полную активность, демонстрируя большинство тех же элементов своего поведения (питание, социальные контакты, хотя и несколько ослабленные, попытки спаривания и т. п.), что и днем. При этом характерной особенностью вторичной вечерней активности является то, что они проявляют почти весь репертуар своей дневной полной активности, но только при температурах тела примерно на 10° ниже, чем днем!
Мы, естественно, задались вопросом о смысле, назначении этого явления. Ведь совершенно очевидно, что если дневной, высокотемпературный вид переходит на время к почти ночной активности, значит, на то должны быть очень веские причины. Судя по всему, на Сарыкуме, как и при переходе дневных видов к ночной активности у ламп освещения, о чем мы рассказывали выше, основная мотивация связана с питанием. По нашим наблюдениям, днем на поверхности песка на бархане не так много беспозвоночных, которые могли бы стать пищей для круглоголовок. Мало того, днем на бархане часто дуют очень сильные ветры, которые просто сдувают с поверхности все мелкие, в том числе и кормовые, объекты. Учитывая это, а также то, что на Сарыкуме большая плотность популяции этих ящериц, можно предположить, что у ушастых круглоголовок может ощущаться дефицит в количестве корма, который можно добыть днем. В сумерках же, особенно летом, идет массовый лёт всевозможных насекомых, особенно жуков (разные виды мелких хрущей и т. п.). Мы наблюдали, что даже в очень плотных сумерках круглоголовки активно за ними охотятся и поедают. Более того, сумеречная активность позволяет охотиться и за пределами «дневного» индивидуального участка (см. рис. 1).
Однако данные о сумеречной активности ушастых круглоголовок в окрестностях пос. Досанг показывают, что во вторичной вечерней активности пищевая мотивация не всегда может быть ведущей. Дело в том, что в песках под Досангом в дневное время достаточно много насекомых, которые могут служить круглоголовкам пищей. Следовательно, здесь пищевые потребности не могут быть мотивационными стимулами для них вести сумеречную активность. Здесь должны быть другие мотивации.
В конце августа 2021 г. нами было выявлено резкое уменьшение частоты социальных контактов между круглоголовками при дневной активности. В ходе сумеречной активности социальные контакты также почти прекратились, однако сама сумеречная активность сохранила свою продолжительность и температурные параметры, сдвинувшись по времени примерно на час раньше вслед за моментом наступления темноты (см. табл. 1 и 2, по освещенности и температурам во всех указанных точках 19 ч. в конце августа соответствовали 20 ч. в июне). Видимо, в качестве дополнительного времени для социальных контактов сумеречная активность имеет меньшее значение, чем для питания. Сеголетки, родившиеся (судя по размерам) в июле – августе, в конце августа вели сумеречную активность так же, как и взрослые особи.
Как мы уже упоминали выше, кроме прочего, часть вторичной вечерней активности ушастых круглоголовок была связана еще и с терморегуляцией.
Можно предложить по крайней мере еще одно соображение по поводу мотивации сумеречной активности ушастых круглоголовок на юге России. Достаточно давно, когда существовала связь песчаных пустынь Средней Азии и территорий Северного Кавказа, ушастые круглоголовки заселили эти регионы. Из результатов наших исследований видно, что термофизиологические характеристики (диапазон температуры полной активности, температуры термостабилизации) у круглоголовок из этих регионов не различаются. Логично предположить, что есть и другие физиологические особенности, которые могут оставаться у них стабильными и неизменными. В частности, таковыми могут быть особенности генетически закрепленных физиологических суточных (циркадных) и сезонных ритмов активности. Из материалов табл. 4 видно, что когда в районе Махачкалы, т. е. на бархане Сарыкум, солнце уже заходит, в Средней Азии (Ашхабад) оно будет светить еще больше часа (рис. 8).
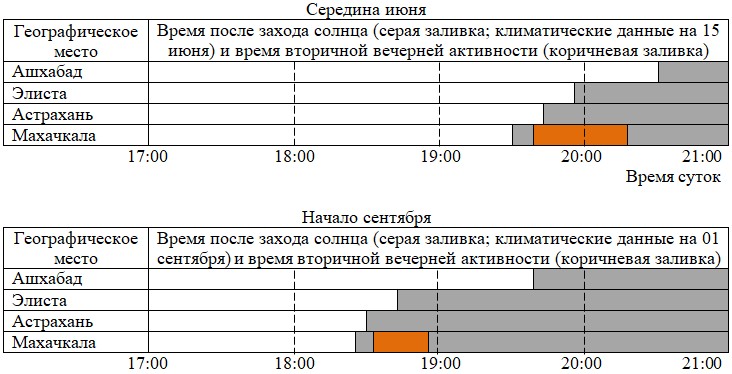
Рис. 8. Время захода солнца и проявлений вторичной вечерней активности в разных регионах, населенных ушастыми круглоголовками
Fig. 8. Time of sunset and manifestations of secondary evening activity in different regions inhabited by toad-headed agamas
Значит, несмотря на то, что солнце в Дагестане уже садится, по физиологическим циркадным суточным ритмам у дагестанских ушастых круглоголовок, скорее всего, еще должна продолжаться активность. Видимо, это может быть важной мотивацией, мешающей им полностью завершить дневной цикл активности адекватно условиям освещенности и температуры.
Можно предположить, что, когда освещенность начинает падать, инсоляция ослабевает и температуры внешней среды понижаются, это является для дневных высокотемпературных ушастых круглоголовок физиологически закрепленным сигналом для завершения дневной активности. Подчиняясь ему, ящерицы уходят с поверхности, зарываясь в верхний слой песка или выкапывая ночную норку. Далее сам по себе сигнал ухода в убежища, реализовавшись, ослабевает, а циркадные ритмы суточной активности ящериц входят в конфликт с внешними условиями и начинающейся фазой ночного сна. И, подчиняясь этой циркадной ритмике, круглоголовки вновь выходят на поверхность, продолжая активность даже в условиях, которые, по нашему представлению, не совсем для этого подходят. Заканчивается эта вторичная вечерняя активность у круглоголовок, видимо, когда температурные условия или какие-то другие непонятные пока обстоятельства становятся слишком неподходящими.
При этом, как уже говорилось выше, у этих ящериц, видимо, имеются и не известные пока физиологические особенности, позволяющие им быть активными при существенно более низких температурах тела, чем днем.
Описанное предположение может касаться не только Дагестана, но и других мест юга России, где живут ушастые круглоголовки (Астраханская область, Калмыкия) и где они встречались иногда вплоть до 21 часа (Бадмаева и др., 1981 и др.).
Таким образом, ушастые круглоголовки, живущие на юге России, проявляют вторичную вечернюю активность в период времени, когда в Средней Азии, т. е. на их исходной территории, солнце еще светит и остальные условия позволяют быть полностью активными (см. рис. 8).
Однозначных экспериментальных доказательств этой закономерности у нас нет, но вся логика наблюдаемых явлений, с одной стороны, не опровергает подобное утверждение, с другой – говорит о том, что существование такой закономерности вполне допустимо.
Из всего сказанного ясно одно – мотивации вести вечернюю и сумеречную активность у ушастых круглоголовок могут быть различными, но при этом явно присутствуют некоторые пока не известные физиологические характеристики, которые позволяют им в это время быть активными в полном объеме, только при температурах тела, существенно ниже, чем днем.
***
В результате наблюдений появился ряд новых вопросов, на которые пока нет ответов.
Обращает на себя внимание одно обстоятельство: во всех регионах, по крайней мере на территории бывшего СССР, населенных ушастыми круглоголовками, они заканчивают свою дневную активность, т. е. первичную вечернюю активность, примерно в 20:00. Это похоже на проявление врожденных, видоспецифичных циркадных ритмов. Мотивации, заставляющие ушастых круглоголовок в отдельных регионах вести вторичную вечернюю активность, требуют отдельного, дополнительного изучения.
Не исключено, что ушастые круглоголовки активны в двух самостоятельных температурных режимах. В дневное время – при температурах тела в диапазоне примерно 34–44º, а поздним вечером – в совершенно другом температурном режиме при температурах тела примерно 27–32º, то есть на 5–12º ниже! Как устроена, как организована термофизиология ушастых круглоголовок, что она позволяет им иметь два температурных режима активности, два диапазона температур тела при полной активности, разделенных хиатусом примерно в 10º? Как может быть организована у них регуляция температуры тела и как это вообще возможно?
Ушастые круглоголовки активны вечером до того времени, когда уже практически окончательно стемнеет. Мы наблюдали, когда круглоголовка уже почти в темноте заметила небольшого жука на расстоянии около 6 м, подбежала к нему и съела. Как они видят в условиях критического недостатка света? Ведь их глаза приспособлены для того, чтобы видеть, наоборот, при очень ярком свете!
Заключение
Ушастые круглоголовки на бархане Сарыкум проявляют необычную для них, как типично дневных видов, вечернюю и даже сумеречную активность. Вначале, заканчивая первичную активность вечером, они уходят с поверхности в убежища. Но затем через некоторое время вновь выходят и начинают вторичную вечернюю активность уже почти в полной темноте (примерно до 21:00).
В течение сумеречной активности круглоголовки делают почти все то же, что они делают днем: питаются, осуществляют социальные коммуникации, самцы пытаются спариваться с самками и т. п.
Температуры тела ушастых круглоголовок при вторичной сумеречной активности находятся в диапазоне, который примерно на 10° ниже, чем при дневной активности.
Мотивация для вечерней и сумеречной активности именно на Сарыкуме связана, видимо, с необходимостью дополнительного питания, поскольку пищевых объектов для них на бархане в дневное время по ряду причин недостаточно, а вечером там наблюдается массовый лёт мелких кормовых насекомых.
Мотивации для вечерней и сумеречной активности у ушастых круглоголовок в разных местах могут быть различными. Но при этом явно присутствуют некоторые пока не известные физиологические характеристики, которые позволяют им в это время быть активными в полном объеме, но при температурах тела существенно ниже, чем днем.
Библиография
Бадмаева В. И., Лебеденко Н. А., Савина Н. А. Суточная активность ушастой круглоголовки в Калмыкии // Вопросы герпетологии: Авторефераты докладов. Пятая Всесоюзная герпетологическая конференция. Ашхабад, 22–24 сентября 1981 г. 1981. Л.: Наука. Ленинградское отделение, 1981. С. 11–12.
Банников А. Г., Даревский И. С, Ищенко В. Г., Рустамов А. К., Щербак Н. Н. Определитель земноводных и пресмыкающихся фауны СССР . М.: Просвещение, 1977. 415 с.
Богданов О. П. Фауна Узбекской ССР. Земноводные и пресмыкающиеся . Ташкент: Изд-во АН УзбССР, 1960. 260 с.
Богданов О. П. Пресмыкающиеся Туркмении . Ашхабад: Изд-во АН Туркм. ССР, 1962. 235 с.
Богданов О. П. Экология пресмыкающихся Средней Азии . М: Наука, 1965. 257 с.
Богданов О. П., Сударев О. Н. Экология пресмыкающихся . Ташкент: Укитвувчи, 1989. 128 с.
Божанский А. Т. Ушастая круглоголовка // Красная книга Астраханской области. Астрахань: Астраханский гос. ун-т: Издательский дом «Астраханский университет», 2014. С. 265–266.
Бондаренко Д. А. Некоторые сведения по экологии хентаунской круглоголовки // Материалы 11 конференции молодых ученых по проблемам охраны живой природы ВНИИ охраны природы МСХ. М., 1979. С. 41–42.
Бондаренко Д. А. Экология хентаунской круглоголовки в летне-осенний период // Вестник зоологии. 1982. № 5. С. 55–59.
Бондаренко Д. А. Хентаунская круглоголовка // Природа. 1984. № 4. С. 48–49.
География Доминики . URL: https://ru.abcdef.wiki/wiki/Geography_of_Dominica (дата обращения 01.10.2021).
Джамирзоев Г. С. и др. Редкие позвоночные животные заповедника «Дагестанский» / Под ред. Г. С. Джамирзоева и С. А. Букреева. Махачкала: Государственный заповедник «Дагестанский», 2013. 372 с. (Труды заповедника «Дагестанский». Вып. 6).
Доронин И. В. Ушастая круглоголовка // Красная книга Ставропольского края. Ставрополь: ООО «Астерикс», 2013. С. 144.
Ждокова М. К. Ушастая круглоголовка // Красная книга Республики Калмыкия. Т. 1. Животные. Элиста: ЗАОр «НПП «Джангар», 2013. С. 94–95.
Киреев В. А. Земноводные и пресмыкающиеся. Животный мир Калмыкии . Элиста, 1983. 112 с.
Климат Доминики . URL: http://www.world-globe.ru/countries/dominica/climate/ (дата обращения: 01.10.2021).
Кропачев И. И. Термобиология пестрой круглоголовки Кулагина, Phrynocephalus versicolor kulagini Bedriaga, 1909 (Squamata: Agamidae) на северной границе ареала вида, в Туве // Известия Самарского научного центра. 2013. Т. 15, № 3 (7). С. 2319–2327.
Крымов Н. Г. Гекконы Австралии и Океании. Распространение. Содержание. Разведение . Барнаул: Алтай, 2021. 596 с.
Литвинов Н. А., Югов М. В. Температура тела и микроклиматические условия обитания двух видов круглоголовок в Северном Прикаспии // Вестник ПГГПУ. Сер. 2. Физико-математические и естественные науки. 2013. С. 19–25.
Лотиев К. Ю. Ушастая круглоголовка // Красная книга Чеченской Республики. Ростов н/Д: ООО «Южный издательский дом», 2020. С. 340–342.
Мазанаева Л. Ф. Ушастая круглоголовка // Красная книга Республики Дагестан. Махачкала, 2009. С. 387–388.
Мазанаева Л. Ф. Ушастая круглоголовка Phrynocephalus mystaceus (Pallas, 1776) // Красная книга Республики Дагестан. Махачкала: Республиканская газетно-журнальная типография, 2020. С. 479–481.
Никольский А. М. Пресмыкающиеся (Reptilia) Chelonia и Sauria (Черепахи и ящерицы). Т. 1. Фауна России . Пг., 1915. 534 с.
Окштейн И. Л. Термобиология круглоголовки-вертихвостки Phrynocephalus guttatus (Gmelin, 1789) в Астраханской области // Вестник СПбГУ. Сер. 3. 2016. Вып. 3. С. 107–112.
Окштейн И. Л., Черлин В. А., Вето Д. В., Кожара В. В., Корнеева А. А., Ложкина А. В., Мироненко-Маренков А. Д., Рябова Е. Д., Тищенко М. А., Турова М. А., Фраерман А. Ю., Яцук А. А. Сумеречная активность ушастых круглоголовок (Phrynocephalus mystaceus) на песчаном массиве Большой Сарыкум в Дагестане // Вопросы герпетологии: Программа и тезисы докладов VIII съезда Герпетологического общества имени А. М. Никольского при РАН «Современные герпетологические исследования Евразии» 3–9 октября 2021 г. М.: КМК Scientific Press, 2021. С. 199–201.
Параскив К. П. Пресмыкающиеся Казахстана . Алма-Ата: Изд-во Акад. наук КазССР, 1956. 228 с.
Саид-Алиев С. А. Земноводные и пресмыкающиеся Таджикистана . Душанбе: Дониш, 1979. 147 с.
Тертышников М. Ф. Пресмыкающиеся Центрального Предкавказья . Ставрополь, 2002. 240 с.
Тертышников М. Ф., Горовая В. И. К изучению западных популяций ушастой круглоголовки и круглоголовки-вертихвостки // Вид и его продуктивность в ареале. Свердловск, 1984. С. 43–44.
Хонякина З. П. Динамика численности ушастой круглоголовки в районе Кумторкалы ДАССР // Зоологический журнал. 1962а. Т. 41. Вып. 5. С. 778–780.
Хонякина 3. П. Сезонная и суточная активность ушастой круглоголовки (Phrynocephalus mystaceus Pall.) в районе Кумторкалы Дагестанской АССР // Ученые записки Дагестанского государственного университета. Биологические науки (зоология, паразитология и физиология). 1962б. Т. XI. С. 133–154.
Хонякина З. П. Ящерицы Дагестана : Автореф. дис. ... канд. биол. наук. Махачкала, 1964. 20 с.
Целлариус А. Ю., Черлин В. А., Меньшиков Ю. Г. Предварительное сообщение о работах по изучению биологии Varanus griseus (Reptilia, Varanidae) в Средней Азии // Герпетологические исследования. Л.: ЛИСС, 1991. № 1. С. 61–103.
Черлин В. А. Рептилии: температура и экология . Saarbrücken: Lambert Academic Publishing, 2014. 442 с.
Черлин В. А., Музыченко И. В. Термобиология и экология сетчатой ящурки (Eremias grammica), ушастой (Phrynocephalus mystaceus) и песчаной (Ph. interscapularis) круглоголовок летом в Каракумах // Зоологический журнал. 1983а. Т. 62, № 6. С. 897–908.
Черлин В. А., Музыченко И. В. Использование нор в терморегуляции рептилий // Прикладная этология: Материалы III Всесоюзной конференции по поведению животных. Т. 3. М.: Наука, 1983б. С. 172–174.
Черлин В. А., Целлариус А. Ю. Зависимость поведения песчаной эфы, Echis multisquamatus Cherlin 1981 от температурных условий в Южной Туркмении // Фауна и экология амфибий и рептилий палеарктической Азии. Л.: Наука, 1981. С. 96–108 (Труды Зоологического института АН СССР. Т. 101).
Черлин В. А., Целлариус А. Ю., Громов А. В. К температурной биологии сцинкового геккона (Teratoscincus scincus) в Каракумах // Экология. 1983. № 2. С. 84–87.
Черлин В. А., Шепилов С. А. Термобиология среднеазиатской гюрзы (Macrovipera lebetina turanica) хребта Нуратау и гюрзы Чернова (Macrovipera lebetina černovi) западных Кызылкумов // Зоологический журнал. 2014. Т. 93, № 2. С. 242–247.
Шаммаков С. Пресмыкающиеся равнинного Туркменистана / Под науч. ред. А. К. Рустамова. Ашхабад: Ылым, 1981. 311 с.
Шнитников В. Н. Пресмыкающиеся Семиречья . Кзыл-Орда: Об-во изучения Казахстана, 1928. 85 с. (Труды Общества изучения Казахстана. 1928. Т. 8, вып. 3).
Югов М. В., Литвинов Н. А., Галиулин Д. М., Окулов Г. А. Термобиология круглоголовки-вертихвостки (Phrynocephalus guttatus, Gmelin, 1789) в Прикаспийской низменности // Известия Самарского научного центра. 2014. Т. 16, № 5 (1). С. 448–450.
Afsar M., Sahin M. K., Afsar B., Çiçek K., Tok C. V. Data on nocturnal activity of Darevskia rudis (Bedriaga, 1886) (Sauria: Lacertidae) in Central Black Sea Region, Turkey // Ecologica Montenegrina. 2018. Vol. 19. P. 125–129.
Amadi N., Luiselli L., Belema R., Nyiwale G. A., Wala C., Urubia N., Meek R. From diurnal to nocturnal activity: a case study of night-light niche expansion in Agama agama lizards // Ethology Ecology & Evolution. 2021. DOI: 10.1080/03949370.2021.1883120.
Böhme W., Ziegler T. Varanus melinus sp. n., ein neuer Waran aus der V. indicus-Gruppe von den Molukken, Indonesien // Herpetofauna. 1997. Vol. 19. P. 26–34.
Brisbane J. L. K., van den Burg M. P. No need for artificial light: nocturnal activity by a diurnal reptile under lunar light // Neotropical Biodiversity. 2020. Vol. 6, No 1. P. 193–196.
Cherlin V. A. Method of Ecology-Physiological Matrix of Species in Researches on Reptile Biology // Герпетологические исследования. Л.: ЛИСС, 1991. № 1. С. 138–146.
Cherlin V. A. Physiology-ecological matrix and its significance in biology of reptiles // Some important aspects of thermal biology of reptiles. Saarbrücken: Lambert Academic Publishing, 2015. P. 117–125.
Cota M., Chan-Ard T., Mekchai S., Laoteaw S. Geographical Distribution, Instinctive Feeding Behavior and Report of Nocturnal Activity of Varanus dumerilii in Thailand // Biawak. 2008. Vol. 2, No 4. P. 152–158.
Christian T. Notes on Varanus glebopalma // Vic. Herp. Soc. Newsletter. 1977. Vol. 6. P. 11–13.
Duncan W. W., Gehlbach F. R., Middendorf III G. A. Nocturnal activity by diurnal lizards (Sceloporus jarrovi, S. virgatis) eaten by small owls (Glaucidium gnoma, Otus trichopsis) // The Southwestern Naturalist. 2003. Vol. 48, No 2. P. 218–222.
Huey R. B., Webster P. T. Thermal biology of Anolis lizards in a complex fauna: the cristatellus group on Puerto Rico // Ecology. 1976. Vol. 57. Р. 985–994.
Irwin S., Lyons B., Frisby T. Nocturnal activity by Varanus panoptes at Cape Melville // Herpetofauna (Sydney). 1996a. Vol. 26, No 2. P. 50.
Irwin S., Engle K., Mackness B. Nocturnal Nesting by Captive Varanid Lizards // Herpetol. Rev. 1996b. Vol. 27, No 4. P. 192–194.
James F. C., Porter W. P. Behavior-microclimate relationships in the African rainbow lizard, Agama agama // Copeia. 1979. No 4. P. 585–593.
Jones A. Nocturnal activity in captive varanid lizards // Herpetofauna (Sydney). 1998. Vol. 28, No 2. P. 50–51.
Lister B. C. The nature of niche expansion in West Indian Anolis lizards. Ecological consequences of reduced competition // Evolution. 1976. Vol. 30. P. 659–676.
Nanhoe L. M. R., Ouboter P. E. The distribution of reptiles and amphibians in the Annapurna-dhaulagiri region (Nepal) // Zool. Verh. Leiden. 1987. Vol. 240. P. 1–105.
Rhind D., Doddy J. S., Ellis R., Ricketts A., Scott G., Clulow S., McHenry. Varanus glebopalma (Black-palmed Monitor). Noctural activity and foraging // Herpetological Review. 2013. Vol. 44, No 4. P. 687–688.
Roitberg E. S., Mazanaeva L. F., Ilyina E. V., Orlova V. F. Die Echsen Dagestans (Nordkaukasus, Russland): Artenliste und aktuelle Verbreitungsdaten (Reptilia: Sauria: Gekkonidae, Agamidae, Anguidae, Scincidae et Lacertidae) // Faunistische Abhandlungen, Staatliches Museum. Tierkunde. Dresden. 2000. Vol. 22, No 8. P. 97–118.
Röll B., Horn H.-G. The structure of the eye in the monitor lizard Varanus griseus caspius (Reptilia: Varanidae) // Advances in Monitor Research II. (Eds. H.-G. Horn and W. Böhme), Mertensiella. 1999. Vol. 1. Р. 291–306. Rheinbach, Germany.
Sweet S. S. Spatial ecology of Varanus glauerti and V. glebopalma in northern Australia // Mertensiella. 1999. Vol. 11. P. 317–366.
Team B. What Kinds of Lizards Are Nocturnal? URL: https://www.cuteness.com/article/kinds-lizards-nocturnal (дата обращения: 01.10.2021).
Trembath D. Nocturnal activity by Gould's Monitor (Varanus gouldii) at Town Common Environmental Park, Townsville Queensland // Herpetofauna (Sydney). 2000. Vol. 30, No 2. P. 52–53.
Yong D. L., Fam S. D., Ng J. J. Rediscovery of Dumeril's monitor Varanus dumerilii (Varanidae) in Singapore // Nature in Singapore. 2008. Vol. 1. P. 21–22.
Vidan E., Roll U., Bauer A., Grismer L., Guo P., Maza E., Novosolov M., Sindaco R., Wagner P., Belmaker J., Meiri S. The Eurasian hot nightlife: Environmental forces associated with nocturnality in lizards // Global Ecology and Biogeography. 2017. Vol. 26. P. 1316–1325.
Wilson S., Knowles D. G. Australia’s Reptiles: A Photographic Reference to the Terrestrial Reptiles of Australia. Sydney: Collins, 1988. 447 p.
Благодарности
Выражаем искреннюю благодарность заведующей кафедрой зоологии и физиологии Дагестанского государственного университета Л. Ф. Мазанаевой и заместителю директора государственного заповедника «Дагестанский» Г. С. Джамирзоеву за возможность организовать и провести подробные исследования, а также Д. В. Вето, В. В. Кожаре, А. А. Корнеевой, А. В. Ложкиной, А. Д. Мироненко-Маренкову, Е. Д. Рябовой, М. А. Тищенко, М. А. Туровой, А. Ю. Фраерман, А. А. Яцук, Р. М. Магомедовой, С. З. Алиевой и А. М. Магомедовой за помощь и участие в работах. Кроме того, благодарим всех тех, кто поделился своими воспоминаниями и материалами, касающимися активности ушастых круглоголовок в разных регионах: Н. А. Литвинова, Г. В. Полынову, Е. А. Дунаева, Э. В. Вашетко, Ф. Мирсалихову, А. Ю. Целлариуса, Ю. Г. Меньшикова, И. Ю. Барсукова, С. Б. Пугача, А. П. Лисачева, Д. А. Бондаренко.

 © 2011 - 2026
© 2011 - 2026