



Введение
Очень редко удается открыть одновременно несколько тайн природы одним и тем же ключом. Здание нашего несколько искусственно созданного благополучия слишком легко может рухнуть, как только в один прекрасный день окажется, что при помощи нескольких магических слов, таких, как «информация», «энтропия», «избыточность», нельзя решить всех нерешенных проблем.
Клод Шеннон (1963, с. 667–668).
В будущем наука будет концентрироваться больше вокруг проблем организации, структуры, языка, информации программирования и управления и меньше – вокруг проблем силы, движения, вещества, реакции, работы и энергии.
Джон фон Нейман (цит. по: Флейшман, 1982, с. 6).
Общепринято, что связь компонентов экосистемы (отдельных особей и популяций видов друг с другом и со средой) осуществляется через три взаимосвязанные, но качественно разные группы процессов (см., напр.: Розенберг и др., 1999, с. 102) – преобразование и перемещение вещества, расходование энергии в ходе совершения работы и ее накопление в связанной форме (второе начало термодинамики и закон сохранения энергии) и, наконец, упорядочение этих процессов, которое может рассматриваться как передача, накопление и перекодирование информации (закон убывания разнообразия и принцип обратной связи). Еще Винер в 1948 г. отмечал: «Информация есть информация, а не материя и не энергия. Тот материализм, который не признает этого, не может быть жизнеспособным в настоящее время» (Винер, 1958, с. 166). Хотя важность информационных процессов для реализации экологических функций (в контексте экосистемных услуг) сегодня широко признана (Margalef, 1968; Розенберг и др., 2002; Ulanowicz, 2002; Пузаченко, 2009; Ulanowicz et al., 2009; Ковалева, Яковлев, 2017; Walker, 2017 и др.), различные варианты теоретических конструкций в экологии явно недостаточно отражают центральную роль обработки информации в синтезе теоретической экологии.
Информация содержится в сложной организации и конфигурации потоков вещества и энергии, возникающих в результате взаимодействий совокупностей атомов, молекул, клеток или организмов (генетическая информация – программа свойств организма, получаемая от предков и заложенная в наследственных структурах в виде генетического кода; иными словами, «гены отличаются друг от друга чередованием пар нуклеотидов, и наследственная информация закодирована в виде последовательности нуклеотидов» (Инге-Вечтомов, 1989, с. 16)). Все эти структуры – их состав, форма и динамика – могут быть описаны с помощью некоторых информационных мер (это самый простой и очевидный способ использования ряда представлений теории информации в генетике, экологии и др.), но некоторые структуры, кроме того, представляют смысловой аспект информации, несут или кодируют семантическую информацию, которую можно интерпретировать и обрабатывать (Patten, 1959; Frey, 1966; Фрей, 1966; Гиляров, 1967; Пузаченко, 1982; Odum, 1988; Пузаченко Ю., Пузаченко А., 1996; Моничев, Гелашвили, 2001; Ulanowicz et al., 2006; Harte, 2011). «Обработка» информации в живых системах[1] влияет, в частности, на динамику популяции (Donaldson-Matasci et al., 2010; Fronhofer et al., 2017; Gil et al., 2018) и эволюционные процессы (Dall et al., 2005; Frank, 2008, 2012; Ulanowicz et al., 2009; Schmidt et al., 2010; Михайловский, 2013; Wagner, 2017) и является частью всех аспектов экологических изменений и взаимодействия человека с остальной биосферой (Commoner, 1971; Хефлинг, 1990; Воронцов, 1999; Моисеев, 2000; Павлов, Букварева, 2007 и мн. др.). Все это свидетельствует о том, что экологическая наука может даже выиграть, включив основы информационных теорий и обработки информации в строящееся «здание» теоретической экологии (Розенберг, 2013) для лучшего понимания и описания нашего изменяющегося мира (Wicken, 1987; Holt, 2007; Farnsworth, 2013; O'Connor et al., 2019).
Завершу это «Введение» словами отечественного специалиста по философским аспектам информатики Б. В. Бирюкова (1974, с. 243): «Информация налицо там, где имеется разнообразие, неоднородность (а биоразнообразие – основное свойство жизни. – Г. Р.). Информация "появляется" тогда, когда хотя бы два "элемента" в совокупности различаются, и она "исчезает", если объекты "склеиваются", "отождествляются"».
Аналитический обзор
Что такое информация?
Понятие «информация» происходит от лат. informatio, что в переводе обозначает сведения, разъяснение, ознакомление; «информация – сведения о лицах, предметах, фактах, событиях, явлениях и процессах независимо от формы их представления» (Глоссарий.., 2009, с. 66)[2]. В современном мире информация – это и ресурс, и одна из движущих сил развития человеческого общества[3]. Естественно, что информация имеет фундаментальное значение для жизни в целом и для живых систем разного уровня организации (от субклеточных до биосферы). В Орхусской конвенции о доступе к информации, участии общественности в процессе принятия решений и доступе к правосудию по вопросам, касающимся окружающей среды (заключена в г. Орхусе (Дания) 25.06.1988 г.), дано такое определение (ООН..., 2021; Оленина, 2018, с. 130):
«"Экологическая информация" означает любую информацию в письменной, аудиовизуальной, электронной или любой иной материальной форме:
1) о состоянии элементов окружающей среды, таких как воздух и атмосфера, вода, почва, земля, ландшафт и природные объекты, биологическое разнообразие и его компоненты, включая генетически измененные организмы, и взаимодействие между этими элементами;
2) факторах, таких как вещества, энергия, шум и излучение, а также деятельность или меры, включая административные меры, соглашения в области окружающей среды, политику, законодательство, планы и программы, оказывающие или способные оказать воздействие на элементы окружающей среды, охватываемые в подпункте 1 выше, и анализ затрат и результатов и другой экономический анализ и допущения, использованные при принятии решений по вопросам, касающимся окружающей среды;
3) состоянии здоровья и безопасности людей, условиях жизни людей, состоянии объектов культуры и зданий и сооружений в той степени, в какой на них воздействует или может воздействовать состояние элементов окружающей среды или, через посредство этих элементов, факторы, деятельность или меры, упомянутые в подпункте 2 выше».
Достижения в ряде областей науки (физика, кибернетика, генетика, молекулярная биология и др.) признают вещество, энергию и информацию в качестве «трех китов», на которых строятся живые системы (Шкловский, 1987; Dick, 1999; Бауместер и др., 2002; Frank, 2012; Davies, Walker, 2016; O'Connor et al., 2019; Малеина, 2020 и др.). При описании экологических принципов, называя все эти три группы процессов, все-таки большая роль приписывается круговороту вещества и энергии, а потоки информации анализируются заметно реже. Именно этим (сбои в обработке информации) можно объяснить, например, некоторые ошибки при прогнозировании динамики популяций гидробионтов при изменении окружающей среды (например, закисление океана (Моисеенко и др., 1999; Munday et al., 2009; Gasyukov, Kasatkina, 2013; Gordon et al., 2018) меняет способность организмов действовать соответствующим образом на основе экологической информации, такой как сигналы сезонных явлений, которые они используют для поиска пищи или партнеров). Более широкое признание роли, которую потоки информации играют, в частности, в динамике популяций, может привести к созданию экологической теории, учитывающей информационные эффекты, что, несомненно, будет способствовать углублению нашего понимания экологических последствий нарушения целостности информации (Бриллюэн, 1980; Schneider, Kay, 1994; Gil et al., 2018).
Еще один момент касается общебиологической и экологической роли информационных потоков для организации биологических полей межклеточных взаимодействий (Гурвич, 1944), стабильности сигнальных полей растений и животных (Уранов, 1965; Наумов, 1973, 1975; Мозговой, 1980; Крышень, 1998; Мозговой и др., 1998; Биологическое сигнальное поле..., 2013; Никольский, 2013; Рожнов, 2013), симбиозов (Douglas, 1994; Бухарин O. и др., 2007; Лысак, 2007; Babikova et al., 2013), социальных групп (Резникова, Рябко, 1988, 1990), информационных полей (Бухарин С. и др., 2009) и социо-, эколого-экономических систем разного масштаба (в частности, на оценке степени риска здоровью из-за снижения качества окружающей природной среды базируется вся система информационной связи человека с окружающим миром (Социо-, эколого-экономическая оценка..., 2013, c. 200)).
Прокомментирую только одну из процитированных выше статей – работу британских исследователей о сигналах через общие мицелиальные сети, которые «предупреждают» соседние растения о нападении тли (Babikova et al., 2013). Корни большинства наземных растений заселены микоризными грибами, которые обеспечивают минеральные питательные вещества в обмен на углерод. Авторы показывают, что микоризный мицелий также может действовать как проводник для передачи сигналов между растениями, как система раннего предупреждения о нападении травоядных. Травоядные насекомые вызывают системные изменения в производстве летучих веществ растений, особенно метилсалицилата (метиловый эфир салициловой кислоты), что делает бобовые растения Vicia faba репеллентом для тли, но привлекательными для врагов тлей (таких, как паразитоиды). Было показано, что эти эффекты могут возникать и у растений, свободных от тли, но только в том случае, если они связаны с зараженными тлей растениями через общую сеть микоризного мицелия. Эта подземная система «обмена сообщениями» позволяет соседним растениям активировать защиту от травоядных животных перед нападением. При этом общие микоризные мицелиальные сети могут определять результат мультитрофных взаимодействий, передавая информацию о нападении травоядных между растениями, тем самым влияя на поведение как травоядных, так и их естественных врагов.
Наконец, биоразнообразие – это одна из форм информации, это информация, хранящаяся в генах, морфологии, поведении, отражающих экологическую и эволюционную историю жизни организмов на Земле. Экология до сих пор не в состоянии полностью понять последствия потери этой информации (биоразнообразия) для ее считывания (обработки) и управления с ее помощью потоками вещества и энергии. Хотя некоторые концепции и методы оценки биоразнообразия неплохо разработаны (Мэгарран, 1992), они еще не встроены в структуры теоретической экологии (общие отношения между информацией и потоками вещества и энергии). Справедливости ради отмечу, что некоторые общетеоретические проблемы в свете теории информации (иерархичность, устойчивость, межвидовые взаимодействия, классификация экосистем и пр.) обсуждаются специалистами (Schneider, Kay, 1994; Ulanowicz et al., 2009; Coscieme et al., 2013; Sentis et al., 2015; Lee et al., 2016; Norton, Ulanowicz, 2017). Таким образом, хотя общепризнано, что биоразнообразие утрачивается (безвозвратно теряется ценная информация), у нас нет основных принципов, которыми можно было бы руководствоваться в понимании последствий этой угрозы. При этом антропогенная деятельность может разрушить целостность информационных сетей и их способность к адаптации еще до того, как они станут предметом изучения в рамках экологической науки (O'Connor et al., 2019, p. 2).
Измерение информации
Экологический аспект представлений об информации и энтропии весьма удачно преподнесен в статье (Моничев, Гелашвили, 2001) и обсуждался нами ранее (Розенберг, 2010; Шитиков и др., 2011, с. 73–81). Не буду описывать историю этих понятий (от статьи Клаузиуса (Rudolf Julius Emanuel Clausius, 1822–1888)), в которой он предложил модификацию второго закона термодинамики через функцию энтропии и которая была опубликована в Лейпциге, в журнале «Annalen der Physik und Chemie» (Clausius, 1854), через физическое осмысление и модификации Больцмана (Ludwig Eduard Boltzmann, 1844–1906), Планка (Max Karl Ernst Ludwig Planck, 1858–1947), кибернетические представления Шеннона (Claude Elwood Shannon, 1916–2001), Бриллюэна (Léon Nicolas Brillouin, 1889–1969), к общебиологическим (Волькенштейн, 1986) и экологическим интерпретациям Макартура (Robert Helmer MacArthur, 1930–1972), Маргалефа (Ramon Margalef i López, 1919–2004) и др.). Отмечу только, что Макартур предложил использовать индекс Шеннона для оценки энтропии (S, в битах) дискретного множества частот p1, …, pn (для измерения неопределенности (выбора) в потоках):
S = -∑pi log2pi.
При этом Макартур не делал никаких попыток заставить свои формализмы соответствовать категориям теории информации. Однако он продемонстрировал свою необычайную проницательность (MacArthur, 1955), когда обсудил ограничения только что предложенного индекса S. Он отметил, что стабильность зависит от видов, которые имеют рацион со многими степенями свободы; ограниченная диета имеет тенденцию к снижению стабильности. Но ограниченная диета важна для эффективности, а эффективность и стабильность необходимы для выживания в естественных условиях. То есть эффективность и стабильность являются фактически антагонистическими, но одновременно необходимыми свойствами устойчивых систем. Но и это не всё. В то время Макартуру казалось, что уравнение для оценки S (через потоки в трофической сети) не включает в себя все атрибуты, необходимые для выживания. После этого он прекратил споры, и его многообещающее преимущество оставалось практически невостребованным в течение следующих 20 лет.
На шесть лет раньше Макартура был прецедент измерения разнообразия сообществ с точки зрения размеров и численности популяций с использованием более традиционной математики (Simpson, 1949; Розенберг, 2007). Таким образом, исследователи после Макартура решили применить формулу Шеннона не к системным потокам, а к каждой конкретной популяции. Так, Маргалеф (Margalef, 1957) выбрал в качестве pi не долю общей активности, а, скорее, долю, которую составляет популяция от общего числа организмов или их общей биомассы; он установил pi = ni / N, где ni – количество организмов в популяции i, а N = ∑i ni.
Еще раз напомню (Розенберг, 2010), что под разнообразием (однородностью, выравненностью) обычно понимают видовое богатство и то свойство, которое отличает, например, сообщество из 12 особей трех видов с распределением (4, 4, 4) от сообщества (10, 1, 1), – выравненность. Стремление представить характер такого распределения, заданного большим набором численностей, в компактном виде одним числом приводит к обобщенным индексам, форма выражения которых, начиная с работ Макартура и Маргалефа, связывается с мерами количества информации.
Некоторые авторы трактуют энтропию еще более широко, считая ее мерой стохастичности и изменчивости, а информацию – мерой упорядоченности и детерминизма (см., напр.: Блюменфельд, 1996). «В такой трактовке энтропия – это то, что руководит эволюцией животных, растительных видов и экосистем в целом. Энтропийность рассматривается как неотъемлемое качество всех организмов, проявляющееся на уровне генов, межклеточных связей, внутривидового и межвидового взаимодействия. Энтропия считается необходимой природе, но и опасной в крайних ее проявлениях. Так, чрезмерная детерминация приводит к тому, что системы лишаются возможности адаптироваться в изменяющихся условиях, а "перебор" энтропии вызывает хаос и ералаш, потерю вектора эволюции» (Моничев, Гелашвили, 2001, с. 54).
Факт изменения видового разнообразия экосистем, находящихся в градиенте воздействия какого-либо фактора, достаточно тривиален. Менее очевидным является вопрос об оценках разнообразия в структурном и информационном аспектах. Применение с этой целью индекса Шеннона не снимает необходимости рассматривать разнообразие в пространственно-временном континууме. И здесь неоценимую помощь оказывает фрактальная геометрия – одна из молодых (менее 50 лет), но очень быстро развивающихся областей современной математики, которая связана с именем Мандельброта (2002). В контексте данной работы меня будет интересовать лишь один аспект применения «фрактальной идеологии» в экологических исследованиях – это фрактальный анализ видового разнообразия (Гелашвили и др., 2013), и даже еще более частный вопрос – измерение разнообразия с помощью мультифрактального спектра (Иудин и др., 2003; Гелашвили и др., 2007).
Одной из особенностей мультифрактального представления видовой структуры сообщества оказывается то, что большая часть известных в экологии индексов разнообразия (Шеннона, Маргалефа, Симпсона, Менхайника, Животовского и др.) «укладываются» на кривую функции мультифрактального спектра. Преимущество этой функции состоит в том, что ее значения представляют собой размерности неких однородных фрактальных подмножеств из исходного множества, которые дают доминирующий вклад для конкретных моментов распределения. Находит свое место на этой кривой и информационный индекс Шеннона – между индексом Симпсона (индексом доминирования) и индексом Маргалефа (максимальное значение функции мультифрактального спектра), иными словами, больше реагирует на наличие доминантных видов, чем учитывает редкие.
На рисунке в качестве примера представлена изменчивость мультифрактальных спектров видовой структуры сообществ зоопланктона Чебоксарского водохранилища в районе городов Нижний Новгород и Чебоксары (фактически влияние разных водных масс рек Волга и Ока (Шурганова и др., 2005)). Фрактальный анализ (индекс и спектр сингулярностей – это характеристики режима хаотических флуктуаций наблюдаемой величины (Иудин и др., 2003)) позволяет легко убедиться в том, что при слиянии рек Ока и Волга сообщества зоопланктона структурно заметно различаются, а около г. Чебоксары водные массы перемешиваются и сходство сообществ возрастает. Здесь же видно, что оценка только по индексу Шеннона (как и по индексу Симпсона) этих различий не вскрывает.
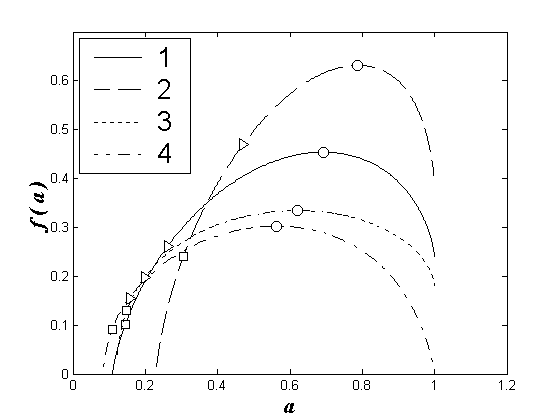
Пространственная динамика мультифрактальных спектров видовой структуры зоопланктонных сообществ Чебоксарского водохранилища для июля 2002 г. По оси абсцисс: a – индекс сингулярности; по оси ординат: f(a) – спектр сингулярностей. Номера графиков отвечают взятию проб: 1 – Нижний Новгород, левобережье (Волга), 2 – Нижний Новгород, правобережье (Ока), 3 – Чебоксары, левобережье, 4 – Чебоксары, правобережье. Условные обозначения фрактальных размерностей соответствуют: Ο – видовому разнообразию по Маргалефу; Δ – видовому разнообразию по Шеннону; Π – видовому разнообразию по Симпсону
Spatial dynamics of multifractal spectra of the species structure of zooplankton communities of the Cheboksary reservoir for July 2002. On the abscissa: a – singularity index; on the ordinate: f(a) is the spectrum of singularities. Graph numbers correspond to sampling: 1 – Nizhny Novgorod, left bank (Volga), 2 – Nizhny Novgorod, right bank (Oka), 3 – Cheboksary, left bank, 4 – Cheboksary, right bank. Symbols of fractal dimensions correspond to: Ο – species diversity according to Margalef; Δ – species diversity according to Shannon; Π – species diversity according to Simpson
Таким образом, фрактальные формализмы позволяют уйти от вычисления отдельных индексов разнообразия и наглядно представить структуру сообщества в виде мультифрактального спектра, что заставляет по-иному взглянуть на роль теории информации в измерении (в частности, по Шеннону) биоразнообразия.
Смысл информации (биосемиотика)
Определение количества информации, которое передается и воспринимается объектами экосистемы (допустим, что мы некоторым образом смогли определить процессы «передачи» и «приема» информации в сообществе), еще не дает полного представления о характере взаимодействия этих объектов. Действительно, два воздействия АВ и ВА несут одинаковое количество информации (при условии независимости этих воздействий), оцениваемое некоторым энтропийным индексом, но могут быть причиной различного поведения системы, находящейся под воздействием. Таким образом, важное значение имеет не только количественная оценка информации, но и ее качественная составляющая (в данном случае – порядок воздействия).
Информация структурно неоднородна и включает в себя синтаксическую (форма), семантическую (значение) и прагматическую (ценность) составляющие, описываемые тремя соответствующими разделами семиотики – общей теории знаковых систем (Kull, 1999, 2001; Emmeche, Kull, 2011; Князева, 2018). Биосемиотика как раздел семиотики исследует свойства знаков и знаковых систем (знаковые процессы) в живых системах. Различают фитосемиотику (изучает знаковые процессы у растений (Krampen, 1981; Kull, 2000)) и зоосемиотику (изучает биокоммуникацию животных – передачу информации от одной особи к другой – с позиции содержания их коммуникативных действий (Sebeok, 1968; Наумов, 1973; Мозговой, Розенберг, 1992; Мозговой и др., 1998; Turovski, 2002; Никольский, 2003, 2013; Kull, 2014)).
А. Д. Арманд (1975) построил интересный гипотетический пример физико-географической системы (снежный покров, залегающий на склоне), который позволил ему продемонстрировать качественное различие информационных воздействий на нее. В нашей работе (Розенберг и др., 1999, с. 105–107) рассмотрен теоретический пример конкурентного взаимодействия четырех видов в растительном сообществе (А1 > A2 ≈ A3 > A4; А1 – самый конкурентоспособный вид; А2 предпочитает холодные и влажные условия среды; A3 – теплые и сухие), взаимодействие анализируется в два временных интервала (весна – лето), виды находятся под воздействием двух факторов: температура воздуха («энергия», Т – тепло, Х – холодно) и осадки («вещество», С – сухо, В – влажно). Было показано, что состояние выхода системы (доминирование того или иного вида летом; см. далее) в большей степени зависит не от количественных значений состояний входов (если считать состояния Т, Х, С, В независимыми, то количество информации в рассмотренных далее четырех случаях одинаково), а от их последовательности, т. е. организация растительного сообщества в данном случае определяется не только потоком вещества (осадки) и энергии (температура), но и информационным процессом, заключенным в последовательности воздействия этих факторов.
ТС (весной) – ХВ (летом) → А3 – А2 → содоминирование А2 и А3
ХС (весной) – ТВ (летом) → А4 – А1 → доминирование А1
ТВ (весной) – ХС (летом) → А1 – А4 → доминирование А1
ХВ (весной) – ТС (летом) → А2 – А3 → содоминирование А2 и А3
Таким образом, количественные и качественные характеристики экосистем нельзя описать и прогнозировать, основываясь только на энергетическом и вещественном взаимодействии объектов. Еще раз подчеркну, что формальное определение количества информации, содержащейся в том или ином сообществе, без качественного анализа или построения дедуктивных моделей информационных процессов вряд ли может рассматриваться как полнокровное приложение теории информации к экологическим исследованиям.
В 70-х гг. прошлого столетия Н. П. Наумов (1973, 1975, 1977 и др.) предложил концепцию биологического сигнального поля млекопитающих, что позволило, к примеру, глубже понять механизм динамики численности популяций. Чуть позже были предприняты попытки «одеть» эту концепцию в количественную форму (Мозговой, Розенберг, 1992; Мозговой и др., 1998; Владимирова, 2002; Мозговой, Владимирова, 2002; Мозговой, 2005). В частности, для измерения биологического сигнального поля млекопитающих был предложен метод информационно-знакового поля и использованы параметры анизотропность (количество всех объектов и событий внешней среды, на которые реагирует исследуемая особь, приведенное к определенной протяженности следовой дорожки), напряженность (общее количество дискретных двигательных реакций, реализованных особью во время прохождения дистанции определенной протяженности) и величина этого поля (количество различных функциональных классов объектов, включенных особью в сферу своей активности, приведенное к определенной протяженности следовой дорожки). Таким образом, анизотропность – это показатель экстенсивности двигательного взаимодействия животных со средой обитания, который указывает на значение информации, полученной особью; напряженность – это показатель интенсивности взаимодействия особи с внешней средой, который указывает на суммарную ценность полученной ею информации; величина знакового поля указывает на особенности формального выражения информации, полученной исследуемой особью (табл. 1).
Таблица 1. Параметры информационно-знаковых полей млекопитающих, вытропленных
в пойменных лесах Самарской области в 1978–1982 гг. (Мозговой, 2005)
| Показатель | Виды | |||
| лисица | куница | горностай | ласка | |
| Количество вытропленных особей | 7 | 14 | 2 | 3 |
| Длина следовой дорожки, м | 37820 | 82160 | 1330 | 1000 |
| Величина поля (на эквив. дистанцию) | 26.4 | 20.5 | 19.0 | 12.0 |
| Анизотропность поля (на 1000 м следов) | 44 | 107 | 235 | 196 |
| Напряженность поля (на 1000 м следов) | 111 | 289 | 468 | 695 |
| Эквивалентная дистанция, м | 900.9 | 346.0 | 213.7 | 143.9 |
В частности, было показано, что обитающие на одной территории виды млекопитающих образуют коинформативный комплекс и реагируют на определенную группу внешних объектов, которые для видов этого комплекса более значимы, чем объекты, специфичные для каждого из видов (Владимирова, Мозговой, 2006).
Наиболее удобным приемом статистической обработки параметров информационных полей является дисперсионный анализ. Построение графиков изменения наблюдаемого признака (величина, анизотропность, напряженность поля и пр.) для отдельных видов, в зависимости от изменения выбранного для ординации фактора, сопровождается расчетом некоторых статистических характеристик (средневзвешенная напряженность фактора, сила влияния фактора и пр.). Результаты однофакторного дисперсионного анализа изменчивости параметров информационно-знаковых полей лисицы (Vulpes vulpes L.) и куницы (Martes martes L.) приведены в качестве примера в табл. 2 (Мозговой, Розенберг, 1992).
Таблица 2. Некоторые параметры информационно-знаковых полей лисицы и куньих,
биотопы Южного Урала и Самарской области, 1978–1992 гг.
| Объект | Фактор | Параметры информационно-знакового поля | Сила влияния фактора, % | Частные средние по градациям | |
| Лисица (на 500 м) | Пол (для взрослых особей) | самцы | самки | ||
| Величина | – | 18 | 19 | ||
| Напряженность | 8.7 | 89 | 133 | ||
| Анизотропность | – | 36 | 39 | ||
| Лисица (на 500 м) | Возраст (самцы) | взрослые | молодые | ||
| Величина | 2.2* | 18 | 20 | ||
| Напряженность | 5.4 | 89 | 119 | ||
| Анизотропность | – | 36 | 40 | ||
| Куница (на 26 объектов) | Возраст (самки) | ||||
| Величина | – | 400 | 395 | ||
| Напряженность | 2.0* | 281 | 311 | ||
| Анизотропность | – | 43 | 82 | ||
Примечание. Звездочкой (*) отмечены значения силы влияния фактора, достоверные на 80 % уровне значимости, остальные – на 95 %, прочерк – недостоверное влияние.
Данный пример (см. табл. 2) подтверждает тот факт, что с возрастом все большее значение для животных приобретает постоянство среды обитания – напряженность информационно-знакового поля уменьшается; таким образом, величина знакового поля, отражающая разнообразие объектов среды, на которые особи реагируют, может выступать, например, мерой постоянства внешней среды. Величина и анизотропность поля свидетельствуют о разнообразии объектов среды, вовлекаемых животным в ходе активности. Анизотропность информационного поля – показатель неравнозначности тех или иных объектов и событий среды для животного; чем больше этот показатель, тем выше адекватность особи данной среде (в ее информационном проявлении; в нашем примере – анизотропность выше для куниц). Если одновременно и величина поля большая, значит, сигнально-информационная среда максимально знакома особи и поведение ее почти стереотипно в этой среде.
Таким образом, метод информационно-знакового поля позволяет конструктивно рассматривать экосистему (с млекопитающими) в качестве информационной системы, оценивать степень сходства параметров таких полей (в пространственном и временном аспекте), характеризовать «информационную нишу» (например, произведением разных объектов среды на частоту реагирования особей на них (Мозговой и др., 1998, с. 81–82)) и др.
Принципы, объединяющие теорию информации и экологию
В работе О’Коннор с соавторами (O'Connor et al., 2019, р. 3) сформулировано пять принципов, которые «наводят мосты» между теорией информации и теоретической экологией. Перечислю их:
- Информация – это фундаментальная характеристика живых систем, а значит, и всех экологических систем (синтаксическая информация и семиотическая информация составляют две основные формы информации, каждая из которых важна для структуры и функционирования живых систем, от молекулярных систем до биосферы);
- Синтаксическая и семиотическая информация взаимодействует посредством обратной связи с энергетическими процессами и материальными циклами, чтобы влиять на структуру, функции и организацию в экосистемах;
- Обработка информации требует вещества и энергии, поэтому их недостаток, а также термодинамические ограничения могут ограничивать обработку информации;
- Обработка информации позволяет компонентам живых систем измерять окружающую среду и свое собственное состояние, а также измерять взаимосвязь между их состоянием и прошлой и ожидаемой средой;
- Наконец, системы обработки информации связаны внутри экосистемы и между экосистемами масштабами биологической организации (в частности, сильные положительные обратные связи в обработке информации могут определять или усиливать уровни организации – от клетки через особь до экосистемы и биосферы).
В этом контексте (пять принципов (O'Connor et al., 2019)) интересна статья С. Шайнера и М. Виллига (Scheiner, Willig, 2008; Розенберг, 2021), в которой сформулированы семь принципов построения теоретической экологии. Так, принципы 3–5 (обработка информации) соответствуют принципам 2 (организмы взаимодействуют со своей абиотической и биотической средой), 4 (условия окружающей среды неоднородны в пространстве и времени) и 5 (ресурсы конечны и неоднородны в пространстве и времени); принцип 2 вполне можно соотнести с «теоретическим» принципом 3 (распределение организмов и их взаимодействия зависят от непредвиденных обстоятельств; именно в этих случаях дополнительная информация необходима). А вот первый принцип и шестой «теоретический» (все организмы смертны) в обоих концепциях декларативны. Таким образом, можно констатировать, во-первых, что «общая теория экологии» мало в чем отличается от принятой мною (Розенберг и др., 1999; Розенберг, 2013) концепции создания теорий (основание [блоки составляющих теорий] – ядро [составляющие теории] – вершина [общая теория экологии]). Несомненно, семь фундаментальных принципов общей теории экологии (Scheiner, Willig, 2008) – это смело, достаточно спорно, но интригующе. Таким образом, в полном соответствии с героем комедии Мольера «Мещанин во дворянстве», мы все «говорим прозой», и наши представления о путях синтеза теоретической экологии сходны. А во-вторых, нетрудно видеть, что эти принципы носят сугубо описательный (зачастую констатирующий) характер и являются, скорее, «добрыми пожеланиями», чем строго формализованными положениями. Попробуем подойти к решению этой проблемы с другой стороны.
В 60–80-х гг. прошлого столетия Б. С. Флейшман (1971, 1982 и др.) развил новое направление в системологии – теорию потенциальной эффективности сложных систем (ТПЭСС), «основной особенностью которой является изучение не того, что есть, а того, что может быть» (Флейшман, 1971, с. 4). ТПЭСС содержит системную интерпретацию известных результатов теории информации Шеннона, теории игр фон Неймана, теории надежности, теории случайных процессов и др. С конца 1970-х гг. ТПЭСС стала применяться в экологии (Флейшман, 1977, 1982; Розенберг и др., 2002; Розенберг, 2003, 2013).
Общая форма предельного закона в системологии имеет следующий вид. Пусть имеется система А, которая взаимодействует со средой В путем (u, v)-обмена, где u и v – некоторые ресурсы, причем u – ресурсы системы А, которые она «платит» («обменивает») системе В за получение от нее некоторых ресурсов v. Эффективность системы всегда ограничивается предельно выгодным для нее (u, v0)-обменом, когда за данное u она получает максимальное v0 (или при фиксированном v0 она тратит предельно малое u (Флейшман, 1982, с. 42)). Таким образом, целью системы А является максимально выгодный (u, v0)-обмен. Чаще всего взаимодействие системы А со средой В носит случайный характер, что заставляет говорить о вероятности P(u, v0), т. е. о вероятности максимально выгодного (u, v0)-обмена, которая и называется потенциальной эффективностью системы. Как было показано Б. С. Флейшманом (1971), эта вероятность описывается асимптотическим соотношением
P(u, v) = либо 0 (при v > v0), либо 1 (при v < v0).
Б. С. Флейшман (1982, с. 29) в порядке возрастания сложности системы обозначил следующий порядок ее качеств: устойчивость (stability, reliability; SR-качество), помехоустойчивость (достигается через информированность системы о среде; information; I-качество), управляемость (controllability; C-качество) и самоорганизация, обучаемость (learning; L-качество). Для экосистем можно добавить и такое качество, как биоразнообразие (biodiversity; В-качество). Рассмотрим их потенциальную эффективность применительно к экосистемам (табл. 3).
Таблица 3. Предельные законы и их интерпретация
| Качество | Формула оптимального (u, v)-обмена |
Интерпретация |
| SR | Оптимальный (n, t)-обмен будет при t0 = exp(Cn · n), где Cn – некоторая фундаментальная величина, зависящая от вероятностей рi – гибели элементов i системы А
|
Экосистема осуществляет со средой оптимальный (n, t)-обмен: система приобретает у среды время жизни t, откупаясь гибелью n своих элементов. Система А будет существовать неограниченно долго, если число ее элементов будет расти экспоненциально со временем. Популяции на ранних стадиях своего развития растут по экспоненте, в дальнейшем среда ограничивает их рост (например, по логистической кривой) |
| I | Оптимальный (t, М)-обмен будет при М0 = exp(CI · t), где Cn – некоторая фундаментальная величина, зависящая от вероятностей рi – пропускная способность системы А в терминологии теории информации
|
Экосистема осуществляет со средой оптимальный (t, М)-обмен: экосистема приобретает у среды некоторое количество сигналов М, платя за это временем t своего существования. Это можно наблюдать в случае коллективного поведения сложной системы (например, стайное поведение при поиске агрегированной пищи (Margalef, 1961; Patten, 1963)). Естественно, I-качество будет широко представлено в «экосистемах с человеком» (социо-, эколого-экономических системах, СЭЭС). Кроме того, ТПЭСС в варианте I-закона по-новому ставит вопрос об интерпретации экосистемы как некоего текста, закодированного с целью повышения помехоустойчивости в «шумящей среде» (Левич, 1977, 1978, 1983; Адильбаев, 2017) |
| С | Оптимальный (ṯ, K0)-обмен будет при K0 = CC ṯ, где CC – некоторая фундаментальная величина, зависящая от функции выигрыша, определяемой вероятностями выигрыша каждой из систем А и В и количеством ресурсов, получаемой системой А и проигрываемой системой В (Флейшман, 1982, с. 214) | Процесс взаимодействия систем А и В состоит в следующем. Системы А и В ведут борьбу за ограниченное количество ресурсов K. Борьба проходит отдельными схватками в последовательные дискретные моменты времени. Борьба не ограничивается во времени и ведется до полного захвата ресурсов одной из систем (соответственно, другая гибнет). Предполагается, что количество ресурсов, получаемое системой на каждом шаге взаимодействия, много меньше K. Фиксированное значение ресурса K может быть выиграно той или иной системой за случайное время t (среднее время ṯ). Можно считать, что имеет место (ṯ, K)-обмен: за количество ресурсов K система расплачивается временем своего существования ṯ. Фактически С-закон является обобщением принципов конкурентного исключения Г. Ф. Гаузе |
| L | Оптимальный (t, М)-обмен (система А приобретает у среды В некоторое количество сигналов М, платя за это временем t своего существования) будет при М0 = exp(CI · t), где Cn – некоторая фундаментальная величина, зависящая от вероятностей рi – пропускная способность системы А в терминологии теории информации
|
Обучение – целенаправленный процесс организации и стимулирования активной познавательной деятельности. Пусть некая популяция животных (система А) взаимодействует со средой В путем изучения информационно-знаковых полей (см. выше). Популяция в каждый дискретный момент времени ti оценивает параметры информационного поля – анизотропность (АТ), напряженность (In) и величину (V). Далее – все, как для описания I-качества, с той только разницей, что
M = M1 + M2 + M3 = MAT + MIn + MV и вероятность P(M) оптимального (t, М)-обмена оценивается с использованием неравенства Буля: 1 – 3 · max{1 – P(Mi)} ≤ P(M) ≤ min P(Mi). Это неравенство позволяет оценить вероятность пересечения зависимых событий, не вдаваясь в сложную природу их зависимости; верхняя оценка может интерпретироваться как экологический закон Либиха |
| В | Оптимальный (ni, mi)-обмен (ответ на вопрос: какое максимальное количество видов mi получит система А при фиксированном количестве видов ni0, которыми она готова пожертвовать ради новых видов?) будет при p (n + m)/m < 1, где все pk = p, nk = n и mk = m; в этом случае Ni будет стремиться к K. Точнее, к величине, близкой к 0.75K (максимальная плотность упаковки равных сфер (Conway, Sloane, 1993))
|
Экосистема осуществляет со средой обмен следующим образом. В каждый дискретный момент времени ti из системы А (в t0 состоит из N0 видов) в систему В (в t0 состоит из M0 видов) переходит ni видов с вероятностью pi, а из системы В в систему А – mi видов с вероятностью (1 – pi). Таким образом, в каждый дискретный момент времени ti имеет место (ni, mi)-обмен. При этом Ni = N0 - Σnkpk + Σmk (1 - pk). Если Ni подчиняется логистическому закону (биоразнообразие не может расти бесконечно), то где r – скорость роста числа видов, K – теоретически предельное биоразнообразие для данной территории (можно говорить и о пространственном ресурсе). Используя принцип плотной упаковки как экстремальный принцип для функции логистического роста, было показано, что «сообщество конкурирующих за жизненное пространство видов эволюционирует к состоянию с максимально плотной упаковкой, причем в процессе эволюции плотность упаковки сообщества всегда возрастает, достигая в равновесном состоянии максимально возможного для данной среды значения» (MacArthur, 1969, 1970; Свирежев, Логофет, 1978, с. 273).
|
Примечание. Первые три примера (SR-, I- и С-качества) рассматривались Б. С. Флейшманом (1982); примеры L- и В-качеств – предложены мною.
Рассмотренные в табл. 3 «одномерные» Х-качества системы А дают оценки ее структуры и поведения через модели оптимальных (u, v)-обменов. Возможно рассмотрение «двумерных» XY-качеств (в частности, В. Ф. Крапивин (1978) и Б. С. Флейшман (1982, с. 217–223) рассматривают (SR; С)-модель как особый вариант активной устойчивости системы со специальным названием живучесть). Более подробно примеры использования теории потенциальной эффективности сложных систем в информатике рассмотрены в работах (Флейшман, 1971, 1982; Fleishman, 1976), в математической экологии – (Флейшман, 1977; Крапивин, 1978; Розенберг, 2003, 2013).
Заключение
В последние годы все чаще стали появляться работы, в которых на полном серьезе обсуждаются основы нового научного направления – «информационной экологии» (Davenport, Prusak, 1997; Ивченко, Мартыщенко, 1998; Мизинцева и др., 2000 и др.). Мне представляется это излишним по целому ряду причин (Розенберг, 2011), среди которых есть и такой аргумент. «Информационный подход» при изучении тех или иных свойств экосистем должен восприниматься только как метод (если вы выполнили исследование по изучению микроорганизмов с помощью микроскопа, то вам не придет в голову защищать эту работу по физике (оптике), скорее, вы будете защищаться по микробиологии. Ситуация чем-то напоминает попытки «захватить» в Интернете звучный домен, чтобы, если повезет, сделать его предметом торга). Таким образом, теория информации (как и математическое моделирование, автоматическое управление и пр.) позволяет увидеть некоторые аналогии в описании экосистем и применить (с четким пониманием возможных ограничений) соответствующий аппарат, а это не есть новая наука.
То же самое можно сказать и в адрес формирующейся «экологической информатики». Вопрос о необходимости «экологической информатики» может быть переформулирован следующим образом: «А надо ли это информатике?» или «Что нового это может дать общим закономерностям передачи информации?». Теория информации в экологических исследованиях пока сведена только к измерению информации с помощью энтропийного показателя. Но теория информации – это не теория измерения информации (точнее, это далеко не только измерение информации); внимание экологов пока не привлекли основные предельные соотношения для систем передачи данных, которые и составляют ее суть. Таким образом, похоже, как и с «информационной экологией», «экологическая информатика» не вправе претендовать на новое и оригинальное направление. Естественный процесс взаимопроникновения различных наук здесь не набрал еще «критической массы».
На сегодняшний день представляется весьма вероятным, что изучение экологии будет вестись как минимум по двум отдельным направлениям – аут- и демэкология, с одной стороны, и синэкология, с другой, которые будут по-разному применять методологию теории информации (Ulanowicz, 2002). Теория информации «исповедует» реляционный подход (Bateson, 1972), согласно которому нельзя сказать, что люди, животные или другие организмы вступают в общение (информационный подход) или участвуют в нем (интеракциональный подход), поскольку они уже являются неотъемлемой частью этого процесса независимо от того, хотят они этого или нет, частью как локальной, так и глобальной системы взаимоотношений. Этот подход вступает в противоречие с методами аут- и демэкологических исследований (акцент на численность и состав популяции, а отношения с другими популяциями остаются второстепенными). Действительно, в такой ситуации многие информационные подходы теряют смысл (исключение – анализ информационных полей). Синэкология, напротив, делает главный упор на взаимоотношения популяций друг с другом, в этом случае теория информации может быть очень естественным образом применена к исследованию взаимосвязанных процессов.
Можно надеяться, что в результате методических успехов, а также опыта, полученного в других областях знания, «информатизация» экологии не потребует большого времени, как это может показаться. Однако представляется, что эта задача по своей трудности превосходит пределы любой индивидуально планируемой программы. Более того, процесс математизации внедрения информационной теории в экологию вовсе не является тривиальным (точнее сказать, он мало или совсем не используется до сих пор в этом процессе).
В заключение отмечу, что чувство неудовлетворенности математическими интерпретациями применения теории информации в экологии в значительной степени объясняется тем, что они часто дают (подтверждают) утверждения, которые не лучше, чем те же утверждения, высказанные в словесной форме. Здесь вполне подходят слова, сказанные полвека тому назад фон Нейманом и Моргенштерном (1970, с. 31) относительно использования теории игр в экономике: «...доказательства отсутствуют потому, что математический аппарат применяется к тем областям, которые настолько обширны и сложны, что еще в течение долгого времени – до тех пор, пока не будет накоплено больше эмпирических фактов, – едва ли можно ожидать серьезного прогресса от одного только увеличения дозы математики. Тот факт, что эти области атакуются таким путем, показывает только, что сопровождающие этот процесс трудности недооцениваются. В действительности эти трудности огромны, и мы не чувствуем себя достаточно подготовленными для их преодоления».
Подводя итог взаимовоздействию теоретической экологии и теории информации, повторюсь и еще раз подчеркну, что «информационный» подход применим в экологии лишь в рамках аналогий, которые могут навести мысль исследователей-экологов на формулирование новых подходов и направлений поиска новых законов и закономерностей. Причем успехов следует ожидать не в прямом использовании информационных мер для оценки биоразнообразия или сходства экосистем, а в глубоком проникновении и экологической интерпретации предельных кибернетических законов потенциальной эффективности сложных систем.
Завершить этот обзор я хочу словами Шеннона из краткой заметки «Бандвагон»: «Что можно сделать, чтобы внести в сложившуюся ситуацию ноту умеренности? Во-первых, представителям различных наук следует ясно понимать, что основные положения теории информации касаются очень специфического направления исследования, направления, которое совершенно не обязательно должно оказаться плодотворным в психологии, экономике и в других социальных науках (к этому списку можно добавить биологию и экологию. – Г. Р.). <…> поиск путей применения теории информации в других областях не сводится к тривиальному переносу терминов из одной области науки в другую. Этот поиск осуществляется в длительном процессе выдвижения новых гипотез и их экспериментальной проверки. <…> Во-вторых, мы должны поддерживать образцовый порядок в своем собственном доме. На понятия теории информации очень большой, даже, может быть, слишком большой спрос. <…> Только последовательно придерживаясь строго научной линии, мы сможем достичь реальных успехов в теории связи и укрепить свои позиции» (Шеннон, 1963, с. 668).
[1] «Не существует такой единицы жизни, которую можно было бы назвать "живой" безотносительно к внешней среде. <…> Биологи должны подчеркивать еще и еще раз, что жизнь – это неизбежное свойство, присущее экосистеме в целом, а не свойство изолированного скопления макромолекул. Мне кажется, что центральный вопрос происхождения жизни – это не вопрос о том, что возникло прежде, ДНК или белки, а вопрос о том, какова простейшая экосистема?» (Патти, 1970, с. 178).
[2] Общепринятого определения понятия «информация» нет (Тарасенко, 1963; Madden, 2000; Хургин, 2007; Пахомова, 2013; Эрнштейн, 2018 и др.). Это не вызывает удивления: одним из свойств, определяющих сложность системы (а биологические системы практически все – сложные), является невозможность дать четкое и однозначное определение такой системы (Флейшман, 1982; Розенберг, 2013). Н. Н. Моисеев (1995) также полагал, что универсального определения информации не только нет, но и быть не может из-за широты этого понятия.
[3] Недаром сегодня и в политике, и в СМИ можно услышать даже об информационных войнах.
Библиография
Адильбаев Т. Ш. Биосемиотика как наука: коды, знаки, логические конфликты и порождение смысла (сводный реферат) // МЕТОД: Московский ежегодник трудов из обществоведческих дисциплин. 2017. № 7. С. 190–203.
Арманд А. Д. Информационные модели природных комплексов . М.: Наука, 1975. 126 с.
Бауместер Д., Экерт А., Цайлингер А. Физика квантовой информации . М.: Постмаркет, 2002. 376 с.
Биологическое сигнальное поле млекопитающих / Под ред. А. А. Никольского, В. В. Рожнова. М.: Тов-во науч. изданий КМК, 2013. 323 с.
Бирюков Б. В. Кибернетика и методология науки . М.: Наука, 1974. 416 с.
Блюменфельд Л. А. Информация, термодинамика и конструкция биологических систем // Соросовский образовательный журнал. 1996. № 7. С. 88–92.
Бриллюэн Л. Термодинамика – кибернетика – жизнь // Кибернетика – неограниченные возможности и возможные ограничения: современное состояние. М.: Наука, 1980. С. 8–27.
Бухарин О. В., Лобакова Е. С., Немцева Н. В., Черкасов С. В. Ассоциативный симбиоз . Екатеринбург: УрО РАН, 2007. 264 с.
Бухарин С. Н., Ковалев В. И., Малков С. Ю. О формализации понятия информационного поля // Информационные войны. 2009. № 4 (12). С. 2–9.
Винер Н. Кибернетика, или управление и связь в животном и машине. М.: Сов. радио, 1958. 216 с. (Wiener N. Cybernetics: Or Control and Communication in the Animal and the Machine. Paris: Hermann & Cie; Cambridge; Massachusetts: M.I.T. Press, 1948. 194 p.).
Владимирова Э. Д. Коммуникативная составляющая информационных сигнальных полей лисицы // Вестник СамГУ. Естественнонаучная серия. 2002. № 4 (26). С. 145–156.
Владимирова Э. Д., Мозговой Д. П. Влияние следов жизнедеятельности лисицы обыкновенной на зимнюю экологию некоторых куньих // Вестник СамГУ. Естественнонаучная серия. 2002. № 6/1 (46). С. 220–234.
Волькенштейн М. В. Энтропия и информация . М.: Наука, 1986. 192 с.
Воронцов Н. Н. Экологические кризисы в истории человечества // Соросовский образовательный журнал. 1999. № 10. С. 2–10.
Гелашвили Д. Б., Иудин Д. И., Розенберг Г. С., Якимов В. Н. Степенной характер накопления видового богатства как проявление фрактальной структуры биоценоза // Журнал общей биологии. 2007. Т. 68, № 2. С. 115–124.
Гелашвили Д. Б., Иудин Д. И., Розенберг Г. С., Якимов В. Н., Солнцев Л. А. Фракталы и мультифракталы в биоэкологии . Н. Новгород: Изд-во Нижегород. гос. ун-та, 2013. 370 с.
Гиляров А. М. Информационная теория в экологии // Успехи современной биологии. 1967. Т. 64, № 4. С. 107–115.
Глоссарий по информационному обществу / Под общ. ред. Ю. Е. Хохлова. М.: Институт развития информационного общества, 2009. 160 с.
Гурвич А. Г. Теория биологического поля . М.: Сов. наука, 1944. 156 с.
Ивченко Б. П., Мартыщенко Л. А. Информационная экология. Ч. 1. Оценка риска техногенных аварий и катастроф. Статистическая интерпретация экологического мониторинга. Моделирование и прогнозирование экологических ситуаций . СПб.: Нордмед-Издат, 1998. 208 с.
Инге-Вечтомов С. Г. Генетика с основами селекции: Учебник для биол. спец. ун-тов . М.: Высш. шк., 1989. 591 с.
Иудин Д. И., Гелашвили Д. Б., Розенберг Г. С. Мультифрактальный анализ структуры биотических сообществ // Доклады РАН. 2003. Т. 389, № 2. С. 279–282.
Князева Е. Н. Биосемиотика: истоки междисциплинарного направления // Вопросы философии. 2018. № 11. С. 86–98.
Ковалева Е., Яковлев А. Экологические функции болотных экосистем (на примере острова Сахалин) // Экология и промышленность России. 2017. Т. 21, № 12. С. 32–37.
Крапивин В. Ф. О теории живучести сложных систем . М.: Наука, 1978. 248 с.
Крышень А. М. К методике изучения фитогенных полей деревьев // Ботанический журнал. 1998. Т. 83, № 10. С. 133–142.
Левич А. П. Понятие устойчивости в биологии (математические аспекты) // Человек и биосфера. Вып. 1. М.: МГУ, 1977. С. 138–174.
Левич А. П. Информация как структура систем // Семиотика и информатика. 1978. Вып. 10. С. 116–132.
Левич А. П. Семиотические структуры в экологии, или существует ли экологический код? // Человек и биосфера: экологический прогноз. Вып. 8. М.: МГУ, 1983. С. 68–77.
Лысак В. В. Микробиология: Учеб. пособие . Минск: БГУ, 2007. 430 с.
Малеина М. Н. Понятие и классификации геномной (генетической) информации // Lex Russica. 2020. Т. 73, № 7. С. 50–58.
Мандельброт Б. Фрактальная геометрия природы . М.: Институт компьютерных исследований, 2002. 656 с.
Мизинцева М. Ф., Королева Л. М., Бондарь В. В. Информационная экология . М.: Россельхозакадемия, 2000. 231 с.
Михайловский Г. Е. Энтропия, информация и эволюция // Библиотека электронных публикаций. 2013. URL: http://www.chronos.msu.ru/old/RREPORTS/michailovsky_entropy.htm.
Мозговой Д. П. Использование концепции информационного биологического поля в биогеоценотических исследованиях // Вопросы лесной биогеоценологии экологии и охраны природы в степной зоне: Межвед. сборник. Куйбышев: КГУ, 1980. С. 119–125.
Мозговой Д. П. Информационно-знаковые поля и поведение млекопитающих: теория и практика // Вестник СамГУ. Естественнонаучная сер. 2005. № 2. С. 238–249.
Мозговой Д. П., Владимирова Э. Д. Сигнальные поля и поведение животных в сигнально-информационной среде // Известия Самарского НЦ РАН. 2002. Т. 4, № 2. С. 207–215.
Мозговой Д. П., Розенберг Г. С. Сигнальное биологическое поле млекопитающих: теория и практика полевых исследований: Учебное пособие . Самара: СамГУ, 1992. 119 с.
Мозговой Д. П., Розенберг Г. С., Владимирова Э. Д. Информационные поля и поведение млекопитающих: Учебное пособие . Самара: Самарский ун-т, 1998. 92 с.
Моисеев Н. Н. Современный рационализм . М.: МГВП КОКС, 1995. 376 с.
Моисеев H. H. Системная организация биосферы и концепция коэволюции // Общественные науки и современность. 2000. № 2. С. 123–130.
Моисеенко Т. И., Шаров А. Н., Вандыш О. И., Яковлев В. А., Лукин A. A. Изменения биоразнообразия поверхностных вод Севера в условиях закисления, евтрофирования и токсичного загрязнения // Водные ресурсы. 1999. № 4. С. 492–501.
Моничев А. Я., Гелашвили Д. Б. Энтропия и информация: экологический аспект // Вестник Нижегородского университета им. Н. И. Лобачевского. Сер.: Биология. 2001. № 1. С. 52–59.
Мэгарран Э. Экологическое разнообразие и его измерение . М.: Мир, 1992. 184 с.
Наумов Н. П. Сигнальные (биологические) поля и их значения для животных // Журнал общей биологии. 1973. Т. 34, № 6. С. 808–817.
Наумов Н. П. Биологические (сигнальные) поля и их значение в жизни млекопитающих // Вестник АН СССР. 1975. № 2. С. 55–62.
Наумов Н. П. Биологические (сигнальные) поля и их значение в жизни млекопитающих // Успехи современной териологии. М.: Наука, 1977. С. 93–110.
Нейман Д. фон, Моргенштерн О. Теория игр и экономическое поведение . М.: Наука, 1970. 708 с.
Никольский А. А. Экологические аспекты концепции биологического сигнального поля млекопитающих // Зоологический журнал. 2003. Т. 82, вып. 4. C. 443–449.
Никольский А. А. Концепция биологического сигнального поля как раздел общей экологии // Биологическое сигнальное поле млекопитающих: Коллективная монография. М.: Тов-во науч. изданий КМК, 2013. С. 7–11.
Оленина Т. Ю. Понятие экологической информации в российском законодательстве // Legal Concept (Правовая парадигма). 2018. Т. 17, № 2. С. 126–133.
ООН. Конвенции и соглашения . 2021. URL: https://www.un.org/ru/documents/decl_conv/conventions/orhus.shtml (дата обращения: 10.09.2021).
Павлов Д. С., Букварева Е. Н. Биоразнообразие, экосистемные функции и жизнеобеспечение человечества // Вестник РАН. 2007. Т. 77, № 11. С. 974–998.
Патти Г. Замечания // На пути к теоретической биологии. Пролегомены. М.: Мир, 1970. С. 177–179.
Пахомова В. А. Понятие термина «информация» и его историческое развитие // Вестник ЮУрГУ. Сер.: Право. 2013. Т. 13, № 4. С. 59–64.
Пузаченко Ю. Г. Биологическое разнообразие, устойчивость и функционирование // Проблемы устойчивости биологических систем. М.: ИЭМЭЖ АН СССР, 1982. С. 5–32.
Пузаченко Ю. Г. Биологическое разнообразие в биосфере: системологический и семантический анализ // Биосфера. 2009. Т. 1, № 1. С. 25–38.
Пузаченко Ю. Г., Пузаченко А. Ю. Семантические аспекты биоразнообразия // Журнал общей биологии. 1996. Т. 57, № 1. С. 5–43.
Резникова Ж. И., Рябко Б. Я. Язык муравьев и теория информации // Природа. 1988. № 6. С. 65–70.
Резникова Ж. И., Рябко Б. Я. Теоретико-информационный анализ «языка» муравьев // Журнал общей биологии. 1990. Т. 51, № 5. С. 601–609.
Рожнов В. В. Место концепции опосредованной хемокоммуникации в теории биологического сигнального поля Н. П. Наумова // Биологическое сигнальное поле млекопитающих: Коллективная монография. М.: Тов-во науч. изданий КМК, 2013. С. 16–23.
Розенберг Г. С. О моделях потенциальной эффективности экологических систем // Известия Самар. НЦ РАН. Спец. вып. "Проблемы современной экологии". 2003. Вып. 1. С. 34–43.
Розенберг Г. С. Несколько слов об индексе разнообразия Симпсона // Самарская Лука: Бюллетень. 2007. Т. 16, № 3 (21). С. 581–584.
Розенберг Г. С. Информационный индекс и разнообразие: Больцман, Котельников, Шеннон, Уивер… // Самарская Лука: проблемы региональной и глобальной экологии: Бюллетень. 2010. Т. 19, № 2. С. 4–25.
Розенберг Г. С. Экология и кибернетика: по следам Маргалефа // Биосфера. 2011. Т. 3, № 4. С. 445–454.
Розенберг Г. С. Введение в теоретическую экологию : В 2 т. Изд. 2-е. Тольятти: Кассандра, 2013. Т. 1. 565 с.; Т. 2. 445 с.
Розенберг Г. С. Комментарий переводчика статьи об общей теории экологии // Самарская Лука: проблемы региональной и глобальной экологии: Бюллетень. 2021. Т. 30, № 4. С. 87–92.
Розенберг Г. С., Мозговой Д. П., Гелашвили Д. Б. Экология. Элементы теоретических конструкций современной экологии (Учебное пособие) . Самара: СамНЦ РАН, 1999. 396 с.
Розенберг Г. С., Мозговой Д. П., Гелашвили Д. Б. Информация и экология: некоторые мысли об информационных процессах в экосистемах // Вестник Волжского университета им. В. Н. Татищева. Серия «Экология». 2002. Вып. 2. С. 48–59.
Свирежев Ю. М., Логофет Д. О. Устойчивость биологических сообществ . М.: Наука, 1978. 352 с.
Социо-, эколого-экономическая оценка состояния территории / Под ред. С. В. Карелова, И. С. Белик. Екатеринбург: УрФУ, 2013. 258 с.
Тарасенко Ф. П. К определению понятия «информация» в кибернетике // Вопросы философии. 1963. № 3. С. 80.
Уранов А. А. Фитогенное поле // Проблемы современной ботаники. Т. 1. М.; Л.: Наука, 1965. С. 251–254.
Флейшман Б. С. Элементы теории потенциальной эффективности сложных систем . М.: Сов. радио, 1971. 224 с.
Флейшман Б. С. Стохастические модели сообществ // Океанология. Биология океана. Биологическая продуктивность океана. М.: Наука, 1977. Т. 2. С. 276–288.
Флейшман Б. С. Основы системологии . М.: Радио и связь, 1982. 368 с.
Фрей Т. Э.-А. Некоторые аспекты фитоценотической значимости вида в растительном сообществе // Ботанический журнал. 1966. Т. 51, № 8. С. 1073–1084.
Хефлинг Г. Тревога в 2000 году: Бомбы замедленного действия на нашей планете . М.: Мысль, 1990. 270 c.
Хургин В. М. Об определении понятия «информация» // Информационные ресурсы России. 2007. № 3 (97). C. 6–14.
Шеннон К. Бандвагон // Шеннон К. Работы по теории информации и кибернетике. М.: Иностр. лит., 1963. С. 667–668. (Shannon C. The Bandwagon // IRE Transactions on Information Theory. 1956. Vol. 2, No 1. P. 3.)
Шитиков В. К., Зинченко Т. Д., Розенберг Г. С. Макроэкология речных сообществ: концепции, методы, модели . Тольятти: Кассандра, 2011. 255 с.
Шкловский И. С. Вселенная, жизнь, разум . 6-е изд. М.: Наука, 1987. 320 с.
Шурганова Г. В., Иудин Д. И., Гелашвили Д. Б., Якимов В. Н. Мультифрактальный анализ видового разнообразия зоопланктоценозов Чебоксарского водохранилища // Актуальные проблемы рационального использования биологических ресурсов водохранилищ. Рыбинск: Рыбинский дом печати, 2005. С. 294–309.
Эвери Д. Теория информации и эволюция . М.; Ижевск: Регулярная и хаотическая динамика: Институт компьютерных исследований, 2019. 252 c.
Эрштейн Л. Б. Об определении понятия информации // Метафизика. 2018. № 3 (29). С. 21–30.
Babikova Z., Gilbert L., Bruce T. J. A., Birkett M., Caulfield J. C., Woodcock C., Pickett J. A., Johnson D. Underground signals carried through common mycelial networks warn neighboring plants of aphid attack // Ecol. Lett. 2013. Vol. 16. P. 835–843.
Bateson G. Steps to an Ecology of Mind: Collected Essays in Anthropology, Psychiatry, Evolution, and Epistemology. Chicago: Univ. Press, 1972. 542 p. (Бейтсон Г. Шаги в направлении экологии разума. 2-е изд. М.: УРСС, 2005. 232 c.)
Clausius R. Über eine veränderte Form des zweiten Hauptsatzes der mechanischen Wärmetheoriein // Annalen der Physik und Chemie. 1854. Vol. 93, No 12. P. 481–506.
Commoner B. The Closing Circle: Nature, Man, and Technology. N. Y.: Knopf., 1971. 326 p. (Коммонер Б. Замыкающийся круг. М.: Гидрометеоиздат, 1974. 280 с.).
Conway J. H., Sloane N. J. A. Sphere Packings, Lattices and Groups. 2nd ed. N. Y.: Springer-Verlag, 1993. 679 p.
Coscieme L., Pulselli F. M., Jørgensen S. E., Bastianoni S., Marchettini N. Thermodynamics-based categorization of ecosystems in a socio-ecological context // Ecol. Model. 2013. Vol. 258. P. 1–8.
Dall S. R., Giraldeau L. A., Olsson O., McNamara J. M., Stephens D. W. Information and its use by animals in evolutionary ecology // Trends Ecol. Evol. 2005. Vol. 20. P. 187–193.
Davenport T. H., Prusak L. Information Ecology: Mastering the Information and Knowledge Environment. N. Y.: Oxford Univ. Press, 1997. 272 p.
Davies P. C., Walker S. I. The hidden simplicity of biology // Rep. Prog. Phys. 2016. Vol. 79. P. 1–19.
Dick S. J. The Biological Universe: The Twentieth Century Extraterrestrial Life Debate and the Limits of Science. Cambridge: Univ. Press, 1999. 578 р.
Donaldson-Matasci M. C., Bergstrom C. T., Lachmann M. The fitness value of information // Oikos. 2010. Vol. 119. P. 219–230.
Douglas A. E. Symbiotic Interaction. Oxford et al.: Oxford Univ. Press, 1994. 148 p.
Emmeche C., Kull K. (Eds.). Towards a Semiotic Biology: Life is the Action of Signs. London: Imperial College Press, 2011. 304 p.
Farnsworth K. D. Living is information processing: from molecules to global systems // Acta Biotheor. 2013. Vol. 61. P. 203–222.
Fleishman B. S. Philosophy of systemology // Cybernetica. 1976. Vol. 19, No 4. P. 261–272.
Frank S. A. Natural selection maximizes Fisher information // J. Evol. Biol. 2008. Vol. 22. P. 231–244.
Frank S. A. Natural selection. V. How to read the fundamental equations of evolutionary change in terms of information theory // J. Evol. Biol. 2012. Vol. 25. P. 2377–2396.
Frey T., Võhandu L. Uus meetod klassifikatsiooniühikute püstitamiseks // ENSV TA Toimet. Biol. Seeria. 1966. Nr. 4. Lk. 565–576.
Fronhofer E. A., Nitsche N., Altermatt F. Information use shapes the dynamics of range expansions into environmental gradients // Global Ecol. Biogeogr. 2017. Vol. 26. P. 400–411.
Gasyukov P. S., Kasatkina S. M. Dynamic of the Krill Fishery in the Area 48 and Its Relation to Climate Variability and Changes in Fishing Technology. Bremerhaven (Germany): Document CCAMLR WGEMM-13/25, 2013. 23 p.
Gil M. A., Hein A. M., Spiegel O., Baskett M. L., Sih A. Social information links individual behavior to population and community dynamics // Trends Ecol. Evol. 2018. Vol. 33. P. 535–548.
Gordon T. A. C., Harding H. R., Wong K. E., Merchant N. D., Meekan M. G., McCormick M. I., Radford A. N., Simpson S. D. Habitat degradation negatively affects auditory settlement behavior of coral reef fishes // Proceedings of the National Academy of Sciences of the U.S.A. 2018. Vol. 115. P. 5193–5198.
Harte J. Maximum Entropy and Ecology. A Theory of Abundance, Distribution, and Energetics. Oxford: Univ. Publ., 2011. 280 p.
Holt R. The unraveling of Nature’s information webs: the next depressing frontier in conservation? // Israeli J. Ecol. Evol. 2007. Vol. 53. P. 229–236.
Krampen M. Phytosemiotics // Semiotica. 1981. Vol. 36, No 3/4. P. 187–209.
Kull K. Biosemiotics in the twentieth century: A view from biology // Semiotica. 1999. Vol. 127, No 1/4. P. 385–414.
Kull K. An introduction to phytosemiotics: Semiotic botany and vegetative sign systems // Sign Systems Studies. 2000. Vol. 28. P. 326–350.
Kull K. Jakob von Uexküll: An introduction // Semiotica. 2001. Vol. 134, No 1/4. P. 1–59.
Kull K. Zoosemiotics is the study of animal forms of knowing // Semiotica. 2014. Vol. 198. P. 47–60.
Lee A. E. G., Ounsley J. P., Coulson T., Rowcliffe J. M., Cowlishaw G. Information use and resource competition: an integrative framework // Proceedings of the Royal Society B. 2016. Vol. 283, No 1825. P. 2015–2550.
MacArthur R. H. Fluctuations of animal populations, and a measure of community stability // Ecology. 1955. Vol. 36. P. 533–536.
MacArthur R. Н. Species packing and what interspecies competition minimizes // Proc. Nat. Acad. Sci. USA. 1969. Vol. 64, No 4. P. 1369–1375.
MacArthur R. H. Species packing and competitive equilibrium for many species // Theor. Popul. Biol. 1970. Vol. 1, No 1. P. 1–11.
Madden A. D. A definition of information // Aslib Proceedings. 2000. Vol. 52, No 9. P. 343–349.
Margalef R. La teoria de la information en ecologia // Mem. Real. acad. cienc. y artes Barcelona. 1957. Vol. 32. P. 373–449. (Margalef R. Information theory in ecology // Gen. Syst. 1958. Vol. 3. P. 36–71.)
Margalef R. Communication of structure in plankton populations // Limnology and Oceanography. 1961. Vol. 6, No 2. P. 124–128.
Margalef R. Perspectives in Ecological Theory. Chicago: Univ. Press, 1968. 111 р. (Маргалеф Р. Перспективы в экологической теории. Тольятти: Кассандра, 2012. 122 с.)
Munday P. L., Dixson D. L., Donelson J. M., Jones G. P., Pratchett M. S., Devitsina G. V., Døvingd K. B. Ocean acidification impairs olfactory discrimination and homing ability of a marine fish // Proceedings of the National Academy of Sciences of the U.S.A. 2009. Vol. 106. P. 1848–1852.
Norton B. G., Ulanowicz R. Scale and biodiversity policy: a hierarchical approach // Ambio. 2017. Vol. 21. P. 244–249.
O'Connor M. I., Pennell M. W., Altermatt F., Matthews B., Melián C. J., Gonzalez A. Principles of ecology revisited: integrating information and ecological theories for a more unified science // Front. Ecol. Evol. 2019. Vol. 7. Article 219. P. 1–20.
Odum H. T. Self-organization, transformity, and information // Science. 1988. Vol. 242, No 4882. P. 1132–1139.
Patten B. C. An introduction to the cybernetics of the ecosystem: the trophic-dynamic aspect // Ecology. 1959. Vol. 40. P. 221–231.
Patten B. C. Information processing behavior of a natural plankton community // Amer. Biol. Teacher. 1963. Vol. 25. P. 489–501.
Scheiner S. M., Willig M. R. A general theory of ecology // Theor. Ecol. 2008. Vol. 1. P. 21–28. (Шайнер С. М., Виллиг М. Р. Общая теория экологии // Самарская Лука: проблемы региональной и глобальной экологии. 2021. Т. 30, № 4. С. 77–86.)
Schmidt K. A., Dall S. R. X., van Gils J. A. The ecology of information: an overview on the ecological significance of making informed decisions // Oikos. 2010. Vol. 119. P. 304–316.
Schneider E. D., Kay J. J. Life as a manifestation of the second law of thermodynamics // Math. Comput. Modell. 1994. Vol. 19. P. 25–48.
Sebeok T. A. Zoosemiotics // American Speech. 1968. Vol. 43, No 2. P. 142–144.
Sentis A., Ramon-Portugal F., Brodeur J., Hemptinne J.-L. The smell of change: warming affects species interactions mediated by chemical information // Global Change Biol. 2015. Vol. 21. P. 3586–3594.
Simpson E. H. Measurement of diversity // Nature. 1949. Vol. 163. P. 688. (Симпсон Э. Х. Измерение разнообразия // Самарская Лука: Бюллетень. 2007. Т. 17, № 3 (21). С. 585–587.)
Turovski A. On the zoosemiotics of health and disease // Sign Systems Studies. 2002. Vol. 30, No 1. P. 213–219.
Ulanowicz R. E. Information theory in ecology // Computers and Chemistry. 2002. Vol. 25. P. 393–399.
Ulanowicz R. E., Goerner S. J., Lietaer B., Gomez R. Quantifying sustainability: resilience, efficiency and the return of information theory // Ecol. Complex. 2009. Vol. 6. P. 27–36.
Ulanowicz R. E., Jørgensen S. E., Fath B. D. Exergy, information and aggradation: an ecosystems reconciliation // Ecol. Modell. 2006. Vol. 198. P. 520–524.
Wagner A. Information theory, evolutionary innovations and evolvability // Philosophical Transactions of the Royal Society. B. Biol. Sci. 2017. Vol. 372. P. 416–418.
Walker S. I., Davies P. С. W., Ellis G. F. R. (Eds). From Matter to Life: Information and Causality. Cambridge: Univ. Press, 2017. 494 p.
Walker S. I., Kim H., Davies P. C. The informational architecture of the cell // Transactions of the Royal Society. A. 2016. Vol. 374. P. 1–21.
Wicken J. S. Entropy and Information: suggestions for common language // Philos. Sci. 1987. Vol. 54. P. 176–193.
Благодарности
ФИНАНСИРОВАНИЕ
Работа финансировалась из средств федерального бюджета в рамках выполнения государственного задания Самарского ФИЦ РАН – Институт экологии Волжского бассейна РАН.
КОНФЛИКТ ИНТЕРЕСОВ
Автор заявляет об отсутствии какого-либо конфликта интересов и собственной финансовой заинтересованности при подготовке данной статьи.
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
Настоящая статья не содержит каких-либо исследований с участием людей и животных в качестве объектов изучения.

 © 2011 - 2026
© 2011 - 2026