



Введение
Пространственная структура популяций является результатом взаимодействия многих факторов и механизмов, в том числе гетерогенности местообитаний, репродуктивного поведения, особенностей дисперсии, внутри- и межвидовой конкуренции и др. (Krebs, 1996; Whitehead, 1997; Stenseth et al., 1998; Kleewein, 1999; Madan, Dobson, 2001; Morrison et al., 2006; Szybiak et al., 2009; Крамаренко и др., 2014; Попов, 2015; Guthery, Strickland, 2015 и др.).
Исследования внутрипопуляционного пространства у мелких млекопитающих традиционно связаны с индивидуальным мечением, отслеживанием перемещений отдельно взятых особей, определением границ индивидуальных участков и степени их перекрывания (Жигарев, 2005; Громов, 2008; Олейниченко, 2008; Жигарев и др., 2017 и др.). Этот подход при всех своих неоспоримых преимуществах имеет и существенные ограничения. Высокая трудоемкость работ по организации живоотлова и мечения затрудняет получение массового материала для относительно больших территорий. Накладывает свой отпечаток «индивидуализация» результатов, связанная с поведенческими особенностями конкретных особей в условиях модельного участка.
В то же время изучение пространственного распределения видов довольно давно и успешно решается при помощи процедур моделирования, которые позволяют предсказывать присутствие или отсутствие вида в зависимости от влияющих на него характеристик (предикторов) среды (Guisan, Zimmerman, 2000). При этом исследователи оперируют анонимными подмножествами животных разных видов, формирующих ассоциации в сообществах (Пузаченко, Кузнецов, 1998; Абрамов, Виноградов, 2012). Как отмечал Ю. Г. Пузаченко (2004), аналогичные методы вполне могут быть применимы и для исследования внутрипопуляционной пространственной структуры видов.
В нашей работе на примере рыжей полевки (Myodes glareolus, Schreber) из коренных экосистем южной тайги центра Русской равнины демонстрируются возможности данного подхода и рассматриваются особенности пространственных отношений между внутрипопуляционными группами особей во второй половине репродуктивного сезона. В этот период популяция вида, как правило, характеризуется высокой суммарной численностью и достаточно сложной демографической структурой.
Материалы
Сбор материала проводился в Центрально-Лесном государственном природном биосферном заповеднике (Тверская область, Россия). Территория заповедника представляет собой моренную равнину Валдайского оледенения с высотами 230–310 м над уровнем моря. Регион относится к подзоне южной тайги. Почвенный покров характеризуется большой сложностью и дифференцированностью (Регуляторная…, 2002). Растительный покров в основном представлен ельниками южно-таежного типа и образовавшимися на их месте вторичными лесами (Факторы…, 1983). На слабо расчлененных водоразделах распространены бореальные еловые леса. Хорошо дренированные склоны водоразделов заняты ельниками неморальными. На слабодренированных склонах распространены ельники неморально-бореальные.
В июле – августе 2010–2014 гг. особенности пространственного размещения мелких млекопитающих в континууме естественной среды лесных экосистем изучали с использованием протяженной «градиентной» ловушко-линии (Истомин, Михалап, 2011, 2012), которая была привязана к трансекте, организованной сотрудниками лаборатории биогеоценологии ИПЭЭ им. А. Н. Северцова РАН для исследований ландшафтно-экологической неоднородности растительного покрова заповедника. Трансекта длиной 2280 м пересекала различные типы коренных экосистем южной тайги, в первую очередь ельники разного генезиса. Трансекта размечена 114 визированными точками в системе координат WGS 84 (UTM Zone 36 North). Расстояние между соседними точками составляет 20 метров. Точки имеют подробные геоботанические описания на площадях 20 × 20 и 5 × 5 м. Описания включают 275 переменных фитоценотической среды, микрорельефа и микроклимата.
В каждой точке с использованием ловушек Геро в течение 2–4 суток облавливалась круговая площадка с радиусом 5 м. При этом центральная ловушка устанавливалась в визированной точке, а четыре другие располагались крестообразно на расстоянии 5 м от нее. Всего было отработано 10488 ловушко-суток, отловлено 2096 экземпляров мелких млекопитающих, принадлежащих к 13 видам, из них 1520 – рыжие полевки.
Для рыжей полевки, как и для многих других видов мышевидных грызунов, характерна поливариантность онтогенеза, которая связана прежде всего со спецификой роста, развития и репродуктивного состояния сезонных генераций (Шварц, 1980; Оленев, 1988, 1991, 2002; Большаков и др., 1991; Оленев, Григоркина, 2005, 2011; Истомин, 2007, 2009 и др.). На основании функционально-онтогенетического подхода традиционно выделяют три внутрипопуляционные группы особей: половозрелые сеголетки (созревшие и размножающиеся в год рождения особи, для которых характерен однофазный рост – первый тип онтогенеза); неполовозрелые сеголетки (несозревающие и неразмножающиеся в год рождения особи с двухфазным ростом – второй тип онтогенеза, первая фаза); перезимовавшие животные (второй тип онтогенеза, вторая фаза). Причем количество особей с различными типами онтогенеза непосредственно связано с уровнем формируемой численности и особенностями популяционных циклов.
В табл. 1 приведен объем материала, использованного для анализа паттернов пространственной внутрипопуляционной организации указанных групп рыжей полевки в условиях лесных экосистем южной тайги.
Таблица 1. Объем материала по внутрипопуляционным группам рыжей полевки
| Годы | Всего особей | Число перезимовавших особей | Число половозрелых сеголетков | Число неполовозрелых сеголетков | |||
| самки | самцы | самки | самцы | самки | самцы | ||
| 2010–2014 | 1520 | 75 | 46 | 107 | 84 | 503 | 705 |
Методы
Для характеристики пространственных отношений между внутрипопуляционными группами рассчитывали математические дистанции, используя метод многомерного непараметрического шкалирования (Nonparametric Multidimensional Scaling). Достоинство многомерного шкалирования заключается в возможности дифференцировать различные типы размещения групп особей без предварительного знания особенностей их взаимной ассоциации и определять размерность и структуру пространства, описывающего дисперсии этих групп (Пузаченко, Кузнецов, 1998; Пузаченко, 2004; Ardente et al., 2016).
Для выполнения процедуры многомерного шкалирования были использованы данные о количестве особей различных функционально-онтогенетических групп, отмеченных в каждой точке трансекты. На основании результатов суммарных отловов рассчитывали коэффициенты гамма-корреляции между местами регистрации особей различных групп рыжей полевки на площадках отлова. Преимущество гамма-корреляций по сравнению с другими корреляционными метриками заключается в том, что в своей основе они опираются на условные вероятности и учитывают совпадающие значения в выборках (Siegel, Castellan, 1988). Подобные совпадения значений неизбежны для суммарных выборок (Пузаченко, 2004). Математические дистанции между группами оценивали по формуле:
D = 1 - Gamma,
где D – дистанция, Gamma – значение коэффициента гамма-корреляции.
На основе полученных математических дистанций была выполнена оценка размерности пространства распределения внутрипопуляционных групп на трансекте и вычислены коэффициенты чувствительности каждой группы к осям многомерного шкалирования. Размерность пространства определяли по функции стресса, которая является мерой различия расстояний между объектами в исходном и вновь созданном математическом пространствах. Задача процедуры заключается в подборе путем итераций таких координат точек в новом пространстве, чтобы величина функции стресса между вновь созданным и исходным (эмпирическим) пространствами была минимальной, а подобие матриц – максимальным. Значения стресса изменяются в диапазоне от 0 до 1, где 0 означает полное сходство, а 1 – абсолютное различие между расположением объектов в исходном и во вновь созданном пространстве. Если значение стресса не превышает критического порога в 0.10, то многомерное шкалирование считается успешным. Процедура получения осей многомерного шкалирования на основе матрицы гамма-корреляций подробно изложена Ю. Г. Пузаченко с соавторами (Пузаченко, Кузнецов, 1998; Пузаченко, 2004).
Анализ, обработку и визуализацию статистических данных осуществляли в программах Statistica 8.0 и R 4.0.5.
Результаты
Рыжая полевка на территории Центрально-Лесного заповедника доминирует во всех лесных экосистемах и формирует обширную континуальную популяцию. Особенности многолетней динамики численности вида подробно описаны в целом ряде публикаций (Истомин, 2007а, 2008, 2009, 2014 и др.). В 2010–2014 гг. летняя численность рыжей полевки поддерживалась на достаточно высоком уровне (23.5–78.3 экземпляра на 100 ловушко-суток – в ельниках неморальных; 6.0–45.3 экземпляра на 100 ловушко-суток – в ельниках бореальных). Соотношение внутрипопуляционных групп с учетом пола и функционального статуса особей в суммарных выборках рыжей полевки в исследованный период времени вполне типично для второй половины репродуктивного сезона в условиях хвойно-широколиственных лесов заповедника (рис. 1).
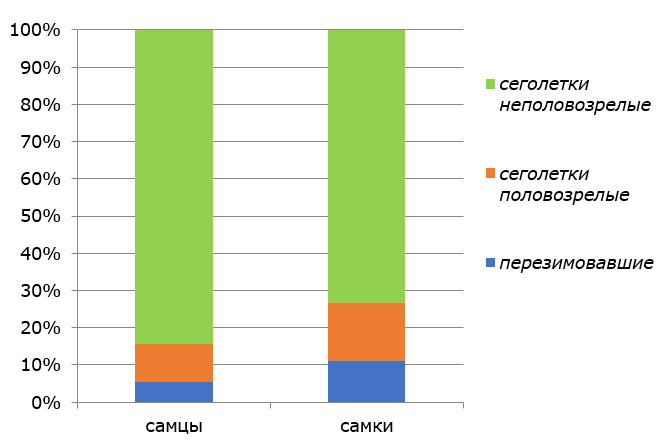
Рис. 1. Структура внутрипопуляционных групп рыжей полевки во второй половине репродуктивного периода
Fig. 1. Structure of intra-population groups of bank voles in the second half of the reproductive period
Проведенное многомерное шкалирование показало, что пространственное размещение внутрипопуляционных групп оптимальным образом воспроизводится двумя осями: на шестой итерации была получена финальная величина стресса, равная 0.028, что свидетельствует о достаточном качестве процедуры многомерного шкалирования. Величины коэффициентов чувствительности отдельных групп к полученным осям дают первичную информацию о структуре пространства (табл. 2.).
Таблица 2. Коэффициенты чувствительности к осям многомерного шкалирования различных внутрипопуляционных групп рыжей полевки
| Внутрипопуляционные группы | Пол | Оси многомерного шкалирования | |
| 1 ось | 2 ось | ||
| Перезимовавшие особи | самцы | -0.039 | -0.986 |
| самки | -0.756 | 0.998 | |
| Половозрелые сеголетки | самцы | 1.280 | 0.600 |
| самки | 0.744 | -0.318 | |
| Неполовозрелые сеголетки | самцы | -0.644 | -0.214 |
| самки | -0.584 | -0.080 | |
Для оценки вклада средовых (в рамках видового экологического пространства) и внутрипопуляционных характеристик в описании каждой из полученных осей было проведено многофакторное регрессионное моделирование. Для этого в качестве независимых переменных использовали число особей каждой внутрипопуляционной группы во всех точках трансекты, а значения отдельно взятых осей многомерного шкалирования – в качестве зависимых переменных (табл. 3).
Таблица 3. Результаты регрессионного анализа зависимости осей многомерного шкалирования от размещения различных внутрипопуляционных групп рыжей полевки (значения Beta-коэффициентов отражают вклад каждой независимой переменной в значение зависимой, R2 – коэффициент детерминации)
| Ось многомерного шкалирования 1 | |||
| Независимая переменная (число особей отдельно взятой половозрастной группы) | Beta ± ошибка | R2 | Уровень значимости |
| Перезимовавшие самцы | 0.002 ± 0.01 | 0.75 | 0.02 |
| Перезимовавшие самки | -0.19 ± 0.01 | < 0.001 | |
| Половозрелые сеголетки (самцы) | 0.37 ± 0.01 | < 0.001 | |
| Половозрелые сеголетки (самки) | 0.22 ± 0.01 | < 0.001 | |
| Неполовозрелые сеголетки (самцы) | -0.46 ± 0.01 | < 0.001 | |
| Неполовозрелые сеголетки (самки) | -0.47 ± 0.01 | < 0.001 | |
| Ось многомерного шкалирования 2 | |||
| Перезимовавшие самцы | -0.28 ± 0.01 | 0.88 | < 0.001 |
| Перезимовавшие самки | 0.50 ± 0.01 | < 0.001 | |
| Половозрелые сеголетки (самцы) | 0.28 ± 0.01 | < 0.001 | |
| Половозрелые сеголетки (самки) | -0.23 ± 0.01 | < 0.001 | |
| Неполовозрелые сеголетки (самцы) | 0.32 ± 0.01 | < 0.001 | |
| Неполовозрелые сеголетки (самки) | 0.18 ± 0.01 | < 0.001 | |
По итогам моделирования методом множественной регрессии было выявлено, что первая ось на 75 % определяется размещением групп животных и на 25 % – прочими переменными среды. В число предикторов, не связанных с особенностями размещения зверьков, в положительной области с наиболее значимым (p < 0.01) вкладом входят: доля широколиственных пород в первом древесном ярусе, развитость и бонитет второго яруса древостоя, размер (возраст) кустов лещины в подлеске, наличие валежа ели. Вторая ось определялась особенностями размещения животных на 88 % и лишь на 12 % – другими характеристиками. Достоверные (p < 0.01) положительные значения β-коэффициентов регрессии получены для переменных, которые характеризуют травяно-кустарничковый ярус, в частности выраженность и высоту второго яруса травянистой растительности, а также наличие валежа широколиственных пород деревьев.
На рис. 2 в двух осях с учетом вероятностей регистрации особей изображены области пространства, занимаемые каждой внутрипопуляционной группой во второй половине лета.
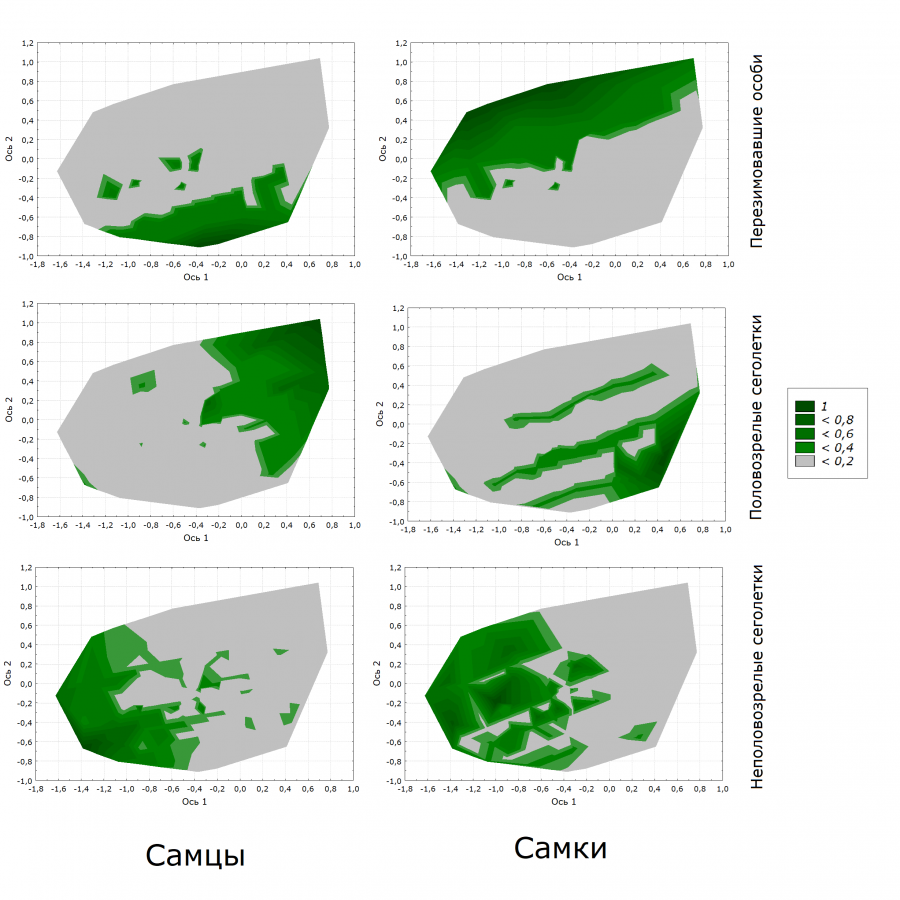
Рис. 2. Размещение исследованных внутрипопуляционных групп в пространстве осей многомерного шкалирования
Fig. 2. Placement of the studied intra-population groups in the coordinate system of multidimensional scaling
Обсуждение
Полученные результаты показывают, что исследованные группы особей рыжей полевки во второй половине репродуктивного сезона приурочены к разным областям экологического пространства (см. табл. 2).
С комплексом условий первой оси положительно и существенно связаны половозрелые сеголетки, причем с наибольшей чувствительностью – самцы. Перезимовавшие самки и неполовозрелые сеголетки обоих полов связаны с первой осью отрицательно. Только перезимовавшие самцы в своем размещении не зависят от нее.
К условиям, определяемым второй осью многомерного шкалирования, более чувствительны перезимовавшие самцы и самки, причем их положение диаметрально противоположно: отрицательная связь отмечена для самцов и положительная – для самок. Для половозрелых сеголетков отмечена обратная зависимость: положительная и более существенная – для самцов, отрицательная – для самок. Неполовозрелые зверьки незначительно зависят от второй оси.
Выполненный регрессионный анализ зависимости значений осей многомерного шкалирования от размещения различных функционально-физиологических групп рыжей полевки показал, что вклад «внепопуляционных» факторов в границах видового экологического пространства составляет не более 25 % для первой оси и не более 12 % для второй оси (см. табл. 3).
Анализ вероятностей регистрации особей различных внутрипопуляционных групп во второй половине репродуктивного периода демонстрирует, что области пространства перезимовавших самцов и самок практически полностью расходятся (см. рис. 2). Перекрывающимися для этих групп являются лишь небольшие участки пространства, характерного для вида. Подобная дифференциация дает основание предполагать, что на этой фазе репродукции вероятность спаривания зимовавших особей друг с другом невелика.
Области пространства, занимаемые самцами и самками из числа половозрелых сеголетков, имеют гораздо больше пересечений друг с другом, чем у зимовавших особей. Это может свидетельствовать о более высокой вероятности спаривания сеголетков друг с другом по сравнению с перезимовавшими зверьками.
Неполовозрелые сеголетки разного пола занимают практически одинаковую область со слабо выраженными оптимумами, но в то же время имеют очень мало пересечений с группой половозрелых молодых особей. Особенно это характерно для самцов.
Обращает на себя внимание почти полное отсутствие перекрывания между перезимовавшими самцами и молодыми половозрелыми самцами, а также между перезимовавшими самками и размножающимися самками-сеголетками. Тогда как степень перекрывания занимаемого пространства в парах: перезимовавшие самцы и участвующие в размножении самки-сеголетки, а также перезимовавшие самки и половозрелые самцы-сеголетки – весьма существенна. Значительная степень пересечения областей перезимовавших самок и половозрелых самцов-сеголетков и практически полное перекрывание пространств, занимаемых перезимовавшими самцами и самками-сеголетками, косвенно свидетельствует о высокой вероятности спаривания особей, принадлежащих к разным возрастным группам (возрастной кросс) во второй половине репродуктивного периода. Это может иметь значение для формирования популяционного разнообразия и динамики популяций в целом.
Некоторые исследователи считают возрастной кросс, в том числе у представителей рода Myodes (Clethrionomys), важной адаптивной стратегией, которая направлена на максимальную реализацию возможности роста численности при одновременном увеличении генетической гетерогенности и эффективном сохранении молодняка (Оленев, Григоркина, 2016). По мнению авторов, возрастной кросс является трансгенерационным путем передачи наследственной информации, значимость которого возрастает в экстремальных условиях (Григоркина, Оленев, 2012; Оленев, Григоркина, 2016). Ранее нами также было показано, что именно в годы активного участия в репродукции сеголетков обоих полов происходят наиболее существенные изменения фенетической структуры популяции к осени по сравнению с весенним периодом (Истомин, 2007б). Наоборот, в годы с низким уровнем участия в размножении сеголетков регистрировались несущественные различия между фенетическими структурами популяции весной, состоящей из перезимовавших особей, и осенью, представленной неполовозрелыми особями разных генераций.
Заключение
Выполненное исследование показало, что метод многомерного шкалирования вполне может быть одним из инструментов для исследования внутрипопуляционной пространственной структуры видов мелких млекопитающих. В условиях коренных экосистем южной тайги Центрально-Лесного заповедника у рыжей полевки во второй половине репродуктивного периода формируется специфическая пространственная внутрипопуляционная структура, которая обусловлена в первую очередь особенностями отношений между зверьками различных функционально-онтогенетических групп. В рамках видового экологического пространства правила размещения детерминированы внутрипопуляционной структурой на 75–88 % и только на 12–25 % определяются факторами иной природы. Причем для некоторых групп (перезимовавшие самцы – перезимовавшие самки, половозрелые самцы-сеголетки – неполовозрелые самцы сеголетки) наблюдается достаточно выраженная пространственная дифференциация. Наоборот, для перезимовавших самцов и половозрелых самок-сеголетков, перезимовавших самок и половозрелых самцов-сеголетков характерна существенная степень перекрывания, что повышает вероятность активного скрещивания разновозрастных партнеров во второй половине сезона размножения. Это может свидетельствовать о наличии возрастного кросса и его существенной роли в популяционной динамике, в первую очередь для поддержания и увеличения гетерогенности популяции. Одним из механизмов обеспечения возрастного кросса, по-видимому, являются обнаруженные особенности пространственной внутрипопуляционной организации рыжей полевки. Поскольку соотношение функциональных групп особей с разными типами онтогенеза достаточно лабильно, более подробный анализ паттернов пространственной внутрипопуляционной организации, безусловно, должен учитывать сезонные и межгодовые аспекты динамики. Также важна и содержательная биологическая интерпретация осей многомерного шкалирования.
Библиография
Абрамов С. А., Виноградов В. В. Экологическая дифференциация мышевидных грызунов лесного пояса гор юга Средней Сибири // Поволжский экологический журнал. 2012. № 4. С. 363–375.
Большаков В. Н., Добринский Л. Н., Кубанцев Б. С., Ищенко В. Г., Оленев Г. В., Горчаковский П. Л., Садыков О. Ф., Безяев В. С. Развитие идей академика С. С. Шварца в современной экологии . М.: Наука, 1991. 276 с.
Григоркина Е. Б., Оленев Г. В. Гетерогенность группировки зимовавших – основа трансгенерационной трансмиссии радиационно индуцированных эффектов у грызунов // Доклады Академии наук. 2012. Т. 443, № 1. С. 136–138.
Громов В. С. Пространственно-этологическая структура популяции грызунов . М.: Товарищество научных изданий КМК, 2008. 582 с.
Жигарев И. А. Локальная плотность и индивидуальные участки рыжей полевки (Clethrionomys glareolus) в условиях южного Подмосковья // Зоологический журнал. 2005. Т. 84, № 6. С. 719–727.
Жигарев И. А., Путилова Т. В., Алпатов В. В. Некоторые особенности пространственной организации рыжих полевок (Myodes glareolus) и малых лесных мышей (Apodemus uralensis) // Научно-практический журнал «Вестник ИрГСХА». 2017. Вып. 82. С. 17–22.
Истомин А. В. Климатические флуктуации и популяционная динамика ценозообразующих видов в эталонных лесных экосистемах главного Русского водораздела // Вестник Псковского государственного педагогического университета. Серия: «Естественные и физико-математические науки». 2007а. № 2. С. 45–60.
Истомин А. В. Популяционная фенетика рыжей полевки (на примере южной тайги) : Монография. Псков, 2007б. 196 с.
Истомин А. В. Мелкие млекопитающие в региональном экологическом мониторинге (на примере Каспийско-Балтийского водораздела) . Псков, 2008. 278 с.
Истомин А. В. Динамика популяций и сообществ мелких млекопитающих как показатель состояния лесных экосистем (на примере Каспийско-Балтийского водораздела) : Дис. … д-ра биол. наук. М., 2009. 481 с.
Истомин А. В. Мелкие млекопитающие в биомониторинге лесных экосистем: комплексный подход // Вестник Тверского государственного университета. Серия «Биология и экология». 2014. № 4. С. 95–113.
Истомин А. В., Михалап С. Г. Градиентный подход и ГИС-анализ при изучении пространственной динамики популяций и сообществ организмов // Вестник Псковского государственного университета. Серия: «Естественные и физико-математические науки». 2012. № 1. С. 49–55.
Истомин А. В., Михалап С. Г. Использование «градиентных» ловушко-линий при изучении мелких млекопитающих в природных условиях // Материалы международной конференции «Актуальные проблемы охраны природы». Чебоксары, 2011. С. 6–7.
Крамаренко С. С., Кунах О. Н., Жуков А. В., Андрусевич Е. В. Анализ паттернов пространственной организации популяций наземных моллюсков: подход с использованием методов геостатистики // Бюллетень Дальневосточного малакологического общества. 2014. Вып. 18. С. 5–40.
Олейниченко В. Ю. Анализ типов пространственного размещения животных по данным мечения на трансектах (на примере землероек-бурозубок) // Известия РАН. Серия биологическая. 2008. № 6. С. 757–764.
Оленев Г. В. Эколого-генетические особенности внутрипопуляционных структурно-функциональных группировок (на примере рыжей полевки) // Экология популяций. М., 1988. Ч. 2. С. 136–138.
Оленев Г. В. Эколого-генетические особенности внутрипопуляционных структурно-функциональных группировок грызунов // Экология популяций. М.: Наука, 1991. С. 54–67.
Оленев Г. В. Альтернативные типы онтогенеза цикломорфных грызунов и их роль в популяционной динамике (экологический анализ) // Экология. 2002. № 5. С. 341–350.
Оленев Г. В., Григоркина Е. Б. Единая неспецифическая адаптивная реакция популяций грызунов на действие факторов различной природы (функционально-онтогенетический подход) // Вестник Нижегородского университета. Сер. «Биология». 2005. № 1. С. 154–162.
Оленев Г. В., Григоркина Е. Б. Динамическое соотношение полов в популяциях цикломорфных млекопитающих (Rodentia, Cricetidae, Muridae) // Зоологический журнал. 2011. Т. 90, № 1. С. 45–58.
Оленев Г. В., Григоркина Е. Б. Эволюционно-экологический анализ стратегий адаптации популяций грызунов в экстремальных условиях // Экология. 2016. № 5. С. 375–381.
Попов С. В. Современные методы исследований пространственной структуры популяций // Структура и разнообразие биоты в природных и антропогенных экосистемах: Материалы XII Всерос. научно-практ. конф., г. Тобольск, 12–13 ноября 2015 г. Тобольск: ООО «Принт-Экспресс», 2015. С. 52–55.
Пузаченко Ю. Г. Математические методы в экологических и географических исследованиях . М.: Изд. центр «Академия», 2004. 416 с.
Пузаченко Ю. Г., Кузнецов Г. В. Экологическая дифференциация грызунов сезонновлажных лесов Южного Вьетнама // Зоологический журнал. 1998. Т. 77, № 1. С. 117–132.
Регуляторная роль почвы в функционировании таежных экосистем . М.: Наука, 2002. 364 с.
Факторы регуляции экосистем еловых лесов . Л.: Наука, 1983. 318 с.
Шварц С. С. Экологические закономерности эволюции . М.: Наука, 1980. 278 с.
Ardente N. C., Ferreguetti Á. C., Gettinger D., Leal P., Mendes-Oliveira A. C., Martins-Hatano F. Diversity and Impacts of Mining on the Non-Volant Small Mammal Communities of Two Vegetation Types in the Brazilian Amazon // PLoS ONE. 2016. Vol. 11. Issue 11. DOI: 10.1371/journal.pone.0167266.
Guisan A., Zimmermann N. E. Predictive habitat distribution models in ecology. Ecological Modeling // Ecological Modelling. 2000. Vol. 135. Issue 2–3. P. 147–186.
Guthery F. S., Strickland B. K. Exploration and critique of habitat and habitat qualityin Wildlife habitat conservation. Concepts, challenges, and solutions. Johns Hopkins University Press, Baltimore, Maryland, 2015. P. 9–18.
Kleewein D. Population size, density, spatial distribution and dispersal in an Austrian population of the land snail Arianta arbustorum styriaca (Gastropoda: Helicidae) // Journal of Molluscan Studies. 1999. Vol. 65. P. 303–315.
Krebs C. J. Population cycles revisited // Journal of Mammalogy. 1996. Vol. 77. P. 8–24.
Madan K. O., Dobson F. S. Population cycles in small mammals: the ahypothesis // Journal of Mammalogy. 2001. Vol. 82, № 2. P. 573–581.
Morrison M. L., Marcot B. G., Mannan R. W. Wildlife‐Habitat Relationships: Concepts and Applications. Third Edition. Island Press. Washington D. C., 2006. 493 p.
Siegel S., Castellan N. J. Jr. Nonparametric Statistics for the Behavioral Sciences (Second Edition). New York: McGraw-Hill, 1988. 399 p.
Stephens R. B., Anderson E. M. Habitat associations and assemblages of small mammals in natural plant communities of Wisconsin // Journal of Mammalogy. 2014. Vol. 95. Issue 2. P. 404–420. DOI: 10.1644/13-MAMM-A-025.
Szybiak K., Bloszyk J., Koralewska-Batura E., Goldin B. Small-scale distribution of wintering terrestrial snails in forest site: relation to habitat conditions // Polish Journal of Ecology. 2009. Vol. 57. Issue 3. P. 525–535.
Whitehead H. Analyzing animal social structure // Anim. Behav. 1997. Vol. 53. P. 1053–1067.
Благодарности
Авторы выражают глубокую признательность Ю. Г. Пузаченко за постоянное внимание к нашей работе на всех ее этапах, методическую помощь и консультации при обработке материала, а также сотрудникам лаборатории биогеоценологии ИПЭЭ им. А. Н. Северцова РАН за предоставленную возможность использовать материалы геоботанических описаний трансекты.

 © 2011 - 2026
© 2011 - 2026