



Введение
Биотические компоненты проточных водных экосистем являются важными критериями оценки качества воды и экологического состояния водоемов. Для этих целей таксономический состав, численность и соотношение чувствительных и нечувствительных таксонов рассматриваются как биологические индикаторы качества воды (Chaïb, Tison-Rosebery, 2012). В России в общегосударственной системе оценки вод водоемов и водотоков (Росгидромет) среди принятых гидробиологических показателей для автотрофного звена водных экосистем рассматриваются параметры фитопланктона (состав, обилие, доминирующие виды, первичная продукция, индекс сапробности), среди бентосных – перифитона. С 2000 г. в странах ЕС регламентами Рамочной водной директивы (ВРД) (European Community..., 2000) значительное внимание при экологической оценке водоемов уделяется бентосу, а среди его автотрофной компоненты – диатомовым водорослям, которые в лотических экосистемах являются наиболее значимой по видовому богатству группой (Round et al., 1990; Diatomeen…, 2013).
В большинстве стран Европы и США фитобентос широко используется для целей мониторинга качества воды в реках (Whitton, Rott, 1996; Prygiel et al., 1999; Оксиюк, Давыдов, 2011, 2012). В России среди биологических компонентов качества доминирует характеристика сапробности в основном по фитопланктону. Река Волга и ее водохранилища в этом отношении изучены достаточно хорошо, продемонстрирована индикационная роль системы сапробности, применение которой позволило отразить происходившие на протяжении ΧΧ века процессы загрязнения и эвтрофирования вод реки (Волга и ее жизнь, 1978; Охапкин, 2011). При этом данные по оценке качества воды по индикаторным видам фитопланктона р. Оки немногочисленны (Охапкин, 1994, 2011).
Поскольку диатомовые водоросли, как наиболее разнообразный и значимый компонент фитобентосного комплекса с хорошо известными экологическими характеристиками (Kelly, 2000), активно применяются для биологического мониторинга лотических экосистем (Bere, Tundisi, 2010), исследования их состава, пространственно-временной динамики и показательных возможностей имеют несомненный интерес для оценки качества воды р. Оки.
Цель работы – на основе исследований видового состава и структуры диатомовых комплексов донных альгоценозов дать их экологическую характеристику, в сравнительном аспекте оценить возможности различных подходов определения качества воды и современного ее состояния по организмам фитобентоса.
Материалы
Пробы фитобентоса отбирали на 3 станциях, расположенных в рипали правобережья р. Оки в пределах г. Нижнего Новгорода (рис. 1) летом (первая декада июля) и осенью (первая декада ноября) 2018 г. на глубине 1.0–1.5 величины прозрачности по диску Секки. Температура воды в июле изменялась от 24 до 27°, а в ноябре была практически одинакова на всех станциях (4.9°). Прозрачность воды (0.5–0.8 м) как летом, так и осенью была невелика, а ее электропроводность осенью колебалась в пределах 634–663.1 μS/cm. Активная реакция среды, достигая в июле 8.3– 8.7, осенью изменялась незначительно (7.8–8.1).
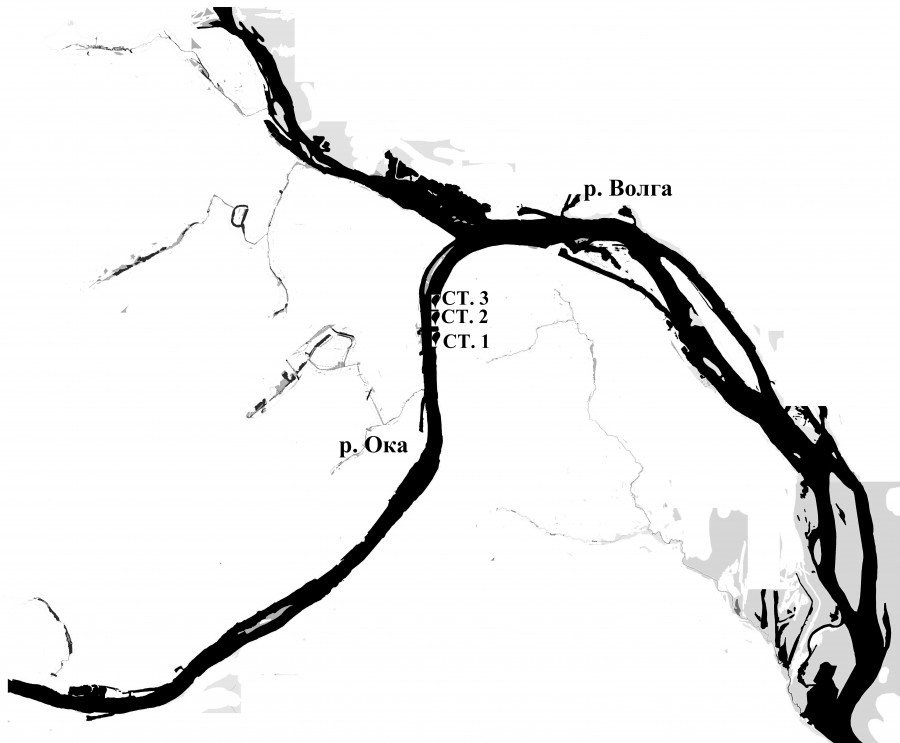
Рис. 1. Карта-схема устья р. Оки с расположением станций отбора проб
Fig. 1. Map of the mouth of the Oka river with the location of sampling stations
Грунты изученного нами участка правобережья р. Оки представлены в основном илами и сильно заиленными песками с вкраплением каменистого субстрата, сформированного преимущественно известняками. Высшая водная и прибрежно-водная растительность развита довольно слабо и представлена отдельными куртинами стрелолиста и немногочисленными особями кубышки и рдеста.
Река Ока – это второй по величине русла и объему стока крупный приток реки Волги (длина 1500 км, площадь водосбора 24500 км2), оказывающий важную роль в формировании гидрологического, гидрохимического и гидробиологического ее режимов, особенно речного участка Чебоксарского водохранилища до устья р. Суры. Качество окских вод на протяжении последних десятилетий оставалось низким и, по данным Верхне-Волжского управления по гидрометеорологии и контролю природной среды (УГКС), оценивалось 4-м классом. Характеристика гидрохимического состава вод р. Оки в пределах города, по данным этой организации (табл. 1), демонстрирует, что воды устьевого участка реки в пределах г. Н. Новгорода отличались высокой мутностью, повышенными концентрациями минеральных форм азота, фосфора, органического вещества, тяжелых металлов и других компонентов загрязнения. Река, как и ранее (Охапкин и др., 2015; Джамалов и др., 2017), относится к категории эвтрофно-гипертрофных водных объектов с высоким уровнем антропогенного влияния.
Таблица 1. Гидрофизические и гидрохимические показатели вод устья р. Оки (2018 г., данные Верхне-Волжского УГКС)
| Показатели | VI–VII 2018 г. |
X–XI 2018 г. |
| pH | 8.23 ± 0.16 | 8.49 ± 0.04 |
| Взвешенные вещества, мг/л | 23.3 ± 3.53 | 12.4 ± 2.03 |
| Растворенный кислород, мг О2/л | 8.60 ± 0.09 | 10.2 ± 0.42 |
| Сумма ионов, мг/л | 349* | 596* |
| Ионы Са2+, мг/л | 60.2* | 85.6* |
| Ионы Mg2+, мг/л | 26.7* | 25.5* |
| Сумма ионов, Na+ + K+, мг/л | 9.6* | 46.0* |
| Гидрокарбонаты, HCO3-, мг/л | 128.0* | 229.0* |
| Сульфаты, SO42-, мг/л | 106.0* | 182.0* |
| Хлориды, Cl-, мг/л | 27.1* | 27.6* |
| БПК5, мг О2/Л | 3.0 ± 0.56 | 2.19 ± 0.31 |
| ХПК, мг О/Л | 24.4 ± 4.6 | 28.8 ± 1.7 |
| Азот нитратов, NO3-, мг N/Л | 1.152* | 0.91 ± 0.04 |
| Азот нитритов, NO2-, мг N/Л | 0.013 ± 0.006 | 0.02 ± 0.006 |
| Азот аммонийный, NH4+, мг N/Л | 0.39 ± 0.16 | 0,20 ± 0,04 |
| Фосфаты, PO42-, мкг P/Л | 30.0* | 91 ± 46 |
| Фосфор общий, Pобщ, мкг P/Л | 99 ± 34 | 95 ± 26 |
| Кремний, Si, мг/л | 0.50* | 1.80* |
| Железо общее, мг/л | 0.08 ±0.01 | 0.02* |
Примечание. Приведены средние арифметические ± ошибка среднего; * – данные единичных измерений.
Методы
Образцы бентосных альгоценозов отбирали с поверхности донных отложений (эпипелон), каменистого субстрата (эпилитон) и высших водных и прибрежно-водных растений (эпифитон) с использованием принятых методических подходов (Водоросли…, 1989; Комулайнен, 2003; Неврова и др., 2015).
Водоросли эпилитона отбирали путем очистки камней жесткой зубной щеткой. Эпифитные диатомовые водоросли счищали с поверхности погруженных макрофитов зубной щеткой, помещая растение в кювету с добавлением дистиллированной воды. Полученные суспензии диатомей помещали в пластиковую бутылку с маркировкой. Пробы населения мягких грунтов (эпипелона) отбирали с помощью пластиковой трубки площадью захвата 14.5 см2 из столба донных отложений, собранных с помощью дночерпателя Петерсена или Экмана – Берджи путем вырезания поверхностного слоя толщиной 5–7 мм. Затем содержимое помещали в маркированный контейнер. Пробы фиксировали 40 % раствором формалина. Всего было отобрано 25 количественных и качественных проб.
В лаборатории образцы диатомовых водорослей обрабатывали перекисью водорода и соляной кислотой (горячее окисление перекисью водорода), постоянные препараты готовили с использованием среды Naphrax (показатель преломления 1.74, Brunel Microscopes Ltd). Идентификация диатомовых водорослей проводилась методами световой микроскопии с применением определителей, основной список которых приведен в работе Е. Н. Невровой с соавторами (2015).
Количественную представленность видов оценивали по модифицированной глазомерной балльной шкале Стармаха (Водоросли…, 1989): 1 – единично, 3 – немного, 5 – умеренно, 7 – много, 9 – массово. Анализировали частоты встречаемости видов (% проб, в которых он обнаружен) и доминирования (% проб, в которых обилие вида составило 7 или 9 баллов). Экологическая характеристика диатомей приведена по С. С. Бариновой с соавторами (2019).
Качество и экологическое состояние окских вод оценивали по индикационным величинам средовых характеристик отдельных видов (Баринова и др., 2019), а также значению сапробности (трофности) посредством расчета индексов загрязнения Т. Ватанабе (DIApo) (Watanabe et al., 1986, 1988, 1990), эвтрофикации / загрязнения А. Дель Уомо (EPI) (Dell' Uomo, 1996) и индекса сапробности Пантле и Букка в модификации В. Сладечека (Sládeček, 1973, 1986). При этом применяли списки индикаторных видов разных авторов (Sládeček, 1973, 1986 – SLA; Wegl, 1983 – WEG; Marvan et al., 2005 – MAR; Баринова и др., 2019 – BAR).
Результаты
В альгоценозах бентали правобережья устьевого участка р. Оки в районе г. Нижний Новгород выявлено 133 вида и внутривидовых таксона диатомовых водорослей, относящихся к 2 классам, 6 порядкам, 17 семействам и 46 родам. Общие характеристики флоры диатомовых бентоса р. Оки довольно схожи с таковыми крупных европейских рек: заметная роль семейств Naviculaceae и Nitzschiaceae, ведущие ранговые позиции родов Navicula, Nitzschia, Gomphonema, Achnanthidium являются чертами сходства и определенной стабильности альгофлоры донных диатомей средних и больших эвтрофированных и заметно загрязненных европейских рек. Преобладание родов Nitzschia, Navicula, Gomphonema и их ведущая роль в составе альгоценозов бентоса установлены ранее для многих европейских водотоков (Волга и ее жизнь, 1978; Владимирова, 1989; Кoziychuk, 2019). Преобладание пеннатных шовных диатомей, ранговое распределение ведущих семейств и родов и в целом состав альгоценозов бентали устьевого района р. Оки свойственны характеристикам диатомовых бентоса крупных речных экосистем с заметным антропогенным влиянием.
Массовые виды, отмеченные более чем в половине проб, составляли лишь одну пятую видового состава, а редкие виды, найденные единично, – 40 %. Состав доминирующих видов со встречаемостью более 50 % и заметной частотой доминирования (20–40 %) небогат (всего 6 видов). Большая часть обычных компонентов бентосных альгоценозов продемонстрировала низкую частоту доминирования, что косвенно свидетельствует о пространственно-временном разнообразии условий обитания диатомей в рипали р. Оки с преобладанием субстратной и сезонной приуроченности (табл. 2).
Таблица 2. Частоты встречаемости (1) и доминирования (2) массовых видов диатомовых водорослей
| Вид | Эпипелон | Эпифитон | Эпилитон | Все альгоценозы бентали | ||||
| 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | |
| Achnanthidium affine (Grunow) Czarn | 14 | 0 | 29 | 0 | 55 | 27 | 36 | 12 |
| Amphora copulata (Kütz.) Schoeman et R. E. M. Archibald | 57 | 0 | 71 | 0 | 73 | 9 | 68 | 4 |
| A. pediculus (Kütz.) Grunow ex A. W. F. Schmidt | 43 | 0 | 9 | 0 | 82 | 9 | 68 | 4 |
| Cocconeis pediculus Ehr. | 43 | 0 | 57 | 14 | 73 | 9 | 68 | 8 |
| C. placentula Ehr. | 100 | 0 | 100 | 29 | 91 | 9 | 96 | 12 |
| Diatoma vulgaris Bory | 71 | 0 | 57 | 14 | 73 | 36 | 68 | 20 |
| Encyonema caespitosum Kütz. | 29 | 0 | 12 | 0 | 55 | 9 | 36 | 4 |
| E. leibleinii (C. Agardh) W. J. Silva, R. Jahn, T. A. V. Ludwig, et M. Menezes | 14 | 0 | 12 | 0 | 45 | 9 | 28 | 4 |
| E. reichardtii (Krammer) D. G. Mann | 0 | 0 | 29 | 0 | 0 | 9 | 4 | 4 |
| E. ventricosum (C. Agardh) Grunow | 57 | 0 | 12 | 0 | 55 | 9 | 44 | 4 |
| Fragilaria vaucheriae (Kütz.) Peters | 57 | 14 | 57 | 0 | 64 | 18 | 60 | 12 |
| Gomphonema pumilum (Grunow.) Reich. et Lange-Bert | 57 | 0 | 86 | 42 | 91 | 18 | 80 | 20 |
| G. parvulum Kütz. | 71 | 0 | 71 | 29 | 82 | 27 | 76 | 4 |
| Navicula cryptocephala Kütz. | 86 | 0 | 86 | 0 | 91 | 18 | 88 | 8 |
| N. tripunctata (O. Müll.) Bory | 86 | 0 | 100 | 14 | 82 | 54 | 88 | 28 |
| Nitzschia dissipata (Kütz.) Rabenh. | 71 | 0 | 43 | 14 | 73 | 54 | 64 | 28 |
| N. linearis W. Sm. | 14 | 0 | 43 | 0 | 18 | 9 | 24 | 4 |
| N. palea (Kütz.) W. Sm. | 29 | 0 | 100 | 29 | 63 | 0 | 64 | 8 |
| Nitzschia sp. | 53 | 0 | 43 | 14 | 34 | 18 | 44 | 12 |
| Pseudostaurasira brevistriata (W. Smith) E. Morales | 43 | 0 | 57 | 0 | 55 | 9 | 52 | 4 |
| Sellaphora seminulum (Grunow) D. G. Mann | 0 | 0 | 0 | 0 | 18 | 9 | 8 | 4 |
| Staurosira venter (Ehr.) Cl. et Möller | 14 | 0 | 0 | 0 | 55 | 9 | 28 | 4 |
| Stephanodiscus hantzschii (Grunow) | 100 | 29 | 100 | 42 | 100 | 9 | 100 | 24 |
| S. invisitatus M. H. Hohn & Hellermann | 71 | 0 | 57 | 14 | 27 | 0 | 48 | 4 |
| S. neoastraea Håk. et Hick | 100 | 29 | 100 | 71 | 100 | 29 | 100 | 40 |
| Tryblionella hungarica (Grunow) Frenguelli | 57 | 0 | 86 | 0 | 73 | 9 | 72 | 4 |
| Ulnaria ulna (Nitzsch.) Compère | 86 | 0 | 100 | 29 | 82 | 0 | 88 | 8 |
Центрические диатомовые водоросли (в основном различные Stephanodiscus, Cyclotella, Aulacoseira) как доминанты и субдоминанты постоянно отмечались во всех типах сообществ бентоса в результате процессов седиментации из водной толщи на дно, демонстрируя максимальную интенсивность доминирования в альгоценозах эпифитона. В сообществах эпилитона более заметная частота доминирования (29 %) наблюдалась у Stephanodiscus neoastraea. В эпипелоне только Fragilaria vaucheriae в 14 % проб выступала в качестве доминанта. Комплекс бентосных видов эпифитона сформирован Nitzschia palea, Cocconies placentula, Ulnaria ulna, Gomphonema pumilum и G. parvulum. В два раза реже в качества доминантов или сопутствующих доминантам компонентов в эпифитоне развивались Navicula tripunctata, Diatoma vulgaris, Cocconies pediculus, Nitzschia dissipata и Nitzschia sp. С максимальными для бентосных альгоценозов показателями доминирования в группировках диатомовых эпилитона отмечены Navicula tripunctata, Nitzschia dissipata, Nitzschia sp., Diatoma vulgaris и Achnanthidium affine (Grun.) Czarn. Реже в состав преобладающих видов входили Gomphonema parvulum, G. pumilum, Navicula cryptocephala и Fragilaria vaucheriae (см. табл. 2).
Сезонные изменения общего видового богатства бентосных диатомовых оказались незначительными, хотя удельные показатели (число видов в пробе) немного возрастали от июля (в среднем 32.6 ± 2.6) к ноябрю (37.5 ± 2.0). Летом среднее по станциям и типам субстратов обилие диатомовых водорослей достигало 77.9 ± 10.0 балла, а в ноябре оно достоверно возрастало (94.9 ± 9.3) (табл. 3). Участие планктонных центрических водорослей в организации донных альгоценозов вслед за снижением их количественных показателей в толще окских вод от лета к осени (Okhapkin et al., 2014) также уменьшалось с 34 % в июле до 18 % в ноябре. Основу суммарного обилия диатомей часто создавали редкие и единичные виды (обилие 1 и 3 балла), осенью – представители с обилием 5–7 баллов (рис. 2).
Таблица 3. Средние показатели обилия диатомовых водорослей донных альгоценозов (М ± m)
| Месяц | Июль | Ноябрь |
| Ст. 1 | 91.6 ± 10.89 | 104 ± 9.18 |
| Ст. 2 | 61.5 ± 17.27 | 97.66 ± 25.45 |
| Ст. 3 | 77.2 ± 8.73 | 96.67 ± 12.44 |
| Эпилитон | 89.4 ± 12.14 | 115.83 ± 7.12 |
| Эпипелон | 44.66 ± 12.99 | 77.25 ± 11.30 |
| Эпифитон | 84.66 ± 5.20 | 99 |
| Суммарное обилие | 77.9 ± 9.97 | 94.85 ± 9.31 |
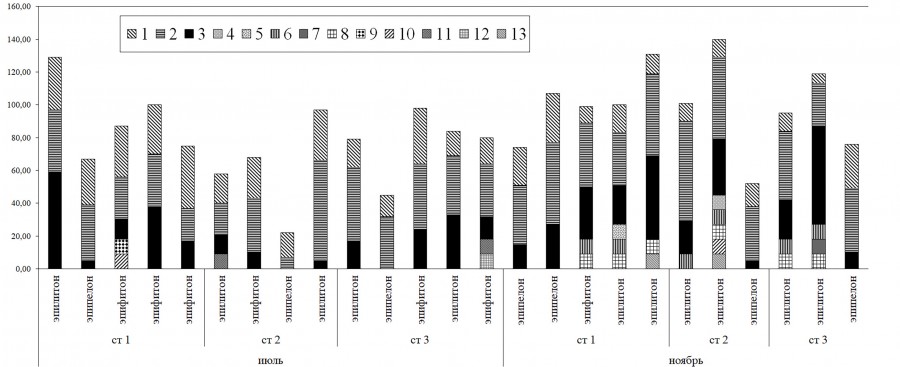
Рис. 2. Пространственно-временная динамика обилия (баллы) сообществ диатомовых водорослей различных субстратов. 1 – центрические, 2 – водоросли с обилием 1 + 3 баллов, 3 – водоросли с обилием 7 + 5 баллов, 4 – Sellaphora seminulum, 5 – Pseudostaurosira brevistriata, 6 – Nitzschia dissipata, 7 – Navicula cryptocephala, 8 – Navicula tripunctata, 9 – Gomphonema parvulum, 10 – Gomphonema olivaceum, 11 – Gomphonema pumilum, 12 – Cocconeis pediculus, 13 – Achnanthidium affine
Fig. 2. Spatial-temporal dynamics of the abundance (points) of communities of diatoms of various substrates. 1 – Centric,
2 – algae with an abundance of 1 + 3 points, 3 – the same, 7 + 5 points, 4 – Sellaphora seminulum, 5 – Pseudostaurosira brevistriata, 6 – Nitzschia dissipata, 7 – Navicula cryptocephala, 8 – Navicula tripunctata, 9 – Gomphonema parvulum, 10 – Gomphonema olivaceum, 11 – Gomphonema pumilum, 12 – Cocconeis pediculus,
13 – Achnanthidium affine
Интенсивность развития диатомовых водорослей эпилитона оказалась несколько большее, чем эпифитона и особенно эпипелона (см. табл. 3). При этом разнообразнее был и общий перечень диатомей, обитающих на каменистом субстрате, хотя вклад основных таксономических групп в формирование видового богатства на уровне классов для разных субстратов был практически одинаковым.
Экологическая характеристика водорослей-индикаторов условий существования донных альгоценозов в р. Оке приведена на рис. 3. По типу местообитания видовой состав диатомовых водорослей отразил одинаковый вклад планктонно-бентосных и бентосных форм (46 и 45 % соответственно), доля компонентов планктона в списке видов невелика.
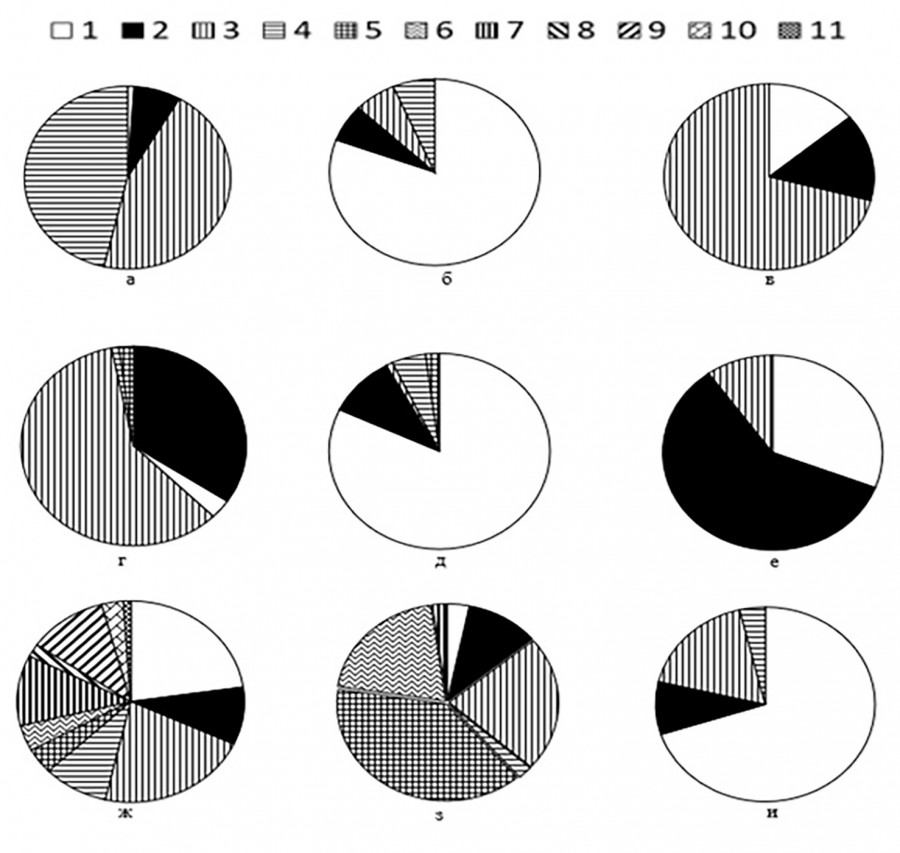
Рис. 3. Экологическая характеристика состава донных Bacillariophyta. Условные обозначения: а: 1 – бентосные; 2 – планктонно-бентосные; 3 – планктонные; 4 – эпибионты; б: 1 – виды умеренного температурного режима; 2 – теплолюбивые; 3 – эвритермные; 4 – холодолюбивые; в: 1 – виды, предпочитающие стоячие воды; 2 – предпочитающие текучие воды; 3 – индифференты; г: 1 – индифференты; 2 – алкалибионты; 3 – алкалифилы; 4 – нейтрофилы; 5 – ацидофилы; д: 1 – индифференты; 2 – галофилы; 3 – галофилы, мезогалобы; 4 – мезогалобы; 5 – галофобы; е: 1 – сапроксены; 2 – эврисапробы; 3 – сапрофилы; ж: 1 – олигосапробионты; 2 – ксено-альфа-мезосапробионты; 3 – бета-мезосапробионты; 4 – альфа-олигосапробионты; 5 – альфа-мезосапробионты; 6 – бета-альфа-мезосапробионты; 7 – альфа-олигосапробионты; 8 – ксено-олигосапробионты; 9 – олиго-бета-мезосапробионты; 10 – ксено-бета-мезосапробионты; 11 – ксеносапробионты; з: 1 – мезотрафенты; 2 – гипертрафенты; 3 – эутрафенты; 4 – гиперэутрафенты; 5 – мезоэутрафенты; 6 – олигомезотрафенты; 7 – олиготрафенты; и: 1 – азотно-автотрофные, выдерживающие повышенные концентрации органически связанного азота; 2 – факультативно азотно-гетеротрофные; 3 – азотноавтотрофные, обитающие при малых концентрациях органически связанного азота; 4 – облигатно-азотногетеротрофные
Fig. 3. Ecological characteristics of the composition of benthic Bacillariophyta. Legend: a: 1 – benthic; 2 – plankton – benthic; 3 – plankton; 4 – epibionts; b: 1 – species of moderate temperature conditions; 2 – thermophilic; 3 – eurythermal; 4 – cold – loving; c: 1 – species that prefer stagnant waters; 2 – preferring flowing waters; 3 – indifferent; d: 1 – indifferent; 2 – alkalibionts; 3 – alkaliphiles; 4 – neutrophils; 5 – acidophiles; d: 1 – indifferent; 2 – halophiles; 3 – halophiles, mesohalobes; 4 – mesohaloba; 5 – halophobes; e: 1 – saproxenes; 2 – evrisaprobes; 3 – saprophiles; g: 1 – oligosaprobic; 2 – xeno – alpha – mesosaprobic; 3 – beta mesosaprobic; 4 – alpha oligosaprobic; 5 – alpha mesosaprobic; 6 – beta – alpha mesosaprobic; 7 – alpha oligosaprobic; 8 – xeno – oligosaprobic; 9 – oligo – beta – mesosaprobic; 10 – xeno – beta – mesosaprobic; 11 – xenosaprobic; h: 1 – mesotrafents; 2 – hypertrafents; 3 – eutrafents; 4 – hypereutrafents; 5 – meso – eutrafents; 6 – oligo – mesotrafents; 7 – oligotrafents; and: 1 – nitrogen autotrophic, withstanding increased concentrations of organically bound nitrogen; 2 – optionally nitrogen heterotrophic; 3 – nitrogen – autotrophic, living at low concentrations of organically bound nitrogen; 4 – obligate nitrogen – heterotrophic
По приуроченности к температурному режиму преобладали обитатели умеренно теплых вод (81 % видов с известными из литературы температурными предпочтениями). Среди них – преобладающие виды из сообществ мягких грунтов (Cocconeis placentula, Gomphonema parvulum), эпифитона (Epithemia sorex Kütz., Eunotia bilunaris (Ehr.) Schaarsch.) и эпилитона (Craticula cuspidata (Kutz.) Mann, Hantzschia amphioxys (Ehr.) Grun.). Единично отмечены эвритермные (Sellaphora pupula (Kütz.) Mereschk., Achnanthidium minutissimum (Kütz.) Czarn.), теплолюбивые (Planothidium lanceolatum (Bréb. ex Kütz.) Lange-Bert., Staurosira venter) и холодноводные диатомеи (Odontidium mesodon (Kütz.) Kütz., Gyrosigma acuminatum (Kütz.) Rabenh).
Индикция отношения к гидродинамике водных масс и кислородному режиму показала, что состав показательных видов почти на две трети сформирован обитателями вод с замедленным течением, умеренно насыщенных кислородом (Surirella angusta Kütz.), представителей, предпочитающих текучие воды, богатые кислородом (Placoneis clementis (Grun.) Cox), и обитатилей стоячих вод (Nitzschia recta Hantzsch ex Rabenh.) значительно меньше (по 14 %).
Среди видов индикторов PH воды преобладали акалифилы (т. е. широко распространенные при PH более 7.0), которые формировали около 60 % состава показательных видов. Среди них основу обилия диатомовых группировок создавали Halamphora veneta (Kütz.) Levkov, Navicula cryptocephala. Немногим более трети состава сформировано индифферентами, предпочитающими PH около 7.0 (например, Meridion circulare (Grev.) Ag.); число алкалибионтов (Ulnaria acus (Kütz.) Aboal), ацидофилов (Eunotia incisa W. Sm. ex Greg.) и нейтрофилов (Odontidium mesodon (Kütz.) Kütz.) много меньше.
По отношению к степени галобности среды 82 % перечня индикаторных видов – это индифференты, типичные обитатели пресных вод с заметным обилием, но в небольших количествах встречающиеся в водах с низкой соленостью. Среди них в качестве ведущих компонентов альгоценозов развивались Fragilaria vaucheriae, Gomphonema italicum Kütz. Галофилы, предпочитающие воды с большой минерализацией, составляли только десятую часть состава индикаторов солености воды, доля галофобов и мезогалобов незначительна (1–5 видов). Среди показателей повышенных значений минерализации вод отметим Meridion constrictum (Ralfs) Kütz., Navicula capitatoradiata Germ и др.
Согласно системе Т. Ватанабе (Watanabe, 1988, 1990), по отношению к степени загрязнения воды органическими веществами преобладали эврисапробы (59 % индикаторных видов, например Navicula radiosa Kütz., Nitzschia clausii Hantzsch., Planothidium rostratum (Østr.) Lange-Bert.) – водоросли, устойчивые к органическому загрязнению, обычно развивающиеся в слабо- и умеренно загрязненных водах. Обитателей чистых и слабо загрязненных вод – сапроксенов много меньше (31 %, например Nitzschia dissipata, Fragilaria vaucheriae). Сапрофилов, преобладающих в водах с сильным органическим загрязнением, немного (10 %). Среди них отметим Luticola goeppertiana (Bleisch) Mann, Nitzschia frustulum (Kütz.) Grun.
Среди индикаторов сапробности, по В. Сладечеку (Sladeček, 1973), по разнообразию видов преобладали обитатели чистых вод (от ксеносапробов до β-олигомезосапробов, 61 % видов списка индикаторов). Показателем средней степени загрязнения – β-мезосапробов значительно меньше (22 %). Диатомей, предпочитающих воды с заметной и значительной степенью содержания органических веществ (от β-α-мезосапробов до α-мезосапробов), немного (17 видов, 17 %, например Craticula cuspidata, Halamphora veneta). Но среди массовых видов с наибольшей представленностью и обилием показатели чистых вод и виды, развивающиеся в водах со средним, повышенным и высоким загрязнением, представлены равным числом индикаторов.
Оценка трофического состояния вод р. Оки продемонстрировала преобладание мезо-эутрафентов (Cocconeis pediculus) и эутрафентов (Diatoma tenuis Ag.) (в сумме более 60 % количества индикаторов) над олиго- (Encyonema caespitosum) и олиго-мезотрафентами (Cymbella cymbiformis Ag.). Несмотря на преобладание в списке видов разнообразия обитателей чистых вод (от ксеносапробов до β-мезо-олигосапробов) распределение диатомовых водорослей по категориям типов питания показало, что подавляющее их большинство — это организмы, требующие для своего развития повышенного (Nitzschia acicularis (Kütz.) W. Sm.) и высокого содержания органических форм азота (Gomphonema parvulum), в том числе и миксотрофы, и только 17 % видов обитателей чистых вод (Neidium dubium (Ehr.) Cl.) предпочитало малые его концентрации.
Из 133 видов диатомовых, зарегистрированных в ходе исследования, для расчета индексов загрязнения использованы только 97. Результаты оценки качества воды устья р. Оки различными биотическими индексами представлены в табл. 4. Пространственно-временные и субстратные изменения показателей, оцененные индексами Пантле и Букка, Ватанабе и Дель Уомо, были невелики, а полученные оценки качества воды несколько различались. В целом степень сапробности вод рипали устьевого участка р. Оки оказалась близкой к средним значениям β-мезосапробной зоны III класса качества с незначительно выраженными тенденциями пространственно-временных изменений.
Таблица 4. Средние показатели (М ± m) различных индексов оценки качества воды по диатомовым водорослям
| Индексы | SLA | BAR | MAR | WEG | WAT | EPI |
| Ст. 1 | 1.87 ± 0.04 | 1.87 ± 0.04 | 1.80 ± 0.03 | 2.09 ± 0.03 | 53.2 ± 3.05 | 2.25 ± 0.05 |
| Ст. 2 | 1.94 ± 0.04 | 1.94 ± 0.04 | 1.80 ± 0.05 | 2.14 ± 0.07 | 50.42 ± 2.30 | 2.29 ± 0.05 |
| Ст. 3 | 1.89 ± 0.02 | 1.89 ± 0.02 | 1.81 ± 0.01 | 2.08 ± 0.02 | 57.5 ± 2.15 | 2.25 ± 0.03 |
| Эпилитон | 1.86 ± 0.02 | 1.87 ± 0.02 | 1.77 ± 0.02 | 2.07 ± 0.02 | 57.18 ± 2.21 | 2.24 ± 0.03 |
| Эпипелон | 1.92 ± 0.04 | 1.92 ± 0.04 | 1.82 ± 0.05 | 2.11 ± 0.05 | 52.28 ± 2.19 | 2.31 ± 0.07 |
| Эпифитон | 1.94 ± 0.04 | 1.94 ± 0.04 | 1.84 ± 0.03 | 2.14 ± 0.03 | 50.00 ± 3.55 | 2.26 ± 0.04 |
| Июль | 1.95 ± 0.02 | 1.95 ± 0.02 | 1.84 ± 0.03 | 2.14 ± 0.03 | 48.57 ± 1.45 | 2.24 ± 0.01 |
| Ноябрь | 1.84 ± 0.02 | 1.84 ± 0.02 | 1.76 ± 0.02 | 2.05 ± 0.02 | 60.45 ± 1.52 | 2.29 ± 0.04 |
Индикация качества вод с помощью индекса эвтрофикации / загрязнения Дель Уомо (EPI), проявляющего значительную корреляцию с химическими и физическими свойствами воды (БПК5, биогенные элементы, электропроводность, хлориды и т. д.) (Зуева и др., 2019), показала, что он изменялся от 2.05 (умеренное загрязнение) до 2.61 (очень сильное загрязнение) (Dell'Uomo, 2003). Средние значения этого индекса (2.24–2.31) характеризуют окские воды как сильно загрязненные и этрофированные.
Обсуждение
В период наблюдения характеристики компонентного состава и уровень загрязнения окских вод в сравнении с данными 2011 г. (Охапкин и др., 2015) изменились незначительно (см. табл. 1). По-прежнему низовья реки можно рассматривать как эвтрофно-гипертрофный водный объект с высоким уровнем антропогенного эвтрофирования и загрязнения (Джамалов и др., 2017) при росте со второй половины XX века к началу XXI минерализации и содержания сульфатов и щелочных металлов.
Динамика химических показателей качества окских вод достаточно четко отразила сезонный ход процессов самоочищения вод в направлении от лета к осени (см. табл. 1). Так, к октябрю – ноябрю в воде отмечен рост (в 1.2 раза) содержания кислорода, снижение количества взвешенных веществ (в 1.9 раза), в том числе и за счет уменьшения интенсивности вегетации фитопланктона (Okhapkin et al., 2014). Кроме того, установлена тенденция к уменьшению содержания в воде лабильного органического вещества (БПК5) и минеральных форм азота (оба показателя в 1.4 раза). При недостоверно выраженных изменениях химического потребления кислорода (ХПК) и концентрации общего фосфора процессы минерализации фосфорсодержащих органических соединений к осени происходили почти полностью. Так, относительное содержание минеральных форм фосфора в общем его запасе с 0.30 летом осенью увеличилось до 0.96. Эти факты отразили тенденцию оптимизации показателей качества воды с возрастанием гидродинамики водных масс и улучшением кислородного режима в реке.
Несмотря на низкий класс качества вод, локальность изученного участка бентали и непродолжительность периода исследования, видовой состав комплексов диатомовых водорослей дна оказался весьма богатым (133 таксона рангом ниже рода). Видовое богатство Bacillariophyta оказалось сопоставимым с перечнем донных и эпифитных представителей диатомей (137 таксонов), обнаруженных нами в составе фитопланктона р. Оки и определенных по результатам многолетних исследований планктонных альгоценозов с 1960-х гг. до настоящего времени (Охапкин, Хедаириа, 2019). Разнообразие видов диатомовых водорослей бентали р. Оки, оцененное нами по результатам рекогносцировочного обследования на основании относительно небольшого числа проб, оказалось сопоставимым с таковым донных диатомей нижнего течения р. Днепр (Владимирова, 1989), но оказалось в 1.7 раза меньше, чем в устьевом районе р. Дунай (Кoziychuk, 2019), и в 1.3 раза ниже, чем в нижнем течении р. Енисей (Левадная, 1986) при значительно большем объеме изученного материала.
Самые многочисленные виды диатомовых водорослей исследованного района р. Оки (Cocconeis placentula, C. pediculus, Gomphonema olivaceum, G. parvulum, Navicula tripunctata, Nitzschia dissipata и др.) являются широко распространенными, эврибионтными по многим экологическим факторам видами – обычными компонентами диатомовой флоры многих рек Европы с заметным антропогенным влиянием (β- и α-мезосапробного типа) (Van Dam et al., 1994).
Таким образом, высокие разнообразие и индикационные свойства диатомовых донных сообществ, а также заметная и часто определяющая их ценотическая роль дают основания для получения надежных и адекватных оценок экологического состояния и качества воды исследованной водной экосистемы.
Оценка экологической целостности проточных вод часто требует разработки комплексных методов, учитывающих сложные взаимосвязи внутри сообществ и между ними на фоне непрерывной изменчивости факторов окружающей среды. Связи между диатомовыми водорослями и экологическими переменными являются достаточно надежными и поддающимися количественной оценке, что делает их подходящими количественными индикаторами экологических условий в лотических экосистемах (Chaïb, Tison-Rosebery, 2012). Диатомовые водоросли используются в качестве альтернативного дополнительного средства оценки качества воды в связи с наличием у многих из них индивидуальных предпочтений к загрязненности или чистоте воды, чувствительности и четкой реакции на физико-химические и биологические изменения (Adewole et al., 2019). При этом считают, что использование диатомовых водорослей для оценки качества воды дешевле, чем обычные химические анализы, и прямо показывает влияние загрязнения на водную биоту (Adewole et al., 2019).
Состав диатомей бентосных альгоценозов устья р. Оки поровну представлен планктонно-бентосными и бентосными видами, обитателями умеренно теплых нейтрально-олигощелочных вод с замедленным течением и умеренно насыщенных кислородом. Их население сформировано главным образом типичными представителями пресных вод с заметным обилием, индифферентами, видами, устойчивыми к органическому загрязнению, показателями как незначительной, так и высокой степени сапробности. Насыщенность вод р. Оки биогенными элементами (фосфор, азот) и органическими веществами отразилась на преимущественном развитии мезо-эутрафентов и эутрафентов – видов, требующих для своей жизнедеятельности повышенного и высокого содержания органических форм азота, в том числе и миксотрофов.
Преобладание в списке видов диатомей представителей, обитающих в чистых водах (60 % от числа индикаторов органического загрязнения), и небольшая доля обитателей высокосапробных вод могут быть связаны c постепенным и незначительным улучшением экологического состояния реки в сравнении с данными 1970–1990-х гг., а также, возможно, с заниженными величинами индивидуальных характеристик многих видов, приведенных в списке С. С. Бариновой с соавторами (2019). Это еще раз подтверждает необходимость работ по региональной адаптации индикаторной значимости отдельных видов. Возможно, такой результат является следствием химического состава грунтов, органическое вещество которых может состоять в основном не из лабильных компонентов (сапробного типа), а из трудно окисляемых, на которые сапробные организмы не реагируют. В связи с этим анализ степени загрязнения вод только на основе списка видов без учета их ценотической значимости может привести к недостоверным оценкам.
Расчеты индексов сапробности Пантле и Букка с использованием списков индикаторных видов разных авторов дали сходные результаты. Средние их значения колебались от 1.76 до 1.95, но определенные по списку R. Wegl (1983) оказались относительно более высокими (2.05 до 2.14) и немного превышающими средние значения для β-мезосапробной зоны. Оценки по индексам Т. Ватанабе аналогичны таковым по индексу сапробности Пантле и Букка. Диатомовый индекс EPI А. Дель Уомо (Dell'Uomo, 2003), на наш взгляд, реальнее отразил уровень качества окских вод (от умеренно до очень сильно загрязненных), что более соответствует оценкам по их химическому составу.
Степень органического загрязнения водной толщи по сезонам колебалась в пределах одного класса качества и имела тенденцию к снижению от июля к ноябрю (см. табл. 4). Эту тенденцию к незначительному улучшению качества воды осенью более четко отразил индекс Ватанабе: средние показатели в ноябре оказались достоверно выше (но сапробность ниже), чем в июле. При этом в ноябре в организации альгоценозов бентали заметно снизилось ценотическое значение обитателей стоячих вод (с 19 до 14 %), что косвенно свидетельствует об усилении процессов гидродинамики. Кроме того, аналогичная тенденция установлена и для факультативных и облигатных гетеротрофов по органическим формам азота, относительное среднее обилие которых с 31 % в июле снизилось в ноябре до 17 %. Таким образом, получено удовлетворительное соответствие оценки временной динамики процессов самоочищения в реке по биоценотическим и химическим показателям.
Сезонные изменения в группировках диатомовых водорослей были обнаружены также и в других реках, например р. Сегре́ (Франция, Испания) (Goma et al., 2005), реках Каталонии (Leira, Sabater, 2005), р. Куарто (Кордова, Аргентина) (Martínez de Fabricius et al., 2003), реках южной Финляндии (Soinine, Eloranta, 2004) и Восточной Фенноскандии (Комулайнен, 2005). Изменения в сообществах диатомовых водорослей в течение года отражают колебания температуры, интенсивности освещения, продолжительности дня, динамики жизненных циклов травоядных, стока (Luis Alberto, 2015) и определяются как природными, так и антропогенными факторами (орошение, забор воды для промышленных целей и др.). Динамика расхода воды может быть основным фактором, контролирующим сезонные изменения состава и структуры фитоперифитона, в том числе и таксоценоза диатомовых водорослей, что отмечалось рядом авторов (Комулайнен, 2005; Boix et al., 2010; Martínez de Fabricius et al., 2003; Tang, Dudgeon, 2013).
Пространственные изменения качества воды были выражены значительно слабее временных, что неудивительно при близком расположении станций отбора проб. Тем не менее, несмотря на это и недостоверность различий индексов в пространственном аспекте, большинство их значений (кроме рассчитанных по спискам Marvan et al., 2005) оказалось выше на ст. 2, особенно индекс Ватанабе. Ст. 2, расположенная ниже нового Окского моста по течению реки, установлена в месте, где река незначительно вдается в береговую полосу правобережья с визуально менее выраженной (в сравнении с другими станциями) гидродинамикой водных масс и ее возможным воздействием на интенсивность процессов самоочищения.
Оценки степени органического загрязнения, полученные с использованием сообществ диатомовых разных субстратов, отобранных на одном участке, в целом были весьма схожими, поскольку в большинстве случаев они были сгруппированы близко друг к другу, что отмечалось также и другими исследователями (Bere, Tundisi, 2011). Тем не менее биоценотические индексы, рассчитанные по сообществам эпилитона, оказались более низкими в сравнении с сообществами других субстратов. Различия структурных и индикационных характеристик альгоценозов, развивающихся на разных субстратах, отмечались рядом авторов (Комулайнен, 2005; Porter et al., 1993; Lowe, Pan, 1996; Kelly et al., 1998), которые говорили об отсутствии единого мнения относительно сравнения оценки качества воды на основе эпифитных и эпилитных сообществ. Предполагалось, что для структуры диатомовых комплексов небольшие различия гидрологических и гидрохимических условий между участками являются более значимыми, чем роль субстратов (Soinine, Eloranta, 2004). Тем не менее в р. Зауэр (Люксембург) и в некоторых реках Финляндии были получены разные результаты в значениях биоценотических индексов по диатомовым водорослям, меняющимся в зависимости от типа субстрата (Torrisi et al., 2006; Bere, Tundisi, 2011). Это, возможно, происходит из-за влияния нескольких абиотических факторов, таких как свет, температура (Tesolin, Tell, 1996) или концентрация нитратов (Coleman, Burkholder, 1995), которые приводят к изменениям в конкретном составе эпилитона. Также было показано, что в сравнении с эпилитоном, видовое разнообразие эпифитона было относительно невелико (Комулайнен, 2005; Cazaubon, 1989), а виды, его составляющие, обладают экологическими характеристиками, которые могут изменять показатели биологического качества окружающей среды.
Ранее на основе оценок сапробнсти по фитопланктону (Охапкин, 2011) было показало, что в устье р. Оки отмечался постепенный рост средневегетационных индексов Пантле и Букка с 1.98 ± 0.07 (по численности индикаторных видов) и 2.16 ± 0.06 (по биомассе) в 1989–1990 гг. до 2.20 ± 0.03 и 2.31 ±.0.05 соответственно в 2011 г. Эти значения получены на основе большого массива материалов, но при подсчете индексов применялась не балльная система определения обилия, а конкретные показатели численности или биомассы видов в пробе. Оценки, полученные по результатам изучения бентосных диатомей, могут быть несколько заниженными в связи с локальностью участка исследований, небольшим периодом исследований, а также недоучетом индикационных свойств водорослей других отделов.
Заключение
- Высокие видовое богатство и индикационные свойства диатомовых донных сообществ, а также заметная и часто определяющая их ценотическая роль дают основания для получения, как правило, надежных и адекватных оценок экологического состояния и качества воды исследованной водной экосистемы.
- Применение различных подходов к оценке качества воды и экологического состояния окских вод отразило пространственно-временные аспекты процессов самоочищения в реке и показало довольно сходные результаты.
- Сообщества эпилитона, как правило, демонстрировали несколько лучшие оценки качества среды обитания, чем таковые эпифитона и эпипелона.
- Индексы сапробности Пантле и Букка в модификации Сладечека отразили средний или немного превышающий средний для β-мезосапробной зоны уровень загрязнения. Индекс эвтрофирования / загрязнения Дель Уомо изменялся в более широких пределах (от умеренного до очень сильного загрязнения), а его средние значения характеризуют окские воды как сильно загрязненные и этрофированные.
- При оценке степени сапробности окских вод по составу и структуре альгоценозов бентали, на наш взгляд, оптимальнее применение списков индикаторных организмов R. Wegl (1983) с последующей их региональной адаптацией и диатомового индекса EPI.
Библиография
Баринова С. С., Бeлоус Е. П., Царенко П. М. Альгоиндикация водных объектов Украины: методы и перспективы . Хайфа; Киев: Изд-во Университета Хайфы, 2019. 367 с.
Владимирова К. С. Фитомикробентос Днепра, его водохранилищ и Днепровско-Бугского лимана . Киев: Наук. думка, 1989. 232 c.
Водоросли: Справочник / С. П. Вассер, Н. В. Кондратьева, Н. П. Масюк и др. Киев: Наук. думка, 1989. 608 c.
Волга и ее жизнь . Л.: Наука, 1978. 348 с.
Джамалов Р. Г., Никаноров А. М., Решетняк О. С., Сафронова Т. И. Воды бассейна Оки: химический состав и источники загрязнения // Вода и экология. 2017. Т. 21. № 3. С. 114–132.
Зуева Н. В., Алексеев Д. К., Куличенко А. Ю., Примак Е. А., Зуев Ю. А., Воякина Е. Ю., Степанова А. Б. Биоиндикация и биотестирование в пресноводных экосистемах: Учебное пособие для высших учебных заведений . СПб.: РГГМУ, 2019. 140 с.
Комулайнен С. Ф. Методические рекомендации по изучению фитоперифитона в малых реках . Петрозаводск: КарНЦ РАН, 2003. 43 с.
Комулайнен С. Ф. Структура и функционирование фитоперифитона в малых реках Восточной Фенноскандии : Автореф. дис. … д-ра биол. наук. СПб.: ЗИН РАН, 2005. 52 с.
Левадная Г. Д. Микрофитобентос реки Енисей . Новосибирск: Наука, 1986. 286 с.
Неврова Е. Л., Снигирева А. А., Петров А. Н., Ковалева Г. В. Руководство по изучению морского микрофитобентоса и его применению для контроля качества среды . Симферополь: Н. Орiанда, 2015. 175 с.
Оксиюк О. П., Давыдов О. А. Санитарно-гидробиологическая характеристика водных экосистем по микрофитобентосу // Гидробиологический журнал. 2011. Т. 47. № 4. С. 66–79.
Оксиюк О. П., Давыдов О. А. Санитарная гидробиология в современный период. Основные положения, методология, задачи // Гидробиологический журнал. 2012. Т. 48. № 6. С. 50–65.
Охапкин А. Г. Фитопланктон Чебоксарского водохранилища . Тольятти: ИЭВБ РАН, 1994. 274 с.
Охапкин А. Г. Опыт применения системы сапробности к оценке качества воды реки Волги // Биоиндикация в мониторинге пресноводных экосистем II: Сборник материалов международной конф. СПб.: Любавич, 2011. С. 91–96.
Охапкин А. Г., Андриянова Н. В., Максимова В. А., Шарагина Е. М., Воденеева Е. Л. Динамика гидрохимического состава вод нижнего течения р. Оки // Вода: химия и экология. 2015. № 5. С. 15–21.
Охапкин А. Г., Хедаириа Т. Диатомовые водоросли как компонент донных альгоценозов устьевого участка реки Оки // Вопросы современной альгологии. 2019. № 2 (20). С. 60–63.
Adewole M. G., Benjamin O. Di., Adewale M. T., Isaac T. O. Application of Diatom-Based Indices in River Quality Assessment: A Case Study of Lower Ogun River (Abeokuta, Southwestern Nigeria) Using Epilithic Diatoms // Limnology – Some New Aspects of Inland Water Ecology. Didem Gökçe, IntechOpen, 2019. DOI: 10.5772/intechopen.86347.
Bere T., Tundisi J. G. Biological monitoring of lotic ecosystems: the role of diatoms // Brazilian Journal of Biology. 2010. Vol. 70. P. 493–502.
Bere T., Tundisi J. G. The Effects of Substrate Type on Diatom-Based Multivariate Water Quality Assessment in a Tropical River (Monjolinho), São Carlos, SP, Brazil // Water Air Soil Pollut. 2011. Vol. 216. P. 391–409.
Boix D., Garcia-Berthou E., Gascon S., Benejam L., Tornes E., Sala J., Benito J., Munne A., Sola C., Sabater S. Response of community structure to sustained drought in Mediterranean rivers // Journal of Hydrology. 2010. Vol. 383. P. 135–146.
Cazaubon A. La florule épiphyte principalement diatomique de diverses plantes-hôtes à la source d’une rivière méditerranéenne (l’Argens, sud-est de la France) // Cryptogamie Algol. 1989. Vol. 10. P. 195–207.
Chaïb N., Tison-Rosebery J. Water quality assessment and application of the biological diatom index in the Kebir-East wadi, Algeria // African Journal of Aquatic Science. 2012. Vol. 37. № 1. P. 59–69.
Coleman V. L., Burkholder J. M. Response of microalgal epiphyte communities to nitrate enrichment in an eelgrass (Zostera marina) meadow // J. Phycol. 1995. Vol. 31. P. 36–43.
Dell'Uomo A. AIgae ofrunning waters in Italy and their importance for monitoring rivers // Bocconea. 1996. Vol. 16. № 1. P. 367–377.
European Community Directive 2000/60/EC of the European parliament and of the Council of 23 October 2000 establishing a framework for Community action in the field of water policy. Official Journal of the European communities L327, 2000.
Goma J., Rimet F., Cambra J., Hoffmann L., Ector L. Diatom communities and water quality assessment in Mountain Rivers of the upper Segre basin (La Cerdanya, Oriental Pyrenees) // Hydrobiologia. 2005. Vol. 551. Issue 1. P. 209–225.
Hofmann G., Werum M. und H. Lange-Bertalot. Diatomeen im süßwasser-benthos von mitteleuropa. Kanigstein: Scientific Books, 2013. 908 S.
Kelly M. G., Cazaubon A., Coring E., Dell’uomo A., Ector L., Goldsmith B., et al. Recommendations for the routine sampling of diatoms for water quality assessments in Europe // Journal of Applied Phycology. 1998. Vol. 10. P. 215–224.
Kelly M. G. Identification of Common Benthic Diatoms in Rivers // Field Studies. 2000. Vol. 9. P. 583–700.
Koziychuk E. Sh. Phytomicrobenthos Diversity in Water courses of the Killya Delta of the Danube River // Hydrobiol. J. 2019. Vol. 55. № 5. P. 44.
Leira M., Sabater S. Diatom assemblages distribution in catalan rivers, NE Spain, in relation to chemical and physiographical factors // Water Research. 2005. Vol. 39. P. 73–82.
Lowe R. L., Pan Y. Benthic algal communities as biological indicators // R. J. Stevenson, M. L. Bothwell, & R. L. Lowe, Eds. Algal ecology, Freshwater benthic ecosystems. San Diego: Academic, 1996. P. 705–739.
Luis Alberto Quevedo Báez. Effects of global change on benthic communities of the lower Ebro River: implications for the assessment of the ecological status: PhD Thesis. … URV. Tarragona, Spain, 2015. 164 p.
Martinez de Fabricius A. L., Maidana N., Gomez N., Sabater S. Distribution patterns of benthic diatoms in a Pampean river exposed to seasonal floods: the Cuarto River (Argentina) // Biodiversity and Conservation. 2003. Vol. 12. P. 2443–2454.
Marvan P., Maršálek B., Heteša J., Sukačova K., Maršálková E., Geriš R., Kozáková M. Comments on the revised tables of algal (and other botanical) water quality indicators listed in CSN 75 7716-discussion material for assessment of trophic status of water bodies. Association Flos Aquae www.cyanobacteria.net on 6th May. 2005.
Okhapkin A. G., Genkal S. I., Sharagina E. M., Vodeneeva E. L. Structure and dynamics of phytoplankton in the Oka river mouth at the beginning of the 21th century // Inland Water Biol. 2014. Vol. 7. № 4. P. 357–365.
Porter S. D., Cuffney T. F., Gurtz M. E., Meador M. R. Methods for collecting algal samples as part of the National Water-Quality Assessment Program. US Geological Survey Open-File Report 93-409. US Geological Survey, Raleigh, North Carolina, 1993. 39 p.
Prygiel J., Whitton B. A., Bukowska J. Use of algae for monitoring rivers III. Douai, France: Agence de l’Eau Artois-Picardie, 1999. P. 224–238.
Round F. E., Crawford R. M., Mann D. G. The Diatoms: biology and morphology of the genera. Cambridge: Cambridge University Press, 1990. 760 p.
Sládeček V. System of Water Analysis from the Biologica Point of View. Archiv fur Hydrobiologie-Beiheft Ergebnisse der Limnologie, 1973. 222 p.
Sládeček V. Diatoms as indicators of organic pollution // Acta Hydrochim. 1986. Vol. 14. № 5. P. 555–566.
Soinine J., Eloranta P. Seasonal persistence and stability of diatom communities in rivers: Are there habitat specific differences? // European Journal of Psychology. 2004. Vol. 39. P. 153–160.
Tang T. S., Niu D. Dudgeon. Responses of epibenthic algal assemblages to water abstraction in Hong Kong streams // Hydrobiologia. 2013. Vol. 703. P. 225–237.
Tesolin G., Tell G. The epiphytic algae on floating macrophytes of a Paraná river floodplain lake // Hydrobiologia. 1996. Vol. 333. P. 111–120.
Torrisi M., Rimet F., Cauchieb H. M., Hoffmann L., Ector L. Bioindication par les diatomees epilithiques et epiphites dans la rivier Sure (Luxembourg) // Royal Botanical Society of Belgium. 2006. Vol. 139. № 1. P. 39–48.
Van Dam H., Mertens A., Sinkeldam J. A coded checklist and ecologicalindicator values of freshwater diatoms from the Netherlands. Netherlands // Journal of Aquatic Ecology. 1994. Vol. 28. № 1. P. 177–133.
Watanabe T., Asai K., Houki A. Numerical water quality monitoring of organic pollution using diatom assemblages // Sci. Total. Environ. 1986. Vol. 55. P. 209–218.
Watanabe T., Asai K., Houki A. Biological information closely related to the numerical index DAIpo (Diatom Assemblage Index to Organic Water Pollution) // Jpn. J. Diatomol. 1988. Vol. 4. P. 49–58.
Watanabe T., Asai K., Houki A. and Michiaki S. Numerical simulation of organic pollution based on the attached diatom assemblage in Lake Biwa // Diatom. 1990. Vol. 5. P. 9–20.
Wegl R. Index fur die Limnosaprobitat. Wasser und Abwasser. German: Print book, 1983. 175 p.
Whitton B. A., Rott E. Use of algae for monitoring rivers II. Innsbruck: Institut für Botanik, Universität Innsbruck, 1996. 196 p.

 © 2011 - 2026
© 2011 - 2026