



Введение
Рептилии эктотермные, но теплолюбивые животные, и в умеренной полосе суточный ход температуры их тела охватывает очень широкий диапазон (примерно от 5 до 40 °С). Величина этого диапазона очень близка даже у разных видов умеренных широт, не говоря о представителях одного вида. Это обстоятельство сильно затрудняет сравнение характеристик терморегуляции разных групп животных.
Ситуацию усугубляет все еще распространенный метод сбора данных (полевые замеры температуры тела у отдельных единичных представителей), который дает очень ограниченные объемы выборок случайных температур. С появлением миниатюрных датчиков температуры (логгеров), которые можно вживить в полость тела, появилась возможность записывать большое количество показаний температуры для многих особей в любое время суток на протяжении длительных периодов жизни.
Большой объем собираемых таким способом данных позволяет вести поиск статистически обоснованных показателей способности рептилий к терморегуляции. Этому вопросу посвящены несколько наших последних публикаций (Коросов, Ганюшина, 2019, 2021). К сожалению, экологическое содержание этих публикаций заставляло сокращать описание достаточно сложных методов, поэтому мы решили посвятить отдельную публикацию только методическим вопросам.
Цель настоящего сообщения состоит в том, чтобы показать подходы к конструированию термобиологических характеристик и предложить ряд показателей, которые позволят выполнять статистически обоснованные сравнения этих показателей для разных групп рептилий.
Материалы
Работы проводились в теплый сезон 2017–2019 гг. на стационаре Института биологии КарНЦ РАН «Гомсельга» (N62.068310°, E33.958824°). Во время эксперимента несколько особей обыкновенной гадюки (Vipera berus L.) жили в вольере размером 8 * 8 м; ночевали в убежище (размером 1.5 * 1.5 * 1 м), сложенном из булыжников 10 на 15 см, в дневное время выползали наружу погреться под лучами солнца. На площадке находился «водоем» в виде вкопанного в землю таза с водой, прикрытого досками. В вольеру периодически выпускали лягушек разного размера в качестве объектов питания, зафиксированы случаи охоты гадюк в вольере. Исследовали температуру 23 взрослых особей обыкновенной гадюки. Температуру регистрировали с помощью логгеров ДТНЗ-28 (масса 1 г, объем 0.8 мл, точность измерения ±0.1 °С, рабочий диапазон от -15 до +60 °С, производство ООО «ЭМБИ РЕСЕРЧ») и ds1921, вживленных под кожу спины примерно посередине туловища (операция проводилась в течение 1–2 мин. на животных, охлажденных до 4–5 °С). Поведение фиксировалось на видео web-камерой, камерами «Ssocco» и «Xiomi YI Lite» в режиме замедленной съемки 1 кадр в 1 секунду. Для идентификации змей их метили разноцветным лаком. Логгеры ds1921 и ДТНЗ-28 были настроены для сбора температуры через каждые 2 мин. в течение всего периода наблюдений. Параллельно с наблюдениями за змеями велась запись температуры воздуха в негустой траве (высотой 10–15 см) и на солнцепеке («черный логгер»). Общая продолжительность наблюдений составила 19 дней в 2017 г., 20 дней 2018 г., 30 дней в 2019 г.
Все наблюдения в среде Excel организованы в базу данных и импортированы в формат CSV. База данных имеет следующие поля: год, месяц, час, минута, номер особи, пол, масса, длина тела, текущая температура особи, текущая температура в траве, в укрытии, на открытом месте («черный логгер»), номер позы (0 – плоская, 1 – круглая, 2 – полукольцо, 3 – кольцо, 4 – клубок), характер движений (0 – в укрытии, 1 – на поверхности, 2 – перемещается), рабочие составные ключи. Одна запись соответствует текущему наблюдению за одной особью в данный момент времени (с обычным шагом 2, часть наблюдений сделана с шагом 1 или 8 мин). Всего для 23 особей число записей составляет 286400.
Вся количественная обработка (поиск, фильтрация данных, составление выборок, расчет показателей) выполнена в среде пакета R (R Core Team, 2012).
Традиционные методы исследований
История изучения термобиологии рептилий достаточно продолжительна (Черлин, 2019а, 2019б). Однако до сих пор нет определенности в адекватных методах описания способности рептилий к активной терморегуляции. Здесь мы будем рассматривать не все показатели термобиологии рептилий, но остановимся только на тех, которые выражают их способность регулировать температуру тела. Характеризуя разные виды и удаленные популяции по их способности поддерживать «оптимальную» температуру тела, обычно применяют ограниченный набор показателей. Как правило, это статистическое обобщение полевых наблюдений, т. е. характеристики малочисленных выборок значений температуры тела, полученных в полевых условиях вручную, а также некоторое множество показателей, полученных при лабораторном содержании животных и их испытаниях в термоградиент-приборах. К типичными характеристиками температурных предпочтений рептилий, которые рассматриваются как показатели способности к терморегуляции, относятся примерно пять показателей. Это диапазон «оптимальных» температур (наблюдаемых обычно в течение теплого времени суток), средняя для этого диапазона температура тела, максимальная зарегистрированная температура тела, преферентная температура (как средняя температура субстрата, выбранная животными в термоградиент-приборе), диапазон температур полной активности (Черлин, 2010). К сожалению, даже появление миниатюрных логгеров, регистрирующих температуру тела рептилии, не привело к появлению новых адекватных методов ее анализа. Обработка этих показаний мало отличается от обработки выборок ручных замеров, выполненных в полевых условиях. Как правило, рассчитывают средние значения температуры тела групп животных, которые относятся уже не ко всему теплому времени, а к определенному часу наблюдений; смысл работы состоит в сравнении этих статистических показателей, полученных для разных видов (Lourdais et al., 2013).
Все перечисленные выше показатели имеют существенные технические или теоретические недостатки и, на наш взгляд, практически не годятся для ответственного сравнения разных групп животных. Полевые замеры максимальной температуры тела из-за своей единичности обладают плохими статистическими свойствами. Преферентные температуры определяются только в искусственных условиях, возможно, меняющих отношение животных к теплоте, к тому же требуют дорогостоящего оборудования. Диапазон размаха типичных температур тела и его средние оценки характеризуют по большей части тепловые условия, воздействующие на особь в данный момент времени, а не их потребности в тепле или терморегуляторное поведение.
На наш взгляд, основная причина конструирования неудачных показателей состоит в том, что теория терморегуляции рептилий находится еще в зачаточном состоянии в отличие от достаточно развитой теории для гомойотермных животных. Герпетологи некритично берут на вооружение понятия из термобиологии маммалий (например, представление об оптимальном диапазоне температур) и пользуются неподходящими методами оценки соответствующих параметров. Между тем принципы терморегуляции, по крайней мере, гелиотермных рептилий существенно отличаются от гомойотермных животных. Применяя адекватную теорию регуляции, можно получить более точные показатели, выражающие способность животных к поддержанию температуры тела, – параметры терморегуляции.
Оригинальные методы исследований
Для выработки показателей терморегуляции гелиотермных рептилий, к которым относится живущая на Севере гадюка обыкновенная, мы воспользовались кибернетико-физиологической теорией регуляции, подробно рассмотренной нами ранее (Коросов, 2008). В самом кратком изложении физиологическая регуляция состоит в выработке организмом реакций, компенсирующих уклонение некой характеристики внутренней среды организма от видовой нормы (Шмидт-Ниельсн, 1981). В поле зрения физиолога находятся две величины: некий нормальный «штатный» параметр, хранящийся в регистрирующих структурах, и текущая характеристика состояния особи. Отдельный акт регуляции состоит, во-первых, в обнаружении уклонения значений текущей характеристики от значения параметра, во-вторых, в выработке компенсаторных реакций.
В термобиологии млекопитающих штатный параметр (хранимый в гипоталамусе) задает уровень необходимой температуры тела. В норме он является неизменным единственным общим критерием для регистрации двух типов уклонения температуры тела – как в сторону больших значений (перегрев), так и в сторону меньших значений (переохлаждение) (Слоним, 1971). В первом случае организм включает реакции усиленного охлаждения («физическая терморегуляция»: потоотделение и пр.), во втором – реакции усиленного разогрева («химическая терморегуляция», тремор и пр.). Распределение уклонений в обе стороны в целом имеет симметричную форму, и для его описания совершенно обоснованно применение средней арифметической и показателей изменчивости температуры тела. Анализ этого распределения позволяет оценить диапазон нормальных температур, который и называют оптимальными температурами.
Терморегуляция гелиотермных рептилий организована совершенно по-другому. Единственный неизменный видовой штатный параметр представляет собой «верхний ограничитель», он служит только для регистрации уклонений температуры тела рептилии в сторону перегрева и при его достижении включает серию реакций, снижающих температуру тела (Коросов, Ганюшина, 2019). В числе таких реакций вазомоторная регуляция, усиленная вентиляция слизистых поверхностей, изменение позы относительно субстрата и потока солнечного излучения, поведенческие реакции, направленные на поиск прохладных субстратов, местообитания или убежищ (Шмидт-Ниельсен, 1982; Черлин, 2012). На диаграмме суточного хода температуры тела значение штатного параметра выглядит как прямая линия, от которой траектории индивидуальной температуры тела должны бы «отскакивать» вниз, в область более низких температур. Гадюка именно таким образом односторонне регулирует температуру тела, избегая перегрева.
При этом у рептилий нет неизменного видового нижнего температурного порога, регулирующего избегание охлаждения. Иначе они, подобно млекопитающим, вынуждены были бы тратить много усилий для поиска температур выше этого значения. Однако этого не происходит: при отсутствии необходимых условий рептилии терпеливо и неподвижно остывают. Вместо этого у рептилий есть относительный температурный порог, побуждающий их переходить в более благоприятные тепловые условия, если они имеются поблизости и доступны для органов чувств. Этот феномен уже давно назван теплолюбивостью. Однако здесь речь идет о температурной чувствительности, которую мы рассматриваем как второй ключевой параметр активной терморегуляции у рептилий (Коросов, 2008, 2010). Его относительный характер состоит в том, что, независимо от температуры тела рептилии, они предпочтают переместиться в местообитание (локацию) с более комфортными тепловыми условиями. Это перемещение осуществляется, когда уровень теплового излучения от окрестных нагретых объектов превысит некоторый порог. Например, гадюка начнет перемещаться, если на расстоянии 30 см температура субстрата будет выше температуры ее тела примерно на 5–8 °С (Коросов, 2010).
Поскольку температура субстрата во многом зависит от уровня солнечной радиации (времени суток, сезона, облачности и пр.), большую часть суток температура тела рептилий будет существенно ниже отмеченного порога перегрева. В силу указанных особенностей терморегуляторного поведения гелиотермных рептилий распределение температуры их тела в любой период теплого сезона будет иметь сложный характер, в целом не связанный с терморегуляцией. В этом случае такие статистические параметры, как средняя арифметическая или диапазоны (доверительные интервалы и пр.), ничего не будут сообщать о существе терморегуляторных реакций рептилий. Попытки различных герпетологов как-то ограничить рассматриваемый диапазон температур внешними критериями, например, замерами только днем, только в ясную погоду, только во время «активности», не меняет сути вопроса – полученный диапазон температур тела будет определяться в той или иной степени тепловыми условиями среды.
Если целью термобиологических исследований принять поиск видовых физиологических параметров терморегуляции, то необходимо выходить на оценку именно пороговых значений температуры тела, достижение которых вызывают терморегуляторные реакции. Такие значения будут оценками физиологических «штатных» параметров терморегуляции (Коросов, 2015). Видимо, температура поверхности тела или внутренних слоев, замеренная логгерами, не будет равна «штатной» критической температуре, которая также, скорее всего, хранится в гипоталамусе. Однако те значения температуры тела, при которых змея осуществляет терморегуляторное поведение, должны жестко коррелировать с физиологической константой терморегуляции и поэтому могут служить ей хорошей оценкой. Аналогичная оценка – «нормальная температура под мышкой человека», хотя «штатный» параметр терморегуляции примерно на 1 °С выше нее. Одна из наших задач – поиск оценок этих «критических» значений температуры тела, характерных для особей определенного статуса, их видовой нормы.
Если же целью становится поиск устойчивых воспроизводимых характеристик терморегуляторных способностей рептилий, такими характеристиками могут стать показатели скорости изменения температуры тела.
Наконец, если в центре внимания стоит поиск особенностей терморегуляторных реакций различающихся по статусу (возрасту, полу, зрелости и пр.) особей, можно попробовать построить относительные метрики «теплолюбивости» животных, живущих в одинаковых условиях.
Наша работа в этом направлении привела к конструированию 10 новых показателей, также использован один ранее известный.
Таблица 1. Термобиологические характеристики
| Название | Имя | Характеризует... |
| Время выхода | t | начало дневного обогрева под лучами солнца |
| Скорость нагревания | V | способность к аккумуляции теплоты |
| Сумма температур | S | теплолюбивость особи |
| Максимальная высшая | Tmh | предельный порог толерантности |
| Максимальная пиковая | Tmp | статистически обоснованный порог толерантности |
| Максимальная типичная | Tmt | статистически обоснованный порог толерантности |
| Максимальная добровольная | Tmg | порог толерантности, оцененный по поведению особи |
| Средняя температура баскинга | Tb | типичную температуру тела на баскинге |
| Средняя температура баскинга на поверхности | Tbs | температуру змеи, греющейся на открытой поверхности |
| Медианные активные температуры | Tact | правую часть распределения температур тела |
| Средняя нормированная температура | Tn | среднее значение отклонения температуры особи от среднего для группы |
Время выхода из укрытия утром
Моментом выхода особи (ai) считали тот отсчет времени (число минут после полуночи) утром данного дня (j = 1, 2 …, N), после которого температура тела отдельной змеи (i = 1, 2 …, n) возрастала на 1.5 °С (очевидно, под лучами солнца) (рис. 1). Этот показатель годится для сравнения реактивности разных змей только для данного дня.
Далее для данного дня по всем особям рассчитывали среднюю (Maj) и стандартное отклонение (Saj). Затем для каждой особи (i) для данного дня (j) отыскивали центрированное-нормированное отклонение: naij = (aij – Maj) / Saj и для каждой змеи значения усредняли Ai = Σ naij / N. Стандартное отклонение рассчитывается по стандартной формуле.
Показатель Ai характеризует относительное время выхода за весь период наблюдений и может использоваться для сравнения разных особей, наблюдавшихся как в данный год, так и в разные годы. Для расчета корреляции времени выхода с другими характеристиками змей (или условиями среды) следует выполнить центрирование-нормирование и этих показателей. Например, сначала рассчитать относительные значения массы тела у n особей: nWi = (wi – Mwij) / Swij, затем выполнить анализ связи между этими относительными значениями разных особей.
Скорость нагревания
Для определения скорости нагревания вручную отбирали только те данные, когда график утренней температуры был восходящим и гладким. Для каждой змеи все траектории собирали в общую выборку и рассчитывали линейную регрессию температуры (T) от времени (t): T = a + V * t; коэффициент пропорциональности принимали за индивидуальный показатель скорости нагревания Vi (в рамках регрессионного анализа определяется и его ошибка) (см. рис. 1).
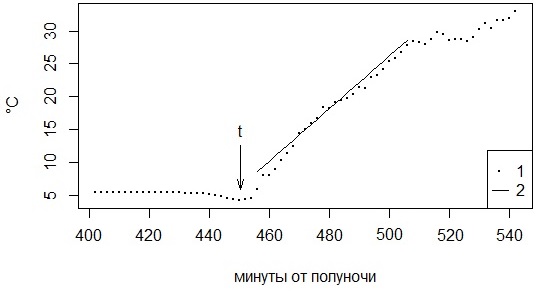
Рис. 1. Фрагмент температурной кривой в момент выхода особи из убежища (t) и в начале нагревания (1 – замеры температуры, 2 – линия регрессии для расчета скорости утреннего нагревания)
Fig. 1. Fragment of the temperature curve at the moment the individual leaves the shelter (t) and the beginning of heating (1 – temperature measurements, 2 – regression line for calculating the rate of morning heating)
Смысл показателя в абсолютном выражении – это количество градусов, приобретенных животным за минуту нагревания. Его значения можно рассматривать как выражение индивидуальных свойств и непосредственно сравнивать у разных особей.
В то же время на скорость нагревания могут влиять условия среды, например ветер, низкие температуры воздуха, субстрата. По этой причине и быстро, и медленно нагревающиеся животные могут иметь одинаковые скорости при разных условиях среды.
Для поиска различий этих показателей уместно использовать бутстрэп (Шитиков, Розенберг, 2014) – как при сравнении пары особей, так и в дисперсионном комплексе.
Максимальная высшая температура
Единичное самое высокое значение температуры тела, зафиксированное логгером у отдельной особи за время наблюдений (Tmh) (рис. 2). Для всех изученных животных можно определить абсолютную максимальную температуру (Tmha) – самое высокое значение из всех зафиксированных для всех особей, и среднюю высшую максимальную температуру (MTmh), рассчитанную как среднее из этих значений. Высшая максимальная температура имеет отчетливый смысл: эта температура неприемлема для особей, ee можно принять как примерную оценку последней витальной температуры. На диаграмме распределения значений температуры эта точка ограничивает последний частотный класс. Показатель для отдельной особи представлен одним числом и не имеет интервальной оценки, однако можно говорить о его высокой репрезентативности, поскольку эти значения получены из выборки объемом 15–25 тысяч измерений для отдельной особи.
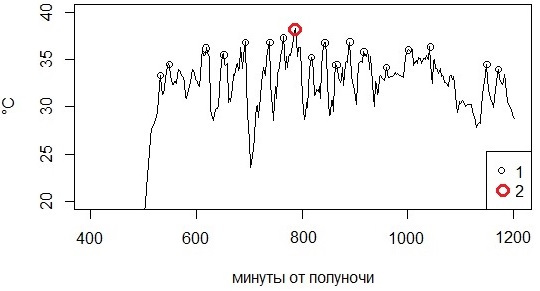
Рис. 2. Максимальные пиковые температуры (1) и максимальная высшая температура (2)
Fig. 2. Maximum peak temperatures (1) and maximum high temperature (2)
Средняя пиковая температура
Показатель, призванный дать хоть какую-нибудь статистическую характеристику значению максимальной температуры. На графике хода дневной температуры тела животного обычно обнаруживается достаточно много «пиков» – высоких значений температуры, после достижения которых температура тела падает (см. рис. 2). Зачастую это связано с переходом животного в полутень или легкие укрытия как способ избежать перегрева, хотя часть пиков вызвана прерыванием теплового потока от солнца облаками. Тем не менее эти пики могут говорить о достижении змеями предела комфортной температуры и включении механизма поведенческой терморегуляции. С этих позиций средняя пиковая температура обретает физиолого-экологический смысл границы толерантного диапазона. Для оценки этих значений требуется найти хорошо выраженные пики, ясно говорящие о смене тепловых потоков.
Поиск пиковых значений (локальных максимумов) велся программой по следующему алгоритму. Для каждой особи в течение одного дня анализировали ряды значений температуры с отсчетами через 2 мин. Для лага длиной от 3 до 15 соседних записей (от 6 до 30 мин.) рассчитывалась скользящая линейная регрессия зависимости температуры от номера отсчета на данном интервале. При этом положительные значения коэффициента регрессии говорят о росте температуры на анализируемом отрезке (наклон линии влево), а отрицательные – о снижении (наклон линии вправо); на пике происходит смена знака коэффициента регрессии, для этого отсчета и брали значение текущей температуры тела как пиковую. Многочисленные испытания показали, что при лаге длиной 10 отсчетов (20 мин.) идентифицируются все хорошо выраженные пики. Наборы пиковых значений служили для расчета статистических параметров – средней пиковой (максимальной) температуры (Tpi) и ее стандартного отклонения (sTpi) для отдельной особи. Эти параметры можно сравнивать между собой с помощью простых статистических методов. Всего обнаружено и включено в расчет 243 значения. Средняя пиковая температура составила 35.1 °С, т. е. в среднем на 1 °С выше, чем максимальная добровольная, это связано с тем, что в первые моменты после начала терморегуляторной реакции температура тела все еще поднимается (см. ниже).
Эти наблюдения позволяют по-новому использовать полевые замеры температуры тела гадюки. Количество пиков, обнаруженных на записи логгерами для 9–17 ч. при ясной солнечной погоде, составляет около 9 % от всех дневных замеров. Если взять 9 % наибольших значений от всех замеров, выполненных в полевых условиях при ясной солнечной погоде (и принять их за пиковые), то можно рассчитать среднюю полевую максимальную температуру тела и ее дисперсию. Объем выборки максимальных пиковых полевых значений из нашей базы данных составил 35 вариант, средняя – 32.5 °С, стандартное отклонение – 5.9 °С. Полевые замеры и замеры логгерами дали не очень близкие значения.
К сожалению, отдельные локальные максимумы температуры тела могут иметь различное происхождение. Помимо терморегуляторных поведенческих реакций пики порождают переменная облачность (прерывистый режим инсоляции) и порывы ветра (переменный режим принудительной конвекции). Таким образом, теоретически средняя пиковая температура должна иметь гораздо более широкую изменчивость, затрудняющую сравнение параметров.
Максимальная типичная температура
Типичная максимальная температура (Tmt) рассчитывается как некий предел в правой части полигона распределения значений температуры тела гадюки. На температурной шкале это такая точка, правее которой температура тела гадюки обычно не встречается. Мы стремимся придать этому показателю вероятностное содержание и, взяв на вооружение статистическое «соглашение о 95 %», отсечь от распределения 5 % «лишних» вариант. В распределении значений дневной температуры тела гадюки только правый скат похож на гауссиану. Иными словами, если в области высоких значений модальный класс (с наибольшей частотой вариант) принять как центральный условного нормального распределения, то на правой половинке можно выделить значение, отсекающее 5 % от этой выборки. Это и будет значение максимальной типичной температуры. Мы разработали три метода для оценки этого значения: метод квантилей (Tmtq), метод регрессии (Tmtr), метод квантилей по регрессии (Tmt).
Метод квантилей
В качестве исходных данных для каждой гадюки послужило распределение температуры тела в течение всех дней наблюдений. От этого распределения отбрасывали часть данных, находящуюся левее модального класса, который практически всегда приходился на 33–34 °С. Общий объем значений температуры тела выше 33 °С для разных змей варьировал от 140 до 2900 вариант, поскольку продолжительность наблюдений и частота считывания температуры тела в разных опытах различались. Далее определяли, сколько вариант составит 5 % от всей выборки (квантиль 5 %) (nq5 = n * 0.05, например, для 300 отсчетов nq5 = 300 * 0.05 = 15), все значения сортировали в порядке уменьшения и отыскивали значение температуры, которое в этом ряду стоит на nq5-месте (в примере – на пятнадцатом справа). Это и есть максимальное типичное квантильное значение. Дальнейший анализ показал, что при не очень больших выборках, во-первых, модальные значения определяются плохо, и исходный объем выборки точно не определить, во-вторых, максимальные значения склонны случайно варьировать, и, как следствие, изменение модального класса ведет к существенным отличиям полученных значений для разных змей. В то же время это самый простой и понятный метод.
Метод регрессии
В этом методе мы отошли от требования сохранять 95 % вариант выборки, ориентируясть только на форму распределения температуры тела. Для отдельной особи за весь период наблюдений распределение разбивали на интервалы шириной 0.5 °С, определяли модальный класс для области самых высоких температур, отбрасывали все значения левее и анализировали только правую часть распределения. Левая граница назначается как первый класс правее модального. Значение максимальной типичной регрессионной температуры количественно определяется как точка пересечения оси температур с линией тренда падения частот в правой ветви распределения (рис. 3). Эта точка отсекает правый «хвост», независимо от его веса в распределении. Технология состоит в построении линейной зависимости средней для класса температуры тела от частоты ее встречаемости: T’ = a + b * F (где T’ – расчетная температура в данном классе, F – частота в классе). Из уравнения следует, что температура равна коэффициенту a при частоте, равной 0; это и есть точка пересечения оси абсцисс линией регресии; искомый параметр (Tmtr) равен коэффициенту a. С целью уменьшить влияние отсекаемого правого «хвоста» распределения при расчете уравнения линейного тренда использовали взвешивание, чтобы наиболее репрезентативные значения температуры (из классов с большими частотами) получали преимущество перед классами с экстремально высокими температурами (но с малой частотой встреч); весами служили частоты в данном классе. В среде R функция записывается так: lm(t~f,weight=f).
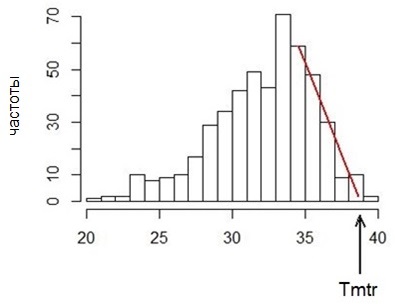
Рис. 3. Линия регрессии для правого ската частот
Fig. 3. Regression line for the right slope of frequency
Для оценки устойчивости полученных значений типичного максимума применена процедура рандомизации (Шитиков, 2014), которая состоит в многократном пересчете коэффициентов регрессии на ограниченных выборках, случайно взятых из исходной. Из этой совокупности данных для одной особи случайным образом извлекали вторичные выборки объемом по 500 значений температуры («с возвратом»), для которых строили гистограмму, подсчитывали частоты классов, строили уравнение регрессии (Т~n) и оценивали Tmtr. Для каждой особи выполняли по 100 оценок типичной максимальной температуры и рассчитывали, во-первых, среднее значение максимальной типичной температуры (Tmtr), во-вторых, стандартное отклонение (mTmtr ), которое имеет смысл ошибки средней.
Этот метод дает достаточно устойчивые результаты, если исходная выборка велика, но при небольших объемах в исходных распределениях намечаются провалы, и регрессионные оценки максимальной типичной температуры, во-первых, получают заниженные оценки (растут при увеличении выборок, рис. 4), во-вторых, ресамплинг дает большие значения ошибки.
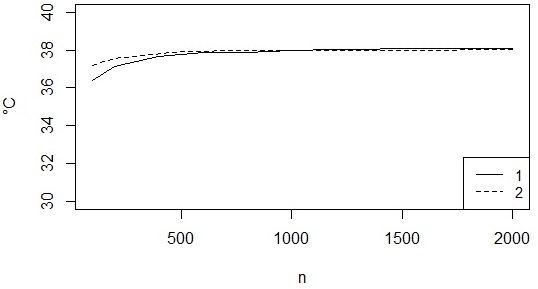
Рис. 4. Соотношение между оценками максимальной типичной (регрессионной) температуры для двух змей (1, 2) с объемами выборки (n)
Fig. 4. Relationship between estimates of the maximum typical (regression) temperature for two snakes (1, 2) with sample sizes (n)
Метод квантилей по регрессии
В поисках процедуры, сглаживающей перепады частот в соседних классах распределения температур, использовали кривую накопления частот, кумуляту. Предварительно исходное частотное распределение сглаживалось скользящей средней по тройкам, и в качестве модального выбирался класс, имеющий наибольшую сглаженную частоту. Далее в обработку включались значения высокой температуры тела, больше значений для модального класса. Для отобранных значений строилась гистограмма с 20 интервалами (шириной около 0.4–0.5 °С), затем этот ряд частот сортировался по возрастанию (i = 1, 2 …, 20 – номер интервала). На основе этого частотного ряда (fi) рассчитывался ряд накопленных частот (Σi), начиная с наименьших частот (от самых высоких значений температуры к меньшим); все значения делятся на общую сумму (pi = 100 * Σi / Σ20). Таким образом, для каждого интервала известен его номер (i), центральное значение температуры (Ti) и относительная накопленная частота (в процентах, pi). Используя эти значения, рассчитывалось уравнение гиперболической зависимости следующего вида: Ti = a1 + a2/pi + a3/(pi ^ a4), которое играет роль кривой сглаживания накопленных частот. В эту формулу подставляли значение 5 % и таким образом рассчитывали значение температуры, соответствующее 5%-й квантили (рис. 5); это и есть максимальная типичная (регрессионно-квантильная) температура (Tmtrq = Tmt). Для расчета ошибки параметра применяется процедура ресамплинга, описанного выше.
Полученные оценки для разных змей имеют гораздо меньшую изменчивость и уже не связаны с объемами выборок, хотя процедура расчета стала заметно сложнее.
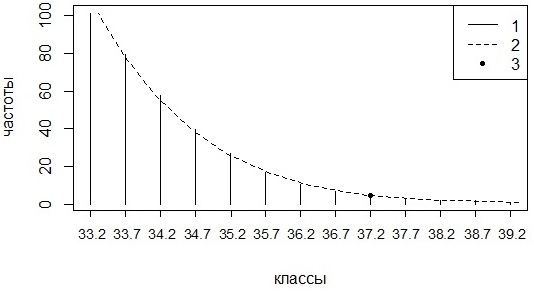
Рис. 5. Оценка температуры, соответствующей квантили 5 %. 1 – значения накопленных частот, 2 – регрессия, 3 –точка с координатой 5 % и 37.7 °С (Tmt = 37.7 °С)
Fig. 5. Estimation of temperature corresponding to quantile 5 %. 1 – values of accumulated frequencies, 2 – regression, 3 – point with coordinates 5 % and 37.7 °С (Tmt = 37.7)
Максимальная добровольная температура
Индивидуальная максимальная добровольная температура – это тот уровень температуры тела, при котором у конкретной особи включается поведенческая терморегуляция, направленная на избегание перегрева. Для определения этого параметра используются расшифровки круглосуточных видеозаписей поведения змей во время всего эксперимента. Алгоритм, реализованный в программе для автоматического определения параметра, следующим. В полной базе данных отыскивались периоды времени (продолжительностью 12 мин. или 6 соседних отсчетов времени), в течение которых змея нагревалась, а затем начинала остывать. Для этого рассчитывали скользящую линейную регрессию (зависимость температуры от номера шести соседних отсчетов 1, 2, ... 6). Смена знака коэффициента скользящей регрессии указывала на точку перегиба температурной кривой, т. е. на момент начала остывания. Далее по базе данных программа определяла, двигалась ли гадюка (код движения – 2) в этот период (12 мин.). Если животное не двигалось и не меняло позу, то принимали, что изменение температуры связано с погодными эффектами (ветер, тень от облака), и такие данные пропускали. Если же в течение этого интервала (6 отсчетов) змея перемещалась или меняла позу, то это поведение рассматривали как терморегуляторную реакцию, и значение температуры тела, соответствующее моменту начала движения, брали как оценку максимальной добровольной температуры, которое стимулирует осуществление поведенческого акта. Зачастую моменты начала движения и достижения пиковой температуры отстояли на 1–2 отсчета, т. е. после начала перемещений для поиска более прохладных условий температура тела змеи продолжала подниматься и формировала температурный пик, который явно является перегревом, но не вполне «добровольным перегревом» (Черлин, 2015). Всего для 6 гадюк было обнаружено 56 значений максимальной добровольной температуры. По всем этим значениям для данной особи рассчитывается значение «средняя максимальная добровольная температура и стандартное отклонение», пригодная для сравнения с другими особями. Это самый стабильный показатель, мало отличающийся у разных змей, средний уровень для гадюк из Карелии (N62) составил 34 °С.
Средняя температура баскинга
Этот показатель (Tb) рассчитывается как средняя арифметическая для всех значений температуры тела гадюки, которые зафиксированы за все моменты времени, когда температура окружающей среды (температура воздуха в траве) превышала 23 °С (рис. 6). Важно отметить, что температура в траве играет роль всего лишь индикатора высокого уровня солнечной радиации, при 23 °С вынуждающего змею осуществлять терморегуляторное поведение. Порог 23 °С принят только по той причине, что на диаграмме зависимости температуры тела от температуры субстрата (см. рис. 6) отчетливо виден изолированный «конус» точек, связанных, очевидно, только с терморегуляторным поведением змей. Именно этот показатель оказывается самым понятным для герпетологов, поскольку близок к часто используемым понятиям «оптимальная» или «предпочитаемая» температура в природных условиях.
К сожалению, в этот период не все змеи остаются на поверхности и прячутся в легкие укрытия, полутень. Поэтому изменчивость этого параметра оказалась довольно велика и значимо связана со средней температурой среды. Таким образом, данный показатель оказался термоэкологическим и не позволяет делать точные оценки физиологических предпочтений гадюки.
В то же время этот параметр можно усилить и уточнить, если его оценку проводить только для тех змей, которые находятся на поверхности субстрата и активно осуществляют терморегуляцию. С этой целью по результатам видеонаблюдения была составлена выборка для наблюдаемых змей и оценены статистические характеристики температуры их тела. Полученные оценки оказались очень близкими у разных особей, составили в среднем 33.1 °С, что почти равно максимальной добровольной температуре. Таким образом, этот параметр также является термофизиологическим, но трудоемким для оценки.
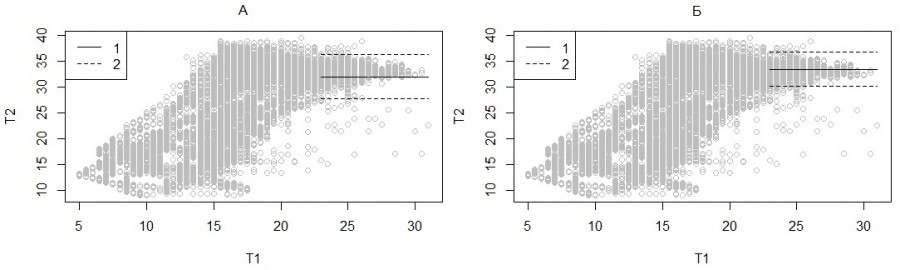
Рис. 6. Средняя температура баскинга (1) и ее стандартное отклонение (2) при температуре травы (T1) выше 23 °С (А), средняя температура баскинга под солнцем (1) и ее стандартное отклонение (2) (Б)
Fig. 6. Average temperature of basking (1) and its standard deviation (2) at grass temperature (T1) above 23 ° С (A), average temperature of basking in the sun (1) and its standard deviation (2) (Б)
Оценка суммы накопленных температур
Этот показатель призван характеризовать особенности использования теплоты среды разными особями, проживающими совместно в одних климатических условиях. Для унификации показаний, полученных разными логгерами, подсчитывали общую сумму значений температуры за весь день (с 8 до 18 ч.) для всех дней наблюдений и делили на общее количество отсчетов за это время. Получаем среднюю дневную температуру тела на один отсчет.
Медианные активные температуры
Показатель «сумма активных температур» (превышающих 10 °С) широко используется в агрономии для сопоставления условий теплообеспечения почв (Синицина и др., 1973). В отношении рептилий показатель призван характеризовать температуры тела змей, когда они находятся вне укрытий в дневное время, стремясь повысить температуру тела, поскольку именно в это время осуществляются основные метаболические реакции, обеспечивающие функционирование организма животного. Для гадюки 10 °С – это очень низкое значение, поэтому были приняты пороги 20, 25, 30 °С (Tact20, Tact25, Tact30). Техника расчетов состоит в том, чтобы сформировать выборку значений температуры тела выше порога и рассчитать медианное значение, имеющее смысл реального срединного значения из данной выборки (рис. 7).
Наши расчеты показали, что медианы для выборок с порогом 20 и даже 25 °С у разных змей имеют широкую изменчивость и, очевидно, несут смысл термоэкологических характеристик.
Медиана для выборок выше 30 °С достаточно стабильна, средняя равна 33.4 °С, т. е. близка по величине к максимальной добровольной температуре Tmg = 34 °С; ее вычисления просты, следовательно, Tact30 и может служить для оценки Tmg. На наш взгляд, эта температура вполне ответствует показателю В. А. Черлина (2012), температуре полной активности (поскольку при этих температурах змеи активно осуществляют терморегуляторное поведение), но более формализованно и точнее.
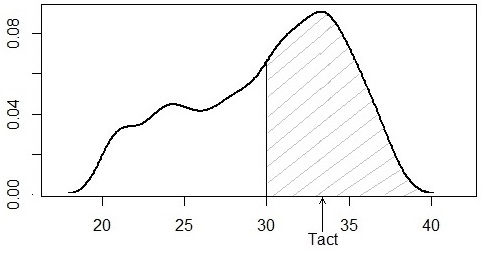
Рис. 7. Оценка медианной активной температуры обыкновенной гадюки при температуре тела 30 °С
Fig. 7. Estimation of the median active temperature of the common viper at temperatures above 30 °С
В такой форме данный показатель можно использовать для сравнения разных змей, изученных в данный сезон, а также для поиска зависимости между теплолюбивостью змей и их морфо-физиологическими характеристиками.
Средняя нормированная температура
Для получения температурных характеристик, которые можно сравнивать у животных, наблюдаемых в разных условиях (место, сезон, погода, год), был рассчитан показатель средней нормированной температуры. Смысл показателя состоит в выявлении индивидуальной теплолюбивости особи, отличающих ее от группы особей, живших с ней (рис. 8).
Этот показатель прост в вычислении. От текущей температуры тела, зафиксированной в данный момент, вычитали среднюю текущую температуру всех особей и делили разницу на стандартное отклонение: Tni = (Ti - Mi)/Si, далее все значения, полученные для конкретной особи, усреднялись. Таким образом, для каждой особи получали одно значение, которое может варьировать от -1 до 1 и характеризует большую или меньшую теплолюбивость этого животного. Препятствием для расчета средней нормированной температуры является отсутствие синхронных измерений температуры тела нескольких особей или их небольшое количество (менее трех).
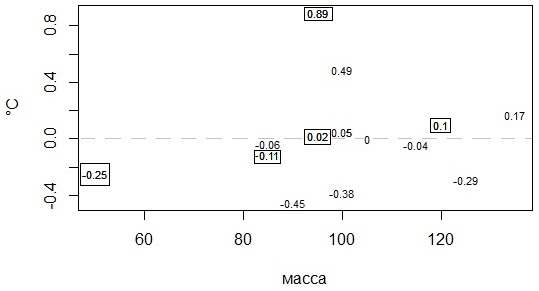
Рис. 8. Соотношение массы и средней нормированной температуры тела (в прямоугольниках – значения для самцов)
Fig. 8. The ratio of mass and average normalized body temperature (values for males are in rectangles)
Соотношение между параметрами терморегуляции
Исходя из физиологической теории регуляции, нами предложено несколько показателей, в той или иной мере приближающихся к «штатному» параметру терморегуляции, включающему терморегуляторные реакции животного (обыкновенной гадюки) (табл. 2, рис. 9). Вероятно, наиболее близки к нему показатели «максимальная добровольная температура» (Tmg) и «медианная активная температура выше для выборки выше 30 °С» (Tact30). В то же время показатель «максимальная типичная температура» (Tmt) хорошо статистически обоснован, а «средняя температура баскинга под солнцем» (Tbs) имеет понятное и привычное содержание. Для оценки работоспособности этих показателей необходимо расширить спектр объектов исследования.
Таблица 2. Величина и изменчивость термобиологических показателей гадюки обыкновенной
| Название | Ник | M | Sm | n |
| Нормированное время выхода | t | 0 | 0.68 | 16 |
| Скорость нагревания | V | 0.8 | 0.3 | 16 |
| Сумма температур | S | 23.4 | 3.37 | 23 |
| Максимальная высшая | Tmh | 39.4 | 1.05 | 23 |
| Максимальная пиковая | Tmp | 35.1 | 0.21 | 7 |
| Максимальная типичная | Tmt | 37.8 | 0.74 | 23 |
| Максимальная добровольная | Tmg | 34 | 0.61 | 6 |
| Средняя температура баскинга | Tb | 32.3 | 2.7 | 22 |
| Средняя температура баскинга под солнцем | Tbs | 33.1 | 0.9 | 6 |
| Медианная активная температура для выборки выше 20 °С | Tact20 | 30.3 | 1.5 | 23 |
| Медианная активная температура для выборки выше 25 °С | Tact25 | 32.3 | 0.8 | 23 |
| Медианная активная температура для выборки выше 30 °С | Tact30 | 33.4 | 0.5 | 22 |
| Средняя нормированная температура | Tn | 0 | 0.34 | 16 |
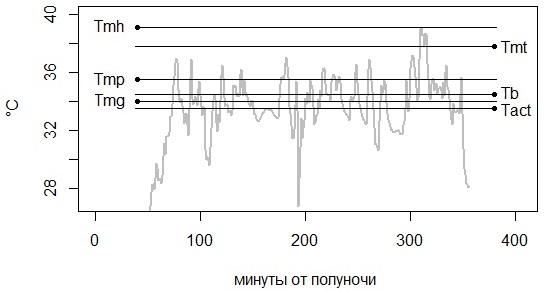
Рис. 9. Соотношение терморегуляторных параметров на фоне типичной траектории дневной температуры тела гадюки
Fig. 9. Ratio of thermoregulatory parameters against the background of the typical trajectory of the daytime body temperature of the viper
Заключение или выводы
Описывая термобиологические особенности рептилий, необходимо иметь четкое представление о тех параметрах и характеристиках, которые выбираются в качестве основных показателей. Это представление должно быть как биологическим (иметь явный физиологический или экологический смысл), так и математическим (для понимания степени значимости полученных значений).
Среди рассмотренных нами показателей наиболее репрезентативным является максимальная типичная температура. Несмотря на сложность расчетов, точность и устойчивость этого показателя позволяет проводить объективные сравнения максимальной типичной температуры у особей из разных ареалов и межвидовые сравнения. Кроме статистической достоверности, биологический смысл такой температуры – верхний порог толерантности, т. е. максимальная температура тела, выше которой гадюки нагреваются в крайне редком случае.
Биологически ясный основной параметр терморегуляции (максимальная добровольная температура) описывает схему поведенческой терморегуляции гадюки – избегание перегрева. Сложность в определении максимальной добровольной температуры вызывает необходимость искать другие, близкие к ней температурные характеристики, но более простые для определения и требующие меньше трудозатрат для расчета. Таким показателем оказалась медианная активная температура для выборки выше 30 °С.
При выборе температурных показателей рекомендуем использовать те, которые имеют низкую изменчивость и отчетливый биологический смысл.
Библиография
Коросов А. В., Ганюшина Н. Д. К оценке максимальной добровольной температуры обыкновенной гадюки // Известия высших учебных заведений. Поволжский регион. Естественные науки. Пенза, 2019. № 2. С. 96–104. DOI: 10.21685/2307-9150-2019-2-10.
Коросов А. В., Ганюшина Н. Д. Максимальная температура тела как параметр терморегуляции рептилий: опыт статистической оценки на примере обыкновенной гадюки (Vipera berus (L. 1758)) // Зоологический журнал. 2021. Т. 100. № 1. С. 1–10.
Коросов А. В. Двухконтурная отрицательная обратная связь и модель терморегуляции гадюки // Ученые записки Петрозаводского государственного университета. Сер. Естественные и технические науки. 2008. № 1. С. 74–82.
Коросов А. В. Экология обыкновенной гадюки (Vipera berus L.) на Севере (факты и модели) . Петрозаводск: Изд-во ПетрГУ, 2010. 264 с.
Коросов А. В. Новое слово в термобиологии рептилий? // Принципы экологии. 2015. № 1. С. 77–85. DOI: http://dx.doi.org/10.15393/j1.art.2015.4162.
Синицина Н. И., Гольцберг И. А., Струнников Э. А. Агроклиматология . Л.: Гидрометеоиздат, 1973. С. 40–42.
Слоним А. Д., Слоним А. Д. Экологическая физиология животных . М.: Высшая школа, 1971. 448 с.
Черлин В. А. Термобиология рептилий. Общие сведения и методы исследований (руководство) . СПб.: Русско-Балтийский информационный центр «БЛИЦ», 2010. 124 с.
Черлин В. А. Изучение термобиологии рептилий в мире и в СССР/РФ. 1. Изучение термобиологии рептилий в зарубежных странах // Принципы экологии. 2019а. № 3. С. 163–179. DOI: 10.15393/j1.art.2019.8963.
Черлин В. А. Изучение термобиологии рептилий в мире и в СССР/РФ 2. Изучение термобиологии рептилий в СССР/РФ // Принципы экологии. 2019б. № 4. С. 96–132. DOI: 10.15393/j1.art.2019.8964.
Шмидт-Ниельсен К. Физиология животных. Приспособление и среда . М.: Мир, 1982. Кн. 1. 416 c. Кн. 2. 384 c.
Шитиков В. К., Розенберг Г. С. Рандомизация и бутстреп: статистический анализ в биологии и экологии с использованием R . Тольятти: Кассандра, 2014. 314 с.
Herczeg G., Saarikivi J., Gonda A., Pera J. la, Tuomola A., Merila J. Suboptimal thermoregulation in male adders (Vipera berus) after hibernation imposed by spermiogenesis // Biological Journal of the Linnean Society. 2007. Vol. 92. P. 19–27. DOI: 10.1111/j.1095-8312.2007.00827.x.
Lourdais O., Guillon M., DeNardo D., Blouin-Demers G. Cold climate specialization: adaptive covariation between metabolic rate and thermoregulation in pregnant vipers // Physiol Behav. 2013. Vol. 119. P. 149–155. DOI: 10.1016/j.physbeh.3013.05.041.
R Core Team (2012). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL: http://www.R-project.org/.

 © 2011 - 2026
© 2011 - 2026