



Введение
Анализ донных сообществ на региональном (субконтинентальном) уровне основан на исследовании речных сетей, т. е. взаимосвязанных совокупностей лотических объектов, размещенных на площади до нескольких тысяч квадратных километров (часто в разных природно-климатических зонах). Изучение таких крупномасштабных экосистем, включающих десятки и сотни локальных сообществ, функционирующих в неоднородных условиях среды, целесообразно проводить на основе современной концепции метасообщества (metacommunity concept) (Leibold et al., 2004; Holyoak et al., 2005), которая позволяет моделировать закономерности пространственного распределения видового состава.
Любая река по-своему уникальна, поэтому условия существования локальных сообществ и видовой состав гидробионтов в каждой из них могут существенно различаться (Алимов, 2001). Это подтверждают исследования на уровне отдельной малой или средней реки (в линейном масштабе от 10 до 300 км), связанные с моделированием распределения таксономической структуры гидробионтов по продольному профилю водотока под влиянием прерывистого градиента гидрологических условий и качества водной среды (Шитиков, Зинченко, 2014). В более крупном географическом масштабе донные сообщества рек обнаруживают специфические пространственные тенденции изменчивости, которые обусловлены воздействием климатических, ландшафтных или геоморфологических факторов и выявляются в ходе проведения комплексных биосферных исследований.
Пространственное распределение отдельных видов, как правило, автокоррелировано: появление таксона в некоторой точке увеличивает вероятность его обнаружения в соседних экотопах. Действительно, планктонные и бентосные организмы способны мигрировать наземным путем либо перемещаться по градиенту течения в пределах речной сети на сотни километров. Отмечено, что потоки миграции инвазивных видов могут быть в известной степени стационарными, и таксономическая структура локальных ценозов в зоне расселения чужеродных организмов может приобрести дополнительное сходство (Курина, Селезнев, 2019). Кроме того, в рамках крупного региона часто удается выделить относительно однородные области с одинаковыми физико-географическими условиями, составом ландшафтных элементов или уровнем антропогенного воздействия, что приводит к сходству видовой структуры локальных сообществ (Manni et al., 2004). Наконец, для каждой таксономической группы объективно существует некоторый географический градиент, относительно которого встречаемость отдельных видов может статистически значимо изменяться (см., например, теорию изменения широтного разнообразия – Koleff, Gaston, 2001).
В большинстве ситуаций монотонные тренды или однородные кластеры можно выделить, ориентируясь на распределение обобщенных показателей структурного и функционального разнообразия метасообществ. Видовое разнообразие обычно оценивается с помощью совокупности индексов гетерогенности, сочетающих в себе богатство видов и соотношение вероятностей их присутствия, а также индексов выравненности, которые были разработаны в попытке выделить компонент эквитабельности сообществ в отдельное измерение (Pielou, 1975; Magurran, 2004). Объективная необходимость учета филогенетических различий между видами обусловила расширение понятия разнообразия и появление целой коллекции таксономических и филогенетических индексов (Faith, 1992; Ricotta, 2005; Chao et al., 2010; Kembel et al., 2010; Шитиков, Зинченко, 2013а, 2013б). Наконец, возникло понимание важности выделения комплекса редких видов как одного из главных компонентов разнообразия, что предопределило разработку индексов редкости (Gaston, 1994; Leroy et al., 2012).
Для проверки предположения о существовании пространственных закономерностей изменения видового состава и разнообразия изучаемых рек используется концепция нулевых моделей, утверждающих, что распределение таксонов по участкам случайно и хаотично, кластеры не образуются, влияние экологических градиентов отсутствует и каких-либо согласованных связей между видами не обнаруживается (Hausdorf, Hennig, 2007). Нуль-модель являлась эталоном, с которым сравниваются модели, основанные на эмпирических данных, где предполагается наличие пространственной автокорреляции, основанной на близости географических координат.
Все эти гипотезы нуждаются в тщательной проверке с использованием репрезентативного массива натурных исследований. Таковыми являются результаты многолетней гидробиологической съемки донных сообществ малых и средних рек на территории Среднего и Нижнего Поволжья. Ранее (Golovatyuk et al., 2018) были сформированы списки индикаторных видов, показывающие закономерную смену ведущих таксонов при смене природно-климатических поясов в последовательности водохранилищ Волжского каскада. Однако за рамками этой работы осталась такая важнейшая проблема биоиндикации, как оценка качества вод и таксономическое разнообразие изученных лотических объектов.
Настоящая работа носит как прикладной, так и методический характер. С одной стороны, приводятся подробные графические и табличные данные о пространственном распределении различных индексов видового разнообразия и отдельных таксонов локальных донных сообществ бассейна Средней и Нижней Волги. Проверяются научные гипотезы о статистической зависимости этих показателей от географических координат, показана их изменчивость между природно-климатическими зонами. С другой стороны, нами представляется набор алгоритмов тестирования нуль-моделей в геоинформатике и различные примеры тематических экологических карт, которые являются важнейшими инструментами анализа процессов в метасообществе на видовом и ценотическом уровнях, а в дополнительном приложении к статье представлены компьютерные модули их построения (Шитиков, 2020).
Материалы
Исследования выполнены на основе результатов гидробиологического мониторинга донных сообществ бассейна Средней и Нижней Волги (Зинченко, 2011) в разные месяцы вегетационного периода 1990–2019 гг. Гидробиологическую съемку макрозообентоса проводили на 90 малых и 12 средних равнинных реках, притоках Куйбышевского, Саратовского и Волгоградского водохранилищ, в том числе на 6 реках аридного региона бассейна оз. Эльтон. Образцы макрозообентоса (1401 проба) собирали в рипали и медиали рек дночерпателем Экмана – Берджи или гидробиологическим скребком. Фиксацию организмов и последующую камеральную обработку собранного материала проводили согласно общепринятым методикам (Зинченко, 2011). Всего было выявлено S = 740 видов и таксонов бентоса рангом выше вида.
Методы
С целью обобщения материала 245 станций 12 средних рек были разделены на приблизительно однородные участки: верхнее, среднее, нижнее течение и устье, а каждая малая река принималась как целостный объект. Таким образом, было выделено 132 локальных сообщества, в каждом из которых было взято в среднем по 10 проб (от 2 до 137). Для дальнейшего анализа была сформирована обобщенная таблица «132 участка рек» – «740 видов», которая включала встречаемость и среднюю численность каждого вида в каждом сообществе.
Главной проблемой корректной оценки сходства таксономического состава двух произвольных участков рек явилось неравное число взятых проб. Для выравнивания выборочного усилия выполнялась следующая нормализующая процедура:
1) подсчитывалось среднее число видов Sср, обнаруженных в одной пробе на каждом участке рек (по всем пробам Sср = 9.548 ± 0.170);
2) подсчитывалась встречаемость каждого вида, полученный ряд ранжировался по убыванию;
3) в качестве нормализованного видового состава для каждого участка принимался список из Sср таксонов с максимальной встречаемостью.
После процедуры нормализации рассчитывалась матрица 132 × 132 таксономических расстояний на основе меры Брея – Кёртиса с использованием значений относительной встречаемости видов на каждом участке рек.
Гипотеза о значимости статистической связи между таксономическим составом водотоков и их пространственным расположением проверялась на основе матричного коэффициента линейной корреляции Мантеля (quadratic assignment) (Mantel, 1967; Шитиков и др., 2005). Дополнительно использовался тест на кластеризацию, проверяющий гипотезу о существовании положительных ассоциаций видов, который проводили на основе параметрического бутстрэпа матрицы дистанций между реками (Hennig, Hausdorf, 2004).
Для анализа пространственного распределения индексов видового разнообразия или численности отдельных таксонов геоинформационными методами выполняли построение нескольких типов карт:
– точечных карт, на которых значение показателя в каждой точке показано различным размером и/или цветом;
– мозаичных карт, состоящих из шестиугольных элементов различных цветов (Hexbin map);
– карт плотности статистического распределения вероятности обнаружения изучаемого таксона;
– карт интенсивности отображаемого показателя Z на основе его пространственной интерполяции (кригинга).
Подробно откомментированные скрипты кодов на языке R и полный комплект исходных данных представлены дополнительными материалами к статье (Шитиков, 2020), с помощью которых можно воспроизвести большинство расчетов и рисунков.
Для анализа расширенного набора показателей биологического разнообразия использовали метод и R-функцию DER (Diversity, Evenness, Rarity) (Guisande et al., 2017), включенную в пакет EcoIndR и выполняющую вычисления по трем последовательным этапам:
– расчет комплекта индексов, оценивающих разнообразие по различным категориям: богатство видов, их редкость, выравненность и таксономическая сложность сообществ;
– селекция наиболее информативной комбинации четырех индексов из различных разделов, которые обладают максимальной разрешающей способностью в условиях выполненного исследования;
– построение диаграммы ординации объектов в полярных координатах.
Был рассчитан 31 индекс разнообразия из разных категорий (оценку функционального разнообразия не проводили из-за отсутствия необходимых данных). Для расчета индексов филогенетического и таксономического разнообразия по каждому из 740 видов выполнили систематическое описание по 11 классификационным уровням вплоть до типа гидробионтов (Шитиков, Зинченко, 2013а). На этой основе последовательно формировались матрица таксономических дистанций между каждой парой видов и филогенетическое дерево в формате Newick.
Для оценки индекса относительной редкости (Leroy et al., 2012) каждому i-му виду с частотой встречаемости Qi был поставлен в соответствие вес редкости (rarity weight):
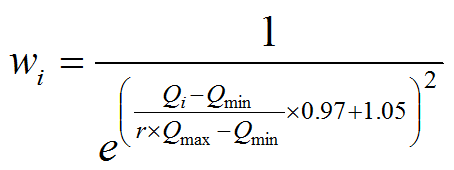
где Omin = 1, Qmax = 448 – минимальная и максимальная встречаемость видов в 1401 пробе, r = 0.01 – порог редкости, принимаемый из эвристических соображений (доля от общей встречаемости, ниже которой вид признается редким). Величина весов уменьшалась по экспоненте от wi = 1 для видов, встретившихся только один раз, до wi = 0.002 при Qi = 7 и далее равна 0. Индекс относительной редкости Irr для локального сообщества вычисляли как сумму весов редкости, нормированную на число видов в сообществе.
Ссылки на публикации, где описан метод вычисления каждого индекса разнообразия, приведены в таблице, а их расчетные формулы – в документации на пакет EcoIndR (Guisande C., 2017. Ecological indicators. R Package Version 1.0. https://cran.r-project.org/web/packages/EcoIndR).
Анализ совокупности вычисленных индексов разнообразия и селекцию наиболее информативной комбинации из них выполняли с использованием следующей процедуры (Guisande et al., 2017):
– индексы преобразовывались в единую шкалу от 0 до 1 и сканировались все возможные их комбинации по четыре (т. е. по одному индексу из групп редкости, гетерогенности, выравненности и филогенетической сложности);
– для каждой комбинации индексов рассчитывались полярные координаты X = ∑|zi| cos(αi) и Y = ∑|zi| sin(αi) каждого водотока, где zi – стандартизированное значение индекса, j = 1, 2, 3, 4, αi - угол, зависящий от zi;
– вычислялись площади выпуклой оболочки и средние евклидовы расстояния между реками в координатах X–Y, в качестве наиболее информативного набора индексов принималась комбинация, доставляющая максимум этим значениям.
Вычисления проводили с использованием среды программирования R версии 3.6 и ее дополнительных пакетов vegan, prabclus, Rarity и EcoIndR.
Результаты
Статистический и картографический анализ пространственного распределения таксономического состава гидробионтов
Для оценки статистической связи между пространственным расположением и видовым составом макрозообентоса сопоставлялись две матрицы 132 × 132 дистанций между каждой парой рек: а) географических расстояний (км) и б) таксономических расстояний на основе меры Брея – Кёртиса. Ядерная оценка функции двумерной плотности распределения значений обоих матриц представлена на рис. 1: прямая линия регрессии очевидно свидетельствует о том, что при увеличении географических расстояний статистически значимо возрастает удаленность в многомерном пространстве между комплексами видов (р ~ 0 для углового коэффициента линейной модели, коэффициент детерминации R2 = 0.097).

Рис. 1. График ядерной оценки двумерной плотности распределения таксономических и географических расстояний. Линия регрессии показывает увеличение различий в видовом составе рек с ростом удаленности между ними
Fig. 1. Graph of two-dimensional kernel density estimation of taxonomic and geographical distances. The regression line shows an increase in differences in the species composition of rivers with an increase in the distance between them
На основе коэффициента матричной корреляции Мантеля оценивали тесноту и статистическую значимость связи между двумя многомерными структурами данных. С помощью рандомизационной процедуры проверяли гипотезу Н0 о том, что расстояния между объектами в географической и таксономической матрицах независимы друг от друга. В ходе циклического процесса из 999 повторностей значения исходных матриц случайным образом перемешивались, и рассчитывалось имитируемое распределение статистики Мантеля при справедливости нулевой гипотезы. Ни одно значение коэффициента Мантеля, полученное в процессе рандомизации, не превысило эмпирическую величину r = 0.284, т. е. с уровнем значимости p = 0.001 можно утверждать, что корреляция между видовым составом и пространственным расположением биотопов статистически значима и не объясняется случайными причинами.
Статистический критерий pdisj (Hennig, Hausdorf, 2004), названный авторами вероятностью объединения (probability of disjunction), оценивает вероятность того, что объединяться в кластер будут объекты, пространственно достаточно удаленные друг от друга. Простейшая неограниченная (unconstrained) нулевая модель предполагала, что все биотопы рек одинаково пригодны или доступны для заселения всех видов независимо от того, в каких местообитаниях они были обнаружены; эта вероятность равна pdisj = 0.476 (т. е. равновероятен исход, близки объекты друг от друга или далеки). Если включить в модель в качестве ограничений географические координаты, то эта вероятность падает до pdisj = 0.2, хотя и не достигает общепринятого порога статистической значимости.
Одной из основных задач биогеографии является выявление регионального тренда изменения видового разнообразия. Естественный подход заключается в попытке аппроксимации данных о числе выявленных видов S (рис. 2) простейшей моделью – плоскостью относительно географических координат X–Y.
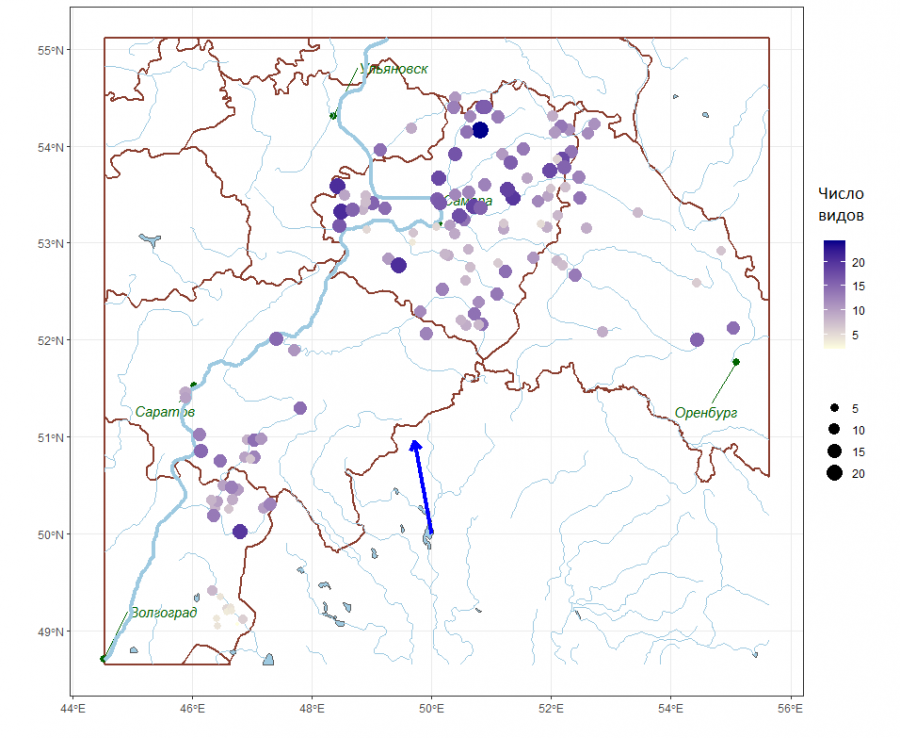
Рис. 2. Распределение видового богатства, оцениваемое по среднему числу видов в пробе (стрелкой показан градиент увеличения числа видов)
Fig. 2. Distribution of species richness estimated by the average number of species in the sample (the arrow shows the gradient of the increase in the number of species)
Для оценки статистической значимости линейного пространственного тренда выполняли следующий пермутационный тест. Аппроксимировали зависимость среднего числа видов S от географических координат X–Y плоскостью:
S = -53.5 - 0.52X + 1.73Y (R2 = 0.191, p = 0.00001).
Находили эмпирическое значение тангенса угла наклона k = 1.81 построенной плоскости относительно XOY. Формировали 999 рандомизированных выборок, в каждой из которых значения числа видов S были случайным образом перемешаны относительно строк X–Y, после чего строили статистическое распределение угла наклона плоскости тренда при справедливости нулевой гипотезы. Средняя величина тангенса угла между аппроксимирующими плоскостями, построенными на имитируемых данных, и плоскостью XOY равна m = 0.39 при стандартном отклонении s = 0.25. Поскольку ни одно из 1000 имитируемых значений не оказалось больше, чем эмпирическое значение, то нулевая гипотеза об отсутствии линейного пространственного тренда отклонена с достигнутым уровнем значимости р = 0.001.
Другим вариантом оценки регионального тренда является аппроксимация пространственного распределения анализируемого показателя нелинейными поверхностями. На рис. 3 показано направление градиента биоразнообразия, вычисляемого по индексу Шеннона, с использованием полиномиальной поверхности 2-й степени.
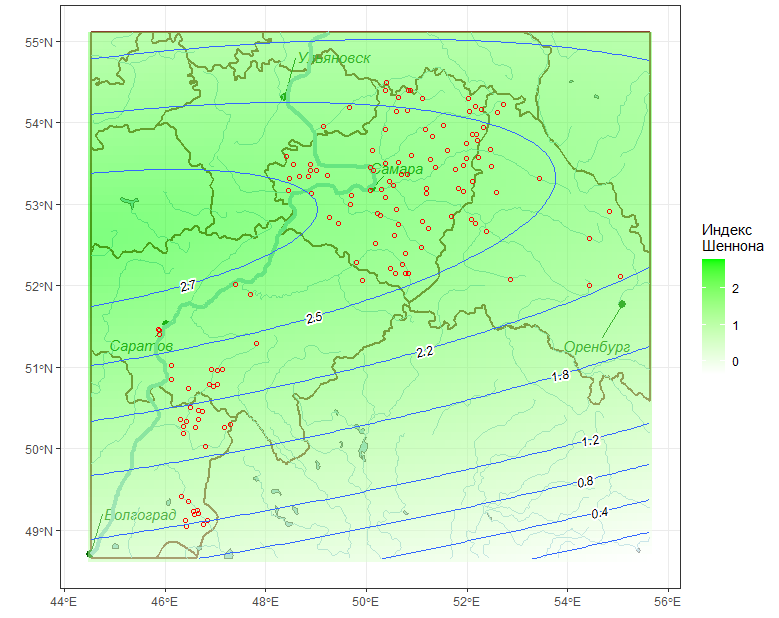
Рис. 3. Полиномиальный тренд пространственного распределения индекса Шеннона
Fig. 3. Polynomial trend of the spatial distribution of the Shannon index
Представляет интерес рассмотреть пространственное распределение рек с различными значениями биотических показателей, используемых для оценки качества вод. Одним из них является, например, показатель EPT, основанный на подсчете численности трех групп гидробионтов Ephemeroptera, Plecoptera и Tricoptera по отношению к общей численности бентоса (%). Считается, что снижение величины индекса отражает реакцию сообщества на загрязнение. Распределение значений индекса представлено на рис. 4 в виде мозаичной карты, на которой каждый участок показан гексагональными ячейками различного цвета.
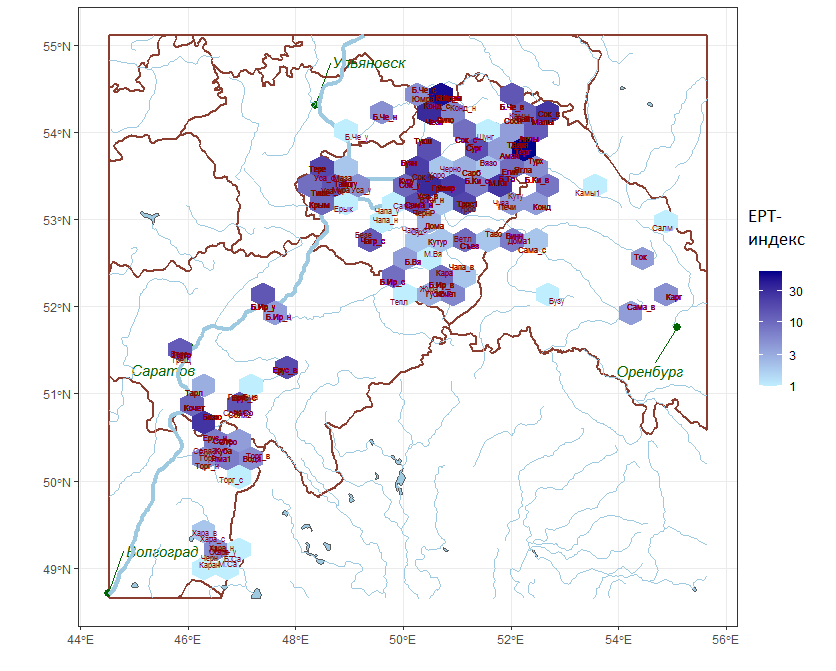
Рис. 4. Пространственное распределение значений индекса качества вод EPT (%)
Fig. 4. Spatial distribution of EPT water quality index (%)
Связь между видовой структурой и географическим местоположением локальных сообществ может проявляться как в форме изменчивости общего богатства видов, так и в появлении ассоциаций видов, не обязательно доминирующих, но характерных для групп пространственно близких рек. Для анализа выраженности этих зависимостей выполняли построение серий тематических карт различных типов, таких как плотности статистического распределения (см. рис. 5 – изолинии вероятности обнаружения в пробе эврибионтного вида хирономид Procladius ferrugineus) или пространственной интерполяции значений анализируемого показателя Z. На рис. 6 показаны изолинии средней численности в пробе реофильных хирономид подсемейства Prodiamesinae, полученные с использованием ординарного кригинга (Шитиков, 2020).
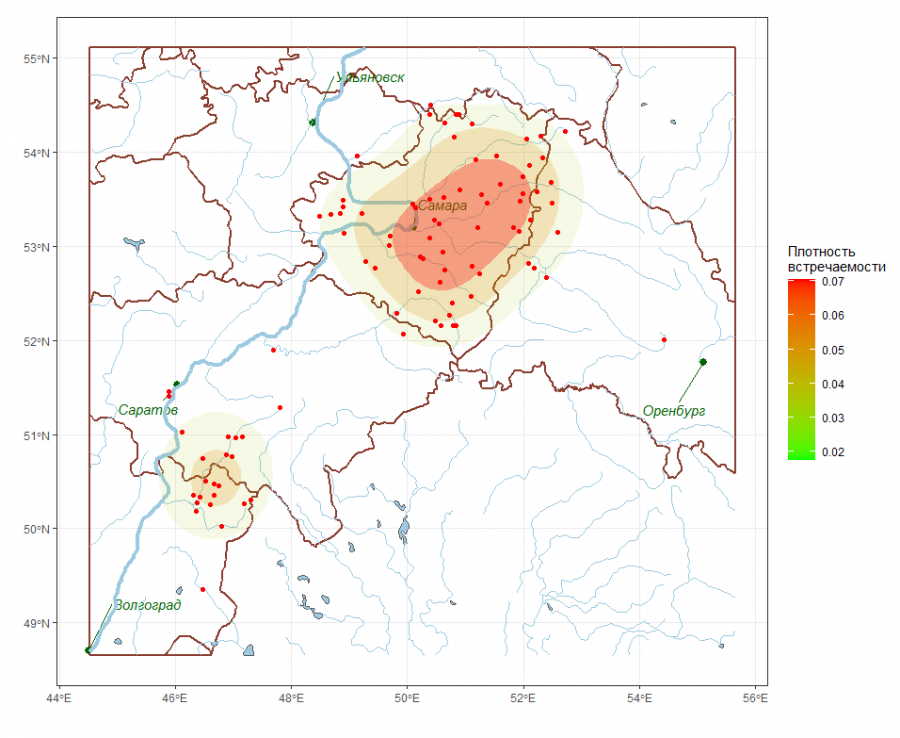
Рис. 5. Плотность распределения вероятности встречаемости Procladius ferrugineus
Fig. 5. Density of probability distribution of occurrence of Procladius ferrugineus
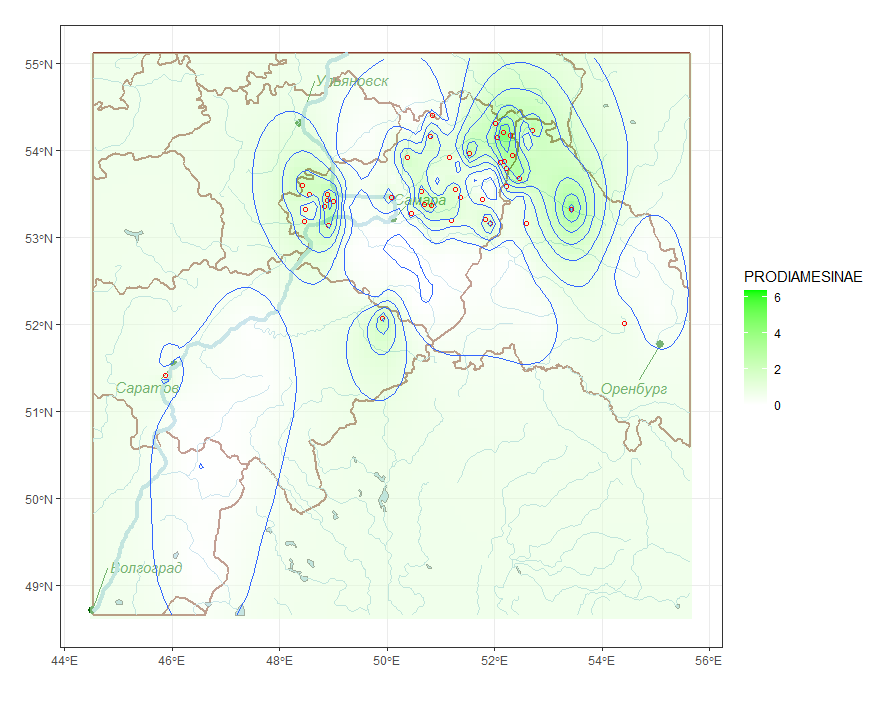
Рис. 6. Пространственное распределение численности хирономид подесемейства Prodiamesinae
Fig. 6. Spatial distribution of the abundance of chironomids subfamily Prodiamesinae
Компоненты биоразнообразия и характер их изменения
Для каждой из 1401 гидробиологической пробы макрозообентоса вычислен 31 наиболее часто используемый индекс разнообразия (таблица), из которых 2 связаны с редкостью, 12 – с видовой гетерогенностью, 7 – с равномерностью обилия, 2 – с таксономическим разнообразием, 8 – с филогенетической дивергенцией. В дальнейшем последние две группы объединяются в одну.
Средние значения индексов биоразнообразия макрозообентоса в реках бассейнов Куйбышевского (Куйб.), Саратовского (Сар. лс – лесостепная и Сар. ст – степная зоны), Волгоградского (Волг.) водохранилищ и оз. Эльтон
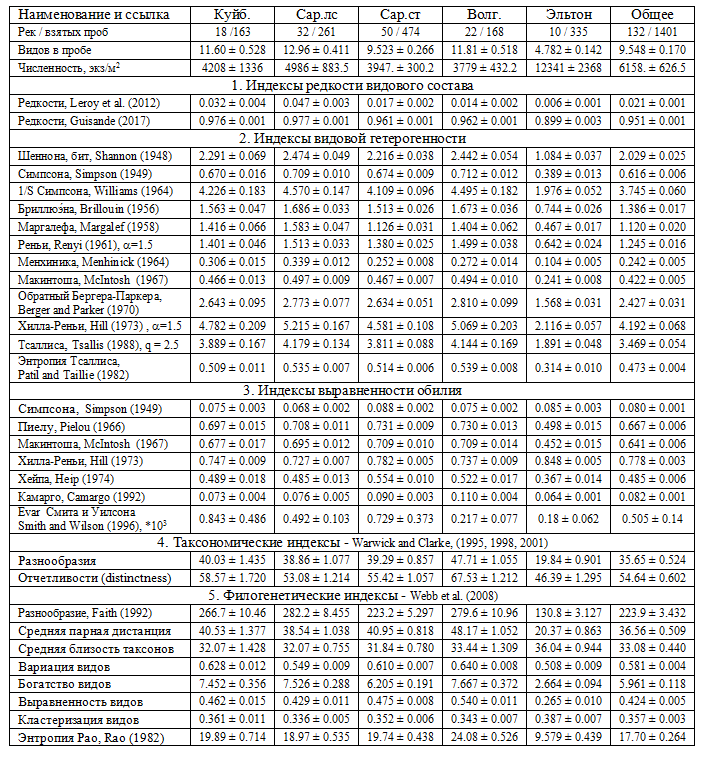
Примечание. В полной форме ссылки на публикации даны в статье (Guisande et al., 2017).
С использованием всей совокупности индексов разнообразия рассчитана матрица нормированных евклидовых расстояний между каждой парой участков рек. Найдена статистически значимая корреляционная связь между этой матрицей и пространственным расположением биотопов (т. е. географическим расстоянием между участками) (см. рис. 1): коэффициент матричной корреляции Мантеля r = 0.175, p = 0.001.
Для решения вопроса о том, какие показатели разнообразия наиболее информативны в условиях выполненного исследования, находили такой набор индексов, который приводил к наибольшей вариации рек, т. е. обладал максимальной разрешающей способностью. Была отобрана следующая оптимальная комбинация индексов:
– индекс относительной редкости Лероя (Rarity.Leroy) – см. выше;
– индекс разнообразия Менхиника (Menhinick):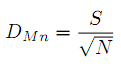
– индекс гомогенности распределения Макинтоша (McIntoshE):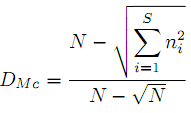
– квадратичная энтропия Рао (raoD):

где S и N – число видов и суммарная численность бентоса в пробе, ni – численность i-го вида, pi = ni/N, dij – расстояние на филогенетическом дереве между видами i и j.
В качестве общего показателя разнообразия рассматривалось среднее значение этих четырех нормированных индексов. Его пространственное распределение на картосхеме региона оказалось во многом идентичным рис. 2–3, хотя имело более сглаженный характер. Выполненный дисперсионный анализ значений обобщенного индекса для разных бассейновых зон (см. таблицу) отклонил нулевую гипотезу о равенстве групповых средних с высоким уровнем значимости (F = 9, p = 0.00002). На основании линейной модели можно сделать вывод, что совокупное биоразнообразие донных сообществ снижается в ряду водохранилищ:
Саратовское (лесостепь) > Куйбышевское > Саратовское степь > Волгоградское > оз. Эльтон.
Поскольку для отдельных сообществ могут быть высокие значения одних индексов и низкие других, то рекомендуется (Guisande et al., 2017) проводить анализ диаграммы в полярных координатах (рис. 7), с помощью которой можно детально оценить различия между реками в терминах редкости, гетерогенности, равномерности и таксономического/филогенетического разнообразия.
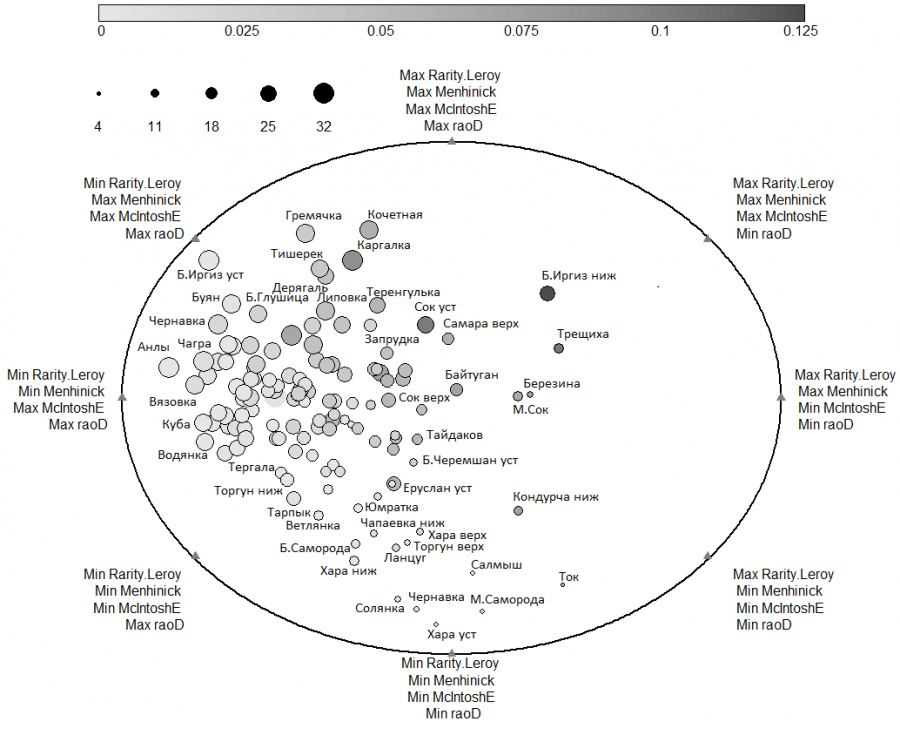
Рис. 7. Диаграмма, полученная в результате использования алгоритма DER на примере донных сообществ Волжского бассейна. Шкала размеров «пузырьков» связана с квадратичной энтропией Рао (raoD). Градиент серого цвета указывает на величину относительного показателя редкости (Rarity.Leroy) в каждой реке. Menhinick – индекс разнообразия Менхиника, McIntoshE – индекс гомогенности Макинтоша
Fig. 7. Diagram obtained as a result of using the DER algorithm on the example of bottom communities of the Volga basin. The bubble size scale is related to the Rao quadratic entropy (raoD). The gray gradient indicates the value of the relative rarity (Rarity. Leroy) in each river. Menhinick – Menhinik diversity index, McIntoshE – Macintosh homogeneity index
Ось ординат на рис. 7, проходящая через центр эллипса, совпадает с градиентом увеличения общего разнообразия по всем четырем индексам. Область справа от нее показывает реки, для которых в большей степени характерны редкие виды, а слева – реки с выраженной вариацией таксонов по филогенетическому дереву (последний показатель, оцениваемый по энтропии Рао, является превалирующим для изученной экосистемы).
Обсуждение
Природно-климатическая и ландшафтная неоднородность изучаемого региона обусловили пространственную автокорреляцию локальных донных сообществ, т. е. наблюдения в точках, расположенных близко друг к другу, вероятнее всего, будут более сходными, чем на участках, расположенных далеко друг от друга. Этот ожидаемый эффект иллюстрируется на рис. 1, а его статистическая значимость подтверждена тестами матричной корреляции Мантеля и пространственного объединения Хеннига – Хаусдорфа.
Какие паттерны являются следствием наблюдаемой автоковариационной структуры – монотонная зависимость или в некотором смысле «хаотические» кластеры, осталось в полной мере неясным. Значимый линейный градиент изменения видового богатства на рис. 2 формально противоречит гипотезе о глобальном направлении широтного градиента разнообразия (Koleff, Gaston, 2001). Однако подобная обратная закономерность была неоднократно описана на примере исследований рыб (Macpherson, Duarte, 1994; Oberdorff et al., 1995), морских моллюсков (Rex et al., 1993; Roy et al., 1998), донных беспозвоночных озер (Bezmaternykh, Vdovina, 2020). Изменения в составе фауны изученного региона при продвижении от севера к югу вызваны, в частности, тем, что в связи с уменьшением уклонов равнинных рек и скорости их течения из состава донных сообществ постепенно выпадают стенобионтные и реофильные таксоны, как это было показано нами для видов подсемейства Prodiamesinae. Кроме того, для аридных регионов бассейна Нижней Волги характерны реки с высокой минерализацией (Golovatyuk et al., 2018), что также приводит к выпадению многих таксонов, функционально не адаптированных к экстремальным условиям среды обитания (Golovatyuk, Shitikov, 2016).
Однако предположение о линейном тренде может оказаться достаточно грубым приближением для интерпретации. В частности, полиномиальная поверхность пространственного распределения на рис. 3 выполняет более точную аппроксимацию видового разнообразия. К аналогичным выводам приводит линейная модель дисперсионного анализа при группировке по водохранилищам, согласно которой наибольшее биоразнообразие наблюдается в районе лесостепной зоны Саратовского водохранилища.
Для оценки качества вод по индексу EPT наиболее характерна кластерная структура пространственного распределения, показанная мозаичной картой на рис. 4. Принимая во внимание реофильность индикаторных групп Ephemeroptera, Plecoptera и Trichoptera, отметим, что индекс EPT не учитывает повышенный естественный уровень минерализации некоторых обследованных рек аридной зоны, поэтому вариация его значений связана в первую очередь с гидрологическими особенностями рек разных географических широт, а не с антропогенным загрязнением.
Отображение распределения обилия различных групп или видов в сообществах макрозообентоса (рис. 5–6) не только позволяет сопоставить пространственное местоположение таксонов с их экологическими предпочтениями, но и создать предпосылки к геоинформационному подходу в оценке разнообразия на основе анализа многоканальных растровых изображений распределения групп гидробионтов.
За последние полвека было разработано большое число индексов видового разнообразия. Однако так и не было предложено единой методики, обобщающей это понятие и универсально применимой ко всем задачам и всем экологическим сообществам (Hurlbert, 1971; Magurran, 2004). Поскольку каждый из традиционных индексов связан с некоторым концептуально специфичным феноменом и подчеркивает только тот или иной компонент разнообразия (богатство, равномерность или таксономическое своеобразие), возникло понимание того, что для его измерения необходимо использовать составную статистическую меру (Stirling, Wilsey, 2001). Метод DER обеспечивает достаточно обоснованный подход к решению этой проблемы, а полученные с его помощью результаты хорошо интерпретируются визуально и позволяют расширить спектр содержательных выводов. В частности, специфицируется дополнительно понятие относительной редкости видов, связанное с возможностью оценки уровня эндемичности, видового состава локальных сообществ, специализацией местообитаний и т. д.
Апробация алгоритма DER на примере изученных донных сообществ позволила провести селекцию набора индексов, который приводил к наибольшей вариации объектов в пространстве полярных координат, т. е. обладал максимальной разрешающей способностью с точки зрения заданных критериев. В то же время сила статистической связи матрицы географических расстояний между участками (а) с матрицей дистанций на основе набора индексов разнообразия участков (б) оказалась существенно слабее, чем с матрицей таксономических расстояний по Брею – Кёртису (в): коэффициент корреляции Мантеля уменьшился с 0.284 до 0.175 для полного состава индесов и 0.201 для «информативной четверки». Вопрос о том, определяется ли этот эффект потерей информации при переходе от эмпирических численностей к расчетным индексам разнообразия или, наоборот, является результатом фильтрации «шума» в наблюдаемых данных, вероятно, послужит предметом дальнейших исследований.
Заключение
1. Видовая структура и биоразнообразие донных сообществ малых и средних рек бассейна Средней и Нижней Волги статистически значимо коррелируют с их пространственным расположением. Анализ показал высокую вероятность гипотезы о том, что градиент возрастания видового богатства ориентирован с юга на север, а максимум разнообразия приходится на лесостепную часть бассейна Саратовского водохранилища.
2. Использование методов обобщения большой совокупности индексов видового разнообразия дает возможность расширить рамки содержательного анализа и сделать дополнительные выводы.
Библиография
Алимов А. Ф. Элементы теории функционирования водных экосистем . СПб.: Наука, 2001. 147 с.
Зинченко Т. Д. Эколого-фаунистическая характеристика хирономид (Diptera, Chhironomidae) малых рек бассейна Cредней и Нижней Волги (Атлас) . Тольятти: Кассандра, 2011. 258 с.
Курина Е. М., Селезнев Д. Г. Анализ закономерностей организации комплексов видов макрозообентоса понто-каспийского и понто-азовского происхождения в водохранилищах Средней и Нижней Волги // Экология. 2019. № 1. С. 62–71.
Шитиков В. К. Интерполяция и визуализация пространственных данных. URL: https://stok1946.blogspot.com/2020/09/blog-post.html (дата обращения: 12.10.2020).
Шитиков В. К., Зинченко Т. Д. Использование чисел Хилла для оценки видового и таксономического разнообразия в группах местообитаний // Принципы экологии. 2013а. № 3. С. 23–36.
Шитиков В. К., Зинченко Т. Д. Изменение таксономического и функционального разнообразия сообществ макрозообентоса по продольному градиенту рек // Успехи современной биологии. 2013б. Т. 133, № 6. С. 566–577.
Шитиков В. К., Зинченко Т. Д. Статистический анализ структурной изменчивости донных сообществ и проверка гипотезы речного континуума // Водные ресурсы. 2014. Т. 41, № 5. С. 530–540.
Шитиков В. К., Розенберг Г. С., Зинченко Т. Д. Количественная гидроэкология: методы, критерии, решения : В 2 кн. М.: Наука, 2005. Кн. 1. 281 с. Кн. 2. 337 с.
Шитиков В. К., Розенберг Г. С., Костина Н. В. Методы синтетического картографирования территории (на примере эколого-информационной системы "REGION-VOLGABAS") // Количественные методы экологии и гидробиологии (Сборник научных трудов, посвященный памяти А. И. Баканова) / Отв. ред. чл.-корр. РАН Г. С. Розенберг. Тольятти: СамНЦ РАН, 2005. С. 167–227.
Bezmaternykh D. M., Vdovina O. N. Composition and structure of macrozoobenthos of lakes in different natural zones and subzones of Western Siberia // Limnology. 2020. Vol. 21. P. 3–13.
Chao A., Chiu C.-H., Jost L. Phylogenetic diversity measures based on Hill numbers // Philos. Trans. R. Soc. Lond. Ser. B – Biol. Sci. 2010. № 365. Р. 3599–3609.
Faith D. P. Conservation evaluation and phylogenetic diversity // Biol. Conserv. 1992. Vol. 61. P. 1–10.
Gaston K. J. Rarity. London: Chapman & Hall, UK, 1994. 205 p.
Golovatyuk L. V., Shitikov V. K., Zinchenko T. D. Estimation of the Zonal Distribution of Species of Bottom Communities in Lowland Rivers of the Middle and Lower Volga Basin // Biology Bulletin. 2018. Vol. 45 (10). Р. 1262–1268.
Golovatyuk L. V., Shitikov V. K. Salinity tolerance of macrozoobenthic taxa in small rivers of the Lake Elton basin // Russ. J. Ecol. 2016. Vol. 47. P. 540–545.
Guisande C., Heine J., García-Roselló E., González-Dacosta J., Vilas L., Perez-Schofield B. DER: An algorithm for comparing species diversity between assemblages // Ecological Indicators. 2017. Vol. 81. P. 41–46.
Hausdorf B., Hennig C. Null model tests of clustering of species, negative co-occurrence patterns and nestedness in meta-communities // Oikos. 2007. Vol. 116. P. 818–828.
Hennig C., Hausdorf B. Distance-based parametric bootstrap tests for clustering of species ranges // Comput. Stat. Data Anal. 2004. Vol. 45. P. 875–895.
Holyoak M., Leibold M., Holt R. (eds.). Metacommunities: spatial dynamics and ecological communities. Univ. of Chicago Press, 2005. 520 p.
Hurlbert S. H. The non-concept of species diversity: a critique and alternative parameters // Ecology. 1971. Vol. 52, № 4. P. 577–586.
Kembel S. W., Cowan P. D., Helmus M. R., Cornwell W. K., Morlon H., Ackerly D. D., Blomberg S. P., Webb C. O. Picante: R tools for integrating phylogenies and ecology // Bioinformatics. 2010. Vol. 26, № 11. Р. 1463–1464.
Koleff P., Gaston K. J. Latitudinal gradients in diversity: real patterns and random models // Ecography. 2001. Vol. 24. Р. 341–351.
Leibold M. A., Holyoak M., Mouquet N., Amarasekare P., Chase J. M., Hoopes M. F., Holt R. D., Shurin J. B., Law R., Tilman D., Loreau M., Gonzalez A. The metacommunity concept: a framework for multi-scale community ecology // Ecology Letters. 2004. Vol. 7. P. 601–613.
Leroy B., Petillon J., Gallon R., Canard A., Ysnel F. Improving occurrence-based rarity metrics in conservation studies by including multiple rarity cut-off points // Insect Conservation and Diversity. 2012. Vol. 5. Р. 159–168.
Macpherson E., Duarte C. M. Patterns in species richness, size, and latitudinal range of East Atlantic fishes // Ecography. 1994. Vol. 17. P. 242–248.
Magurran A. E. Measuring Biological Diversity. Oxford: Blackwell, 2004. 256 р. (Рус. изд.: Мэгарран Э. Экологическое разнообразие и его измерение. М.: Мир, 1992. 181 с.)
Manni F., Guerard E., Heyer E. Geographic patterns of (genetic, morphologic, linguistic) variation: how barriers can be detected by using Monmonier’s algorithm // Human Biol. 2004. Vol. 76, № 2. Р. 173–190.
Mantel N. The detection of disease clustering and a generalized regression approach // Cancer Res. 1967. Vol. 27. P. 209–220.
Oberdorff T., Guégan J.-F., Hugueny B. Global scale patterns of fish species richness in rivers // Ecography. 1995. Vol. 18. P. 345–352.
Ohio EPA. Addendum to biological criteria for the protection of aquatic life. Users manual for biological field assessment of Ohio surface waters // Ohio Environmental Protection Agency, Columbus, Ohio, 1989. Vol. 2.
Pielou E. C. Ecological Diversity. N.Y.: Wiley, 1975. 165 p.
Ricotta C. Through the jungle of biological diversity // Acta Biotheor. 2005. Vol. 53. P. 29–38.
Rex M. A., Stuart C. T., Hessler R. R., Allen J. A., Sanders H. L., Wilson G. D. F. Global-scale latitudinal patterns of species diversity in the deep-sea benthos // Nature. 1993. Vol. 365. P. 636–639.
Roy K., Jablonski D., Valentine J. W., Rosenberg G. Marine latitudinal diversity gradients: tests of causal hypotheses // Proceedings of the National Academy of Sciences. 1998. Vol. 95 (7). P. 3699–3702.
Stirling G., Wilsey B. Empirical relationships between species richness, evenness, and proportional diversity // Am. Natur. 2001. Vol. 158, № 3. P. 286–299.
Благодарности
Работа выполнена при поддержке гранта РФФИ № 15-04-03341 и в рамках государственного задания ФГБУН Институт экологии Волжского бассейна РАН, тема (проект) № AAAA-A17-117112040039-7 «Экологические закономерности структурно-функциональной организации, ресурсного потенциала и устойчивого функционирования экосистем Волжского бассейна» (направление 51 «Экология организмов и сообществ»).

 © 2011 - 2026
© 2011 - 2026