



Введение
Обладая высокой численностью и значительным видовым разнообразием, рептилии играют серьезную роль в аридных экосистемах и могут служить показателем происходящих в них сукцессионных процессов. Сукцессионные изменения сказываются прежде всего на узкоспециализированных видах, к которым здесь относятся псаммофилы. Среди рептилий астраханских полупустынь на первом месте в этом плане стоит ушастая круглоголовка (Phrynocephalus mystaceus mystaceus, Pallas, 1776), обитающая исключительно на открытых барханных и слабо закрепленных песках во всех частях своего ареала. В случае зарастания барханных песков ее численность значительно снижается, и популяция распадается на отдельные изолированные группировки. Экологические особенности этих группировок представляют особый интерес, поскольку дают возможность понять механизмы переживания видом значительных изменений характерного биотопа.
Судьба одной из таких группировок стала предметом нашего исследования в рамках изучения структуры полупустынных герпетокомплексов. Целью работы было изучение процесса и причин сокращения численности ушастой круглоголовки, которое шло вплоть до ее полного исчезновения на данной территории.
Материалы
Исследование проходило в нескольких километрах от поселка Досанг Красноярского района Астраханской области (N 46° 54'08.7264" E 47° 54'52.5312"). Время исследований – первая декада мая пяти последовательных полевых сезонов с 2010 по 2014 г. Далее исследования возобновили в начале мая 2017 г. На изученной территории поселения за время работы отловили, измерили и пометили всех встреченных ушастых круглоголовок, всего 57 особей.
Поселение ушастой круглоголовки, соответствующее уровню внутрипопуляционной группировки (Шилов, 1977), выделили для работы в результате картирования территории популяции, равной по площади примерно 7 км2. Рекогносцировочный этап исследования показал, что пригодных для псаммофилов участков немного. Ближайшее поселение вида обитало на расстоянии около 1 км. Кроме того, на расстоянии 2 км тянулась барханная гряда, также пригодная для ушастых круглоголовок. На всех отмеченных территориях вид имел низкую численность.
Исследованная группировка обитала на изолированном участке полузакрепленных песков площадью 0.4 га. Закрепленные пески, окружавшие площадь группировки, постепенно наступали на ее территорию.
Для оценки изменений, происходивших в биотопе, использовали геоботанические методы, главным образом показатель общего проективного покрытия растительности. За время исследований геоботанические описания провели три раза: в 2011, 2014 и 2017 гг. Всего заложили 90 геоботанических площадок, по 30 в каждый отмеченный сезон.
Методы
В процессе исследования использовали набор стандартных методов, подробно описанный нами в предыдущих публикациях (Полынова, Бажинова, 2012; Полынова и др., 2012).
У пойманных животных измеряли длину тела и хвоста с точностью до 1 мм и метили. Временную метку наносили с помощью спиртового маркера на спину ящерицы, а постоянную ставили с помощью отрезания кончиков фаланг пальцев по классической схеме (Mayhew, 1963; Tinkle, Woodward, 1967).
Для оценки привязанности животных к территории использовали метод картирования встреч, перемещений и жилых нор. Данные картирования переносили на электронную карту в программе MapInfo Professional 11.5. Уточнение и дополнение полученных материалов проводили с помощью двух дополнительных методов, тропления и осторожного преследования, а также визуальных наблюдений.
Метод осторожного преследования основан на выявлении знакомства и привязанности особи к индивидуальному участку. Известно, что оседлое животное уверенно передвигается по своему участку и пользуется имеющимися на нем укрытиями и норами. Когда наблюдатель осторожно, без резких движений на расстоянии 3–5 или 5–10 м (в зависимости от вида) следует за оседлыми особями, они бегают в пределах индивидуального участка, занимаясь рутинной деятельностью, прячутся в знакомые кусты и норы, а доходя до границы участка, сворачивают назад. У мигрирующих особей в случае осторожного преследования поведение резко отличается. Они сразу пугаются и убегают на далекое расстояние от места встречи с наблюдателем. Описанный метод осторожного преследования мы не раз проверяли на ящерицах многих видов. Он позволяет не только выявить привязанность животного к определенному участку, но также получить сведения о размерах и форме участка и этологический материал.
Животных, встреченных несколько раз в пределах небольшой территории, знакомых с ней и имеющих здесь постоянные норы, мы считали оседлыми. Остальных особей относили к мигрантам, которые приходили на участок группировки с уже упомянутых выше песчаных массивов. Основным путем перемещения мигрантов служили песчаные дороги.
Для оценки изменений, происходивших в биотопе, использовали описание геоботанических площадок по упрощенной схеме. Площадь поселения изначально разделили по степени зарастания на три типа: слабо закрепленные (площадь 0.12 га), полузакрепленные (площадь 0.22 га) и закрепленные песчаные участки (площадь 0.06 га). Такое описание биотопа широко встречается в зоологических исследованиях. Слабо закрепленные пески в основном располагались по гребням песчаных гряд и вокруг самого высокого песчаного бугра с кустом тамарикса (Támarix sp.) на вершине. Полузакрепленные участки шли по склонам песчаных гряд, а закрепленные – по межгрядовым понижениям. Ушастые круглоголовки использовали слабо закрепленные и полузакрепленные территории. В первый сезон границы этих биотопов нанесли на карту и в пределах каждого из них заложили по 10 расположенных равномерно геоботанических площадок площадью 1 × 1 = 1 м2. При описании площадок главным показателем служила степень проективного покрытия с точностью до 5 % (Воронов, 1973). Оценка достоверности различий полученных данных провели на основе непараметрического критерия Манна – Уитни.
Результаты
Особенностью исследования было то, что на наших глазах в течение пяти полевых сезонов происходило зарастание открытых песков и постепенное сокращение численности ушастой круглоголовки. Для измерения скорости зарастания исследуемой территории мы использовали показатель проективного покрытия. Процесс зарастания шел во всех трех типах биотопов: слабо закрепленных, полузакрепленных и закрепленных песках (табл. 1). В результате в 2017 г. слабо закрепленная территория превратилась в полузакрепленную, полузакрепленная – в закрепленную, а проективное покрытие закрепленного участка увеличилось в 1.6 раза, что в целом можно назвать процессом остепнения.
Таблица 1. Изменение проективного покрытия растительности исследуемой территории
| Название биотопа | Проективное покрытие (%) 2011 г. | Проективное покрытие (%) 2014 г. | Проективное покрытие (%) 2017 г. |
| Слабо закрепленные пески | 2.2 ± 1.83 | 5.5 ± 1.5 | 9.0 ± 2.0 |
| Полузакрепленные пески | 9.0 ± 2.0 | 23.0 ± 8.1 | 48.5 ± 10.3 |
| Закрепленные пески | 29.5 ± 6.5 | 43.0 ± 9.0 | 67.0 ± 20.0 |
Расчет достоверности различий по непараметрическому критерию Манна – Уитни показал, что отмеченные изменения проективного покрытия в целом достоверны (табл. 2).
Таблица 2. Достоверность различий проективного покрытия растительности исследуемой территории
| Вариант сравнения | Критерий Манна – Уитни U эмп. |
Уровень значимости |
| Слабо закрепленные пески, 2011–2014 гг. | 9 | p ≤ 0.01 |
| Слабо закрепленные пески, 2014–2017 гг. | 15 | p ≤ 0.01 |
| Полузакрепленные пески, 2011–2014 гг. | 14.5 | p ≤ 0.01 |
| Полузакрепленные пески, 2014–2017 гг. | 6 | p ≤ 0.01 |
| Закрепленные пески, 2011–2014 гг. |
14.5 | p ≤ 0.01 |
| Закрепленные пески, 2014–2017 гг. |
19.5 | 0.01 ≤ p ≤ 0.05 |
Изменение характерного биотопа негативно повлияло на численность группировки. Материалы, представленные на диаграмме (рис. 1), показывают, что за первые три года наблюдений общая численность группировки снизилась практически в два раза, в течение следующих двух лет она оставалась на нижнем уровне, а после двухлетнего перерыва наблюдений ушастую круглоголовку на данной территории мы больше не встретили.
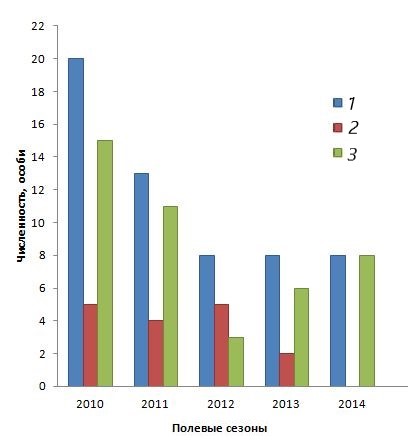
Рис. 1. Динамика численности группировки ушастой круглоголовки Phrynocephalus mystaceus mystaceus, май 2010–2014 и 2017 гг. 1 – общее число; 2 – оседлые особи; 3 – мигранты
Fig. 1. Population dynamics of the grouping of the lizard Phrynocephalus mystaceus mystaceus, May 2010–2014 and 2017. 1 – total number; 2 – sedentary individuals; 3 – migrating individuals
Во всех приводимых диаграммах использовался показатель абсолютной численности, а не плотности населения, поскольку материал касался только малочисленной группировки.
Снижение общей численности группировки шло, прежде всего, за счет уменьшения числа мигрантов (см. рис. 1). Численность оседлых ящериц оставалась первые три года примерно на одном уровне. На четвертый год оседлых ящериц стало в два раза меньше, а на пятый они полностью исчезли. Такой ход изменения численности говорит о том, что существование малочисленной группировки напрямую зависело от притока мигрирующих особей. Подобный вариант поддержания мозаичной пространственной структуры популяции хорошо известен у позвоночных животных, как в случае расселения, так и в случае падения численности (Шилов, 1977).
Анализ динамики половозрастной структуры мигрирующей части популяции позволил понять, за счет какой популяционной группы поддерживалось существование исследованной группировки (рис. 2).
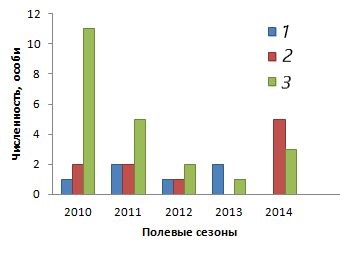
Рис. 2. Динамика половозрастной структуры мигрирующих особей группировки ушастой круглоголовки Phrynocephalus mystaceus mystaceus, май 2010–2014 и 2017 гг. 1 – самцы; 2 – самки; 3 – неполовозрелые особи
Fig. 2. Dynamics of sex and age structure of migrating individuals of the lizard Phrynocephalus mystaceus mystaceus grouping, May 2010–2014 and 2017. 1 – males; 2 – females; 3 – immature individuals
Основу потока мигрантов составляли неполовозрелые особи, и их численность резко снижалась год от года. Половозрелые самцы и самки составляли незначительную долю неоседлых особей и, по нашим наблюдениям, на участке группировки никогда повторно не встречались. Снижение потока мигрантов в целом и мигрирующего молодняка в частности зависело от отмеченного нами общего падения численности популяции на прилежащей территории.
Анализ половозрастной структуры оседлой части группировки позволил понять основу ее стабильности в изученный период, а также проследить процесс сокращения численности группировки и ее изчезновения (рис. 3).
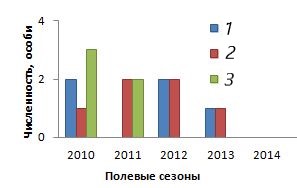
Рис. 3. Динамика половозрастной структуры оседлых особей группировки ушастой круглоголовки Phrynocephalus mystaceus mystaceus, май 2010–2014 и 2017 гг. 1 – самцы; 2 – самки; 3 – неполовозрелые особи
Fig. 3. Dynamics of sex and age structure of sedentary individuals of the lizard Phrynocephalus mystaceus mystaceus grouping, May 2010–2014 and 2017. 1 – males; 2 – females; 3 – immature individuals
В основе группировки стояли половозрелые животные обоих полов. Неполовозрелые ящерицы пропали на территории на третий год наблюдений. Мечение позволило обнаружить тот факт, что все оседлые самки держались на участке группировки минимум два года подряд, а одна особь встречалась даже три года. Напротив, состав оседлых самцов каждый год обновлялся. Из этого можно сделать вывод, что в основе стабильности изученной группировки стояли половозрелые самки.
Снижение общей численности поселения, таким образом, шло в основном за счет сокращения молодняка. Такое положение, несомненно, свидетельствовало о постепенном сокращении размножения на всей территории популяции. Несмотря на то что на пятый сезон в пределах площади группировки существовал еще значительный поток мигрантов, сама группировка практически исчезла. Основной причиной ее исчезновения послужило изменение характерного для вида биотопа – процесс зарастания всех типов песчаных участков и остепнение биоценоза, показанное нашими геоботаническими наблюдениями.
Обсуждение
Зарастание песчаных пустынь и полупустынь – масштабное явление на территории нашей страны (Лотиев, Батхиев, 2019; Сараев, Пестов, 2010; Табачишин и др., 2006 и др.). Оно представляет собой сукцессионный процесс. Обработка данных основных климатических факторов исследуемой территории показала, что зарастание песков имело положительную коррелятивную зависимость от увеличения общей суммы осадков за последнее десятилетие (Полынова, Мишустин, 2020). Ему также способствовало повсеместное сокращение поголовья скота и исчезновение диких копытных. Зарастание песков отрицательно повлияло не только на численность типичных псаммофилов, но, как показывают наши материалы (Полынова и др., 2019) и работы ряда авторов, и на численность других видов рептилий (Лотиев, Батхиев, 2019; Сараев, Пестов, 2010; Табачишин и др., 2006 и др.). Негативными последствиями зарастания для ящериц, очевидно, являются ухудшения условий передвижения и коммуникации.
Анализ полученных материалов позволил не только ответить на вопрос о причинах сокращения численности исследованной группировки ушастой круглоголовки, но и понять механизмы переживания видом значительных изменений характерного биотопа. В фундаменте этого механизма лежит поддержание группировки за счет притока мигрантов с близлежащей территории. Основу кочующих животных составляли неполовозрелые особи. Известно, что именно молодые и неполовозрелые особи большинства видов животных обладают повышенной подвижностью и склонностью к расселению (Наумов, 1956, 1963 и др.). Выполняя функцию расселения, молодые особи подпитывают изолированные группировки, что особенно важно в условиях низкой численности популяции. В группу мигрантов могут входить и взрослые животные. Наиболее очевидная причина этого – обострение социальных отношений во внутрипопуляционных группировках. В нашем случае доля половозрелых особей была невелика и причиной их миграций, скорее всего, был поиск пригодных местообитаний.
Поток мигрантов реализует еще одну очень важную функцию, являясь источником внутрипопуляционной информации, главным образом о размерах популяции. Иссякающий поток мигрантов служит показателем общего снижения численности. В этом случае для поиска внутривидовых контактов подвижность животных может значительно увеличиться. В нашем материале такой процесс наблюдался в 2014 г., когда все отмеченные ящерицы оказались мигрантами. В ситуации высокой численности значительный поток расселяющихся особей включает механизмы ее авторегуляции (Шилов, 1977).
Основу внутрипопуляционных группировок составляли половозрелые особи. Это закономерно, поскольку основная функция пространственной структуры с интенсивным типом использования территории – поддержание процесса размножения (Шилов, 1977). Именно поэтому в изученной нами группировке исчезновение оседлых половозрелых ящериц обоих полов прошло одновременно и стало концом обитания вида на данной территории. Особый интерес в наших материалах представляет тот факт, что стабильность группы в течение всего периода наблюдений обеспечивала привязанность к участку самок, а не самцов. Можно предположить, что подобная оседлость самок обусловлена тем, что в пределах территории группировки находились наиболее пригодные места для откладки яиц.
Полученные результаты являются примером общих закономерностей, известных для наземных позвоночных, но недостаточно изученных у пресмыкающихся и ранее не описанных для ушастой круглоголовки. Возможность их экстраполяции требует дальнейших исследований.
Заключение
- Пятилетние наблюдения за внутрипопуляционной группировкой ушастой круглоголовки (Phrynocephalus mystaceus mystaceus Pallas, 1776) показали сокращение ее численности вплоть до полного исчезновения на данной территории.
- Основной причиной деградации группировки явилось зарастание слабо закрепленных и полузакрепленных песчаных участков характерного для вида биотопа.
- Поддержание существования внутрипопуляционной группировки в течение ряда лет шло за счет потока мигрантов, проходивших через исследованную территорию.
- Поток мигрантов в основном состоял из неполовозрелых особей, возрастной группы, обладающей повышенной подвижностью и обычно служащей целям расселения.
- Ядро внутрипопуляционной группировки составляли оседлые половозрелые особи, причем стабильность группировки определялась привязанностью к территории самок.
Библиография
Воронов А. Г. Геоботаника . М.: Высшая школа, 1973. 384 с.
Лотиев К. Ю., Батхиев А. М. О деградации туранского герпетофаунистического комплекса в Терском песчаном массиве (Восточное Предкавказье) // Известия высших учебных заведений. Поволжский регион. Естественные науки. 2019. №2. С. 115–128. DOI: 10.21685/2307-9150-2019-2-12
Наумов Н. П. Межвидовые и внутривидовые отношения у животных (преимущественно позвоночных) // Зоологический журнал. 1956. Т. 41. Вып. 1. С. 74–89.
Наумов Н. П. Экология животных . М.: Высшая школа, 1963. 618 с.
Полынова Г. В., Бажинова А. В. Учет особенностей активности при оценке численности популяции круглоголовки-вертихвостки (Phrynocephalus guttatus guttatus) // Зоологический журнал. 2012. Т. 91. № 11. С. 1411–1414.
Полынова Г. В., Бажинова А. В., Гриб Е. В. Материалы по пространственной структуре популяции круглоголовки-вертихвостки (Phrynocephalus guttatus guttatus) в полупустынях Астраханской области // Наземные позвоночные аридных экосистем: Материалы международной конференции, посвященной памяти Н. А. Зарудного. Ташкент: CHINOR ENK, 2012. С. 260–264.
Полынова Г. В., Мишустин С. С. Изменение пространственной структуры популяции разноцветной ящурки Eremias arguta deserti (Gmelin, 1789) в полупустынях Астраханской области // Принципы экологии. 2020. № 2. С. 87–96. DOI: 10.15393/j1.art.2020.10303
Полынова Г. В., Мишустин С. С., Полынова О. Е. Динамика герпетокомплекса песчаных пустынь Астраханской области // Известия высших учебных заведений. Поволжский регион. Естественные науки. 2019. № 2. С. 150–163. DOI: 10.21685/2307-9150-2019-2-15
Сараев Ф. А., Пестов М. В. К кадастру рептилий Северного и Северо-Восточного Прикаспия // Герпетологические исследования в Казахстане и сопредельных странах: Сборник научных статей / Под ред. Т. Н. Дуйсебаевой. Алматы: АСБК – СОПК, 2010. С. 172–191.
Табачишин В. Г., Завьялов Е. В., Табачишина Е. И. Пространственное размещение разноцветной ящурки – Eremias arguta (Pallas, 1776) на севере ареала в Поволжье // Современная герпетология. 2006. Т. 5/6. С. 117–124.
Шилов И. А. Эколого-физиологические основы популяционных отношений у животных . М.: Изд-во Московского университета, 1977. 263 с.
Mayhew W. W. Biology of the granite spring lizard, Sceloporus orcutti // Amer. Midl. Nat. 1963. Vol. 69, № 2. P. 310–327. DOI: 10.2307/2422913
Tinkle D. W., Woodward D. W. Relative movements of lizards in natural populations as determined from receptive radii // Ecology. 1967. Vol. 48, № 1. P. 166–168. DOI: 10.2307/1933431
Благодарности
Публикация выполнена при поддержке Программы стратегического академического лидерства РУДН. Авторы благодарят студентов экологического факультета, принимавших участие в сборе полевого материала.

 © 2011 - 2026
© 2011 - 2026