



Введение
Кровососущие комары (Diptera: Culicidae) имеют важное практическое значение как переносчики и промежуточные хозяева возбудителей заболеваний человека и животных. Представитель данного семейства, Aedes dorsalis (Meigen, 1830) переносит калифорнийский энцефалит, вирус лихорадки долины Рифта (RVFV) (Turell et al., 2010). По данным Национального центра биологической информации (NCBI), с данным видом связаны еще 5 вирусов: Лловиу вирус (Liao ning (LNV)), вирус восточного лошадиного энцефаломиелита, Тахина вирус (Tahyna virus (TAHV)), Банна вирус (Banna virus (BAV)) и Уидби вирус (Whidbey virus) (Cao et al., 2011; Lu et al., 2011; Greninge et. al., 2016; Hughes et. al., 2016; Liu et. al., 2016).
Ранее A. dorsalis считался подвидом A. caspius Pallas, 1771 – палеарктического переносчика туляремии и вируса Тягиня (Гуцевич и др., 1970). Комара A. caspius считали политипичным по морфологическим признакам, а A. dorsalis рассматривали в качестве его подвида (Гуцевич, 1977). Научная дискуссия о политипичности вида A. caspius была основана на его морфологии и зоогеографии, без использования генетических данных. Самостоятельность вида A. dorsalis была подтверждена генетическими характеристиками. В каталоге комаров мира A. caspius и A. dorsalis были приведены как самостоятельные виды (Minař, 1990). Исследователи особенностей генетики A. dorsalis отмечали, что приморские и внутриконтинентальные популяции данного вида не имели генетических различий, т. к. характер генетической дифференциации не соответствовал географическому расположению изученных популяций в Северной Америке (Gimnig, Eldridge, 1999). Генетическая самостоятельность A. dorsalis не вызывала сомнений, а разобщенность внутриматериковых и прибрежных популяций объяснялась топическими особенностями. На территории Евразии сравнительных исследований генетики внутриматериковых и приморских популяций A. dorsalis не проводили.
До настоящего времени мало изучены экология и распространение комара A. dorsalis на территории Евразии, в особенности его внутриконтинентальные популяции. Находки данного вида известны на побережье Белого моря в Мурманской области и Республике Карелия. Были выполнены экологические исследования прибрежных популяций A. dorsalis (Тамарина, Георгиева, 1981; Георгиева, 2004). В материковой части Евразии внутриконтинентальные популяции A. dorsalis были обнаружены в местах выхода подземных рассолов, где исторически велась добыча поваренной соли и соленые воды использовались в лечебных целях. Опубликованы краткие сведения о находках внутриконтинентальных популяций A. dorsalis в засоленных водоемах города-курорта Старая Русса Новгородской области (Кункова, 2003), а также в окрестностях бальнеологического курорта «Серегово» в Республике Коми (Панюкова, 2007).
Целью данной работы стал обзор особенностей распространения и экологии комара Aedes dorsalis на территории России и стран ближнего зарубежья. В задачи исследования входили рассмотрение особенностей генетики и морфологии, а также реконструкция ареала данного вида.
Материалы
Экологические и морфологические исследования комара A. dorsalis выполнены нами в разные годы в Республике Коми (с. Серегово 62°19' с. ш. 50°41' в. д.), Новгородской области (г. Старая Русса 57°59' с. ш. 31°21' в. д.), Краснодарском крае (г. Ейск 46°42' с. ш. 38°16' в. д.) и Украине (окрестности г. Одессы 46°33' с. ш., 30°46' в. д.). Сборы A. dorsalis проведены в Старой Руссе в 1999, 2002–2003 гг., в Серегово в 2006 г., в Одессе в 2018 и 2019 гг., в Ейске в 2019 г. Использованы все доступные материалы коллекций наколотых экземпляров имаго самцов и самок A. dorsalis Зоологического института (Санкт-Петербург), Научного музея Института биологии (г. Сыктывкар) и коллекция Украинского научно-исследовательского противочумного института имени И. И. Мечникова. Для данного исследования из серии сборов были случайным образом отобраны один или несколько экземпляров (таблица).
Сборы Aedes dorsalis (Meigen, 1830) на территории России и стран ближнего зарубежья
| № | Название места сбора, как на этикетке (современное название) | Координаты | Дата | Пол | Автор сбора | Место хранения материала |
| 1 | Старый Петергоф (Петергоф) | 59°52′ с. ш., 29°53′ в. д. | 7.07.1924 | ♀ | Мончадский | ЗИН РАН |
| 1 | г. Ленинград, Сосновая поляна (г. Санкт-Петербург) |
59°35′ с. ш., 30°20′ в. д. | 29.07.1970 | ♀ | Пастухов | ЗИН РАН |
| 2 | г. Старая Русса | 57°59′ с. ш., 31°21′ в. д. | 16–27.07.1924 | ♀ | Мончадский | ЗИН РАН |
| 2 | г. Старая Русса | 57°59′ с. ш., 31°21′ в. д. | 28.05.2002 | ♀ | Панюкова | ИБ УрО РАН |
| 2 | г. Старая Русса | 57°59′ с. ш., 31°21′ в. д. | 29.07.2002 | ♀ | Панюкова | ИБ УрО РАН |
| 3 | г. Киев | 50°27′ с. ш., 30°31′ в. д. | 5.12.1928 | ♀ | Рыбинский | ЗИН РАН |
| 4 | Куяльницкий лиман, левый берег, Одесская обл., Лиманский р-н | 46°40′ с. ш., 30°43′ в. д. | 06.08.2018 | ♂ | Ахраменко | УНИПИ |
| 4 | г. Одесса, микрорайон Лузановка, побережье Черного моря | 46°33′ с. ш., 30°46′ в. д.
|
04.06.2019 | ♀ | Ахраменко | УНИПИ |
| 5 | Карудиол лиман, верховья, Одесская обл., Татарбунарский р-н | 45°49′ с. ш., 30°04′ в. д. | 30.05.2018 | ♀ | Ахраменко | УНИПИ |
| 5 | оз. Сасык, Одесская обл., Татарбунарский р-н | 45°25′ с. ш., 29°24′ в. д. | 29.05.2018 | ♀ | Ахраменко | УНИПИ |
| 6 | г. Екатеринослав (г. Днепр) |
48°28′ с. ш., 35°01′ в. д. | 1926 | ♀ | Гуцевич | ЗИН РАН |
| 7 | г. Чугуев, «бассейные» | 49°50′ с. ш., 36°40′ в. д. | 15.05.1928 | ♀ | ? | ЗИН РАН |
| 8 | д. Семеновка, р. Кальмиус (г. Донецк) | 48°00′ с. ш., 37°48′ в. д. | 1928 | ♀ | ? | ЗИН РАН |
| 9 | г. Ейск | 46°42′ с. ш., 38°16′ в. д. | 28.08.2019 | ♀ | Панюкова | ИБ УрО РАН |
| 10 | г. Ессентуки, копани садов | 44°02′ с. ш., 42°51′ в. д. | 1927 | ♀ | Месс | ЗИН РАН |
| 11 | г. Пятигорск | 44°03′ с. ш., 43°03′ в. д. | 22.08.1927 | ♀ | Месс | ЗИН РАН |
| 11 | г. Пятигорск | 44°03′ с. ш., 43°03′ в. д. | 22.08.1927 | ♂ | Месс | ЗИН РАН |
| 12 | г. Воронеж, Шиловский лес | 51°29′ с. ш., 39°10′ в. д. | 1973 | ♀ | Камолов | ЗИН РАН |
| 13 | г. Саратов | 51°32′ с. ш., 46°00′ в. д. | 1917 | ♀ | Кузнецов | ЗИН РАН |
| 14 | Жигулевский гос. заповедник, Самарская обл. | 53°24′ с. ш., 49°49′ в. д. | 18.07.2004 | ♀ | Халин | ЗИН РАН |
| 15 | с. Серегово, Коми | 62°19′ с. ш., 50°41′ в. д. | 13.07.2006 | ♀ | Панюкова | ИБ УрО РАН |
| 16 | г. Свердловск (г. Екатеринбург) |
56°51′ с. ш., 60°36′ в. д. | ? | ♀ | Попов | ЗИН РАН |
| 17 | г. Старая Бухара (г. Бухара) |
39°46′ с. ш., 64°25′ в. д. | 20.04.1925 | ♀ | Мончадский | ЗИН РАН |
| 18 | г. Омск | 54°58′ с. ш., 73°23′ в. д. | 1926 | ♀ | Мальцев | ЗИН РАН |
| 19 | р. Барынкол, с. Нарынкол |
42°43′ с. ш., 80°10′ в. д. | 17.08.1962 | ♀ | ? | ЗИН РАН |
| 19 | р. Барынкол, с. Нарынкол |
42°43′ с. ш., 80°10′ в. д. | 17.08.1962 | ♀ | ? | ЗИН РАН |
| 19 | р. Барынкол, с. Нарынкол |
42°43′ с. ш., 80°10′ в. д. | 17.08.1962 | ♀ | ? | ЗИН РАН |
| 20 | гора Кош-Агач, 1750 м, Алтай |
49°59′ с. ш., 88°40′ в. д. | 4.08.1964 | ♂ | Грунин | ЗИН РАН |
| 20 | гора Кош-Агач, 1750 м, Алтай |
49°59′ с. ш., 88°40′ в. д. | 4.08.1964 | ♂ | Грунин | ЗИН РАН |
| 21 | с. Ташанта, Чуйская степь | 49°42′ с. ш., 89°11′ в. д. | 3.07.1964 | ♂ | Грунин | ЗИН РАН |
| 21 | с. Ташанта, Чуйская степь | 49°42′ с. ш., 89°11′ в. д. | 3.07.1964 | ♂ | Грунин | ЗИН РАН |
| 21 | с. Ташанта 10 км к юго-западу, Чуйская степь | 49°42′ с. ш., 89°11′ в. д. | 5.07.1964 | ♀ | Грунин | ЗИН РАН |
| 22 | г. Минусинск, сады | 53°18′ с. ш., 90°20′ в. д. | 23.07.1926 | ♂ | Скотобой, Кравченко | ЗИН РАН |
| 22 | г. Минусинск, сады | 53°18′ с. ш., 90°20′ в. д. | 23.07.1926 | ♂ | Скотобой, Кравченко | ЗИН РАН |
| 22 | г. Минусинск | 53°18′ с. ш., 90°20′ в. д. | 1926 | ♀ | Внуковский | ЗИН РАН |
| 23 | Ачинский округ | 56°38′ с. ш., 90°25′ в. д. | 16.08.1926 | ♀ | Внуковский | ЗИН РАН |
| 24 | Красноярск, о. Татышев |
56°01′ с. ш., 92°56′ в. д. | 1925 | ♀ | Троицкий | ЗИН РАН |
| 24 | г. Красноярск, о. Татышев, малярийная станция |
56°01′ с. ш., 92°56′ в. д. | 18.08.1926 | ♀ | ? | ЗИН РАН |
| 25 | г. Усть-Кут на р. Лене, Иркутская обл. | 56°48′ с. ш., 105°50′ в. д. | 7.06.1957 | ♀ | Городков | ЗИН РАН |
| 26 | г. Иркутск | 52°17′ с. ш., 104°18′ в. д. | ? | ♀ | ? | ЗИН РАН |
| 26 | д. Мельниково, Кайск. горн. обл. Иркутск | 52°17′ с. ш., 104°18′ в. д. | 1912 | ♀ | Максимович | ЗИН РАН |
| 27 | ст. Онохой, Забайкалье | 51°55′ с. ш., 108°10′ в. д. | ? | ♀ | ? | ЗИН РАН |
| 27 | ст. Онохой, Забайкалье | 51°55′ с. ш., 108°10′ в. д. | ? | ♀ | ? | ЗИН РАН |
| 28 | пос. Муя, Читинская обл. | 52°02′ с. ш., 113°30′ в. д. | ? | ♀ | ? | ЗИН РАН |
| 29 | ур. Куланжа, Читинская обл., Борз. р-н | 50°24′ с. ш., 116° 32′ в. д. | 9.06.1959 | ♀ | Грунин | ЗИН РАН |
| 30 | г. Якутск | 62°02′ с. ш., 129°43′ в. д. | 28.07.1927 | ♀ | Москвин | ЗИН РАН |
| 31 | Уссурийский край | 43°48′ с. ш., 131°57′ в. д. | 1927 | ♀ | Штакельберг | ЗИН РАН |
| 32 | Заповедник Кедровая Падь, Приморская обл. | 43°09′ с. ш., 131°30′ в. д. | 28.07.1940 | ♀ | Мончадский | ЗИН РАН |
| 33 | Судзухинский заповедник, Приморский край (Лазовский заповедник) | 43°14′ с. ш., 133°24′ в. д. | 20.07.1977 | ♀ | ? | ЗИН РАН |
| 34 | г. Владивосток, окрестности | 43°07′ с. ш., 131°54′ в. д. | 16.07.1903 | ♀ | Гавронский | ЗИН РАН |
Примечание. Номера (№№) соответствуют номеру на карте (рис. 1); ? – данные отсутствуют. Сокращения в графе «Место хранения материала»: ЗИН РАН – Зоологический институт Российской академии наук (г. Санкт-Петербург); ИБ УрО РАН – Институт биологии Коми научного центра Уральского отделения РАН (г. Сыктывкар); УНИПИ – Украинский научно-исследовательский противочумный институт им. И. И. Мечникова (г. Одесса).
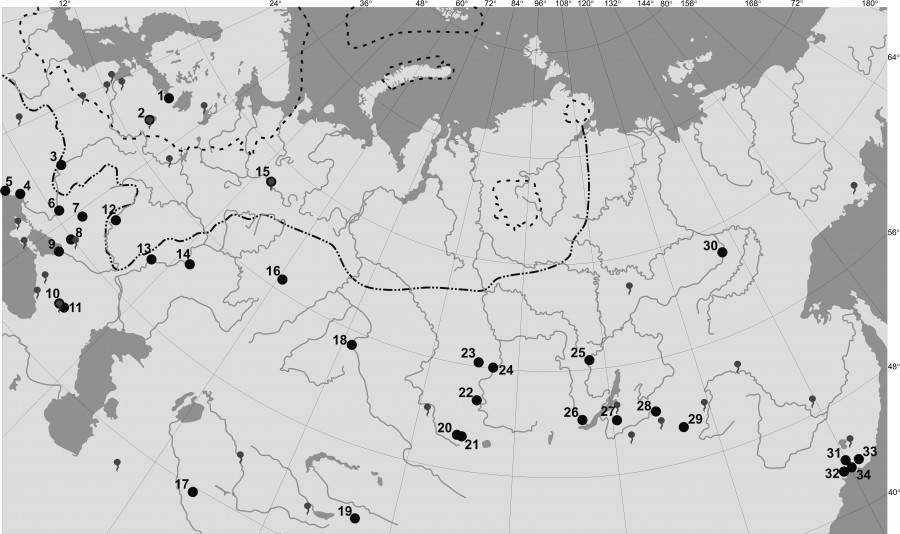
Рис. 1. Карта-схема точек сбора A. dorsalis на территории России и стран ближнего зарубежья.
Пунктиром на карте указаны границы Валдайского оледенения; пунктиром с двумя точками – границы Днепровского оледенения (по: Герасимов, Марков, 1939; Оледенение…, 1984; Svendsen et al., 1999; Смульский, Иванова, 2018). Топографический знак «источник» обозначает минеральные источники (по: Полезные ископаемые…, 1984). Точки соответствуют географическим объектам: 1 – г. Санкт-Петербург (Петергоф), 2 – г. Старая Русса, 3 – г. Киев, 4 – г. Одесса, 5 – оз. Сасык, 6 – г. Днепр, 7 – г. Чугуев, 8 – г. Донецк, 9 – г. Ейск, 10 – г. Ессентуки, 11 – г. Пятигорск, 12 – г. Воронеж, 13 – г. Саратов, 14 – Жигулевский заповедник, 15 – с. Серегово, 16 – г. Екатеринбург, 17 – г. Бухара, 18 – г. Омск, 19 – с. Нарынкол, 20 – гора Кош-Агач, 21 – с. Ташанта, 22 – г. Минусинск, 23 – Ачинский округ, 24 – г. Красноярск, 25 – г. Усть-Кут, 26 – г. Иркутск, 27 – ст. Онохой, 28 – пос. Муя, 29 – ур. Куланжа, 30 – г. Якутск, 31 – Уссурийский край, 32 – заповедник «Кедровая Падь», 33 – Лазовский заповедник, 34 – г. Владивосток
Fig. 1. The map of the collection points of the A. dorsalis on the territory of Russia and neighboring countries.
On the map the dashed lines indicate the borders of the Valdai glaciation; the dotted line with two points is the border of the Dnieper glaciation (after: Gerasimov, Markov, 1939; Glaciations..., 1984; Svendsen et al., 1999; Smulsky, Ivanova, 2018). The topographic sign "source" refers to mineral springs (according to: Minerals..., 1984). Points correspond to geographical objects: 1 – St. Petersburg (Peterhof), 2 – Staraya Russa, 3 – Kiev, 4 – Odessa, 5 – Lake Sasyk, 6 – the city of Dnieper, 7 – Chuguev, 8 – Donetsk, 9 – Yeysk, 10 – Essentuki, 11 – Pyatigorsk, 12 – Voronezh, 13 – Saratov, 14 – Zhigulevsky Reserve, 15 – village Seregovo, 16 – Yekaterinburg, 17 – Bukhara, 18 – Omsk, 19 – v. Narynkol, 20 – Mount Kosh-Agach, 21 – v. Tashanta, 22 – the city of Minusinsk, 23 – Achinsky district, 24 – Krasnoyarsk, 25 – Ust-Kut, 26 – Irkutsk, 27 – st. Onokhoy, 28 – v. Muya, 29 – tract Kulanzha, 30 – Yakutsk, 31 – Ussuri Territory, 32 – Kedrovaya Pad Reserve, 33 – Lazovsky Reserve, 34 – Vladivostok
Методы
Внутриконтинентальные популяции A. dorsalis изучали в Новгородской области и Республике Коми. Собраны имаго и личинки данного вида, личинки встречены в слабосоленых водоемах. Приморские популяции данного вида комара исследованы в городах Одесса и Ейск (побережье Черного и Азовского морей соответственно). В г. Ейске собраны только имаго комаров, развитие личинок, возможно, происходит в Ейском лимане, на берегу которого проведены учеты. Отмечены активные нападения в сумерках видов A. dorsalis и A. caspius в соотношении 1 к 3. В окрестностях г. Одессы также отмечены оба вида, собраны их личинки и имаго. Сбор личинок выполняли сачком и фотокюветой. Из живых собранных личинок и куколок выплаживали имаго самцов и самок. Сбор имаго самок осуществляли на себе пробиркой-морилкой, заполненной 70 % спиртом, в течение 10–20 минут. Монтирование имаго на энтомологические иголки осуществляли в условиях лаборатории. Наколотые экземпляры имаго определяли по ключам (Гуцевич и др., 1970; Becker et al., 2010). Для проведения молекулярно-филогенетического анализа A. dorsalis и A. caspius использовали последовательности первой субъединицы гена цитохром оксидазы (COI) митохондриальный ДНК, взятые из баз генетических данных Bold Systems и NCBI на 08.04.2020. Нуклеотидные последовательности выравнивали с использованием алгоритма ClustalW и редактировали в программном пакете Mega 7.0 (Kumar et al., 2016). Филогенетические деревья строили с вычислением поддержки начальной загрузки для узлов ветвления (1000 повторений) с использованием метода максимального правдоподобия (ML) в программном пакете Mega 7.0. В результате нами построена дендрограмма, которая показывает видовые различия A. caspius и A. dorsalis на примере разных территорий. Для A. dorsalis проанализировано 27 последовательностей гена COI из восьми стран: Канада, Англия, США, Россия, Япония, Китай, Южная Корея и Швеция. Для A. сaspius проанализировано 7 последовательностей гена COI из пяти стран: Испания, Италия, Бельгия, Иран и Пакистан.
Результаты
Генетика A. dorsalis
На молекулярно-филогенетическом дереве, построенном на основании сравнения последовательностей гена COI, взятых в анализ видов A. dorsalis и A. сaspius разделились на две клады. В первую кладу вошли образцы A. dorsalis из Северной Америки и Евразии, во вторую кладу – образцы A. сaspius. На полученной дендрограмме образцы A. dorsalis и A. сaspius образовали отдельные клады с бутстреп-поддержкой (коэффициент бутстрепа = 87). При этом в кладе A. dorsalis с невысокой поддержкой обособились две подгруппы по принадлежности к материкам (рис. 2).
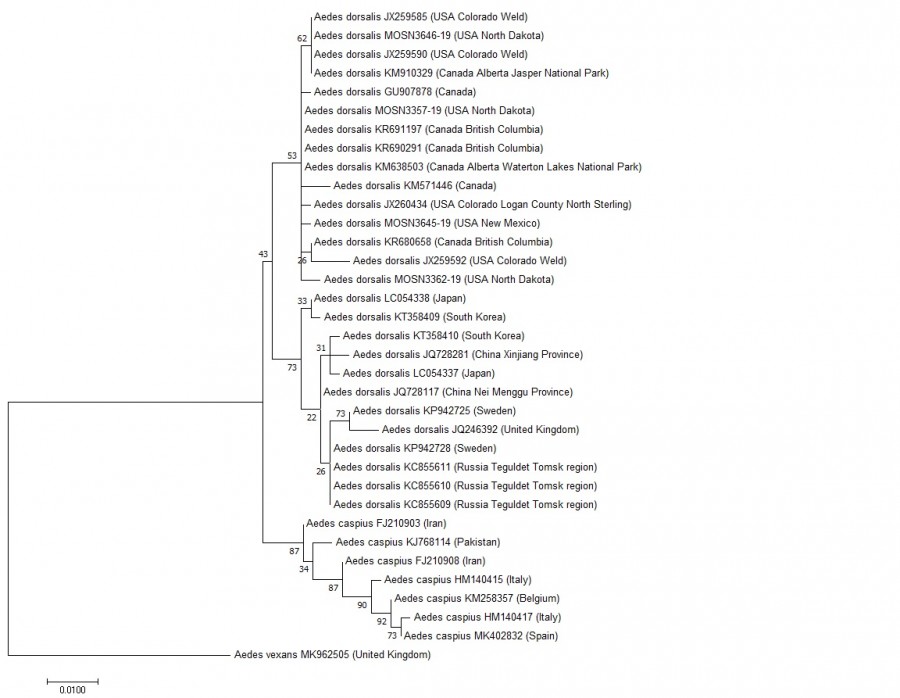 Рис. 2. Филогенетическое дерево максимального правдоподобия для представителей A. dorsalis и A. caspius, построенное по участку гена COI на основе данных GenBank
Рис. 2. Филогенетическое дерево максимального правдоподобия для представителей A. dorsalis и A. caspius, построенное по участку гена COI на основе данных GenBank
Fig. 2. Phylogenetic tree of maximum likelihood for the representatives of A. dorsalis and A. caspius, built on the region of the COI gene based on GenBank data
Для сравнения укажем, что в работе Н. В. Храбровой с соавторами (2013) данные виды также разнесены в две соседние клады с относительно высокой бутстреп-поддержкой (коэффициент бутстрепа = 90). Других исследований, позволяющих оценить генетический полиморфизм между двумя сравниваемыми видами на основании анализа последовательности COI митохондриальной ДНК, в доступной нам литературе не обнаружено. В целом проведенный нами молекулярно-филогенетический анализ, выполненный на основании сравнения последовательностей гена COI, разнес виды A. dorsalis и A. caspius в разные клады, что может являться дополнительным признаком, указывающим на генетическую разобщенность этих видов, и вносит дополнительные сведения в дискуссию о полиморфизме A. caspius, предложенную А. В. Гуцевичем (1977). Тот факт, что представители вида A. dorsalis, хотя с невысокой бутстреп-поддержкой, поделились по принадлежности к материкам, может указывать на то, что изоляция группировок на разных материках привела к их генетической дифференциации после разъединения в плейстоцене Берингийского сухопутного перешейка, Берингии (Кожевников, Железнов-Чукотский, 1995), соединяющего материки в мезозое и начале кайнозоя. Экологически и генетически A. dorsalis самостоятельный вид, распространенный на разных материках, но близкий к A. caspius морфологически.
Морфология A. dorsalis
Известно, что имаго A. dorsalis хорошо определяются по белым чешуйкам на лапках, образующим светлые кольца, при этом каждое кольцо опоясывает два членика лапки: вершину одного и основание последующего. Брюшко сверху с продольной полосой светлых чешуек (Гуцевич и др., 1970). Жилки крыла (в особенности костальная жилка) покрыты светлыми и темными чешуйками. У A. dorsalis светлые чешуйки на костальной жилке сформированы в группы, у A. caspius светлые чешуйки перемешаны с темными. На переднеспинке у A. dorsalis светлые чешуйки образуют полукруг, а вдоль среднеспинки тянется полоса темных чешуек (Becker et al., 2010). По данным литературы, A. caspius хорошо отличается от близких видов по окраске спинки и крыла (рис. 3).
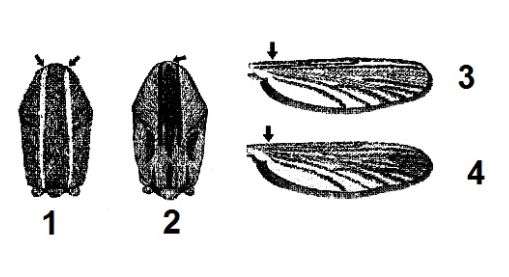
Рис. 3. Морфологические отличия видов по окраске спинки (1, 2) и крыла (3, 4): 1, 3 – A. caspius; 2, 4 – A. dorsalis (Becker et al., 2010)
Fig. 3. Morphological differences of species in color of the back (1, 2) and wing (3, 4): 1, 3 – A. caspius; 2, 4 – A. dorsalis (Becker et al., 2010)
Однако признак окраски крыла нельзя назвать надежным. Просмотренный нами коллекционный материал показал, что белые чешуйки в основании крыла есть у обоих видов и их расположение группами и единичное также свойственно обоим видам. При этом чешуйки на крыльях могут часто осыпаться и повреждаться при сборе и транспортировке. Окраска спинки – более надежный признак, при этом цветовая гамма чешуек очень изменчива у экземпляров из разных географических пунктов. Еще более надежным признаком может служить место расположения чешуек спинки. Имеющиеся коллекционные экземпляры (из фондовой коллекции Зоологического института РАН), определенные как A. caspius и A. dorsalis, мы сравнили по 7 морфологическим признакам, часто используемым для определения видов комаров: расположению белых чешуек на хоботке, щупиках, спинке, костальной жилке крыла, брюшке сверху, а также по окраске последнего членика задней лапки и форме коготка задней лапки. Статистически достоверных различий по данным признакам обнаружено не было. Была отмечена географическая изменчивость в окраске брюшка. У южных экземпляров имаго комаров A. dorsalis из городов Ейск, Одесса и Ессентуки имелись сплошные светлые чешуйки на тергитах брюшка, только по бокам располагались темные чешуйки в виде неясных пятен. Экземпляры комаров из северных популяций с. Серегово Республики Коми и г. Старая Русса Новгородской области имели незначительное количество светлых чешуек на брюшке, которые образовывали тонкую светлую продольную линию (рис. 4). В целом морфологические различия по имаго позволяют диагностировать вид A. dorsalis. Диагностические различия проявляются у самцов и самок.

Рис. 4. Географическая изменчивость в окраске брюшка у A. dorsalis: 1 – южный вариант (г. Одесса, самка); 2 – северный вариант (г. Старая Русса, самец)
Fig. 4. Geographical variability in the color of the abdomen in A. dorsalis: 1 – southern option (Odessa, female); 2 – Northern option (Staraya Russa, male)
Экология A. dorsalis
Развитие комара A. dorsalis происходит с полным превращением и включает следующие стадии: зимующее яйцо, личинку, куколку и имаго. Яйца самки откладывают на почву возле водоемов, в которых развивались личинки данного вида, что свойственно для представителей подрода Ochlerotatus рода Aedes. Диапауза на стадии яйца также характерна для представителей данного рода. Личинка A. dorsalis может встречаться в пресных, слабосоленых и соленых водоемах, вид является факультативным галофилом. В Новгородской области личинки A. dorsalis встречались в пресных водоемах совместно с личинками видов комаров, отмечающихся только в пресных водоемах, таких как Aedes cantans, A. communis, A. cyprius, A. excrucians и A. leucomelas. При этом в пресных водоемах отмечены единичные находки A. dorsalis. В слабосоленых и соленых водоемах вид встречался массово при отсутствии межвидовой конкуренции. Куколка данного вида не питается, ее развитие происходит за счет питательных веществ, накопленных на личиночной стадии, как у всех куколок Culicidae. Продолжительность стадии куколки составляет несколько дней, в зависимости от температуры и уровня воды в водоеме. Выплаживаются из куколок первыми самцы, через день или два происходит вылет из водоема имаго самок. Имаго A. dorsalis встречались в сборах с мая по сентябрь в пойменных (луговых и кустарниковых) и лесных биотопах совместно с комарами Aedes cinereus, A. diantaeus, A. excrucians, A. leucomelas, A. cantans, A. vexans и Anopheles messeae. Для северных внутриконтинентальных и приморских популяций отмечены два поколения данного вида. На юге – более двух поколений. Первое поколение развивается в апреле – мае из перезимовавших яиц, оно малочисленное. Развитие первого поколения проходит в конкурентных условиях с другими весенними видами комаров, в особенности в пресных водоемах. Второе поколение развивается в июле, из яиц, отложенных самками первого поколения. Второе поколение, как у большинства полицикличных видов, более многочисленное. На севере наблюдали развитие второго поколения в июле и августе, при активном нападении имаго в водоемах обнаруживали личинок младших возрастов. На юге возможны три поколения, а имаго встречаются до декабря. Так, по коллекционным материалам известно, что доктор Рыбинский собирал в г. Киеве самок A. dorsalis 5.12.1928 (см. таблицу). При средней продолжительности жизни данного комара в 60 дней данный факт говорит о третьем или последующем поколении данного вида на юге ареала, с развитием яиц в сентябре – октябре.
По литературным данным известно, что в природных популяциях развитие личинок A. dorsalis происходит в широком диапазоне солености, жесткости, сухого осадка, щелочности и других гидрохимических показателей (Георгиева, 2004), что также подтверждает высокую степень экологической пластичности вида. Экологические исследования северной популяции A. dorsalis проведены на побережье Белого моря между 66° и 67° с. ш. (Тамарина, Георгиева, 1981). В результате было отмечено, что A. dorsalis дает два поколения за северное лето благодаря колебаниям уровня моря – морских сизигиев, которые наполняют прибрежные водоемы каждые 28 дней. Считается, что проникновение лесостепного комара A. dorsalis в приполярные районы Белого моря связано с интразональными элементами ландшафта: прибрежными литоральными и пойменными водоемами, имеющими особенности гидрологического режима. Такие водоемы заполняются водой как в период весеннего таяния снега и выпадения атмосферных осадков, так и во время регулярных сизигийных приливов. Обнаружено, что в водоемах с пресной водой в присутствии неспециализированных хищников (личинок стрекоз, личинок плавунцов, гладышей, имаго жуков-ильников и тинников, мальков и взрослых особей колюшек трехиглой и девятииглой) элиминируется до 98 % популяции преимагинальных стадий комаров A. dorsalis. При отсутствии указанных хищников в соленых водоемах погибает только 60 % личинок A. dorsalis (Георгиева, 2004).
Известно, что самки A. dorsalis активно нападают на теплокровных животных и людей (Сазонова, 1959). В Новгородской области наблюдали нападения имаго A. dorsalis на человека и кролика на пойменных, суходольных лугах и лугах, используемых под выпас животных, на мелиоративных участках, в ивняках, на верховых болотах и в городских парках, расположенных на незначительном удалении от соленых источников или скважин (Панюкова, 2005). Нами не отмечены нападения данного вида на человека и животных внутри помещений, весьма вероятно, что A. dorsalis относится к облигатным экзофилам, которые охотятся и переваривают пищу только в природных условиях. При постановке эксперимента с открытым окном возле соленых озер в г. Старая Русса в течение суток для питания и нападения в жилое помещение не проник ни один экземпляр данного вида. В Республике Коми и г. Одессе также имаго собраны в природных биотопах. Таким образом, к экологическим особенностям изучаемого вида можно отнести полицикличность, факультативную галофильность и облигатную экзофильность.
Обсуждение
Особенность большинства видов кровососущих комаров, обитающих в северной части Палеарктики, – трансконтинентальные ареалы, которые охватывают значительную часть Палеарктики и даже Голарктики (Медведев и др., 2017). Вид A. dorsalis также имеет трансголарктический полизональный тип ареала (Панюкова, Остроушко, 2017), согласно классификации типов ареалов К. Б. Городкова (1984).
Реконструкция ареала A. dorsalis
Анализ и обобщение имеющихся материалов о распространении и экологических предпочтениях изучаемого вида позволили провести реконструкцию его ареала на территории европейской части России. Известно, что ареал A. dorsalis охватывает север Африки, Северную Америку и Евразию. При этом в Евразии вид имеет трансконтинентальное распространение, находки A. dorsalis известны от Британских до Японских островов. В лесной полосе вид приурочен к смешанным и лиственным лесам. По поймам, как интразональным элементам ландшафта, A. dorsalis может проникать в другие природные зоны: на юге – в степную и полупустынную, на севере – в таежную (Гуцевич и др., 1970). Отмечалось интразональное проникновение данного вида в тундровую зону на побережье Белого моря (Тамарина, Георгиева, 1981). Вид локально встречается в Субарктической зоне европейской части России. Считается, что продвижение данного вида на север ограничивается южной тундрой (Остроушко, 1987). При анализе распространения вида A. dorsalis на Евразийском континенте прослеживается его приуроченность к местам выхода соленых источников для внутриматериковых популяций (см. рис. 1). Из рассматриваемого материала вблизи минеральных соленых источников собраны имаго данного вида в с. Серегово, городах Старая Русса, Донецк, Пятигорск и Ессентуки. Весьма вероятно обнаружение внутриматериковых популяций данного вида в местах выхода минеральных источников. Северные находки A. dorsalis в континентальных условиях воркутинских южных тундр (Остроушко, 1987) связаны, по нашему мнению, с наличием благоприятных условий. В г. Воркуте имеются сероводородные соленые источники – потенциальные места развития личинок данного вида. В целом на севере европейской части России A. dorsalis встречается точечно, это связано с тем, что максимальная численность популяции данного вида достигается только в условиях приморских или внутриконтинентальных засоленных водоемов при отсутствии межвидовой конкуренции.
Выполнить реконструкцию ареала данного вида позволил географический и палеонтологический анализ. По достоверным данным род Aedes, к которому относится изучаемый вид, известен с эоцена (56 млн лет назад) из янтарей и отпечатков из различных регионов мира (Evenhuis, 1994). Палеонтологические материалы свидетельствуют о большой древности возникновения гематофагии у двукрылых насекомых: так, появление кровососания у комаров существует несколько десятков миллионов лет с конца мезозоя или начала кайнозоя (Балашов, 1999). Палеонтологические материалы о возникновении кровососания в семействе Culicidae и находки древних представителей рода Aedes, сохранившиеся в янтаре, позволяют судить об изменениях ареала A. dorsalis. Известно, что до начала оледенений территория севера Европейской равнины была покрыта широколиственными лесами – дубравами (Козубов, 1999). Здесь был теплый влажный климат, а видовой состав сем. Culicidae, вероятно, был близок к фауне современной Южной Европы. Примером этого служат островные участки типичных дубрав, которые сохранились в поймах крупных рек (Мста, Вятка) и озер (Ильмень, Нургуш). К началу наступления глобальных похолоданий в плейстоценовый (четвертичный) период была большая вероятность широкого распространения A. dorsalis по всей территории современной европейской субарктической зоны, включая берингийский сухопутный перешеек, который соединял материки. Плейстоценовые оледенения, охватывавшие значительную часть Евразии, привели к исчезновению здесь большей части фауны кровососущих комаров. Предположительно в местах выхода соленых или термальных подземных вод поверхностное оледенение было более слабым и протаивало, в таких условиях мог сохраниться на севере в рефугиумах вид A. dorsalis. После разъединения берингийского перешейка часть популяций данного вида стали развиваться в измененных условиях и незначительно отличаются генетически от европейских, о чем свидетельствует проведенное нами исследование. В Евразии, южнее максимальных границ Днепровского материкового оледенения, встречаются оба вида: A. dorsalis и A. caspius. При этом севернее максимальной границы Валдайского оледенения обнаружен только A. dorsalis. Теплолюбивый A. caspius не смог проникнуть на Северо-Американский континент по берингийскому «мосту» и остался только в южной части Евразии. Под воздействием похолодания большинство видов комаров были вытеснены с территории Субарктики на юг и за Уральскую гряду. Центрами формирования и восстановления фауны комаров европейской части в послеледниковый период, возможно, стала Средиземноморская область и Кавказ, а в азиатской части – Тянь-Шань и Тибет.
Заключение
Вид A. dorsalis встречается в широком диапазоне солености, и выживание данного вида в условиях высоких широт возможно благодаря его широкой экологической пластичности. Данный вид занял свободную экологическую нишу, развиваясь в водоемах с повышенной концентрацией солей (хлориды, сульфаты). Такие водоемы формируются в окрестностях выходов подземных вод разного химического состава на поверхность или в приливно-отливной зоне северных морей. Морские трансгрессии и регрессии в мезозое могли определять цикличность развития поколений, быть причиной полицикличности A. dorsalis. Подтверждением этого могут быть проведенные ранее исследования на побережье Белого моря, где отмечалась связь числа поколений данного вида с морскими приливами (Георгиева, 2004). Для большинства типичных представителей зональной фауны насекомых эктремальные условия засоления водоемов губительны. Вид A. dorsalis может занимать засоленные водоемы при низкой пищевой конкуренции и при сниженном количестве водных хищников, образуя локальные популяции в европейской части Субарктики. В южной части Русской равнины вид встречается совместно с A. caspius. В конкурентных условиях A. dorsalis уступает по частоте встречаемости и нападению на учетчика виду A. caspius. Морфологически, экологически и генетически виды достаточно хорошо различаются. Основные морфологические отличительные черты этих двух видов – в окраске спинки имаго. Географическая изменчивость окраски A. dorsalis проявляется в увеличении числа светлых чешуек на брюшке у южных популяций. Вероятно, увеличение светлых чешуек у южных популяций связано с функцией отражения солнечного света при его избытке для сохранения влаги, как защита от высыхания. Преобладание темных чешуек на брюшке северных популяций можно рассматривать в качестве приспособления к увеличению поглощения солнечного тепла, как защита от переохлаждения, при недостатке тепла на севере. Комар A. dorsalis развивается на севере при сниженном количестве водных хищников и низкой межвидовой конкуренции в соленых и содержащих сероводород водоемах. Количество поколений данного вида зависит от широты местности. На севере и северо-западе Русской равнины отмечены два поколения, на юге возможны более двух поколений.
В результате исторической реконструкции ареала комара A. dorsalis на основе анализа его прерывистого распространения и экологических особенностей вида можно предположить, что A. dorsalis присутствовал на изучаемой территории задолго до плейстоценовых оледенений. Вид проник на территорию Северной Америки по сухопутному берингийскому перешейку. После разъединения материков обособленные популяции вида эволюционировали самостоятельно. В плейстоценовый ледниковый период вид A. dorsalis был вытеснен на юг и юго-восток, а после отступления ледника распространился на север до побережья Субарктики по свободным экологическим нишам. Ядром ареала данного вида можно считать южную часть Палеарктики.
Библиография
Балашов Ю. С. Эволюция гематофагии среди насекомых и клещей // Энтомологическое обозрение. 1999. Т. 78. С. 749‒763.
Георгиева Е. К. Роль солености в распространении полизонального комара Aedes caspius Pall (Díptera, Culicidae) : Автореф. дис. ... канд. биол. наук. М., 2004. 22 с.
Герасимов И. П., Марков К. К. Четвертичная геология: палеогеография четвертичного периода : Учебное пособие для университетов и педагогических институтов. М.: Учпедгиз, 1939. 364 с.
Городков К. Б. Типы ареалов насекомых тундры и лесных зон европейской части СССР // Ареалы насекомых европейской части СССР. № 5. Л.: Наука, 1984. С. 3–20.
Гуцевич А. В. О политипических видах комаров (Culicidae) II. Aedes caspius (Pallas, 1771) // Паразитология. 1977. Т. 11. № 1. С. 48–51.
Гуцевич А. В., Мончадский А. С., Штакельберг А. А. Комары сем. Culicidae // Фауна СССР. Насекомые Двукрылые. Т. 3. Вып. 4. Л.: Наука, 1970. 384 c.
Кожевников Ю. П., Железнов-Чукотский Н. К. Берингия: история и эволюция . М.: Наука, 1995. 383 с.
Козубов Г. М. История лесного покрова Европейского Северо-Востока // Леса Республики Коми / Отв. ред. А. И. Таскаев. М.: Дизайн. Информация. Картография, 1999. С. 61–70.
Кункова Е. В. Гидробионты соленых вод курорта «Старая Русса» // Биоразнообразие, функционирование, продуктивность и охрана биосистем в Новгородской области: Материалы региональной научной конференции, 12–15 мая 2003 г., Великий Новгород. Великий Новгород: Изд-во Новгородского ун-та, 2003. С. 100.
Медведев С. Г., Халин А. В., Айбулатов С. В. Пути происхождения фауны кровососущих насекомых Северной Палеарктики на примере блох (Siphonaptera), комаров семейства Culicidae и мошек (Diptera: Simuliidae) // Паразитология. 2017. Т. 51. Вып. 6. С. 499‒516.
Оледенение и многолетняя мерзлота : Карта М. 1:24000000 // Атлас СССР. М: ГУГиК при Совете Министров СССР, 1984. С. 97.
Остроушко Т. С. Эколого-фаунистический обзор кровососущих комаров (Diptera, Culicidae) Большеземельской тундры // Труды Коми филиала Академии наук СССР. Сыктывкар: Изд-во Коми науч. центра Уральского отделения АН, 1987. № 86. С. 58–67.
Панюкова Е. В. Видовой состав кровососущих комаров (Diptera, Culicidae) населенных пунктов Северо-Запада России (на примере Новгородской области) // Структурно-функциональные особенности биосистем Севера (особи, популяции, сообщества): Материалы конференции. Петрозаводск, 2005. С. 76–79.
Панюкова Е. В. Кровососущие комары (Diptera, Culicidae) окрестностей курорта «Серегово» в Республике Коми // Достижения энтомологии на службе агропромышленного комплекса, лесного хозяйства и медицины: Тезисы докладов XIII съезда Русского энтомологического общества, 9–15 сентября 2007 г., Краснодар. Краснодар: Изд-во Кубанского гос. аграрного ун-та, 2007. С. 159–160.
Панюкова Е. В., Остроушко Т. С. Кровососущие комары (Diptera: Culicidae) / Отв. ред. М. М. Долгин. Т. XI. Ч. 2. М.: Товарищество научных изданий КМК, 2017. 209 с.
Полезные ископаемые : Карта М. 1:16000000 // Атлас СССР. М.: ГУГиК при Совете Министров СССР, 1984. С. 92.
Сазонова О. Н. Комары рода Aedes Рыбинского водохранилища и обзор фауны рода Aedes лесной зоны Европейской части РСФСР // Труды Дарвинского государственного заповедника. Вологда, 1959. № 5. С. 209–303.
Смульский И. И., Иванова А. А. Опыт реконструкции палеоклимата по изменению инсоляции на примере Западной Сибири в позднем плейстоцене // Климат и природа. 2018. № 1 (26). С. 3‒21.
Тамарина Н. А., Георгиева Е. К. Интразональное проникновение комара Aedes caspius dorsalis Mg. (Diptera, Culicidae) на север и зависимость его развития от ритма морских приливов на побережье Белого моря // Биологические науки. 1981. № 4. С. 36–41.
Храброва Н. В., Андреева Ю. В., Ваулин О. В., Алексеева С. С., Сибатаев А. К. Изменчивость нуклеотидной последовательности митохондриального гена субъединицы I цитохромоксидазы С у видов родов Aedes и Ochlerotatus (Diptera, Culicidae) // Вавиловский журнал селекции и генетики. 2013. Т. 17. № 1. С. 114–122.
Becker N., Petric D., Zgomba M., Boase C., Dahl C., Madon M., Kaiser A. Mosquitoes and their сontrol. Second Edition. Heidelberg; Dordrecht; London; New York, 2010. 577 p.
Cao Y., Fu S., Tian Z., Lu Z., He Y., Wang H., Wang J., Guo W., Tao B., Liang G. Distribution of mosquitoes and mosquito-borne arboviruses in Inner Mongolia, China // Vector Borne Zoonotic Dis. 2011. Vol. 11 (12). P. 1577–1581. DOI: 10.1089/vbz.2010.0262.
Evenhuis N. L. Catalogue of the fossil flies of the world (Insecta: Diptera). Leiden, 1994. 654 p.
Gimnig J. E., Eldridge B. F. Genetic and Morphological Characterization of the Aedes (Ochlerotatus) dorsalis (Diptera: Culicidae) Group in North America // Journal of Medical Entomology. 1999. Vol. 36. Issue 6. P. 685–694. DOI: 10.1093/jmedent/36.6.685.
Greninger A. L., Makhsous N., Shean R., Jerome K. R., Kangiser D., Dykstra E., Haman K. Submitted (23-SEP-2016) Laboratory Medicine, UW, 1616 Eastlake Ave East, Suite 320, Seattle, WA 98102, USA. URL: https://www.ncbi.nlm.nih.gov/nuccore/KX898491.1 (дата обращения: 8.04.2020).
Hughes H. R., Lanciotti R. S., Blair C. D., Lambert A. J. Submitted (02-SEP-2016) Division of Vector-borne Diseases, Centers for Disease Control and Prevention, 3156 Rampart Rd, Fort Collins, CO 80521. URL: https://www.ncbi.nlm.nih.gov/nuccore/KX817313.1 (дата обращения: 8.04.2020).
Liu H., Gao X. Y., Fu S. H., Li M. H., Zhai Y. G., Meng W. S., Sun X. H., Lv Z., Wang H. Y., Shen X. X., Cao Y. X., He Y., Liang G. D. Molecular evolution of emerging Banna virus // Infect. Genet. Evol. 2016. Vol. 45. P. 250–255.
Lu Z., Liu H., Fu S., Lu X., Dong Q., Zhang S., Tong S., Li M., Li,W., Tang Q., Liang G. Liao ning virus in China // Virol. J. 2011. Vol. 8. P. 282. DOI: 10.1186/1743–422 X–8–282.
Kumar S., Stecher G., and Tamura K. MEGA7: Molecular Evolutionary Genetics Analysis version 7 for bigger datasets // Molecular Biology and Evolution. 2016. Vol. 33. P. 1870–1874.
Mihalyi F., Gulyas М. Magyarorszag csipo szunyogjai. Budapest: Akademiai Kiadó, 1963. 229 p.
Minař J. Family Culicidae. Catalogue of Palaearctic Diptera. Vol. 2. Psychodidae – Chironomidae. Budapest: Akadémiai Kiadó, 1990. P. 74–113.
Svendsen J. I., Astakhov V. I., Bolshiyanov D. Yu., Demidov I., Dowdeswell J. A., Gataflin V., Hjort C., Hubberten H. W., Larsen E., Mangerud J., Melles M., Moller P., Saarnisto M., Siegert M. J. Maximum extent of the Eurasian ice sheets in the Barents and Kara Sea region during the Weichselian // Boreas. 1999. Vol. 28. № 1. Р. 234‒242.
Turell M. J., Wilson W. C., Bennett K. E. Potential for North American mosquitoes (Diptera: Culicidae) to transmit rift Valley fever virus // Journal of Medical Entomology. 2010. Vol. 47. № 5. P. 884–889. DOI: 10.1093/jmedent/47.5.884.
Благодарности
Искренне благодарим сотрудников Института биологии ФИЦ Коми научного центра УрО РАН А. Н. Панюкова и Е. С. Кузьмину, оказавших неоценимую помощь при подготовке и оформлении статьи. При выполнении работы использована фондовая коллекция кровососущих комаров Зоологического института РАН (УФК ЗИН рег. № 2-2.20, контракт с Роснаукой «02.452.11.7031» (2006-РИ-26.0/001/070)). Работа выполнена при финансовой поддержке госбюджетной темы отдела экологии животных Института биологии ФИц УрО РАН: «Систематика, распространение и пространственная организация фауны и населения наземных и водных животных таежных и тундровых экосистем европейского северо-востока России» (2018–2020 гг., № гос. регистрации: АААА-А17-117112850235-2).

 © 2011 - 2026
© 2011 - 2026