



Введение
Палеонтологическая летопись предоставляет обширную информацию о прошлом разнообразии млекопитающих и видовом богатстве. Эти данные имеют особое значение для понимания исторических закономерностей формирования наземных экосистем в позднем плейстоцене – среднем голоцене (MIS 3 – MIS 1: ≈ 50–5 тыс. лет назад) в период резких изменений глобального климата Земли, но еще без экстенсивного воздействия хозяйственной деятельности человека. Эта информация лежит в основе гипотез о воздействии на биоту аналогичных климатических изменений в будущем.
История европейской фауны млекопитающих в позднем плейстоцене – голоцене Европы достаточно полно документирована на основе анализа сотен и тысяч местонахождений (Смирнов и др., 1990; Маркова и др., 2008; Данукалова и др., 2018; Bachura, Kosintsev, 2007; Ponomarev et al., 2012; Puzachenko, Markova, 2014; Álvarez-Lao and Méndez, 2016; Pavelková Řičánková et al., 2015; Crees et al., 2016; Dinnis et al., 2016; Royer et al., 2016; Fernández-García et al., 2016; Kosintsev et al., 2016; Discamps and Royer, 2017 и мн. др.).
Одним из ключевых климатических событий рассматриваемого временного интервала был переход от последнего оледенением позднего плейстоцена к межледниковью голоцена. Формальная стратиграфическая граница между плейстоценом и голоценом принимается по (Walker et al., 2009) и находится в районе 11700 тыс. лет с максимальной шибкой 99 лет. Однако и до и после этой условной границы фауна претерпевала разнообразные трансформации.
Наши исследования состава региональных фаун крупных и среднеразмерных млекопитающих Восточной Европы (Puzachenko, Markova, 2014, 2016) показали, что изменения видового состава в конце позднего плейстоцена были нелинейными и необратимыми. Переход между комплексами крупных млекопитающих плейстоцена и голоцена был очень резким, практически скачкообразным. Последующие исследования, включающие всю фауну млекопитающих Европы, датируемую интервалом ≈ 50–5 тыс. лет назад, выявили гораздо более сложную и неоднозначную реакцию комплексов млекопитающих на изменения глобального климата, в том числе на уровне отдельных регионов (Маркова, Пузаченко, 2017; Puzachenko, Markova, 2019).
Удивительный феномен «дисгармоничных»/«безаналоговых» географических ассоциаций видов млекопитающих (Semken, 1988; Semken et al., 2010) позднего плейстоцена не только на территории Европы заключался в сочетании высокого видового разнообразия млекопитающих с высокой устойчивостью всего комплекса на фоне очень сурового климата валдайского (= вислинского, вюрмского) оледенения. Кроме низких температур и аридности, климат последнего оледенения характеризовался высокой вариабельностью климатической обстановки, не характерной для климата современного межледниковья (Svendsen et al., 2004). «Дисгармоничность» фаун оценивается относительно современных фаунистических комплексов. Многие виды, входившие в состав евразийских фаунистических комплексов «мамонтовой степи» в плейстоцене, в голоцене имеют сильно различающиеся экологические ниши, занимают аллопатрические ареалы, не встречаются в составе одних и тех же сообществ. Характерной чертой плейстоценовых комплексов было присутствие в них крупных травоядных и хищных млекопитающих («мегафауна»). Этот важнейший экосистемный компонент практически полностью исчез в конце плейстоцена, а немногие растительноядные представители мегафауны, пережившие границу плейстоцена и голоцена (мамонт, зубр, бизон, первобытный бык, гигантский олень), не смогли пережить экстремально теплый период климатического оптимума среднего голоцена или были истреблены человеком уже в средние века. Трансформация евразийских экосистем, и в том числе формирование сплошной лесной зоны в последние 10 тыс. лет, обусловили эволюцию современных зональных межледниковых экосистем, которые в подавляющем большинстве случаев не обнаруживают аналогов в позднем плейстоцене. В наибольшей степени эти преобразования природной среды затронули территорию Европы.
Понимание механизмов саморазвития экосистем в прошлом путем выявления ограничений (правил) варьирования их важнейших параметров должно лежать в основе создания современных эффективных технологий управления (природопользования) и вероятностного прогноза эволюции экосистем в будущем (Пузаченко, 2017). В связи с этим особый интерес представляет период трансформации/перестройки позднеплейстоценовых экосистем, сопровождавшийся не только хорошо известным вымиранием мегафауны, но в гораздо большей степени изменением ареалов видов, которые в настоящее время либо отсутствуют на территории Европы, либо их ареалы резко сократились или, наоборот, расширились в голоцене.
На основе обширного палеонтологического материала нами была реконструирована эволюция фаун млекопитающих 19 регионов Европы (Puzachenko, Markova, 2019).
Результаты этой работы послужили основой данного исследования. Материал позволяет дать довольно точную оценку временного интервала «перехода» между двумя глобальными состояниями региональных фаун на рубеже плейстоцена и голоцена. Кроме этого мы определили ряд ключевых параметров биологического разнообразия фаунистических комплексов до перехода, в период трансформации фаун и в первой половине голоцена.
Основная цель исследования состояла в поиске общих закономерностей (инвариантов), отражающих процесс эволюционных изменений параметров биологического разнообразия (видовое богатство, энтропия, индекс доминирования (Simpson, 1949), мера организации или избыточности (Shannon, 1948; Foerster, 1960)), широко используемых в экологических исследования современных экосистем. Кроме перечисленных выше, в работе применялись параметр β ранговых распределений и индекс «оборота видов» Вильсона и Шмида βt (Mourelle, Ezcurra, 1997), в качестве аналога индекса бета-разнообразия Уиттекера. При этом мы исходили из рабочей гипотезы о том, что «переход» фаунистических комплексов млекопитающих от их состояния в плейстоцене к состоянию в голоцене был неравновесным, необратимым процессом, сопровождавшимся потерей устойчивости сообществ млекопитающих.
Материалы
Зоогеографическое зонирование Европы, принятое в этой работе (рис. 1), основано на анализе современных ареалов наземных млекопитающих, представленных в базе данных МСОП (Terrestrial Mammals. IUCN, 2016. The IUCN Red List of Threatened Species. Version, 2016–1. http://www.iucnredlist.org/technical-documents/spatial-data). Принципы зонирования с использованием ГИС-технологий описаны в (Heikinheimo et al., 2007; Kreft, Jetz, 2010). Детализация методики нашего варианта зонирования приведена в работах (Puzachenko, Markova, 2016; Markova, Puzachenko, 2018). Основанием для использования современного зоогеографического зонирования для палеореконструкций послужили результаты исследований, обобщенных в (Маркова и др., 2008), где была показана относительная устойчивость положения основных зональных границ на территории Европы в конце позднего плейстоцена и в голоцене.
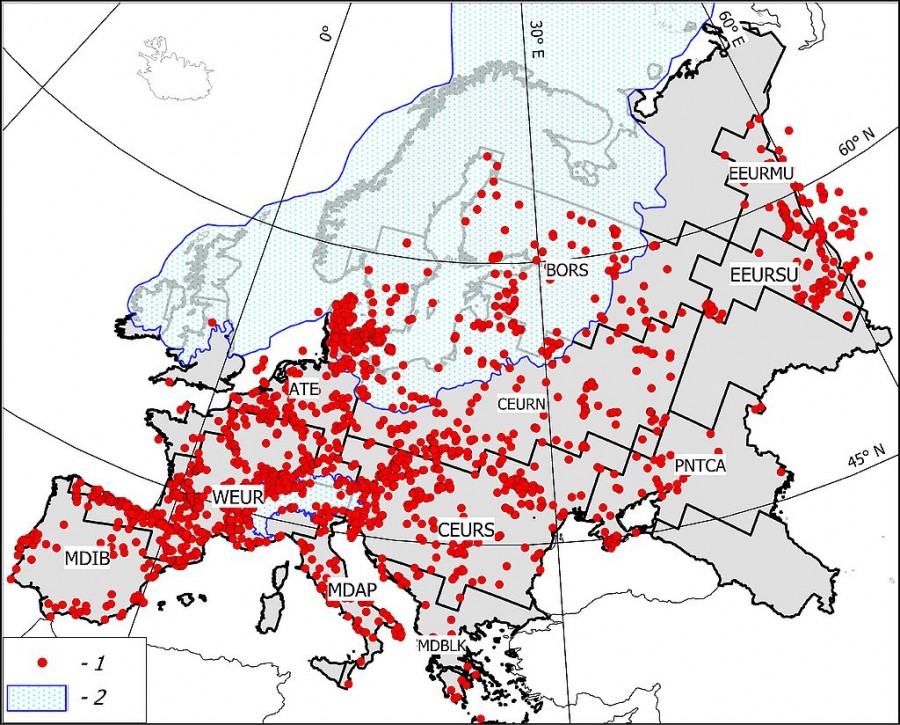
Рис. 1. Схема 11 регионов, упомянутых в статье, и местоположения палеонтологических местонахождений, датированных второй половиной позднего плейстоцена – средним голоценом. Расшифровка аббревиатур регионов см. в табл. 1. 1 – схема скандинавского ледникового щита в максимум валдайского оледенения (изотопная стадия MIS 2), 2 – местонахождения
Fig. 1. A diagram of the 11 regions mentioned in the paper and the paleontological locations dated the second half of the Late Pleistocene – the Middle Holocene. Explanation of the abbreviations of the regions, see table. 1. 1 – Scandinavian ice sheet at the maximum of the Valdai Glaciation (isotopic stage MIS 2), 2 – locations
В данной работе представлены результаты изучения параметров разнообразия по 11 региональным комплексам млекопитающих (табл. 1). Информационная база исследования взята из палеонтологической библиографической базы данных PALEOFAUNA (Markova et al., 1995), которая была собрана и поддерживается авторами на протяжении последних 25 лет (регистрационный номер 2011620493, Институт географии РАН). В целом был проанализирован состав 2593 местонахождений млекопитающих, датированных в основном радиоуглеродным методом.
Таблица 1. Названия и аббревиатура биорегионов, число местонахождений и общее число видов (NSP) в палеонтологической летописи
Table 1. Names of the biogeographic regions and their abbreviations, number of locations, and number of species in a paleontological record
| Биорегион | Аббревиатура | Число местонахождений |
NSP |
| Атлантический восточный (Atlantic East) | ATE | 597 | 109 |
| Бореальный южный (Boreal South) | BORS | 208 | 41 |
| Центрально-Европейский северный (Central European North) | CEURN | 385 | 119 |
| Центрально-Европейский южный (Central European South) | CEURS | 795 | 134 |
| Восточно-Европейский среднеуральский (East European Middle Ural) | EEURMU | 134 | 75 |
| Восточно-Европейский южноуральский (East European Southern Ural) | EEURSU | 137 | 81 |
| Средиземноморский апеннинский (Mediterranean Apennine) | MDAP | 304 | 81 |
| Средиземноморский балканский (Mediterranean Balkan) | MDBLK | 109 | 75 |
| Средиземноморский пиренейский (Mediterranean Iberian) | MDIB | 381 | 92 |
| Черноморско-Каспийский (Ponto-Caspian) | PNTCA | 145 | 65 |
| Западноевропейский (West European) | WEUR | 2593 | 143 |
Методы
Все радиоуглеродные (14С) даты калибровались с использованием калибровочной кривой Intcal13 в программы OxCal 4.3 (Ramsey, Lee, 2013; Reimer et al., 2013). Используется неравномерная временная шкала, что обусловлено гиперболическим ростом «ошибки» (среднеквадратическое отклонение) дат в зависимости от возраста местонахождения (Royer et al., 2016; Puzachenko, Markova, 2019). В итоге для каждого региона все данные о встречаемости местонахождений млекопитающих были агрегированы по 30 временным интервалам с длиной последних от 620 лет (поздний голоцен) до 5.6 тыс. лет (~ < 50 тыс. лет). Практически мы использовали меньшее число интервалов, начиная со среднего голоцена, поскольку более поздние и соответственно короткие интервалы шкалы не были обеспечены репрезентативным палеонтологическим материалом.
Для каждого интервала временной шкалы определяли общее число видов и для каждого вида – частоту встречаемости местонахождений, в которых он был обнаружен. Распределение относительных «нормализовывалось», для чего использовали арксинусное преобразование (Sokal and Rohlf, 1995). Каждый интервал был охарактеризован набором частот местонахождений разных видов («описательные переменные»). Отношения сходства/различия между интервалами задавились метрикой. В качестве метрики использовали индекс Брея – Куртиса (Bray, Curtis, 1957; Ricotta, Podani, 2017), который описывает отношения между всеми парами интервалов по соотношению встречаемости местонахождений с разными видами млекопитающих.
Таким образом, «система» как объект моделирования определяется как совокупность множества элементов (интервалы временной шкалы), их описательных переменных и метрики, задающую отношения между всеми элементами. Задача моделирования состоит в выделении упорядоченных отношений между элементами системы на фоне случайного шума. При этом исходные данные для модели содержатся в матрице значений индекса Брея – Куртиса. Далее эта матрица обрабатывалась методом неметрического многомерного шкалирования (Davison, 1983) с целью ординации временных интервалов в евклидовом пространстве небольшой размерности (подробнее см. в (Puzachenko, Markova, 2014, 2016)). По условию получения, координаты модели (обозначаются в работе BC1, BC2 и BC3) содержат основную информацию об упорядоченных (не стохастических) компонентах временной динамики структуры региональной фауны. Размерность моделей варьировала от 2 до 5. Координаты моделей являются инвариантами изменчивости встречаемости отдельных видов комплексов. Также они могут интерпретироваться и как параметры порядка «системы» в контексте синергетической модели.
Было показано (Puzachenko, Markova, 2019), что BC1 и/или BC2 коррелируют δ18O (‰) (медианное значение показателя рассчитывалось для каждого временного интервала по ряду North Greenland Ice Core Project Oxygen Isotope Data, NGRIP (Andersen et al., 2004)) и видовым богатством.
Результаты моделирования используются нами здесь только для определения границ временного интервала, на котором происходила перестройка регионального фаунистического комплекса. Для этого строились графики между последовательными значениями координат модели (диаграмма Ламерея, но без аппроксимации данных полиномом). Так как по условию координаты BC1 и BC2 не коррелируют друг с другом, то интервал «перехода» для каждой из них может и не совпадать или может быть выражен только по одной из них. Другими словами, предполагается, что по одним свойствам системы границы перехода могут быть выражена, а по другим, не зависимым от них свойствам, может не наблюдаться никаких изменений (подробнее см., например (Пузаченко, 2017)).
Для характеристики биологического разнообразия трех предполагаемых состояний регионального фаунистического комплекса («плейстоцен», переход, «голоцен») применяли: видовое богатство (NSP) – число видов, зафиксированное в региональной палеонтологической летописи, индекс доминирования
D = ∑i(ni/N)2, 0 < D < 1,
где ni – число местонахождений с видом i, N – общее число местонахождений,
энтропию Шеннона, H = -∑i(ni/N) ∙ ln(ni/N), меру организации или «избыточности» (redundancy), где NSP – число видов.
Для сравнения таксономического разнообразия разных состояний фаунистических комплексов использовали модификацию индекса «оборота видов» Вильсона – Шмида (Mourelle, Ezcurra, 1997):
βt = (g + l)/(2α ∙ (n-1),
где g – прирост числа обнаруженных таксонов на «градиенте» между последовательными состояниями, l – число «исчезнувших» таксонов на том же «градиенте», α – среднее число таксонов для всех состояний, n – число одновременно сравниваемых состояний (0 ≤ βt. ≤ 1).
В современной экологии модели ранговых распределений являются традиционным инструментом исследования структуры сообществ (Левич, 1978, 1980, 2007; Булгаков, Левич, 2005; Пузаченко, 2009; 2013; 2016; Hutchinson, 1957; Margalef, 1957; Tokeshi, 1993; Hubbell, 1997; Marquet, 2000, 2005; Magurran, 2004; McGill et al., 2007 и мн. др.). Наиболее общее теоретическое обоснование ранговых распределений дано А. П. Левичем как следствие применения экстремального принципа к системам, реализация которых ограничена некоторыми широко трактуемыми переменными – «ресурсами среды» (Левич, 1978; Булгаков, Левич, 2005).
В нашем исследовании упорядоченные по убыванию распределения частот встречаемости местонахождений с разными таксонами для трех рассматриваемых состояний фаунистического комплекса сравнивались с тремя типами модельных ранговых распределений. Относительно простые модели ассоциируются с гиперболическим распределением Гиббса (или геометрическую модель Мотомуры), известного из статистической механики (далее обозначается аббревиатурой G):
p(r) = exp(λ + β ∙ r),
где p(r) – частота встречаемости местонахождений вида с рангом r (местонахождения с наиболее часто встречаемым видом имеет ранг 1), λ и β – параметры модели.
Параметр β характеризует степень выравненности распределения. В этой модели предполагается линейный вид функции чувствительности вида к варьированию ресурсов.
Распределения Ципфа (Z) и Ципфа – Мандельброта имеют следующий вид:
(Z) p(r) = exp(λ + β ∙ ln(r)) и (ZM) p(r) = exp(λ + β ∙ ln(r + d)) (d – дополнительный параметр, предложенный Б. Мандельбротом (1954)).
Последние два распределения характеризуют системы, в которых чувствительность видов к варьированию ресурса подчиняется логарифмической зависимости. К модели ZM распределения очень близка по смыслу и результатам применения модель разломанного стержня Мак-Артура, в которой «вид» – «ресурс» подчиняется двойной логарифмической зависимости (~ ln(ln(r))).
В рамках решаемой здесь задачи выбор между ZM распределением и моделью «разломанного стержня» на основе статистических критериев был затруднен в большинстве случаев, когда оба распределения удовлетворительно описывали наблюдения. Поэтому мы остановили свой выбор на более простой ZM модели.
Ю. Г. Пузаченко (2016) предложил модификации рассмотренных моделей, допустив, что продолжительность существования вида в сообществе может быть пропорциональна (с разным знаком) его рангу. Мы предварительно рассмотрели соответствующие модификации G, Z и ZM распределений и выяснили, что часто модификация ZM распределения (ZMst: p(r) = exp(λ + ln(r + d) ∙ (с + β ∙ ln(r + d))) давала наилучший результат аппроксимации. ZMst распределение не соответствует модели равновесия, а отражает стационарное состояние сообщества, когда «направленные» изменения в нем происходят достаточно медленно, не катастрофично, т. е. скорости этих изменений не сильно отличаются от 0.
Для статистической обработки данных использовали программные продукты STATISTICA v. 8.0 (StatSoft, Tulsa, OK, USA) и PAST v. 3.12 (Hammer et al., 2001).
Результаты
Время и длительность перестройки региональных фаунистических комплексов на рубеже плейстоцена и голоцена
Длительность временного интервала, в течение которого происходил переход фаунистических комплексов млекопитающих от состояния, характерного для позднего плейстоцена, к состоянию, реализованному в первой половине голоцена, определялась на графиках изменений инвариантов (координат модели) BC1 или BC2. Для большинства региональных комплексов только координата BC1, которая содержит основную информацию об упорядоченном изменении частот встречаемости местонахождений с разными видами, демонстрировала черты динамики, позволяющие рассматривать весь процесс как последовательное во времени изменение «состояния» сообщества (табл. 2). В большинстве случаев эти изменения четко ассоциировались с параллельным изменением климатической обстановки и динамикой видового богатства.
Таблица 2. Оценка времени (средняя, время начала и завершения процесса, тыс. лет назад) изменений состояния региональных фаун в конце позднего плейстоцена – голоцене Европы относительно первой (BC1) и второй (BC2) координат модели; коэффициенты корреляции Спирмена и их статистическая значимость (p) с климатической переменной δ18O (‰) и видовым богатством (SPN)
Table 2. Meantime, start and end time of the transformation (Kyr. BP) in a European regional faunal assemblage' state between the End of the Late Pleistocene and the first part of the Holocene, which reproduced in BC1 and BC2 axes of non-metric multidimensional scaling model; Spearman correlation coefficient with its statistical significance (p) for the climate variable δ18O (‰) or species richness (SPN) and BC1/ BC2 axis is presented
| Регион | BC1 | BC2 | BC1 | BC2 | ||
| δ18O, p | SPN, p | δ18O, p | SPN, p | |||
| ATE | 7.6, 8.7–6.6 | – | 0.53, <0.01 | -0.69, <0.01 | -0.78, <0.01 | не достов. |
| BORS | 13.4, 15.8–11.0 | 13.3, 14.7–11.9 | 0.74, <0.01 | 0.81, <0.01 | 0.45, <0.05 | не достов. |
| CEURN | 12.0, 13.7–10.2 | – | 0.74, <0.01 | -0.81, <0.01 | не достов. | не достов. |
| CEURS | 13.0, 15.8–10.2 | – | 0.88, <0.01 | -0.60, <0.01 | не достов. | 0.55, <0.01 |
| EEURMU | 16.9, 18.0–15.8 | – | 0.81, <0.01 | не достов. | не достов. | 0.62, <0.01 |
| EEURSU | – | 14.8, 15.8–13.7 | не достов. | не достов. | -0.84, <0.01 | 0.55, <0.01 |
| MDAP | – | 8.3, 9.4–7.2 | 0.47, <0.05 | -0.51, <0.01 | -0.65, <0.01 | не достов. |
| MDBLK | 12.3, 12.8–11.9 | 8.0 (?), 8.7–7.4 | 0.85, <0.05 | не достов. | не достов. | не достов. |
| MDIB | 11.1, 11.9–10.2 | - | 0.77, <0.01 | -0.92, <0.01 | -0.44, <0.05 | не достов. |
| PNTCA | 11.9, 12.8–11.0 | - | 0.38, <0.04 | -0.62, <0.05 | не достов. | 0.48, <0.05 |
| WEUR | 11.6, 13.7–9.4 | 3.0, 15.7-10.2 | 0.78, <0.01 | -0.87, <0.01 | -0.86, <0.01 | 0.52, <0.01 |
| Примечание. «–» – изменения структуры комплекса между плейстоценом и голоценом не выражены. | ||||||
Можно выделить три варианта изменений состояния комплексов, характеризующих разные региональные комплексы. Для первого варианта характерен очень резкий относительно осей абсциссы и ординаты диаграммы Ламерея, почти скачкообразный в масштабе шкалы времени переход между двумя состояниями плейстоцена и голоцена (рис. 2А, 2Б, 2В). В эту группу входят регионы PNTCA, CEURN, MDBLK, ATE и BORS.
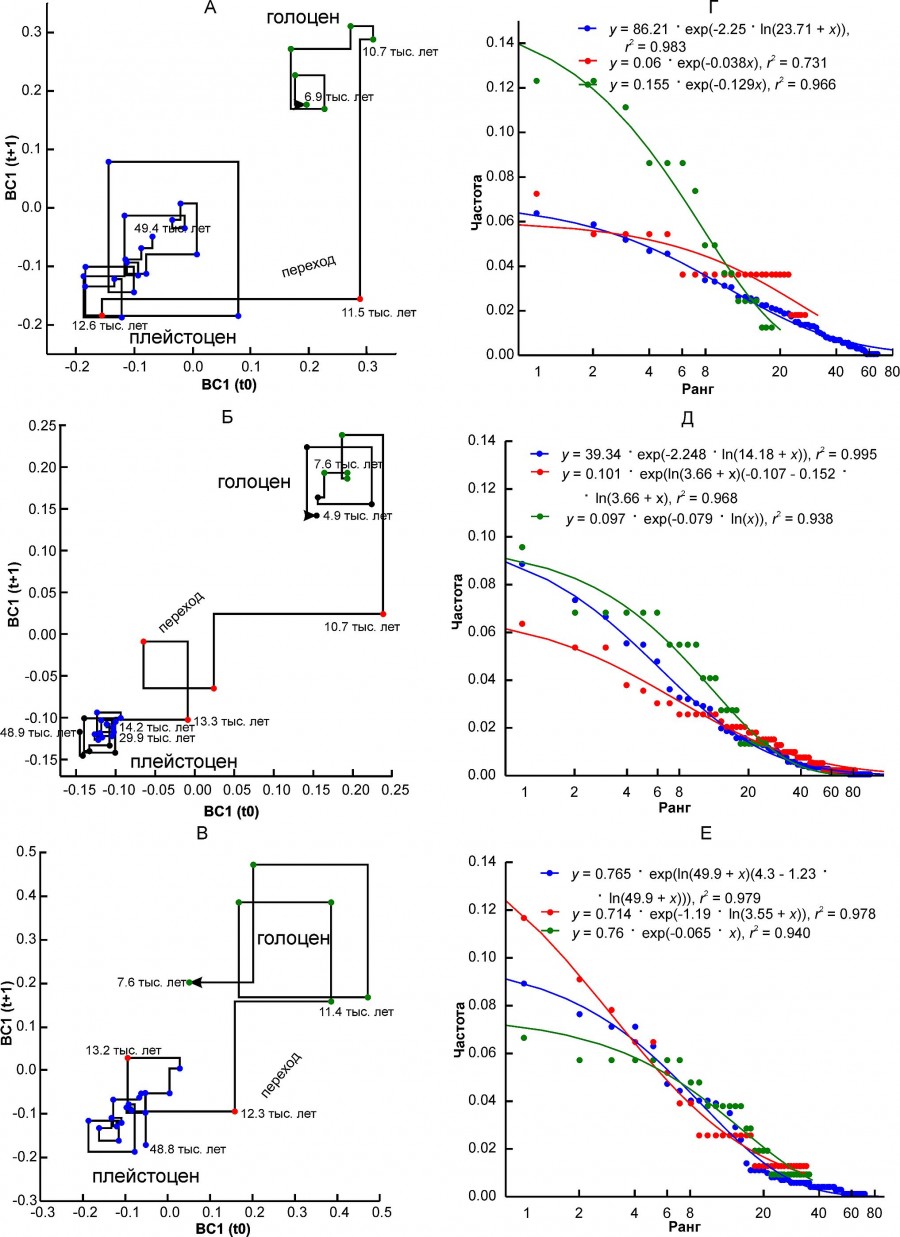
Рис. 2. Диаграмма Ламерея первой координаты BC1 и ранговые распределения встречаемости местонахождений, соответствующих разным состояниям регионального фаунистического комплекса. A, Г – регион PNTCA, Б, Д – регион CEURN, В, Е – регион MDBLK. Разным цветом выделены точки, соответствующие трем состояниям комплекса: синий – «плейстоцен», красный – переходный, зеленый – «голоцен». Цифры на графиках обозначают возраст в тыс. лет
Fig. 2. Lamerey diagram of BC1 coordinate and rank distributions for three states of a regional faunal assemblage. A, Г – region PNTCA, Б, Д – region CEURN, В, Е – region MDBLK. The points corresponding to the three states of the assemblage are highlighted in different colors: blue – "Pleistocene", red – "transitional", green – "Holocene".The numbers on the graphs indicate the age in thousand years
На графиках видно, что состояние фаунистических комплексов в плейстоцене было относительно стабильно. Амплитуда колебаний значений BC1 в голоцене выше, что, однако, может быть и артефактом укорочения шага нашей временной шкалы.
Во время «перехода» происходит потеря устойчивости комплекса, что хорошо отражено на графиках. В целом такой тип графика внешне, а возможно, и содержательно соответствует схеме фазового перехода первого рода. Длительность периода неустойчивости варьирует в разных регионах от 4.8 тыс. лет до ~ 1 тыс. лет (см. табл. 2). Переход мог завершиться до формальной границы плейстоцена и голоцена (MDBLK) или после нее. В регионе ATE перестройка комплекса началась с большой задержкой, в бореальной подстадии голоцена, а завершилась только в среднем голоцене (атлантическая подстадия). Отметим, что время окончания завершения постплейстоценовой трансформации фауны в голоцене для европейских экосистем очень условно. Начиная с позднего неолита драйвером эволюции, особенно западноевропейской фауны, выступала деятельность человека, предотвращающая достижение равновесного состояния на уровне региональных комплексов.
Другой вариант динамики характеризуется постепенными изменениями в направлении от состояния плейстоцена к состоянию голоцена (рис. 3A, 3Б, 3В). При этом относительно устойчивые начальное и конечное состояния комплекса иногда трудно поддаются выделению на графике. Регион EERMU (рис. 3А) занимает промежуточное положение между регионами с первым-вторым вариантами динамики фаунистических комплексов. Кроме региона EERMU, в последнюю группу входят регионы MDIB, WEUR и CEURS. Длительность перехода варьирует от 4.3 тыс. лет (WEUR) до ~ 1.7 тыс. лет (MDIB). Период основных преобразований в основном попадает на интервал от 13.7 тыс. лет до 10.2 тыс. лет, т. е. от первого плейстоценового интерстадиального потепления беллинга – аллереда до начала бореальной подстадии голоцена. Перестройка комплекса среднего Урала (EEURMU) произошла примерно на 2 тыс. лет раньше, в позднеледниковье (см. табл. 2).
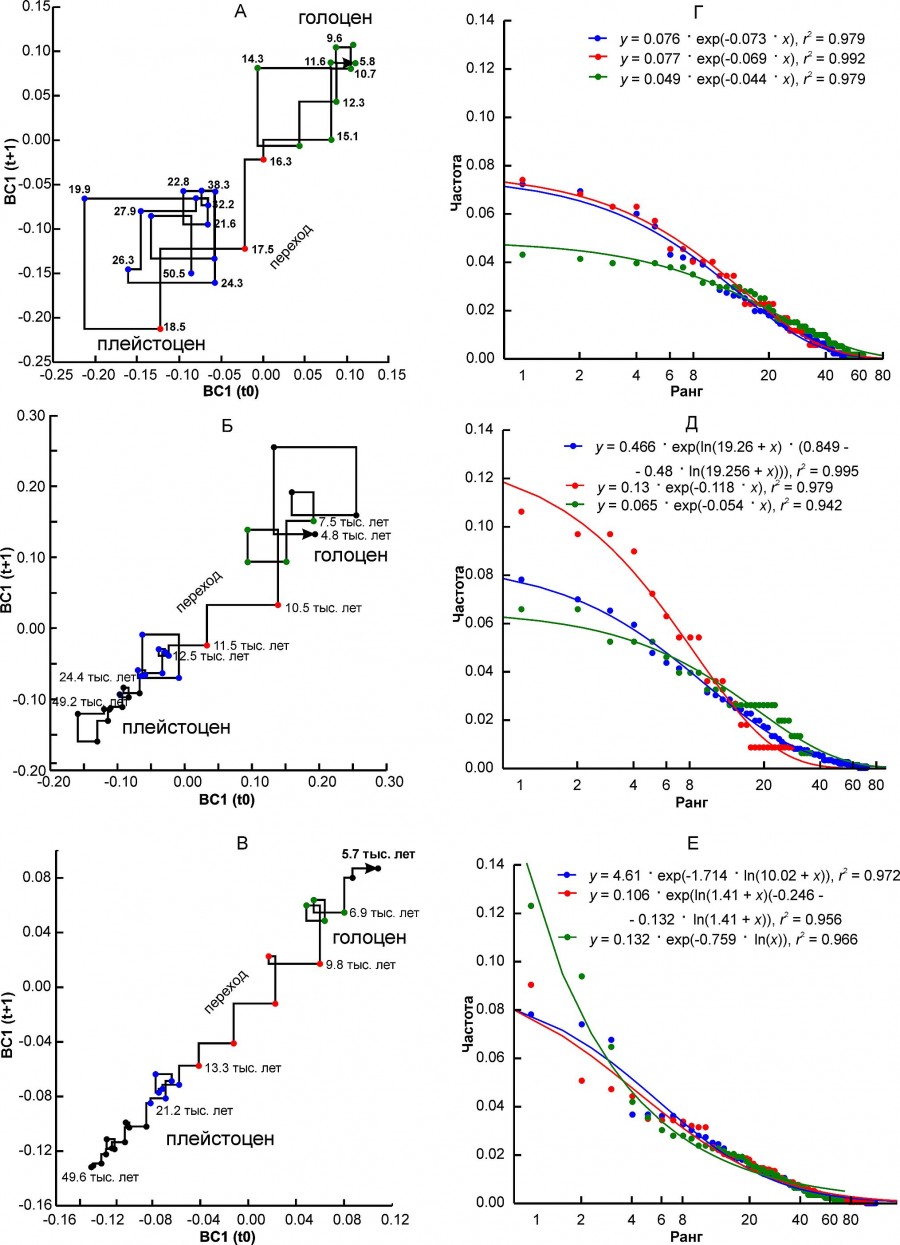
Рис. 3. Диаграмма Ламерея первой координаты BC1 и ранговые распределения встречаемости местонахождений, соответствующих разным состояниям регионального фаунистического комплекса. A, Г – регион EEURMU, Б, Д – регион MDIP, В, Е – регион WEUR. Разным цветом выделены точки, соответствующие трем состояниям комплекса: синий – «плейстоцен», красный – переходный, зеленый – «голоцен». Цифры на графиках обозначают возраст в тыс. лет
Fig. 3. Lamerey diagram of BC1 coordinate and rank distributions for three states of a regional faunal assemblage. A, Г – region EEURMU, Б, Д – region MDIP, В, Е – region WEUR. The points corresponding to the three states of the assemblage are highlighted in different colors: blue – "Pleistocene", red – "transitional", green – "Holocene". The numbers on the graphs indicate the age in thousand years
Комплексы млекопитающих Апеннинского полуострова (MDAP) и Южного Урала (EEURSU) составляют третью группу, в которой мы не обнаружили признаков перестройки комплекса на границе плейстоцена и голоцена по координате BC1 (рис. 4А, 4Б, см. табл. 2). Это означает, что те изменения, которые естественно происходили в это время (вымирание, миграции и т.п.), не существенно повлияли на параметры структуры. Вторая координата моделей BC2 отражает эти изменения (см. табл. 2), которые на Апеннинском полуострове произошли в первой половине голоцена, а на Урале – гораздо раньше, в позднеледниковье. Причем в обоих случаях ведущим внешним фактором были изменения климата, о чем свидетельствуют коэффициенты корреляции BC2 c δ18O.
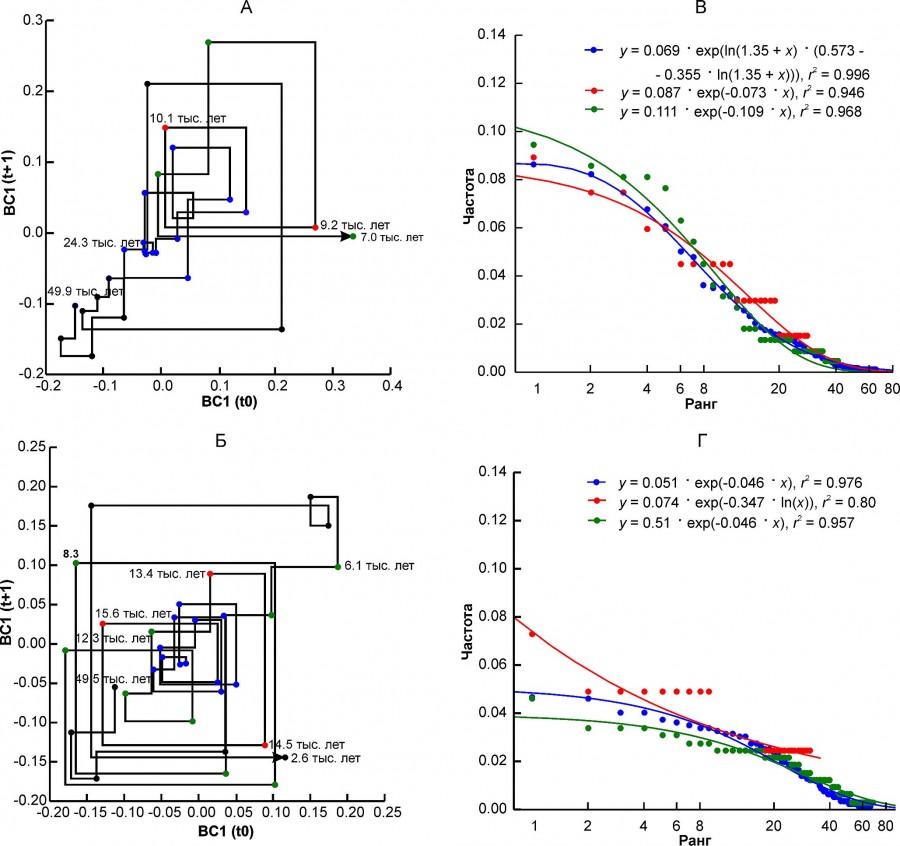
Рис. 4. Диаграмма Ламерея первой координаты BC1 и ранговые распределения встречаемости местонахождений, соответствующих разным состояниям регионального фаунистического комплекса. A, В – регион MDAP, Б, Г – регион EEURSU. Разным цветом выделены точки, соответствующие трем состояниям комплекса: синий – «плейстоцен», красный – переходный, зеленый – «голоцен». Цифры на графиках обозначают возраст в тыс. лет
Fig. 4. Lamerey diagram of BC1 coordinate and rank distributions for three states of a regional faunal assemblage. A, В – region MDAP, Б, Г – region EEURSU. The points corresponding to the three states of the assemblage are highlighted in different colors: blue – "Pleistocene", red – "transitional", green – "Holocene". The numbers on the graphs indicate the age in thousand years
Из данных, приведенных в табл. 2, следует, что изменения глобального климата выступали в качестве основного драйвера эволюции структуры региональных фаун. В основном климатический фактор влиял на колебания видового богатства фаун (переменная NSP). Координата BC2 содержит дополнительную независимую информацию о влиянии климата на встречаемость отдельных видов или основной климатически обусловленный тренд, как в случае с регионами MDAP и EEURSU.
Общие закономерности и вариабельность параметров биологического разнообразия региональных фаунистических комплексов млекопитающих на рубеже плейстоцена и голоцена
Параметры биологического разнообразия (табл. 3) демонстрируют существенную изменчивость в разных регионах и для разных состояний фаунистических комплексов. На этом фоне общие для большинства комплексов закономерности их изменчивости заключаются в следующем: 1) относительно резкое снижение видового богатства между состоянием комплексов до перехода (плейстоцен) и переходным состоянием (рис. 5А), которое в целом коррелирует с энтропией (H) (рис. 5В); 2) для состояния «голоцен» более характерен высокий индекс доминирования по сравнению с состоянием «плейстоцен» (рис. 5Г); 3) наблюдается четкий тренд снижения организованности (R) от состояния «плейстоцен» к состоянию «голоцен» (рис. 5Б). Таким образом, основные изменения в структуре фаунистических комплексов на границе плейстоцена и голоцена отражены в снижении видового богатства/разнообразия региональных фаун и меры организации.
Таблица 3. Параметры разнообразия региональных комплексов млекопитающих: NSP – видовое богатство, H – энтропия (nit), D – индекс доминирования, R – избыточность/организованность
Table 3. Diversity parameters of regional mammalian assemblages: NSP – species richness, H – entropy (nit), D – dominance index, R – redundancy/organization
| Регион | Состояние комплекса | |||||||||||
| «плейстоцен» | переходное | «голоцен» | ||||||||||
| NSP | H | D | R | NSP | H | D | R | NSP | H | D | R | |
| ATE | 89 | 3.86 | 0.028 | 0.14 | 32 | 2.90 | 0.058 | 0.16 | 15 | 1.94 | 0.17 | 0.29 |
| BORS | 9 | 2.08 | 0.141 | 0.05 | 19 | 2.61 | 0.10 | 0.13 | 36 | 3.00 | 0.058 | 0.16 |
| CEURN | 99 | 3.71 | 0.036 | 0.19 | 75 | 3.87 | 0.020 | 0.12 | 25 | 2.87 | 0.04 | 0.11 |
| CEURS | 98 | 3.82 | 0.032 | 0.17 | 85 | 3.88 | 0.024 | 0.13 | 54 | 3.42 | 0.036 | 0.14 |
| EEURMU | 57 | 3.53 | 0.035 | 0.08 | 36 | 3.79 | 0.035 | 0.09 | 63 | 3.76 | 0.025 | 0.10 |
| EEURSU | 66 | 3.75 | 0.026 | 0.11 | 31 | 2.99 | 0.013 | 0.13 | 68 | 3.87 | 0.019 | 0.08 |
| MDAP | 65 | 3.44 | 0.042 | 0.18 | 28 | 2.98 | 0.033 | 0.11 | 41 | 3.17 | 0.05 | 0.15 |
| MDBLK | 70 | 3.44 | 0.043 | 0.19 | 34 | 3.03 | 0.039 | 0.14 | 35 | 3.16 | 0.032 | 0.11 |
| MDIB | 70 | 3.59 | 0.036 | 0.15 | 28 | 2.83 | 0.056 | 0.15 | 35 | 3.29 | 0.031 | 0.07 |
| PNTCA | 66 | 3.72 | 0.029 | 0.11 | 27 | 3.00 | 0.024 | 0.09 | 18 | 2.66 | 0.081 | 0.08 |
| WEUR | 112 | 3.86 | 0.031 | 0.18 | 107 | 3.88 | 0.029 | 0.17 | 70 | 3.62 | 0.041 | 0.15 |
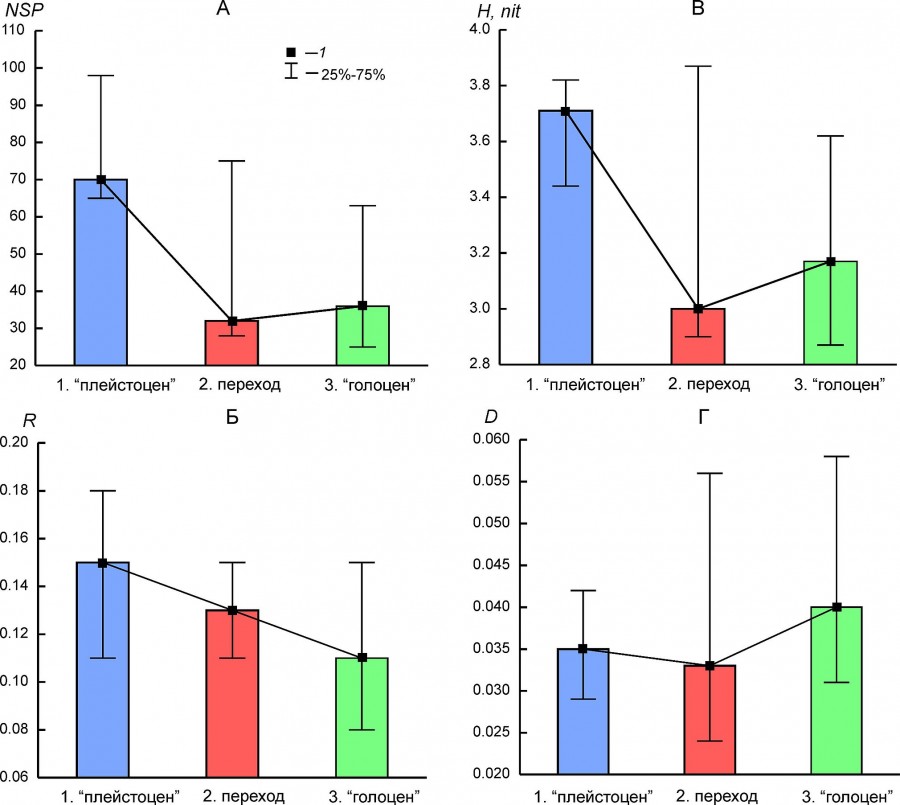
Рис. 5. Медиана (1) и межквартильный интервал (25–75 %) параметров биологического разнообразия разных состояний региональных комплексов млекопитающих (1 – «плейстоцен», 2 – переход, 3 – «голоцен»). A – видовое богатство (NSP), Б – мера организации (R), В – энтропия (H), Г – индекс доминирования (D)
Fig. 5. Median (1) and interquartile range (25%–75%) of the biodiversity parameters for the different states of regional mammalian assemblages (1 – “Pleistocene”, 2 – transition, 1 – “Holocene”). A – species richness (NSP), Б – index of organization (R), В – entropy (H), Г– dominance index (D)
Не во всех региональных комплексах в полной мере проявляются описанные выше временные тренды параметров разнообразия. Видовое богатство региона BORS в голоцене стало в 4 раза выше, чем в плейстоцене. Увеличение разнообразия комплекса сопровождалось ростом параметра R и снижением параметра D (табл. 3). Видовое богатство южно-уральского региона в плейстоцене было не выше такового в раннем и среднем голоцене, но при этом мера организованности и индекс доминирования в голоцене были существенно ниже. Параметры разнообразия среднеуральского комплекса (EEURMU) слабо отличаются для состояний «плейстоцен» и «голоцен». Самое высокое значение R отмечено для комплекса ATE в голоцене при очень низком значении энтропии, низком видовом богатстве и высоком значении индекса доминирования (см. табл. 3).
Индекс «оборота видов» отражает скорость изменения качественного состава фаунистических комплексов между состояниями «плейстоцен» – переход – «голоцен» (табл. 4, рис. 6). В среднем изменения в составе комплексов были несколько больше при их переходе в состояние «голоцен», чем между состоянием «плейстоцен» и переходным состоянием. На уровне медианных значений индекса эти различия не выглядят существенными.
Таблица 4. Индекс «оборота видов» между разными состояниями региональных комплексов млекопитающих
Table 4. “Species turnover" index for different states of regional mammalian assemblages
| Регион | Состояния комплекса | |||
| «плейстоцен» – переходное |
переходное – «голоцен» |
«плейстоцен» – переходное – «голоцен» |
«плейстоцен» – «голоцен» | |
| ATE | 0.488 | 0.447 | 0.882 | 0.712 |
| BORS | 0.643 | 0.418 | 0.961 | 0.733 |
| CEURN | 0.149 | 0.551 | 0.626 | 0.645 |
| CEURS | 0.169 | 0.352 | 0.506 | 0.408 |
| EEURMU | 0.290 | 0.313 | 0.558 | 0.233 |
| EEURSU | 0.381 | 0.394 | 0.691 | 0.239 |
| MDAP | 0.398 | 0.362 | 0.694 | 0.358 |
| MDBLK | 0.346 | 0.333 | 0.318 | 0.486 |
| MDIB | 0.429 | 0.302 | 0.688 | 0.390 |
| PNTCA | 0.419 | 0.778 | ~ 1.0 | 0.690 |
| WEUR | 0.132 | 0.279 | 0.405 | 0.297 |
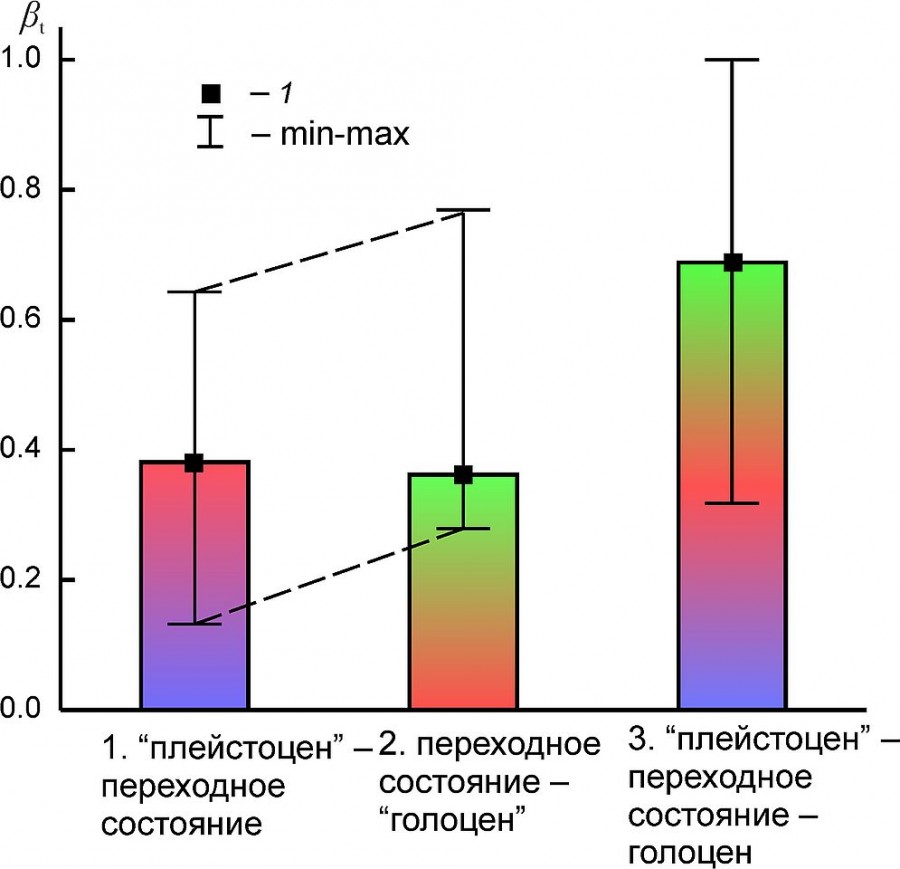
Рис. 6. Медиана (1) и минимальные и максимальные значения (min–max) индекса «оборота видов» (βt) между состояниями региональных комплексов млекопитающих (1 – «плейстоцен» – переход, 2 – переход –«голоцен», 3 – «плейстоцен» – переход – «голоцен»)
Fig. 6. Median (1), the minimum and maximum values (min–max) of the “species turnover” index (βt) between the states of regional mammalian assemblages (1 – “Pleistocene” – transition, 2 – transition – “Holocene”, 3 – “Pleistocene”– transition–“Holocene”)
Видовой состав региональных фаун на рубеже плейстоцена и голоцена изменился в среднем на ~ 68 %. При сравнении состояний комплексов в «плейстоцене» и «голоцене» без учета переходного периода эта величина существенно меньше, ~ 47 %. Относительно слабые изменения состава претерпели комплексы EEURMU, EEURSU, WEUR, MDAP (βt: 0.233– 0.358), а наиболее радикальные – комплексы PNTCA, BORS и ATE (βt: 0.690–0.733) (см. табл. 4).
Абсолютное значение параметра β ранговых снижается в ряду состояний «плейстоцен» – переход – «голоцен» (рис. 7A). В целом эта тенденция изменений формы ранговых распределений хорошо соотносится с трендом снижения организованности структуры фаунистических комплексов и с падением видового богатства.
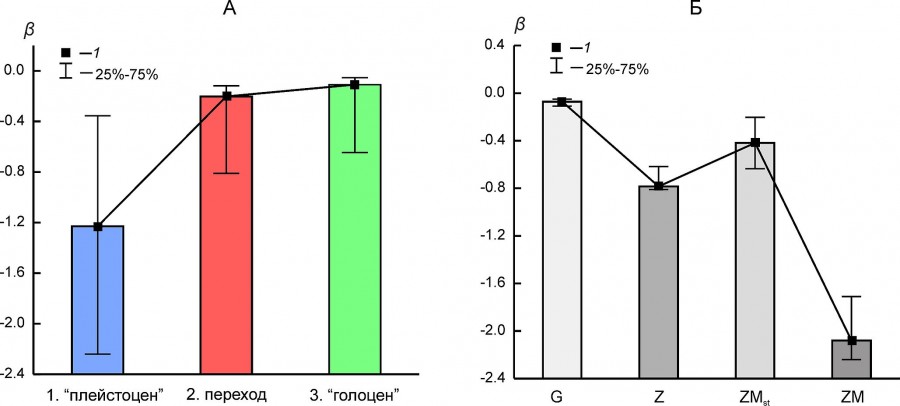
Рис. 7. Медиана (1) и межквартильный интервал (25–75 %) параметра β ранговых распределений для (A) трех состояний региональных комплексов млекопитающих (1 – «плейстоцен», 2 – переход, 3 – «голоцен») и (Б) разных типов ранговых распределений (G, Z, ZM и ZMst)
Fig. 7. Median (1) and interquartile range (25–75 %) of β parameter of rank distributions (А) for the different states of regional mammalian assemblages (1 – “Pleistocene”, 2 – transition, 3 – “Holocene”) and (Б) for different types of rank distribution models (G, Z, ZM, ZMst)
На рисунках 2Г–2Е, 3Г–3Е, 4В и 4Г представлены графики эмпирических ранговых распределений (точки) и их наилучшие аппроксимации (табл. 5) модельными ранговыми распределениями (линии) для разных состояний фаунистических комплексов. Чем больше различия в геометрии распределений, тем четче выражены различия и в структуре самих комплексов на рассматриваемых этапах их эволюции.
Таблица 5. Тип рангового распределения (DSTR) и его параметр β, доля воспроизведенной дисперсии (r2)
Table 5. A type of rank distribution (DSTR) and its parameter β, reproduced variance (r2)
| Регион | Состояние комплекса | ||||||||
| «плейстоцен» | переходное | «голоцен» | |||||||
| DSTR | β | r2 | DSTR | β | r2 | DSTR | β | r2 | |
| ATE | ZM | -1.92 | 0.975 | ZMst | -0.253 | 0.988 | Z | -1.181 | 0.966 |
| BORS | Z | -0.617 | 0.809 | Z | -0.810 | 0.981 | G | -0.110 | 0.986 |
| CEURN | ZM | -2.24 | 0.995 | ZMst | -0.152 | 0.968 | Z | -0.079 | 0.938 |
| CEURS | ZM | -1.43 | 0.988 | ZMst | -0.625 | 0.994 | ZMst | -0.646 | 0.986 |
| EEURMU | G | -0.073 | 0.979 | G | -0.069 | 0.992 | G | -0.044 | 0.979 |
| EEURSU | G | -0.046 | 0.976 | Z | -0.347 | 0.800 | G | -0.046 | 0.957 |
| MDAP | ZMst | -0.355 | 0.996 | G | -0.073 | 0.946 | G | -0.109 | 0.968 |
| MDBLK | ZMst | -1.23 | 0.979 | ZM | -1.19 | 0.978 | G | -0.065 | 0.938 |
| MDIB | ZMst | -0.481 | 0.995 | G | -0.118 | 0.979 | G | -0.054 | 0.942 |
| PNTCA | ZM | -2.25 | 0.983 | G | -0.038 | 0.731 | G | -0.129 | 0.996 |
| WEUR | ZM | -1.71 | 0.972 | ZMst | -0.132 | 0.956 | Z | -0.759 | 0.966 |
Величина параметра β зависит от типа рангового распределения, хотя для одного и того же варианта распределения может варьировать в широких пределах (см. рис. 7Б). Распределения типа ZM или ZMst чаще всего характеризуют фаунистические комплексы в состоянии «плейстоцен» (см. табл. 5). Исключение составили северный комплекс BORS и два уральских комплекса – EEURSU, EEURSU. Параметр β для распределений типа ZM варьировал от -2.25 (PNTCA) до -0.355 (MDAP). В первом случае ранговое распределение полого (см. рис. 2Г), без выраженного доминирования местонахождений с наиболее распространенными видами (кабалоидная лошадь, сайга, первобытный бизон, благородный олень, европейский осел, степная пеструшка и др.) и с коротким «хвостом» из видов, отмеченных единично (росомаха, хомячок Эверсмана и др.). Соответствующее распределение MDAP имеет s-образную форму (см. рис. 4В) и характеризуется относительно четко выраженным доминирование немногих видов (благородный олень, кабан, первобытный бык, косуля, горный козел, заяц-русак и др.) и «хвостом» экзотических или редких для региона видов (узкочерепная полевка, красный волк и др.). Ранговые распределения типа G (см. рис. 4Г) характеризуются плавным снижением частот встречаемости видов и относительно коротким «хвостом» редких таксонов.
На переходной стадии региональные комплексы MDIB, MDAP, PNTC меняют тип рангового распределения с ZM на G. Эта смена отражает, вероятно, упрощение структуры комплексов и экологических отношений видов со средой, снижение общей устойчивости. В регионе EEURSU формально ранговое распределение соответствует модели Z. Для переходного состояния комплекса (оценка по динамике BC2) всего отмечен 31 вид, причем единично – 22 вида, дважды – 8 видов и один (сурок байбак) отмечен три раза (см. рис. 4Г). Таким образом, фактически, учитывая низкое значение r2 (см. табл. 5), это ранговое распределение не может быть описано ни одной из рассмотренных моделей, а состояние системы интерпретируется как сильно нестационарное.
Комплексы СEURS, CEURN, ATE и WEUR перешли из «равновесного» состояние в «стационарное», сопровождающееся снижением абсолютных значений β. Распределение для регионального комплекса MDB формально сохранило тип модели рангового ZM с небольшим уменьшением параметра β. Изменения коснулись таксонов первых рангов (горный козел, благородный олень, кабан), частота местонахождений с которыми существенно возросла (см. рис. 2E).
Переход региональных комплексов из состояния «плейстоцен» в состояние «голоцен» выразился в смене доминирующего типа рангового распределения (см. табл. 5). Преобладании распределения типа G с параметром β от -0.129 до -0.044 в сочетании со снижением меры организованности указывает на упрощение структуры региональных комплексов MDAP, MDIB, MDBLK, PNTCA, наметившееся еще на предыдущей, переходной, стадии. Для региональных комплексов ATE, CEURN и WEUR изменение стационарного варианта ZM распределения на Z не принципиально с точки зрения отношений «вид» – «ресурс», а отражает только снижение видового богатства и/или рост индекса доминирования и увеличение абсолютных величины параметра β. Комплекс центрально-европейского региона CEURS не изменил своей структуры по параметрам рангового распределения.
Выше были приведены сведения об изменении во времени параметров разнообразия региональных комплексов млекопитающих. Эти же данные позволяют проверить гипотезу о существовании пространственного паттерна их изменчивости. Гипотеза предполагает, что какая-то часть дисперсии параметров разнообразия определяется географическим положением регионов. Для проверки гипотезы мы применили метод главных компонент для стандартизованных параметров биологического разнообразия (H, D, R, β, βt) (рис. 8А). Проекция региональных комплексов на PC1 и PC2 и их классификация UPGMA методом по первым трем главным компонентам (метрика – дистанция Эвклида) (рис. 8Б) показала наличие трех географических групп регионов (см. рис. 1): 1) средиземноморская группа (MDIB, MDAP, MDBLK); 2) западно-центрально-европейская группа (WEUR, CEURN, CEURS); 3) уральская группа (EEURMU, EEURSU). Три региональных комплекса (ATE, BORS и PNTCA) демонстрируют своеобразие параметров биологического разнообразия, а их изменчивость отражает кардинальные перестройки региональных фаун млекопитающих на рубеже плейстоцена и голоцена (см. табл. 3–5).
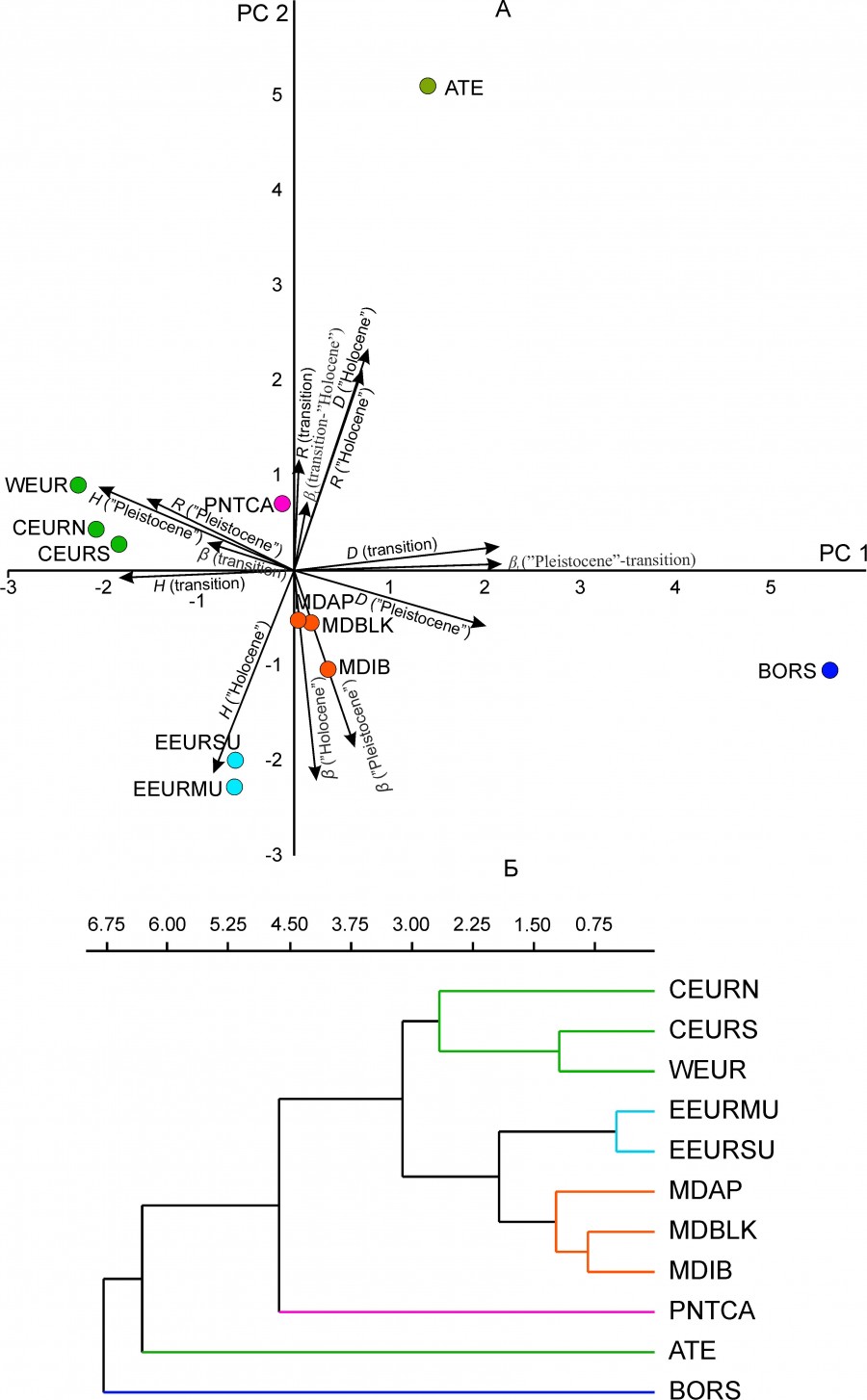
Рис. 8. Относительное положение региональных комплексов млекопитающих. A – в пространстве первых главных компонент (PC1, PC2) показана проекция переменных модели – параметров биологического разнообразия (H, D, R, β, βt ), Б – UPGMA дендрограмма по трем главным компонентам (PC1, PC2, Pc3), кофенетическая корреляция 0.95
Fig. 8. The relative position of regional mammalian assemblages. А – projection on the first principal components (PC1, PC2); the projections of parameters of biological diversity are shown (H, D, R, β, βt ), Б – UPGMA tree based on three principal components (PC1, PC2, PC3); cophenetic correlation 0.95
Обсуждение
Моделирование динамики состава фаун позволило установить, что трансформация региональных комплексов млекопитающих Европы на границе плейстоцена и голоцена происходила асинхронно в разных частях субконтинента в интервале примерно от 18 до 6.6 тыс. лет назад. Продолжительность переходного состояния фаунистических комплексов варьировала от ~1 до 5.6 тыс. лет (в среднем – около 3 тыс. лет). Таким образом, в каждом из регионов, где этот процесс был отчетливо выражен, речь может идти об очень коротких временных интервалах. В масштабе истории этих комплексов в позднем плейстоцене (начиная с конца пятой морской изотопной стадии и до конца максимума последнего оледенения, 74–16 тыс. лет назад) время качественного изменения их состава составило в среднем около 5 % от времени существования.
Эволюция комплексов Апеннинского полуострова (MDAP) и Южного Урала происходила без выраженного переходного периода. В регионе MDAP основная трансформация, которую можно рассматривать в качестве аналога переходного состояния фаунистических комплексов в других регионах, произошла раньше, в максимум последнего оледенения (см. рис. 4А). На Южном Урале фаунистический комплекс не претерпел каких-либо качественных перестроек в конце позднего плейстоцена. Своеобразие эволюции этих комплексов обусловлено прежде всего их географическим положением. На эволюцию комплекса MDAP влиял «островной эффект», выразившийся в обеднении фауны в максимум последнего оледенения (Puzachenko, Markova, 2019) и ограничивший иммиграцию в позднеледниковье и раннем голоцене. Особое положение южноуральского комплекса (EEURSU) на границе европейских и сибирских фаун обеспечило этому региону роль рефугиума для видов с очень разными экологическими нишами. В конце плейстоцена его состав обогатился некоторыми представителями азиатской фауны и лесными видами (колонок, бурундук, белка и т.п.).
Изменения глобального климата выступали в качестве основного драйвера эволюции структуры региональных фаун. Существенно, что повышение температуры в целом было неблагоприятно для позднеплейстоценовых комплексов млекопитающих, что отражено в разных знаках коэффициентов корреляции координаты BC1 с переменными δ18O (+) и NSP (–). Исключением является наиболее северный регион (BORS), ареал которого частично перекрывался скандинавским ледником (см. рис. 1). Поэтому естественно, что потепление было здесь предпосылкой для роста видового разнообразия. В остальных случаях на территории западной, центральной и восточной Европы происходил распад дисгармоничных региональных фаун, ядром которых выступали холодовыносливые и ксерофильные виды открытых пространств, прежде всего травоядных млекопитающих.
Индекс «оборота видов» показывает степень обновления состава фаун за счет двух процессов – вымирания и иммиграции. Сравнение голоценовых и плейстоценовых состояний региональных комплексов показало, что регионы южного и среднего Урала, а также западноевропейский регион могут рассматриваться как рефугиумы плейстоценовой фауны (без учета современной антропогенной трансформации этих территорий). В них «оборот» видов составил не более 30 %. В подавляющем числе регионов отмечено снижение видового богатства, что указывает на преобладание процесса вымирания видов. Исключение составляют только уральские регионы и самый северный из рассмотренных нами регионов – BORS.
Параметры биологического разнообразия (H, D, R) демонстрируют в разной степени чувствительность к изменению эволюционного состояния фаунистических комплексов.
Энтропия (H), в соответствие с зависимостью от числа элементов в «системе» (Foerster, 1960), строго положительно коррелирует с логарифмом числа обнаруженных таксонов (общая зависимость: H = 0.31 + 1.82 · log10(NSP), r = 0.934, p < 0.00001). Эта связь определяет низкие величины разнообразия фаунистических комплексов в голоцене по сравнению с плейстоценом. На уровне устойчивой тенденции отметим отрицательную корреляцию H с индексом доминирования D (к = -0.66, p < 0.0001). Эта зависимость отражает тенденцию к полидоминантности в комплексах с высоким видовым богатством.
Параметр организации (R) имеет важное теоретическое значение. Напомним, что имеется прямое биологическое приложение меры избыточности, например, к линейному генетическому коду, когда одной кодируемой аминокислоте соответствуют от 1 до 6 триплетов нуклеотидов в молекуле ДНК. Такие триплеты по аналогии называются синонимами. Как известно, замены синонимичных триплетов (мутации) не приводят к изменениям в структуре кодируемого белка. Избыточность на уровне элементов, функциональных связей и т. п. является необходимым условием устойчивости на уровне системы, не только в контексте передачи информации. Этот принцип по аналогии можно распространить и на сообщество организмов, в котором, например, совместно обитают множество видов с относительно узкими и перекрывающимися экологическими нишами (функциональная избыточность), которые в совокупности эффективно используют ограниченные ресурсы. Безвозвратная или временная потеря одного или нескольких таких видов не приведет к заметным изменениям в функционировании сообщества в целом с точки зрения использования ресурсов и структуры пищевых цепей за счет расширения или смещения ниш оставшихся видов.
Многочисленные исследования относительного содержания изотопов азота и углерода (d15N, d13С) в костных тканях плейстоценовых млекопитающих показывают высокую устойчивость пищевых ниш представителей мегафауны (пещерный медведь, мамонт, шерстистый носорог, бизон) в позднем плейстоцене и в разных регионах Голарктики (Kuitems et al., 2019; Schwartz-Narbonne et al., 2019 и мн. др.). В частности, в работе (Schwartz-Narbonne et al., 2019) обосновывается гипотеза функциональной экологической избыточности экосистем «мамонтовой степи» (resilient system model). При этом некоторые виды фаунистического комплекса (лошадь, северный олень, сайга) демонстрируют способность менять пищевую нишу в зависимости от условий, свидетельствуя об экологической пластичности (dietary flexibility).
Наши данные демонстрируют относительно высокую организованность фаунистических комплексов млекопитающих в плейстоцене. Отметим, что R не коррелирует ни с NSP, ни с H, т. е. представляет собой независимый параметр разнообразия. Начиная с переходного периода во многих региональных комплексах млекопитающих происходило снижение параметра R. Снижение организованности в голоценовых комплексах предполагает: упрощение экологических отношений, преобладание видов с широкими экологическими нишами, снижение функциональной избыточности и, как следствие, снижение устойчивости. С другой стороны, у таких комплексов довольно высок потенциал восстановления после разрушительного воздействия.
В контексте гипотезы функциональной избыточности интересен феномен уральских регионов, в которых комплексы млекопитающих демонстрируют устойчиво низкие значения параметра R (0.08–0.13). Их устойчивость поддерживалась, вероятно, не столько за счет процессов самоорганизации и взаимодействий между видами, сколько за счет высокой мозаичности и разнообразия местообитаний. Последнее типично для горных экосистем, но в данном случае важно также географическое положение регионов и меридиональное простирание горной системы Урала. Сочетание этих условий обеспечивает обитание на относительно небольшой территории степных, лесостепных и лесных видов как холодовыносливых, так и в разной степени теплолюбивых.
В заключение необходимо отметить, что параметр R играет важную роль в классификации систем и их динамики, независимо от их природы. Так, например, региональные комплексы млекопитающих, для которых R R > ~ 0.11 можно отнести к категории «квазидетерминированных систем» (Крамаренко, 2005). Значения R > 0.31 для естественных (не созданных человеком) сложных систем маловероятны, а более высокие значения параметра, скорее, указывают на присутствие в выборке элементов, относящихся к двум или большему числу разных систем.
Параметрами ранговых распределений частот встречаемости местонахождений с разными видами млекопитающих являются коэффициенты, число которых изменяется от двух до четырех. В данном исследовании мы использовали только один из них – параметр -β. Величина 1/|-β| в «термодинамической интерпретации» сообщества пропорциональна «температуре» системы (Булгаков, Левич, 2005; Пузаченко, 2016). Чем меньше абсолютное значение β, соответственно выше «температура», тем менее выражен вклад в ранговое распределение таксона первого ранга и выше вклад «обычных» и «редких» таксонов (распределение более полого). Величина β зависит от модели распределения. В соответствие с подходом А. П. Левича, в случае линейной зависимости между встречаемостью «вида» и объемом потребляемого им гипотетического «ресурса» (G распределение) β обычно меньше, чем при нелинейной (логарифмической) связи между ними (распределения типа Z и ZM). Можно предположить, что в неустойчивых («горячих») системах с низкими значениями R и со стохастической динамикой смены состояний абсолютное значение β будет ниже, чем в более устойчивых («холодных») системах с более детерминированной динамикой и высокой организованностью. Таким образом, параметр β может содержать обобщенную информацию о доминирующих экологических отношениях в сообществе и применяться как один из «индексов» видового разнообразия. Для большинства региональных комплексов млекопитающих в плейстоцене параметр β был ниже (выше по абсолютному значению), чем в голоцене. Этот эмпирический факт соответствует содержанию гипотезы об их устойчивости и высокой организации. При этом изменчивость β не коррелирует с другими параметрами, т. е. содержит независимую компоненту информации о феномене биологического разнообразия. Ранговые распределения G, Z и ZM характеризуют гипотетическую систему в состоянии равновесия (по условию математического вывода этих моделей при допущении, что система равновесна и все производные равны нулю, а энтропия максимальна (Пузаченко, 2016)). Поэтому в случае хорошего статистического совпадения реального распределения с этими моделями можно предполагать, что в моделируемом сообществе организмов в течение «периода наблюдений» не происходит процессов, изменяющих тип отношений между видами и ресурсами среды. Модификации модели «стационарного» состояния предполагают снижение энтропии системы при производных, не сильно отличных от 0. С этой точки зрения независимо от типа рангового распределения большинство региональных фаунистических комплексов млекопитающих находятся либо в относительно равновесном (чаще), либо в стационарном состоянии на рассматриваемых пространственно-временных масштабах. Масштаб в данной работе определяется размером географического региона и интервалом времени, фиксированным для каждого из трех состояний. Исключение представляют уральские регионы EEURSU и EEURSU, в которых отмечена высокая амплитуда колебаний параметров структуры, отраженная в динамике первой координаты региональных моделей (BC1, см. рис. 4E). Эти обратимые колебания происходили вокруг довольно широкой области значений BC1, без выраженного тренда.
Мы показали присутствие географического паттерна во временной изменчивости параметров биологического разнообразия региональных фаунистических комплексов второй половины позднего плейстоцена и первой половины голоцена. Сами биорегионы были выделены в результате анализа современных ареалов млекопитающих, конфигурация которых для видов, переживших границу плейстоцена и голоцена, существенно изменилась по сравнению с прошлым. Факт географического упорядочивания регионов на основе параметров биологического разнообразия ископаемых фаунистических комплексов косвенно подтверждает нашу гипотезу об относительной инвариантности их границ в голоцене – позднем плейстоцене Европы (Маркова и др., 2008).
Завершая обсуждение результатов, остановимся на некоторых ограничениях, связанных с использованием параметров биологического разнообразия. В основном они состоят в зависимости результата от выбранного пространственно-временного масштаба исследования. Зависимость параметров разнообразия (например, видового богатства и энтропии) от объема выборки или обследованной площади – известная эмпирическая закономерность, имеющая хорошие теоретические обоснования. Мы предполагаем, что аналогичная по природе зависимость будет наблюдаться при анализе палеонтологического материала. По крайней мере, на основе данных, лежащих в этой работе, можно полагать, что существует нелинейная корреляция между длиной временного интервала и числом обнаруженных видов с выходом соответствующей кривой на «плато». Эти ограничения ставят проблему «репрезентативности» исходных данных при оценке параметров разнообразия. Также изменение пространственного масштаба исследования может существенно сказаться на параметрах модели динамики структуры исследуемой «системы» (число координат, смена ведущих управляющих внешних переменных). Этот эффект обусловлен иерархической организацией природных систем (Пузаченко, 2017). Например, из того, что на уровне «большого региона» доказана ведущая роль динамики глобального климата как фактора, ограничивающего изменчивость видового богатства, не следует, что эта роль останется неизменной для отдельных внутрирегиональных выделов. Аналогичные ограничения распространяются и на временной масштаб.
Заключение
Трансформация структуры фаунистических комплексов млекопитающих на рубеже плейстоцена и голоцена происходила в разное время в разных регионах Европы в интервале 18–6.6 тыс. лет назад и не совпадала со стратиграфической границей между плейстоценом и голоценом (11.7 тыс. лет назад).
Длительность переходного состояния фаунистических комплексов варьировала от ~1 до 5.6 тыс. лет (в среднем – около 3 тыс. лет).
Выделено три варианта эволюции фаунистических комплексов: 1) с резкой перестройкой фауны в относительно короткие сроки, 2) с постепенной растянутой во времени направленной трансформацией и 3) без существенных изменений в структуре комплексов.
Ведущими внешними факторами варьирования видового богатства и структуры фауны млекопитающих были изменения глобального климата.
Основные изменения структуры фаунистических комплексов выражались в снижении видового богатства/разнообразия и упрощении внутренней структуры.
Функциональная избыточность большинства европейских фаунистических комплексов в голоцене ниже, чем в плейстоцене, что предполагает их относительно низкую устойчивость к воздействию внешних факторов, но и относительную высокую способность к самовосстановлению.
Устойчивость фаунистических комплексов Среднего и Южного Урала поддерживается не столько за счет процессов самоорганизации на экосистемном уровне, сколько за счет высокой мозаичности и разнообразия горных местообитаний.
Параметры биологического разнообразия, использованные в работе в совокупности с многомерными моделями динамики и моделями ранговых распределений впервые позволили провести содержательное макроскопическое описание паттернов эволюционной перестройки региональных фаун Европы на рубеже плейстоцена и голоцена и их сравнительный анализ.
Библиография
Бир С. Кибернетика и управление производством . М.: Физматгиз, 1963. 275 с.
Булгаков Н. Г., Левич А. П. Описание, происхождение и применение ранговых распределений в экологии сообществ // Вестник Московского университета. 2005. Сер. 16. Биология. № 1. С. 18–24.
Данукалова Г. А., Юрин В. И., Косинцев П. А., Осипова Е. М., Курманов Р. Г. Биостратиграфические исследования отложений верхнего плейстоцена и голоцена пещеры Сикияз-Тамак 7 (Южный Урал, Россия) // Геологический вестник. 2018. № 1. С. 144–161.
Крамаренко С. С. Метод использования энтропийно-информационного анализа для количественных признаков // Известия Самарского научного центра РАН. 2005. Т. 7 (1). С. 242–247.
Левич А. П. Информация как структура систем // Семиотика и информатика. 1978. Т. 10. С. 116–132.
Левич А. П. Структура экологических сообществ . М.: Изд-во Моск. ун-та, 1980. 180 с.
Левич А. П. Описание, происхождение и применение ранговых распределений в экологии сообществ // Общая и прикладная ценология. 2007. № 5. С. 14–19.
Мандельброт Б. Теория информации и психологическая теория частот слов // Математические методы в социальных науках. М.: Прогресс, 1973. С. 326–337.
Маркова А. К., Кольфсхотен Т. ван, Бохнкке Ш., Косинцев П. А., Мол И., Пузаченко А. Ю., Симакова А. Н., Смирнов Н. Г., Верпоорте А., Головачев И. Б. Эволюция экосистем Европы при переходе от плейстоцена к голоцену . (24–8 тыс. л. н.) / Отв. ред. А. К. Маркова, Т. ван Кольфсхотен. М.: Товарищество научных изданий КМК, 2008. 556 с.
Маркова А. К., Пузаченко А. Ю. Реконструкция изменений палеосреды в районе Костенок и прилегающих территорий в течение позднего плейстоцена по данным млекопитающих // Естественно-научные методы в изучении и сохранении памятников Костенковского-Борщевского археологического района. Воронеж: Воронежcкий университет, 2017. С. 48–57.
Пузаченко Ю. Г. Термодинамическая основа учения о биосфере и ноосфере В. И. Вернадского (к 150-летию академика В. И. Вернадского) // Известия РАН. Сер. географическая. 2013. № 4. С. 5–20.
Пузаченко Ю. Г. Ранговые распределения в экологии и неэкстенсивная термодинамика // Аспекты биоразнообразия. Сборник трудов Зоологического музея МГУ. Т. 54 (2). М.: Товарищество научных изданий КМК, 2016. С. 401–830.
Пузаченко Ю. Г. Биологическое разнообразие в биосфере: системологический и семантический анализ // Биосфера. 2009. Т. 1 (1). С. 025–038.
Пузаченко Ю. Г. Теоретико-методологические основы долговременных эколого-географических исследований на территории заповедников // Вопросы географии. 2017. Т. 143. С. 192–233.
Смирнов Н. Г., Большаков В. Н., Косинцев П. А., Панова Н. К., Коробейников Ю. И., Ольшванг В. Н., Ерохин Н. Г., Быкова Г. В. Историческая экология животных гор Южного Урала . Свердловск: УрО АНСССР, 1990. 244 с.
Álvarez-Lao D. J., Méndez M. Latitudinal gradients and indicator species in ungulate paleoassemblages during the MIS 3 in W Europe // Palaeogeography, Palaeoclimatology, Palaeoecology. 2016. № 449. P. 455–462.
Andersen K. K., Azuma N., Barnola J. M., Bigler M., Biscaye P., Caillon N., Chappellaz J., Clausen H. B., Dahl-Jensen D., Fischer H., Fluckiger J., Fritzsche D., Fujii Y., Goto-Azuma K., Gronvold K., Gundestrup N. S., Hansson K.-M., Huber C., Hvidberg C. S., Johnsen S. J., Jonsell U., Jouzel J., Kipfstuhl S., Landais A., Leuenberger M., Lorrain R., Masson-Delmotte V., Miller H., Motoyama H., Narita H., Popp T., Rasmussen S. O., Raynaud D., Rothlisberger R., Ruth U., Samyn D., Schwander J., Shoji H., Siggard-Andersen M.-L., Steffensen J. P., Stocker T., Sveinbjornsdottir A. E., Svensson A., Takata M., Tison J.-L., Thorsteinsson T., Watanabe O., Wilhelms F. & White J. W. C. High-resolution record of Northern Hemisphere climate extending into the last interglacial period // Nature. 2004. Vol. 431 (7005). P. 147–151.
Bachura O., Kosintsev P. Late Pleistocene and Holocene small- and large-mammal faunas from the Northern Urals // Quaternary International. 2007. Vol. 160. P. 121–128.
Bazzaz F. A. Plant Species Diversity in Old-Field Successional Ecosystems in Southern Illinois // Ecology. 1975. Vol. 56 (2). (Early Spring, 1975). P. 485–488.
Bray J. R., Curtis J. T. An ordination of the upland forest communities of southern Wisconsin // Ecological Monographs. 1957. Vol. 27. P. 325–349.
Crees J. J., Carbone C., Sommer R. S., Benecke N., Turvey S. T. Millennial-scale faunal record reveals differential resilience of European large mammals to human impacts across the Holocene // Proceedings of the Royal Society of London B. 2016. Vol. 283. DOI: doi.org/10.1098/rspb.2015.2152
Davison M. L. Introduction to Multidimensional Scaling and Its Applications // Applied Psychological Measurement. 1983. Vol. 7. P. 373–379.
Dinnis R., Pate A., Reynolds N. Mid-to-late Marine Isotope Stage 3 mammal faunas of Britain: a new look // Proceedings of the Geologists’ Association. 2016. Vol. 127. P. 435–444.
Discamps E., Royer A. Reconstructing palaeoenvironmental conditions faced by Mousterian hunters during MIS 5 to 3 in southwestern France: A multi-scale approach using data from large and small mammal communities // Quaternary International. 2017. Vol. 433. P. 64–87.
Fernández-García M., López-García J. M., Lorenzo C. Palaeoecological implications of rodents as proxies for the Late Pleistocene-Holocene environmental and climatic changes in northeastern Iberia // Comptes Rendus – Palevol 2016. Vol. 15. P. 707–719.
Foerster von H. On self-organizing systems and their environments // The Interdisciplinary Symposium on Self-Organizing Systems, May 5, 1959, Chicago, Illinois / Yovits M. C., Cameron S. (eds.). Self-Organizing Systems. London: Pergamon Press, 1960. P. 31–50.
Hammer Ø., Harper D. A. T., Ryan P. D. PAST: paleontological statistics software package for education and data analysis // Palaeontologia Electronica. 2001. Vol. 4. P. 1–9.
Heikinheimo H., Fortelius M., Eronen J., Mannila H. Biogeography of European land mammals shows environmentally distinct and spatially coherent clusters // Journal Biogeography. 2007. Vol. 34. P. 1053–1064.
Hubbell S. P. The unified neutral theory of biodiversity and biogeography. Princeton: Princeton Univ. Press, 2001. 375 p.
Hutchinson G. E. Concluding remarks // Cold Spring Harbor Symposia on Quantitative Biology. 1957. Vol. 22 (2). P. 415–427.
Kosintsev P. A., Gasilin V. V., Gimranov D. O., Bachura O. P. Carnivores (Mammalia, Carnivora) of the Urals in the Late Pleistocene and Holocene // Quaternary International 420. 2016. P. 145–155.
Kreft H., Jetz W. A framework for delineating biogeographical regions based on species distributions // Journal Biogeography. 2010. Vol. 37. P. 2029–2053.
Kuitems M., Kolfschotena T. van, Tikhonovd A. N., Plichta J. van der. Woolly mammoth δ13C and δ15N values remained amazingly stable throughout the last ∼50,000 years in north-eastern Siberia // Quaternary International. 2019. Vol. 500. P. 120–127.
Magurran A. E. Measuring biological diversity. Oxford: Blackwell Publishing, 2004. 256 p.
Margalef R. Information theory in ecology // Memorias de la Real Academia de Ciencias y Artes de Barcelona. 1957. Vol. 23. P. 373–449.
Markova A. K., Puzachenko A. Y., Kolfschotena T. van. The North Eurasian mammal assemblages during the end of MIS 3 (Brianskian-Late Karginian-Denekamp Interstadial) // Quaternary International 2010. Vol. 212. P. 149–158.
Markova A. K., Smirnov N. G., Kozharinov A. V., Kazantseva N. E., Simakova A. N., Kitaev L. M. Late Pleistocene distribution and diversity of mammals in Northern Eurasia (PALEOFAUNA database) // Paleontologia i Evolucio. 1995. Vol. 28/29. P. 5–143.
Markova A., Puzachenko A. Preliminary Analysis of European Small Mammal Faunas of the Eemian Interglacial: Species Composition and Species Diversity at a Regional Scale // Quaternary. 2018. Vol. 1 (9). DOI: doi:10.3390/quat1020009
Marquet P. A. Invariants, scaling laws, and ecological complexity // Science. 2000. Vol. 289. № 5484. P. 1487–1488.
Marquet P. A. Scaling and power-laws in ecological systems // Journal of Experimental Biology. 2005. Vol. 208. P. 1749–1769.
McGill B. J., Etienne R. S., Gray J. S., Alonso D., Anderson M. J., Benecha H. K., Dornelas M., Enquist B. J., Green J. L., He F., Hurlbert A. H., Magurran A. E., Marquet P. A., Maurer B. A., Ostling A. S., Candan U. U., Ugland K. I., White E. P. Species abundance distributions: moving be yond single prediction theories to integration within an ecological framework // Ecology Letters. 2007. Vol. 10. P. 995–1015.
Mourelle C., Ezcurra E. Differentiation diversity of Argentine cacti and its relationship to environmental factors // Journal of Vegetation Science. 1997. Vol. 8. P. 547–558.
Pavelková Řičánková V., Robovský J., Riegert J., Zrzavý J. Regional patterns of postglacial changes in the Palearctic mammalian diversity indicate retreat to Siberian steppes rather than extinction // Scientific Reports. 2015. Vol. 5. DOI: doi.org/10.1038/srep12682
Puzachenko A. Yu., Markova A. K. Diversity dynamics of large- and medium-sized mammals in the late Pleistocene and the Holocene on the east European Plain: systems approach // Quaternary International. 2016. Vol. 420. P. 391–401.
Puzachenko A. Yu., Markova A. K. Evolution of mammal species composition and species richness during the Late Pleistocene – Holocene transition in Europe: A general view at the regional scale // Quaternary International 2019. URL: https://doi.org/10.1016/j.quaint.2018.12.025 (дата обращения: 10.08.2019).
Puzachenko A. Yu., Markova A. K. Mammal diversity during the Pleistocene–Holocene transition in Eastern Europe // Integrative Zoology. 2014. Vol. 9. P. 461–470.
Ramsey B. C., Lee Sh. Recent and planned developments of the program OxCal // Radiocarbon. 2013. Vol. 55. P. 720–730.
Reimer P. J., Bard E., Bayliss A., Beck J. W., Blackwell P. G., Ramsey C. B., Buck C. E., Cheng Edward H. L. R., Friedrich M., Grootes P. M., Guilderson T. P., Haflidason H., Hajdas I., Hatté C., Heaton T. J., Hoffmann D. L., Hogg A. G., Hughen K. A., Kaiser K. F., Kromer B., Manning S. W., Niu M., Reime, R. W., Richards D. A., Scott E. M., Southon J. R., Staff R. A., Turney C. S. M., Plicht J. van der. Intcal13 and Marine13 radiocarbon age calibration curves 0–50,000 years cal BP // Radiocarbon. 2013. Vol. 55. P. 1869–1887.
Ricotta C., Podani J. On some properties of the Bray-Curtis dissimilarity and their ecological meaning // Ecological Complexity. 2017. Vol. 31. P. 201–205.
Royer R. R., Montuire S., Legendre S., Discamps E., Jeannet M., Lécuyer Ch. Investigating the influence of climate changes on rodent communities at a regional-scale (MIS 1–3, Southwestern France) // PLoS ONE. 2016. Vol. 11 (1). DOI: doi.org/10.1371/journal.pone.0145600
Schwartz-Narbonne R., Longstaffe F. J., Kardynal K. J. Druckenmiller P., Hobson K. A., Jass C. N., Metcalfe J. Z., Zazula G.. Reframing the mammoth steppe: Insights from analysis of isotopic niches // Quaternary Science Reviews. 2019. Vol. 215. P. 1–21.
Semken H. A. Jr. Environmental interpretations of the “disharmonious” Late Wisconsinian biome of South-eastern North America // Laub R. S., Miller N. G., Steadman D. W. (eds). Late Pleistocene and Early Holocene Paleoecology and Archeology of the Eastern Great Lakes Region // Bulletin of Buffalo Society of Natural Sciences. 1988. Vol. 33. P. 185–194.
Semken H. A. Jr., Graham R. W., Stafford T. W. Jr. AMS 14C analysis of Late Pleistocene non-analog faunal components from 21 cave deposits in southeastern North America // Quaternary International. 2010. Vol. 217. P. 240–255.
Shannon C. E. A mathematical theory of communication // Bell System Technical Journal. 1948. Vol. 27. P. 379–423, 623–656.
Simpson E. H. Measurement of diversity // Nature. 1949. Vol. 163. P. 688–690.
Sokal R. R., Rohlf F. J. Biometry: The principles and practice of statistics in biological research. 3rd Edition. NY: W.H. Freeman and Co., 1995. 887 p.
Svendsen J. I., Alexanderson H, Astakhov V. I., Demidov I., Dowdeswell J. A., Svend F., Gataullin V., Henriksen M., Christian H., Houmark-Nielsen M., Hubberten H.W., Ingólfsson Ó., Jakobsson M., Kjær K. H., Larsen E., Lokrantz H., Lunkka J. P., Lyså A., Mangerud J., Matiouchkov A., Murray A., Möller P., Niessen F., Nikolskaya O., Polyak L., Saarnisto M., Siegert Ch., Siegert M.J., Spielhagen R. F., Stein R. Late Quaternary ice sheet history of northern Eurasia // Quaternary Science Reviews. 2004. Vol. 23 (11–13). P. 1229–1271.
Tokeshi M. Species abundance patterns and community structure // Advances in Ecological Research. 1993. Vol. 24. P. 111–187.
Torre D., Abbazzi L., Ficcarelli G., Masini F., Mezzabotta C., Rook L. Changes in mammal assemblages during the Lateglacial-eariiest Holocene // Il Quaternario: Italian Journal of Quaternary Sciences. 1996. Vol. 9. № 2. P. 551–560.
Walker M., Johnsen S., Rasmussen S. O., Popp T., Steffensen J.-P., Gibbard P., Hoek W., Lowe J., Andrews J., Bjӧrck S., Cwynar L. C., Hughen K., Kershaw P., Kromer B., Litt T., Lowe D. J., Nakagawa T., Newnham R., Schwander, J. Formal definition and dating of the GSSP (Global Stratotype Section and Point) for the base of the Holocene using the Greenland NGRIP ice core, and selected auxiliary records // Journal of Quaternary Science. 2009. Vol. 24. P. 3–17.
Благодарности
Исследование выполнено по программе Государственного задания Института географии РАН «Оценка физико-географических, гидрологических и биотических изменений окружающей среды и их последствий для создания основ устойчивого природопользования» № 0148-2019-0007 (АААА-А19-119021990093-8).

 © 2011 - 2026
© 2011 - 2026