



Введение
Онежское озеро – это крупнейший после Ладожского озера пресноводный водоем Европы и европейского северо-запада России. Его экосистема испытывает влияние климатических факторов и антропогенной нагрузки. Основными источниками точечного загрязнения являются промцентры – Петрозаводск, Кондопога, Медвежьегорск. В целом на большей части акватории планктонные системы находятся в естественном состоянии, имеют олиготрофный статус. Онежское озеро отличается высоким качеством воды и малой продуктивностью (Онежское озеро…, 2010). В последние десятилетия на территории Карелии наблюдаются климатические колебания, повышение среднегодовой температуры воздуха и количества осадков (Назарова, 2015). В результате этого увеличиваются сроки безледоставного периода и продолжительности периода «биологического лета» (Филатов и др., 2014). В некоторых районах озера отмечается увеличение содержания в воде органического вещества, взвешенных веществ, цветности, общего фосфора, железа (Калинкина и др., 2019). Подобные изменения вызывают реакцию как всего планктона в целом, так и отдельных популяций. Жизненные циклы веслоногих копепод синхронизированы с закономерными сезонными колебаниями в окружающей среде (Nilssen, Wærvågen, 2000). Например, в озере Мюггельзе в Берлине в связи с потеплением климата для рачков Thermocyclops oithonoides и Mesocyclops leuckarti отмечено увеличение продолжительности активной планктонной фазы (сдвиг на более ранние сроки весной и на более поздние осенью), а также увеличение годового пика численности (Gerten, Adrian, 2002).
Вид Thermocyclops oithonoides (Sars, 1863) относится к типу членистоногие (Arthropoda), классу ракообразные (Crustacea), подклассу Copepoda, отряду Cyclopiformes, семейству Cyclopidae (подсемейство Cyclopinae), роду Thermocyclops Kiefer, 1927 (Определитель…, 2010).
T. oithonoides – палеарктический, эвриотопный вид, относится к холодноводному комплексу умеренных широт 50–60º с. ш. (Пидгайко, 1984; Куликова, 2017). Рачок расселился из Понто-Каспийского региона после последнего ледникового периода (Nilssen, Wærvågen, 2000).
Оптимальные условия рачок находит при высоких содержаниях растворенного кислорода. Он обитает при рН = 6.4–7.6. В дистрофных водоемах отсутствует, обитая главным образом в эвтрофных (мезотрофных) и олиготрофных озерах (Рылов, 1948). T. oithonoides относится к видам со средней толерантностью к нарушению ионного состава воды (Калинкина, Куликова, 2009). Вид отмечен в Норвегии (Nilssen, Wærvågen, 2000), Эстонии (Haberman, Virro, 2004), Германии (Adrian et al., 2006), Финляндии (Lehtovaara et al., 2014), в Псковско-Чудском озере на границе между Эстонией и Россией (Хаберман и др., 2012). Массовый вид в Ладожском озере (Родионова, 2013). T. oithonoides встречается в озерах Воже и Лача (Смирнова, 1978), в Куйбышевском (Тимохина, 2000), Горьковском (Гусаков, 2005) и Рыбинском водохранилищах (Лазарева, 2010), в озере Неро (Лазарева, Смирнова, 2008) и др. Самый характерный компонент карельских озер, по данным Т. П. Куликовой (2004, 2007, 2010, 2012), вид отмечен в 409 водоемах из 556 исследованных. Рачок распространен повсеместно от литорали до пелагиали.
T. oithonoides является важным элементом трофической цепи. Это факультативный хищник. Питается простейшими и молодью ракообразных. Науплиальные стадии используют в пищу водоросли, простейших и детрит (Монаков, 1976). Рачок обеспечивает рыбопродуктивность в озерах тайги и подтайги, составляет основу питания ряпушки и корюшки (Пидгайко, 1984). В уловах Онежского озера рыбы-планктофаги составляют 50–85 % (Онежское озеро…, 2010).
Цель работы – изучение жизненного цикла веслоногого рачка T. oithonoides в пелагиали различных районов Онежского озера.
Материалы
В основу работы положен материал комплексных съемок лаборатории гидробиологии ИВПС КарНЦ РАН с 1988 по 2017 г. в период вегетации (июнь – октябрь) (Сярки, Куликова, 2012; Сярки и др., 2015). Пробы зоопланктона центральной части озера были отобраны в пелагиали районов, сходных по термическим и трофическим свойствам (всего 5 станций, 826 проб). Зоопланктон заливов изучался на примере Петрозаводской губы (2 станции, 641 проба) (рис. 1).
Зимнее состояние описано по материалам, полученным в рамках российско-швейцарского мультидисциплинарного проекта «Lake Ladoga: life under ice interplay of under-ice processes by global change» в марте 2015–2017 гг. (в одни и те же даты) на станции в центральной части Петрозаводской губы.
Рис. 1. Схема расположения станций на Онежском озере: 1 – станции в Петрозаводской губе; 2 – станции в центральном районе озера
Fig. 1. Location of stations on Lake Onega: 1 – stations in Petrozavodsk Bay; 2 – stations in the central part
Термический режим в центральной части озера и заливе различается из-за высокой пространственной гетерогенности озера. Так, термобар в Петрозаводской губе проходит в третьей декаде мая, а в центральном районе – в середине июня (рис. 2). Сезонная динамика температуры Петрозаводской губы имеет свои особенности, связанные с активной гидродинамикой залива.

Рис. 2. Средняя температура воды (скользящие средние): 1 – Петрозаводская губа; 2 – центральный район
Fig. 2. Average water temperature (variable averages): 1 – Petrozavodsk Bay; 2 – central part
Центральный район озера в настоящее время имеет олиготрофный статус планктонной системы, что подтверждается низким уровнем общего фосфора (10 мкг/л) и Chl a (2.7 мкг/л).
Химический состав вод Петрозаводской губы формируется в основном за счет речного стока (95 % – река Шуя), обогащенного биогенами, железом и гумусом. Зимой и весной водные массы Петрозаводской губы представлены на 52–62 % шуйскими водами. Особенно велико влияние речных вод на залив в период прохождения термобара и после него, когда водообмен с центральным плесом озера ограничен, формируются неблагоприятные для зоопланктона гидрохимические условия. После прохождения термобара показатели химического состава воды в заливе близки к озерным. Летом и осенью доля озерных вод достигает 82–86 %. Петрозаводская губа испытывает значительную антропогенную нагрузку (промышленно-бытовые стоки, дренажные, ливневые воды города, водный транспорт). Концентрация общего фосфора составляет 18 мкг/л, содержание Chl a 3.3 мкг/л, по этим показателям залив имеет мезотрофный статус (Крупнейшие…, 2015; Лозовик и др., 2019).
Методы
Пробы зоопланктона отбирали планктонной сетью Джеди (с диаметром отверстий 100 мкм) фракционно по слоям (0–5, 5–10, 10–25, 25–50, 50–75, 75–100 м) и фиксировали 4 % формалином. Сбор и обработку материала проводили по стандартной методике (Методические рекомендации…, 1984).
Численность T. oithonoides просчитывали по размерно-возрастным группам: младшие копеподиты (I–III стадии), старшие копеподиты (IV–V стадии), самцы, самки, самки с яйцами. Науплии не идентифицировались до вида и не учитывались.
При вычислении биомассы зоопланктона использовались размеры и индивидуальные веса, рассчитанные для Онежского озера (Куликова, Сярки, 1994).
Для сопоставимости данных из районов с различными глубинами анализировались величины численности в столбе воды под квадратным метром.
Плавные кривые сезонной динамики показателей получены сглаживанием временных рядов (с 1988 по 2017 г.) методом скользящего среднего с шагом в 7 элементов. Для этого ряды данных ранжировались по показателю сезонности (сутки с начала года). Ввиду нерегулярности рядов применялся метод скользящего среднего в модификации двойного сглаживания (Сярки, 2013).
Результаты
T. oithonoides в планктоне встречается круглогодично. Зимой отмечены только копеподиты IV–V стадии, которые зимуют в придонном слое. В подледный период численность рачка составляет 673.5 ± 125.7 экз./м2 (25.3 ± 4.8 экз./м3), биомасса 7.61 ± 1.42 мг/м2 (0.286 ± 0.055 мг/м3). Доля T. oithonoides в планктоне по численности в среднем 3.5 %, по биомассе – 1.5 %.
В первой декаде июня в центральном районе озера наблюдается численность рачка в среднем 4.2 тыс. экз./м2 (0.06 тыс. экз./м3), биомасса 0.05 г/м2 (0.001 г/м3). Основную роль в популяции в это время играют старшие копеподиты, доля которых составляет около 90 %. В Петрозаводской губе отмечены близкие значения численности 4.3 тыс. экз./м2 (0.16 тыс. экз./м3) и биомассы – 0.07 г/м2 (0.003 г/м3), в первую очередь за счет взрослых рачков и младших копеподитных стадий (около 40 % каждая возрастная группа) (рис. 3, 4).
В конце июня – начале июля в центре озера увеличивается количество самцов и самок, и в первой декаде июля отмечается их максимальное количество по численности – 5.8 тыс. экз./м2 (0.08 тыс. экз./м3), по биомассе – 0.09 г/м2 (0.001 г/м3). В первой декаде августа наблюдается второй максимум взрослых рачков, который почти в два раза больше первого (численность 10.6 тыс. экз./м2, или 0.26 тыс. экз./м2, биомасса 0.15 г/м2, или 0.004 г/м3). Для младших и старших копеподитов волны генераций тоже хорошо выражены.
В Петрозаводской губе активное развитие популяции T. oithonoides начинается с первой декады июля. Подобная задержка летнего развития в заливе была отмечена ранее для веслоного рачка Eudiaptomus gracilis (Фомина, Сярки, 2018). Задержка в развитии в июне здесь может быть связана с неблагоприятными гидрохимическими условиями в период термобара и после его прохождения. Активное развитие популяции в начале июля может быть связано с гидродинамикой вод в заливе и с переносом рачков течениями из центрального плеса озера. Волны генерации слабо выражены для старших копеподитов, для младших копеподитов и взрослых рачков сливаются.
Максимум развития популяции T. oithonoides в центральном районе и Петрозаводском заливе приходится на начало августа. В центре озера численность рачка может достигать 46.4 тыс. экз./м2 (1.31 тыс. экз./м3), биомасса – 0.39 г/м2 (0.009 г/м3), в популяции T. oithonoides в Петрозаводской губе отмечены близкие значения – 35.6 тыс. экз./м2 (1.36 тыс. экз./м3) и 0.34 г/м2 (0.013 г/м3) соответственно.
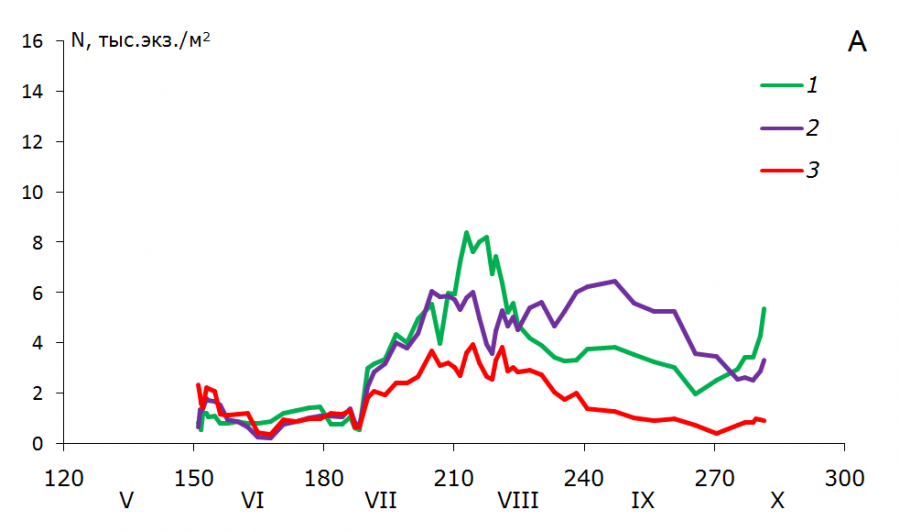
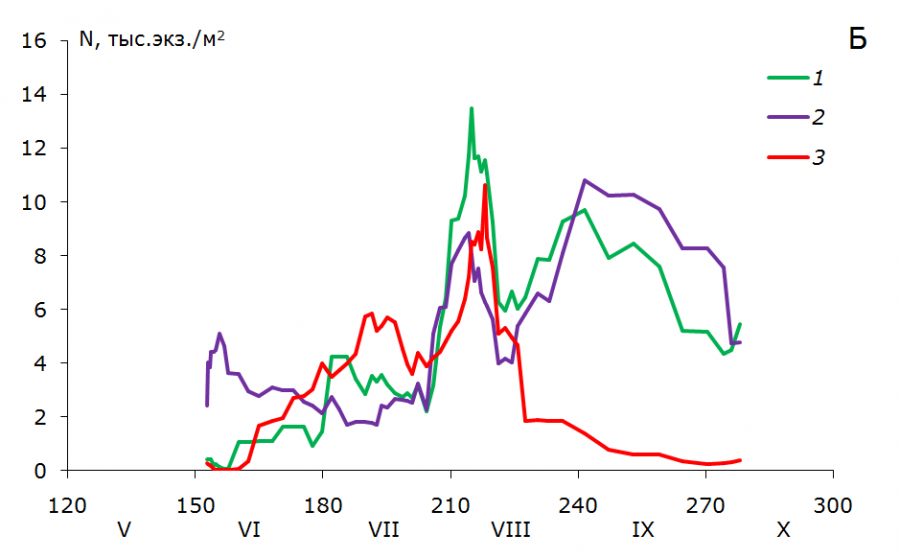
Рис. 3. Динамика численности (N, тыс. экз./м2) возрастных групп T. oithonoides в Петрозаводской губе (А) и центральном районе (Б): 1 – взрослые рачки; 2 – старшие копеподиты; 3 – младшие копеподиты
Fig. 3. Abundance dynamics (N, th. ind./m2) of Thermocyclops oithonoides age groups in Petrozavodskaya Bay (А) and in the central part of Lake Onega (Б): 1 – adults; 2 – senior copepodites; 3 – young copepodites
Осенью доля взрослых рачков снижается, доля копеподитов, наоборот, увеличивается (рис. 4). Старшие копеподиты уходят в диапаузу и мигрируют в придонный слой. В это время в структуре популяции рачка в Петрозаводской губе доля младших копеподитов больше, чем в центральном районе озера. Возможно, это связано с более благоприятными термическими условиями в заливе, когда рачок в теплые годы может давать не две генерации, как в центре озера, а три. По данным В. М. Рылова (1948), T. oithonoides дицикличен, в некоторых озерах он моноцикличен. В карельских озерах вид может быть полицикличен (Филимонова, 1965). Nilssen и Wærvågen (2000) отмечают, что в озерах Норвегии в жизненном цикле T. oithonoides может быть две или три генерации в год в зависимости от ежегодных колебаний температуры воды.


Рис. 4. Структура популяции (%) T. oithonoides в Петрозаводской губе (А) и центральном районе (Б): 1 – взрослые рачки; 2 – старшие копеподиты; 3 – младшие копеподиты
Fig. 4. Population structure (%) of Thermocyclops oithonoides in Petrozavodskaya Bays (А) and in the central part o Lake Onegoa (Б): 1 – adults; 2 – senior copepodites; 3 – young copepodites
Петрозаводский залив прогревается примерно на месяц раньше, чем центральный район, поскольку скорость развития T. oithonoides зависит от температуры, размножение рачка в губе начинается уже в мае. Доля яйценосных самок в заливе в начале июня и до конца августа составляет около 50 % от общего количества самок. В центре озера в начале июня доля яйценосных самок составляет в среднем 5 %, но в июле их число резко возрастает. Короткий вегетационный период вызывает синхронизацию размножения самок (рис. 5).

Рис. 5. Доля яйценосных самок T. oithonoides: 1 – Петрозаводская губа; 2 – Центральный и глубоководный районы
Fig. 5. Proportion of oviparous females of Thermocyclops oithonoides: 1 – Petrozavodskaya Bays; 2 – the central parts of Lake Onega
Значение T. oithonoides в зоопланктоне меняется в течение вегетационного периода. Наименьшую роль в сообществе Петрозаводской губы и в центральной части озера рачок играет в конце июня – начале июля. Наибольшее значение T. oithonoides в заливе наблюдается в первой декаде августа и в начале октября (по численности до 24 %, по биомассе до 8 %), в центральном районе – в начале июня и в начале сентября (по численности до 26 %, по биомассе до 11 %). В связи с небольшими размерами рачка (средний вес взрослых рачков 0.015 мкг) по биомассе он играет меньшую роль.
Обсуждение
Полученные результаты показали, что в последние годы (2014–2017) сезонная динамика рачка T. oithonoides находится в пределах ее межгодовой изменчивости. Определенных трендов в изменении численности и биомассы вследствие изменения климата и сдвигов сезонных сроков, а также колебаний антропогенной нагрузки отмечено не было. В сезонной динамике T. oithonoides из центрального района озера и Петрозаводского залива обнаружены различия. Так, в холодноводном центральном районе жизненный цикл рачка имеет дициклический характер, а в заливе с благоприятными температурными условиями зафиксирована его ди- и трицикличность. Настоящая работа является начальным этапом подробного исследования годовой цикличности планктонных рачков и в перспективе позволит использовать современные модели жизненных циклов (регрессионные, стохастические или имитационные), позволяющие делать прогноз сезонного развития популяции при различных температурных режимах и трофических условиях.
Заключение
Веслоногий рачок T. oithonoides является основным компонентом зоопланктона Онежского озера и важным кормовым объектом для рыб-планктофагов. Жизненный цикл рачка в холодноводном олиготрофном районе озера имеет дициклический характер, в крупном Петрозаводском заливе из-за благоприятного температурного режима возможна ди- и трицикличность.
Среднемноголетние максимумы популяций в центре озера по численности достигают 1.31 тыс. экз./м3, биомассе – 0.009 г/м3. Величины для популяции в Петрозаводской губе сходные, по численности – 1.36 тыс. экз./м3, по биомассе – 0.013 г/м3 соответственно. В июне неблагоприятные гидрохимические условия в заливе угнетают развитие рачка и вызывают задержку его количественного развития. Колебания антропогенной нагрузки и климатические изменения пока не оказывают влияния на характер годовой цикличности рачка в Онежском озере.
Библиография
Гусаков В. А. Мейобентос Горьковского водохранилища // Биологические ресурсы пресных вод: беспозвоночные. Рыбинск: Изд-во ОАО «Рыбинский дом печати», 2005. С. 98–141.
Калинкина Н. М., Теканова Е. В., Сабылина А. В., Рыжаков А. В. Изменения гидрохимического режима Онежского озера с начала 1990-х годов // Известия РАН. Сер. географическая. 2019. № 1. С. 62–72.
Калинкина Н. М., Куликова Т. П. Эволюционная обусловленность реакции гидробионтов на изменение ионного состава воды (на примере пресноводного планктона) // Известия РАН. Сер. биологическая. 2009. № 2. С. 243–248.
Крупнейшие озера-водохранилища Северо-Запада европейской территории России: современное состояние и изменения экосистем при климатических и антропогенных воздействиях . Петрозаводск: Карельский научный центр РАН, 2015. 375 c.
Куликова Т. П. Зоопланктон водных объектов бассейна Белого моря . Петрозаводск: КарНЦ РАН, 2010. 325 с.
Куликова Т. П. Зоопланктон водных объектов бассейна Онежского озера . Петрозаводск: КарНЦ РАН, 2007. 223 с.
Куликова Т. П. Зоопланктон водных объектов бассейна реки Шуя . Петрозаводск: КарНЦ РАН, 2004. 124 с.
Куликова Т. П. Зоопланктон водных объектов Республики Карелия (Россия): История изучения, основные направления исследований, видовой состав, библиография . LAP LAMBERT Academic Publishing, 2017. 125 с.
Куликова Т. П. Зоопланктон водных объектов северной части бассейна Ладожского озера . Петрозаводск: КарНЦ РАН, 2012. 192 с.
Куликова Т. П., Сярки М. Т. Размерно-весовая характеристика массовых видов ракообразных и коловраток Онежского озера (справочно-информационный материал) . Петрозаводск: Карельский научный центр РАН, 1994. 16 с.
Лазарева В. И. Структура и динамика зоопланктона Рыбинского водохранилища . М.: Товарищество научных изданий КМК, 2010. 183 с.
Лазарева В. И., Смирнова С. М. Ракообразные и коловратки // Состояние экосистемы озера Неро в начале XXI века. М.: Наука, 2008. С. 175–210.
Лозовик П. А., Зобков М. Б., Бородулина Г. С., Токарев И. В. Оценка внешнего водообмена заливов озер по химическим показателям воды // Водные ресурсы. 2019. Т. 46. № 1. C. 91–101.
Методические рекомендации по сбору и обработке материалов в гидробиологических исследованиях на пресноводных водоемах. Зоопланктон и его продукция . Л.: ГосНИОРХ, 1984. 33 с.
Монаков А. В. Питание и пищевые взаимоотношения пресноводных копепод . Л.: Наука, 1976. 170 с.
Назарова Л. Е. Атмосферные осадки в Карелии // Труды Карельского научного центра РАН. 2015. № 9. С. 114–120.
Онежское озеро. Атлас . Петрозаводск: Карельский научный центр, 2010. 151 с.
Определитель зоопланктона и зообентоса пресных вод Европейской России. Т. 1. Зоопланктон . М.: Товарищество научных изданий КМК, 2010. 495 с.
Пидгайко М. Л. Зоопланктон водоемов европейской части СССР . М.: Наука, 1984. 208 с.
Родионова Н. В. Зоопланктон // Ладога. СПб.: Нестор-История, 2013. С. 298–306.
Рылов В. М. Фауна СССР. Ракообразные, Cyclopoida пресных вод . Т. III. Вып. 3. М.; Л.: Изд-во Академии наук СССР, 1948. 318 с.
Смирнова Т. С. Зоопланктон озер Воже и Лача // Гидробиология озер Воже и Лача в связи с прогнозом качества вод, перебрасываемых на юг. Л.: Наука, 1978. С. 102–130.
Сярки М. Т. Изучение траектории сезонной динамики планктона с помощью метода двойного сглаживания // Принципы экологии. 2013. № 1 (5). С. 61–67.
Сярки М. Т., Куликова Т. П. «Зоопланктон Онежского озера» : Свидетельство о государственной регистрации базы данных № 2012621150. Правообладатель: Федеральное государственное бюджетное учреждение науки Институт водных проблем Севера Карельского научного центра РАН (ИВПС КарНЦ РАН) (RU). Дата регистрации в реестре баз данных 9 ноября 2012 г.
Сярки М. Т., Теканова Е. В., Чекрыжева Т. А. «Планктон пелагиали Онежского озера» : Свидетельство о государственной регистрации базы данных № 2015620274. Правообладатель: Федеральное государственное бюджетное учреждение науки Институт водных проблем Севера Карельского научного центра РАН (ИВПС КарНЦ РАН) (RU). Дата регистрации в реестре баз данных 13 февраля 2015 г.
Тимохина А. Ф. Зоопланктон как компонент экосистемы Куйбышевского водохранилища . Тольятти: ИЭВБ РАН, 2000. 193 с.
Филатов Н. Н., Руховец Л. А., Назарова Л. Е., Георгиев А. П., Ефремова Т. В., Пальшин Н. И. Влияние изменения климата на экосистемы озер Севера Европейской территории России // Ученые записки Российского государственного гидрометеорологического университета. 2014. № 34. С. 48–55.
Филимонова З. И. Низшие ракообразные планктона озер Карелии // Фауна озер Карелии. Беспозвоночные. М., Л.: Наука, 1965. С. 111–146.
Фомина Ю. Ю., Сярки М. Т. Жизненный цикл рачка Eudiaptomus gracilis (Sars, 1863) в Онежском озере // Принципы экологии. 2018. № 3. С. 92–105. DOI: 10.15393/j1.art.2018.7842.
Хаберман Ю., Вирро Т., Бланк К. Зоопланктон // Псковско-Чудское озеро. Тарту: Eesti Loodusfoto, 2012. С. 285–306.
Adrian R., Wilhelm S. and Gerten D. Life-history traits of lake plankton species may govern their phenological response to climate warming // Global Change Biology. 2006. Vol. 12. P. 652–661.
Gerten D., Adrian R. Species-specific changes in the phenology and peak abundance of freshwater copepods in response to warm summers // Freshwater Biology. 2002. Vol. 47. P. 2163–2173.
Haberman J., Virro T. Zooplankton // Lake Võrtsjärv. Tallinn: Encyclopedia Publishers Ltd, 2004. P. 233–251.
Lehtovaara A., Arvola L., Keskitalo J., Olin M., Rask M., Salonen K., Sarvala J., Tulonen T., Vuorenmaa J. Responses of zooplankton to long-term environmental changes in a small boreal lake // Boreal environment research. 2014. 19 (suppl. A). P. 97–111.
Nilssen P., Wærvågen S. B. Superficial ecosystem similarities vs autecological stripping: the "twin species" Mesocyclops leuckarti (Claus) and Thermocyclops oithonoides (Sars) – seasonal habitat utilisation and life history traits // J. Limnol. 2000. Vol. 59 (2). P. 79–102.
Благодарности
Работа выполнена в рамках Государственного задания Института водных проблем Севера КарНЦ РАН.

 © 2011 - 2026
© 2011 - 2026