



Введение
Оценка характеристики «плодовитость» или «рождаемость» является важнейшим этапом и начальным звеном любых исследований в области популяционной экологии. Самки подавляющего числа видов бесхвостых земноводных умеренной климатической зоны ежегодно откладывают единственную и компактную порцию яиц, и их количество в кладке соответствует величине реальной плодовитости данной самки. Следовательно, задача состоит в том, чтобы подсчитать количество икринок в кладках с точностью, достаточной для поставленных целей исследования конкретной популяции. Варианты существующих способов оценки плодовитости состоят в том, чтобы сократить трудоемкость и минимизировать затраты времени на процедуры, которые необходимо проделывать (см., например, Bohenek, Resetarits, 2017).
Цель настоящей работы – представление метода, не нарушающего целостность икряного комка и максимально упрощающего процедуру подсчета икринок в кладках лягушек в полевых условиях.
Материалы
Кладки икры, всего 49 шт., взяты из репродуктивных водоемов в период массового икрометания травяной лягушки. Возраст кладок составлял 1–3 дня. За это время студенистые оболочки яиц набухали, сцепление между ними ослабевало, а развитие эмбрионов не шло дальше формирования у них нервной трубки (стадия 16 по Терентьеву, 1950). В таком состоянии комки икры удобно помещать в узкие емкости, не деформируя при этом студенистые оболочки и не травмируя эмбрионы.
Традиционные методы исследований
Очевидны простота и точность поштучного пересчета икринок в кладке, однако этот способ имеет два недостатка. Во-первых, он требует значительных затрат времени. В одном из предшествующих экспериментов мы провели хронометраж работы группы учетчиков из трех человек: двое последовательно отделяли от кладки небольшие порции и подсчитывали в них икринки, а третий записывал результаты счета и суммировал их. Время на подсчет икринок в одной кладке, в зависимости от ее состояния (степень набухания слизистых оболочек) и объема (от 930 до 1611 икринок в одной кладке), занимало от 12 до 20 минут.
Второй, более существенный недостаток заключается в неизбежном полном разрушении кладки и повреждении слизистых оболочек в процессе подсчета. Последствия могут быть фатальными для эмбрионов, и возвращение такой икры в репродуктивный водоем бессмысленно. В итоге данный способ оказывается совершенно непригодным в случае работы, например, с кладками икры редких видов земноводных или слишком трудоемким, когда требуется произвести подсчет яиц в большом количестве кладок.
Усовершенствованием метода ручного пересчета послужило сплющивание комка икры с помощью двух прозрачных пластиковых контейнеров. Кладку помещали на дно одного и плавно прижимали сверху дном второго контейнера, добиваясь распределения икринок в один слой. Небольшие кладки икры Ambyostoma maculatum помещали целиком, но крупные икряные массы Rana sylvatica приходилось разделять на несколько порций. Разлиновка на дне верхнего контейнера облегчала подсчет и позволяла повышать его точность (Karraker, 2007). Физические повреждения эмбрионов и слизистых оболочек были невелики, и после пересчета икру возвращали в водоем.
Широко и издавна применяется объемный метод оценки, когда измеряют объем всей кладки и объем отделенной от кладки порции с определенным числом икринок, а затем путем пересчета на общий объем кладки получают количество яиц, выметанных одной самкой (Щупак, 1970; Cooke, 1975; Ляпков, 2003; Räsänen et al., 2008 и мн. др.). Вариантом является метод весовых проб, когда определяют не объем, а вес кладки и ее фрагмента с установленным количеством икринок (Loman, 2001). Объем устанавливают, используя мерные цилиндры, и по завершении процедуры кладку икры возвращают на место. Время, затрачиваемое на обработку, зависит от диаметра цилиндра, используемого для определения объема всей кладки. В нашем эксперименте отделение от кладки и подсчет 100 икринок, определение объема этой пробы и объема всей кладки занимало 7–12 минут при диаметре мерного цилиндра 45 мм или 4–6 минут при диаметре 90 мм.
В. Г. Ищенко (2008) обратил внимание на существенные различия между значениями плодовитости, полученными объемным методом, и истинной плодовитости, определенной прямым пересчетом яиц в кладке. Мы рассчитали эту разницу, обработав двумя способами 29 комков икры травяной лягушки (Кутенков, 2013). Отклонение расчетных величин от полученных при поштучном пересчете составляло от –20 до +30 %, однако при усреднении значение отклонения составило всего 3–7 % в зависимости от диаметра мерного цилиндра.
А. Хаапанен (Haapanen, 1982) применил следующий способ. Взятую из водоема кладку травяной лягушки распределяли по разлинованному фону так, чтобы икринки образовали один слой. После фотографирования и печати фотографий (работа проведена в 1960-е годы) рассчитывали плотность размещения икринок и, с учетом занимаемой кладкой площади, устанавливали их суммарное количество. Трудоемкость такого способа в те годы очевидна, однако несомненным преимуществом являлось то, что работа в поле занимала немного времени и не требовала дополнительного оборудования – только фотоаппарат и палетка. Все дальнейшие процедуры – проявка, печать, подсчет – неспешно осуществлялись в комфортной обстановке.
В последнее время исследователи стали использовать преимущества компьютерной техники и цифровой фотографии в целях сокращения времени на процедуру подсчета. На четырех видах североамериканских земноводных (Ambyostoma maculatum, Hyla chrysoscelis, Lithobates (Rana) sylvaticus и L. pipiens), откладывающих компактные шарообразные порции икры, была отработана методика автоматического подсчета величины кладки по фотографиям (Bohenek, Resetarits, 2017; Moraga, Pervin, 2018).
В одном случае кладку доставляли в лабораторию и распределяли икру в один слой в большом часовом стекле. Затем ее фотографировали с помощью специально оборудованного копировального штатива с подсветкой и с зафиксированной фотокамерой. Во втором случае кладку фотографировали прямо в поле, используя описанное выше приспособление из двух пластиковых сосудов, после чего ее возвращали в водоем (вся процедура занимала около 30 сек.).
Дальнейшую процедуру автоматического подсчета яиц осуществляли по общей схеме. Фотографию икряной массы импортировали в среду программы обработки изображений ImageJ (создана для нужд микробиологии и биологии клетки). Добиваясь наивысшего качества изображения и производя окончательный подсчет, осуществляли многоступенчатый процесс с использованием до 33 команд. Контролем (или теоретическим значением) служил поштучный пересчет яиц, присутствующих на тех же тщательно обработанных изображениях кладок. Для этого использовали компьютерную программу подсчета с помощью многоточечной функции либо ручной счетчик для визуального счета на множественных подвыборках.
В процессе компьютерной обработки основные усилия авторов были направлены на достижение высокого уровня качества изображения каждой икряной массы: ликвидация «шумов» фона, бликов и отражений, удаление лишних объектов определенных размеров и т. д. В конце выясняли степень согласованности результатов автоматического и «ручного» пересчетов. На разных видах получаемая средняя разница величины кладки составляла -2.6...+23.8 %, а значения коэффициента корреляции оказывались в пределах 0.75–0.99 (p < 0.001).
Оригинальные методы исследований
Изготовлена конструкция из стекла в форме параллелепипеда (рис. 1). Размеры емкости: высота 15 см, длинa 26 см, ширина 2.5 см, расстояние между широкими стенками 2 см, внутренний объем 975 мл. Пропорции подобрали с таким расчетом, чтобы туда помещалась вся кладка целиком (предварительно установили, что объем набухшей кладки травяной лягушки не превышал 910 мл), а икринки внутри прибора располагались в 1–2 слоя по вертикали. При этом условии и благодаря упругим слизистым оболочкам яйца редко заслоняют друг друга.
На погружение одной кладки в емкость тратили от 10 до 40 секунд в зависимости от степени набухания слизистых оболочек икринок, еще примерно 15 секунд уходило на съемку цифровой фотокамерой «с рук».

Рис. 1. Внешний вид устройства для фотографирования кладки икры лягушек
Fig. 1. The appearance of the device to photograph frogs’ spawn clutches
Используя любой соответствующий компьютерный редактор, полученное цветное изображение следует перевести в черно-белый формат, увеличив затем яркость и контрастность (рис. 2).

Рис. 2. Вид кладки икры, помещенной в сосуд, в цветном (слева) и черно-белом (справа) изображении
Fig. 2. A view of the spawn clutch in the vessel in color (left) and black and white (right) image
Дальнейшую процедуру осуществляли после импорта файла с изображением кладки икры в среду векторного графического редактора CorelDRAW X6 (рис. 3). Нажатием правой кнопкой мыши на изображение вызывали меню, где выбирали «Трассировка абрисом», затем «Изображение низкого качества» (рис. 4). Далее с помощью бегунка в разделе «Деталь» выполняли приближение изображения и выравнивали степень детализации до того момента, когда на нижнем изображении оставались хорошо видимыми только икринки (рис. 5). После этого здесь же, в разделе «Кривые», отображается число ограниченных векторных объектов (рис. 6), в нашем случае это количество икринок, обнаруженное на фотографии кладки. Серые области перехода тонов в данном случае не учитываются, так как программа рассматривает их как часть фона из-за близкого по оттенку серого цвета.
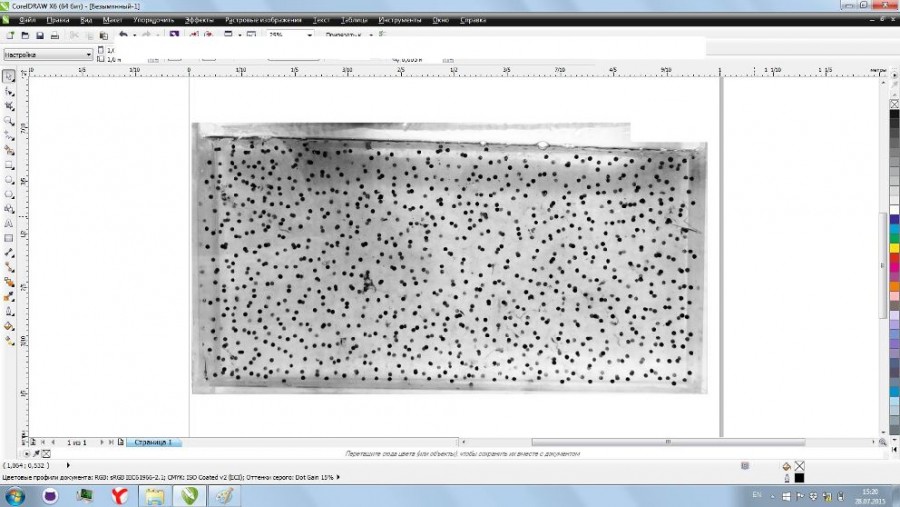
Рис. 3. Изображение кладки икры, импортированное в пакет CorelDRAW X6
Fig. 3. An image of the spawn clutch, uploaded into CorelDRWA X6 software
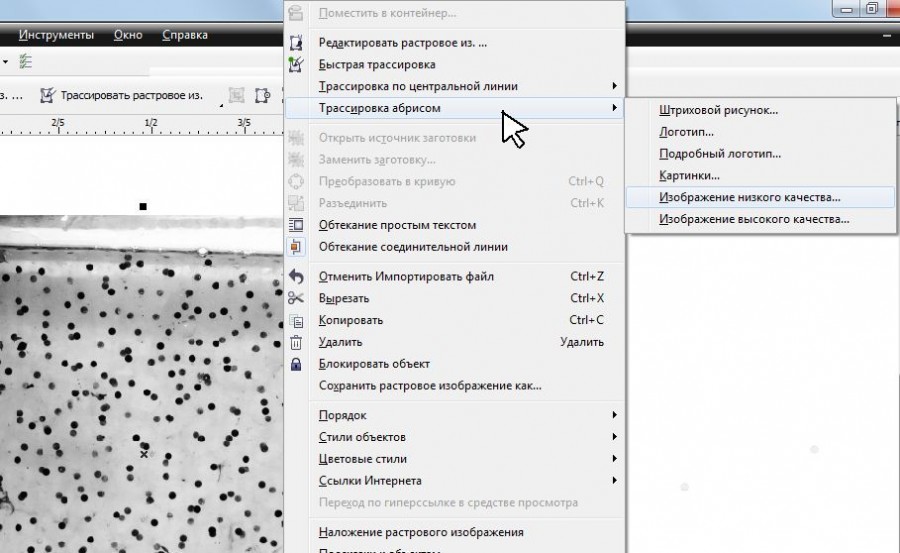 Рис. 4. Процедура «трассировка абрисом», выбор «изображение низкого качества»
Рис. 4. Процедура «трассировка абрисом», выбор «изображение низкого качества»
Fig. 4. "Trace Bitmap" procedure, "Low Quality Image" has chosen
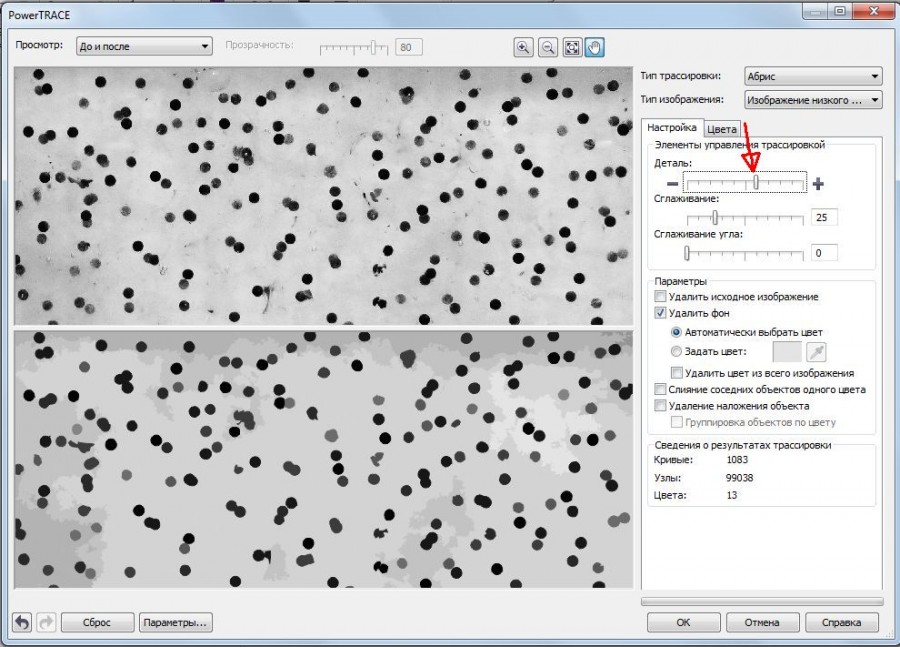
Рис. 5. Использование раздела «Деталь» для выравнивания степени детализации на приближенном изображении. Красная стрелка указывает на бегунок
Fig. 5. The usage of the division "Detail" to flatten the rate of itemization on a zoomed image. The roller is marked with red arrow
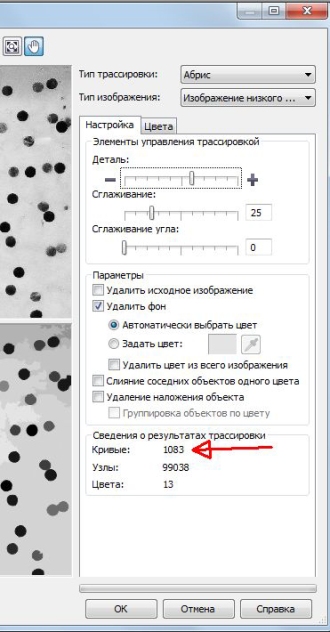
Рис. 6. Результат подсчета количества икринок находится в разделе «Кривые» (указан стрелкой)
Fig. 6. The result of counting the number of ova is in the "Curves" division (marked with arrow)
Кладки икры травяной лягушки приносили в лабораторию, где каждую из них сначала фотографировали в нашем устройстве, затем определяли количество икринок методом объемных проб и, наконец, делали поштучный пересчет. Анализ изображений кладок в графическом редакторе проводили позже, в удобное время. Статистическая обработка полученных результатов выполнена в среде MS Exel с использованием общепринятых алгоритмов (Ивантер, Коросов, 2011).
Сравнивали результаты применения трех способов оценки величины кладок (в скобках даны аббревиатуры, используемые в последующем тексте). Контролем считали поштучный пересчет (ПП). С контролем сравнивали результаты предлагаемого метода оценки с помощью векторного графического редактора CorelDRAW X6 (ВГР) и данные, полученные методом объемных проб с использованием мерного цилиндра диаметром 90 мм и с ценой деления 100 мл (МОП).
В выборке из 49 комков икры среднее число яиц в кладке при разных способах оценки составило: ПП — 1016 ± 45 шт., ВГР — 1025 ± 41 шт. и МОП — 1053 ± 47 шт. Диапазон отклонений от контроля числа яиц в кладках нашей выборки находился в пределах от –291 до +108 (в среднем +9) шт. при оценке с помощью ВГР и от –253 до +357 (в среднем +37) шт. при оценке с помощью МОП. В процентном выражении это составило от –14 до +12 %, в среднем –2 % (ВГР) и от –20 до +33 %, в среднем +4 % (МОП) от величины конкретной кладки. Диапазон выявленных значений плодовитости и графики зависимости величин кладок, полученных способами ВГР и МОП, от контрольного поштучного пересчета приведены на рис. 7 и 8.
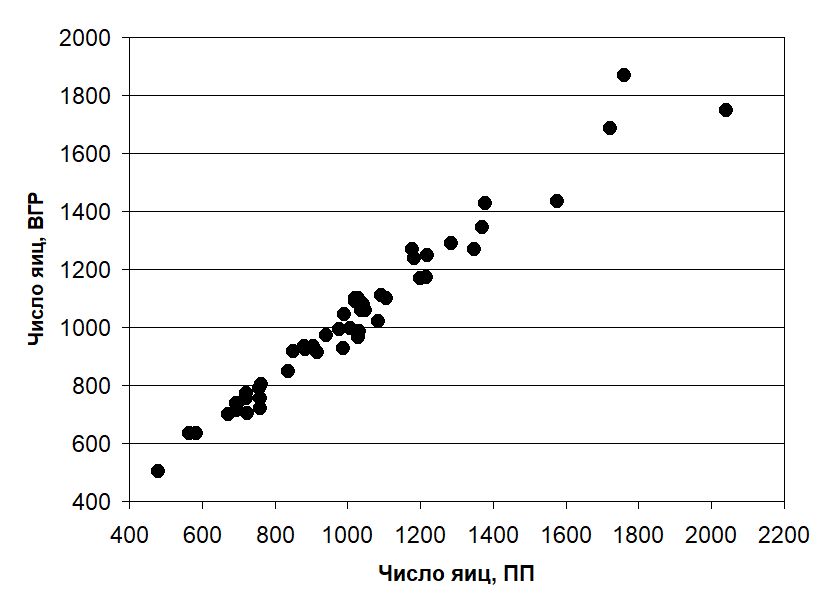
Рис. 7. Соотношение количества яиц в 49 кладках икры R. temporaria, полученное при поштучном пересчете (ПП) и с использованием векторного графического редактора (ВГР) (rs = 0.98, p < 0.0001)
Fig. 7. Relationship between the number of ova in 49 clutches of R. temporaria estimated using manual counting (ПП) and vector graphic editor software (ВГР) (rs = 0.98, p < 0.0001)
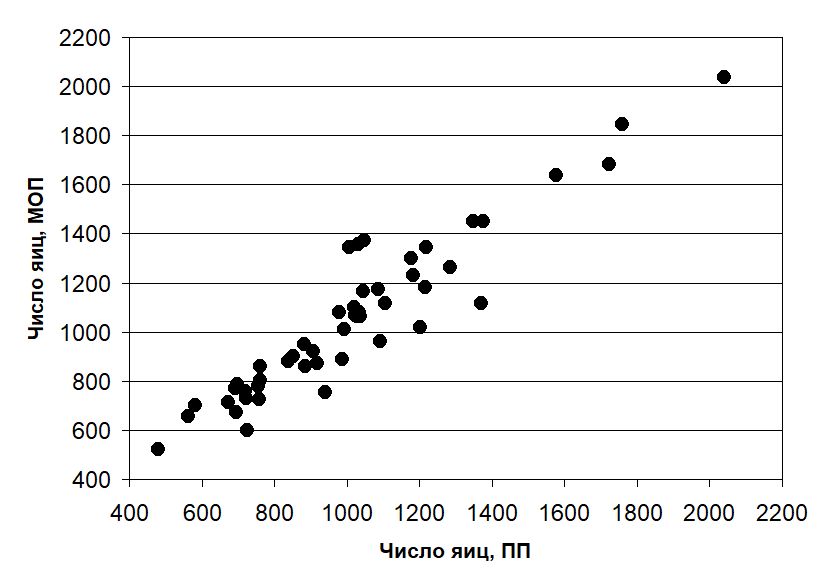
Рис. 8. Соотношение количества яиц в 49 кладках икры R. temporaria, полученное при поштучном пересчете (ПП) и методом объемных проб (МОП) (rs = 0.92, p < 0.0001)
Fig. 8. Relationship between the number of ova in 49 clutches of R. temporaria estimated using manual counting (ПП) and bulk sampling method (МОП) (rs = 0.92, p < 0.0001)
Отмечаем очень высокую степень согласованности оценок плодовитости, выразившуюся в значениях коэффициентов ранговой корреляции. Однако, как следует из приведенных графиков, зависимость в случае с ВГР оказывается более тесной.
Оценку различий результатов подсчета разными способами провели с помощью однофакторного дисперсионного анализа (влияющий фактор – способ подсчета, контроль – поштучный пересчет). Различия с контролем оказались недостоверными: значения F-критерия составили при оценке с помощью ВГР 0.019, при МОП — 0.327 (F0.05 = 3.9).
Данные статистики, таким образом, показывают, что результаты поштучного пересчета икринок в каждой кладке и полученные при оценке с помощью графического редактора достаточно близки.
Возможные причины отклонений числа яиц (кроме вполне вероятных ошибок при ПП: повторных контрольных пересчетов не делали) состоят в следующем. Метод объемных проб дает закономерно завышенную оценку (Кутенков, 2013). Это связано в основном с тем, что в процессе отделения от кладки объемной пробы слизистые оболочки отдельных яиц лопались в неопределенной пропорции, и измеренный объем пробы оказывался заниженным в той или иной степени. При подстановке в формулу данная величина служит делителем, что и ведет к завышенному в целом результату. При подсчете в среде графического редактора излишек возникает из-за того, что при анализе изображения происходит трассировка мелкого мусора, попадающего в сосуд вместе с икрой. Недоучет же, очевидно, возникает при фотографировании «молодых» кладок с не полностью набухшими оболочками. Тогда икра в сосуде располагается более чем в два вертикальных слоя, небольшая часть икринок полностью или частично заслоняют друг друга, и программа в этих случаях считает их за одну. Чтобы снять проблему, достаточно такую кладку оставить на несколько часов в водоеме, после чего сфотографировать повторно.
Заключение или выводы
Предлагаемый способ оценки плодовитости, апробированный на Rana temporaria, не трудоемок и пригоден для обработки большого количества кладок икры на местах нереста лягушек. Естественное и почти не нарушаемое сцепление слизистых оболочек яиц позволяет после фотографирования возвращать кладку в репродуктивный водоем практически неповрежденной. Варьируя размерами конструкции для фотографирования (ширина и внутренний объем), ее можно использовать для подсчета яиц у разных видов лягушек, откладываемые порции икры у которых существенно различаются по объему.
Предлагаемый метод имеет два основных преимущества. Во-первых, в отличие от перечисленных выше способов, использование в качестве инструмента нашего «аквариума» дает возможность фотографировать кладки с большим количеством яиц целиком. Во-вторых, обработка изображений с помощью распространенного и доступного графического редактора предельно проста, при этом точность получаемых оценок оказывается не ниже, чем при работе в среде громоздких специальных программ.
Библиография
Ивантер Э. В., Коросов А. В. Введение в количественную биологию . Петрозаводск, 2011. 302 с.
Ищенко В. Г. Долговременные исследования демографии популяций амфибий: современные проблемы и методы // Вопросы герпетологии: Материалы III съезда Герпетол. об-ва им. А. М. Никольского. Пущино; М., 2008. С. 151–169.
Кутенков А. П. Сравнение двух способов оценки плодовитости травяной лягушки (Rana temporaria L.) // Труды Государственного природного заповедника «Кивач». Вып. 6. Петрозаводск, 2013. С. 164–166.
Ляпков С. М. Сохранение и восстановление разнообразия амфибий европейской части России: разработка общих принципов и эффективных практических мер . М.: КМК, 2003. 116 с.
Терентьев П. В. Лягушка . М.: Сов. наука, 1950. 346 с.
Щупак Е. Л. Динамика биологической продуктивности популяции остромордой лягушки // Экология. 1970. № 1. С. 83–86.
Bohenek J. R., Resetarits W. J. An optimized method to quantify large numbers of amphibian eggs // Herpetology notes. 2017. Vol. 10. P. 573–578.
Cooke A. S. Spawn clumps of the common frog Rana temporaria: number of ova and hatchability // Brit. j. of herpetology. 1975. Vol. 5. № 5. P. 505–509.
Haapanen A. Breeding of the common frog (Rana temporaria L.) // Ann. zool. fennici. 1982. Vol. 19. № 2. P. 75–79.
Karraker N. E. A new method for estimating clutch sizes of ambystomatid salamanders and ranid frogs: introducing the ovagram // Herpetological revive. 2007. Vol. 38. № 1. P. 46–48.
Loman J. Local variation in Rana temporaria egg and clutch size: adaptation to pond drying? // Alytes. 2001. Vol. 19. № 1. P. 45–52.
Moraga A. D., Pervin E. Efficient estimation of amphibian clutch size using image analysis of compressed globular egg masses // Herpetological conservation and biology. 2018. Vol. 13. № 2. P. 341–346.
Räsänen K., Söderman F., Laurila A., Merilä J. Geographic variation in maternal investment: acidity affects egg size and fecundity in Rana arvalis // Ecology. 2008. Vol. 89. № 9. P. 2553–2562.
Благодарности
Значительную часть работы по подсчету яиц в кладках травяной лягушки поштучным пересчетом и методом объемных проб выполнили учащиеся эколого-биологического центра «Крестовский остров» Санкт-Петербургского городского Дворца творчества юных, а также учащиеся и учителя средних школ г. Кондопоги (Республика Карелия). В разработке и исполнении конструкции для фотографирования принимал участие А. С. Мамонтов. А. С. Уткин предложил идею использовать для подсчета икринок векторный графический редактор CorelDRAW X6 и помог в нем разобраться. Всем этим людям мы искренне благодарны. Мы также весьма признательны рецензентам В. Г. Ищенко и С. М. Ляпкову за ценные замечания, практические советы и другую помощь при работе над статьей.

 © 2011 - 2026
© 2011 - 2026