



Введение
Оценка экологических последствий массовой промышленной лесоэксплуатации, и в частности ее влияния на население млекопитающих и других животных, продолжает вызывать большой интерес. Между тем результаты исследований в этой области чаще всего публикуются в виде кратких обзоров, практически лишены каких-либо обобщений и в основном относятся ко второй половине XX в. (Динесман, 1961; Ельшин, Каратаев, 1988; Калинин, 1973; Керзина, 1956; Ларин, 1955; Наумов, Руковский, 1979; Diffendorfer et al., 1978; Hansson, 1992; Keith et al., 1993; Lindzey, 1989).
Цель работы — восполнить этот пробел результатами многолетних исследований (1958–2017 гг.), выполненных в Восточной Фенноскандии (Кольский п-ов, Карелия, Финляндия и Карельский перешеек Ленинградской обл.) и сочетавших работу на полевых стационарах (Гомсельгский, Ладожский, заповедник «Кивач») с широкими экспедиционными обследованиями.
Материалы
Материалы и методы
Основной объект исследований – весьма важная в биоценотическом плане группа мелких млекопитающих: землеройки-бурозубки и лесные мышевидные грызуны. Всего на исследованной территории добыто 11 видов мелких млекопитающих, которые по степени доминирования в суммарных уловах располагаются следующим образом: обыкновенная бурозубка (Sorex araneus L.), рыжая полевка (Cletrionomus glareolus Schreb.), малая бурозубка (S. minutus L.), средняя бурозубка (S. caecutiens Laxm.), темная полевка (Microtus agrestis L.), лесная мышовка (Sicista betulina Pall.), лесной лемминг (Myopus schisticolor Lill.), равнозубая бурозубка (S. isodon Turov), полевка-экономка (M. oeconomus Pall.), мышь-малютка (Micromys minutus Pall.), крошечная бурозубка (S. minutissimus Zimm.).
Эти виды, занимая особое место в структуре трофических связей таежных экосистем и будучи основными потребителями растительности и беспозвоночных нижнего яруса леса, служат одновременно важнейшим объектом потребления многими хищниками. Учеты численности этих зверьков заключались в расстановке параллельных (на расстоянии 25 м одна от другой) линий давилок (капканчиков Геро) – по 25–50 шт. в каждой. Они равномерно распределялись по всем обследуемым биотопам (и модельным участкам) и действовали по 2–4 суток. Приманкой служили кусочки смоченного растительным маслом ржаного хлеба. За показатель обилия принимали число зверьков, попавших за 1-е сутки работы в 100 ловушек (на 100 лов./сут.), и выраженную в процентах долю данного вида в общем улове ловушками (относительное обилие в населении мелких млекопитающих, или индекс доминирования). Учет и отлов канавками проводился с помощью 30-метровых траншей, имевших по три металлических конуса, сужающиеся к горловине и врытые таким образом, что верхний их край находился вровень с дном канавки. Показатель обилия – число зверьков, попавших в конусы за 10 суток работы одной канавки (на 10 кан./сут.), и относительное число зверьков данного вида, выраженное в долях общего числа добытых (индекс доминирования, %). Общий объем проведенных учетов превышает 455000 лов./сут. и 7800 кон./сут.
Один из используемых нами показателей видового разнообразия сообществ – индекс Симпсона, рассчитываемый по формуле I = 1 – ∑(ni / N)2, где ni – оценка значимости одного вида, N – сумма оценок значимости всех видов (Керзина, 1956). Поскольку в нашем случае этот индекс вычислялся только на основе относительных показателей (в экз. на 100 лов./сут.), он используется здесь лишь как способ грубого сравнения видового разнообразия биоценотических комплексов мелких млекопитающих экспериментальной и контрольной территорий.
Результаты
Результаты и их обсуждение
Анализ изменений видовой структуры и разнообразия биоценотических группировок мелких млекопитающих показал, что с сокращением размеров фрагментов хвойных лесов сокращалось и число видов, зафиксированных в среднем за год наблюдений (фактически это индекс видового богатства), причем наиболее резкий переход (в 2 раза) наблюдался от крупных к небольшим (6–7 га) массивам (рис. 1). Характерно, что показатель обилия мелких млекопитающих на небольших и мелких участках сосняков оказался также примерно в 1.5 раза ниже, чем в крупных массивах. Заметные изменения наблюдаются и при сокращении размеров фрагментов от 20–30 до 6–7 га.
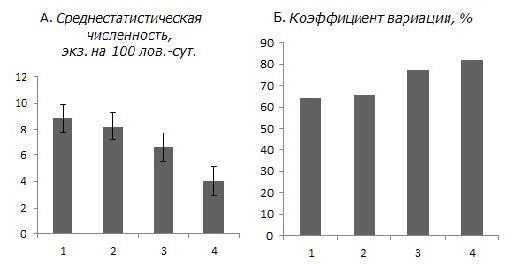
Рис. 1. Численность мелких млекопитающих и ее динамика в сосняках разной степени фрагментации. А – среднестатистическая численность, экземпляров на 100 ловушко-суток, Б – коэффициент вариации, %, 1 – заповедные территории, 2 – крупные массивы (> 30 га), 3 – небольшие массивы (6–7 га), 4 – мелкие выделы до 2 га
Fig. 1. The number of small mammals and its dynamics in pine forests of different fragmentation degree of . A – the average number of specimens per 100 trap-days, B – coefficient of variation,%, 1 – protected areas, 2 – large tracts (> 30 ha), 3 – small tracts (6–7 ha), 4 – small areas up to 2 hectares
Тренды динамики численности мелких млекопитающих в сосняках различной степени фрагментированности в целом имеют сходную направленность. При этом следует отметить два аспекта: во-первых, размах колебаний (особенно максимумы), который даже в слабофрагментированных сосняках (20–30 га) оказался несколько шире, чем на контрольной (заповедной) территории; во-вторых, закономерность, согласно которой дальнейшая фрагментация сосняков (до 6–7 га и особенно до 1–2 га) резко усиливает флуктуации динамики популяций мелких млекопитающих от общих тенденций, характерных для крупных лесных массивов. Это прежде всего проявляется в ином чередовании пиков обилия и в размахе колебаний.
Особенно четкие различия наблюдаются при анализе численности двух фоновых видов (обыкновенной бурозубки и рыжей полевки), которые вместе составляют > 90 % суммарного отлова. С сокращением размеров лесных фрагментов показатель обилия этих видов сокращается, а коэффициент вариации (Cv) возрастает.
Наблюдаются различия и в динамике численности. В крупных таежных массивах заповедника волны популяционной динамики обыкновенной бурозубки имеют более сглаженный характер и спады численности выражены слабее, чем в небольших по площади сосняках. В мелких лесных массивах периоды депрессии численности видов выражены более резко и нередко принимают многолетний характер.
Таким образом, фрагментация сосняков в целом приводит к обеднению видового состава, сокращению численности видов, усилению нестабильности населения и упрощению структуры сообществ мелких млекопитающих. Максимальная степень фрагментации (сокращение площадей до 1–2 га) приводит к формированию довольно нестабильных и упрощенных сообществ с 1–2 абсолютно доминирующими видами.
В процессе дальнейшей антропогенной трансформации таежных лесов через некоторое время после начала их фрагментации формируется ландшафт, представляющий собой мозаику лесных биотопов, находящихся на разных этапах вторичной сукцессии после сплошных рубок. В этой связи большой интерес представляет изучение экологических последствий для животных формирования именно такого мозаичного ландшафта. Результаты учета мелких млекопитающих (табл. 1 и 2) с достаточной точностью отражают как общий относительно невысокий уровень численности видов, так и совпадение волн динамики численности в опыте и контроле: годы пиков и депрессий почти синхронны, совпадает и средняя амплитуда колебаний обилия зверьков в смежные годы.
Таблица 1. Численность мелких млекопитающих на экспериментальном (1) и контрольном (2) участках за 11 лет исследований, экз. на 100 лов./сут. (Численность и распределение..., 2013)
| Участок | Исследованная группа | Численность | Размах колебаний численности в смежные годы, M ± SE
|
||
| суммарная | средняя,
M ± SE |
Cv,
% |
|||
| 1. Гомсельгский стационар | Мелкие млекопитающие | 9.1 | 9.5 ± 1.8 | 64 | 2.1 ± 0.2 |
| бурозубки | 3.1 | 3.1 ± 0.8 | 88 | 3.6 ± 1.0 | |
| мышевидные грызуны | 6.0 | 6.4 ± 1.2 | 63 | 2.1 ± 0.4 | |
| 2. Заповедник «Кивач» | Мелкие млекопитающие | 7.2 | 7.1 ± 1.2 | 55 | 2.1 ± 0.3 |
| бурозубки | 2.9 | 2.7 ± 0.6 | 71 | 1.8 ± 0.2 | |
| мышевидные грызуны | 4.3 | 4.4 ± 0.9 | 68 | 3.2 ± 0.9 | |
Примечание. М – средняя арифметическая, SE – стандартная ошибка средней для p = 0.05, Cv – коэффициент вариации.
Таблица 2. Численность (в числителе, экз. на 100 лов./сут.) и доля (в знаменателе, %) фоновых видов мелких млекопитающих в коренных и трансформированных рубками биотопах средней тайги Восточной Фенноскандии
| Биотопы | Число лов./сут. | Общая численность | Численность по видам | ||||||
| Бурозубка | Полевка | Лесная мышовка | |||||||
| обыкновенная | средняя | малая | рыжая | темная | экономка | ||||
| Спелые сосняки зеленомошные | 7640 | 4.8 | 1.7/35.5 | 0.06/1 | 0.06/1 | 2.8/60.9 | 0.14/2.5 | – | 0.04/0.05 |
| Спелые ельники зеленомошные | 3795 | 4.6 | 1.6/34.5 | 0.08/1.2 | 0.08/1.2 | 2.8/62 | 0.02/0.8 | – | – |
| Открытые вырубки | 7031 | 8.9 | 3.7/41.3 | 0.01/0.1 | 0.1/1.1 | 1.9/22.5 | 2.3/26.1 | 0.8/8 | 0.07/1 |
| Свежие (до 1 года) | 725 | 1.0 | 0.4/37.7 | – | – | 0.5/50.5 | – | 0.1/13 | – |
| От 1 до 5 лет | 6306 | 9.8 | 4.1/41.1 | 0.01/0.1 | 0.1/1.2 | 2.1/22.3 | 2.6/26.2 | 0.8/8.2 | 0.05/1 |
| Молодняки, лет | |||||||||
| 6–20 | 9551 | 7.4 | 2.8/39 | 0.04/0.6 | 0.12/1.5 | 2.9/39 | 1.0/14 | 0.4/5 | 0.04/0.5 |
| 20–40 | 3004 | 2.7 | 0.6/23 | 0.1/4 | – | – | – | – | – |
| Вторичные лиственные и смешанные леса (50–80 лет) | 4891 | 4.8 | 2.2/46 | 0.02/0.4 | 0.1/3 | 2.1/43 | 0.06/1 | 0.1/3 | 0.03/0.4 |
| Семенные куртины | 1140 | 4.1 | 1.6/38.0 | – | 0.1/2 | 2.3/58 | 0.1/2 | – | – |
| Недорубы | 1655 | 7.1 | 2.8/39.0 | – | – | 3.8/55 | 0.4/5 | – | – |
| Прочие | 1415 | 4.7 | 2.5/53.0 | – | 0.1/1 | 2.2/47 | – | – | – |
| Экотоны (лес-вырубка) | 960 | 11.7 | 3.0/26.0 | 0.1/1 | – | 4.7/39 | 2.9/25.2 | 0.9/8.1 | – |
Примечание. «–» – вид отсутствовал в уловах.
Более заметны различия между контролем и опытом лишь у мышевидных грызунов. Число видов этой группы, фиксируемых в среднем за год учетов давилками, оказалось значительно (в 1.6 раза) и достоверно (р < 0.05) выше, чем в заповеднике, в основном за счет регулярного присутствия в уловах зеленоядных видов серых полевок рода Microtus (темная, экономка). Это особенно характерно для ранних этапов вторичной сукцессии лесной экосистемы.
Если брать всю группу мелких млекопитающих, то различия показателей видового разнообразия между опытом и контролем незначительны (соответственно 0.55 и 0.46). При сравнении же отдельных групп видов (землеройки и мышевидные грызуны) различия вновь оказываются существенными: индекс видового разнообразия землероек в заповеднике примерно в 3 раза выше, чем в опыте (0.21 и 0.07), а для мышевидных грызунов, наоборот, значения индекса Симпсона в трансформированном рубками ландшафте в 5.4 раза выше (р < 0.01). Таким образом, трансформация таежных лесов приводит к сокращению видового разнообразия землероек-бурозубок, но к росту видового разнообразия мышевидных грызунов.
При анализе особенностей динамики численности (обилия) мелких млекопитающих на экспериментальной и контрольной территориях следует отметить, что в обоих случаях основной фон создают доминирующие виды – обыкновенная бурозубка и рыжая полевка, составляющие в опыте соответственно 33 и 43 % улова, а в контроле соответственно 35 и 56 %. Волны флуктуации численности обыкновенной бурозубки в заповеднике имеют более сглаженный вид с менее выраженными пиками и депрессией численности по сравнению с опытом (рис. 2). Для рыжей полевки соотношение иное: при примерно равных показателях обилия зверьков периоды депрессий и пиков в опыте выражены значительно слабее, чем в лесах заповедника, а депрессия 1983–1985 гг. вообще отсутствовала.
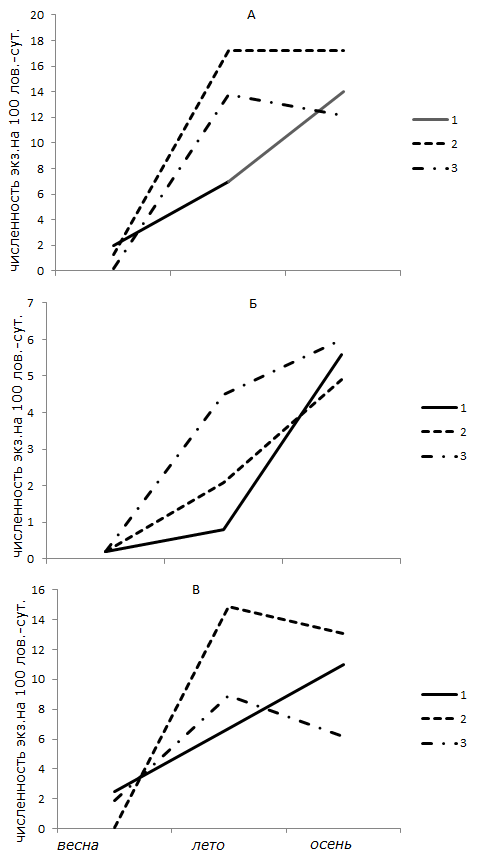
Рис. 2. Сезонная динамика численности мелких млекопитающих в смежных – коренных и трансформированных рубками биотопах. а – все виды, б – обыкновенная бурозубка, в – рыжая полевка. 1 – спелый сосняк черничный, 2 – вырубка (1–10 лет), 3 – сосново-лиственный молодняк (10–20 лет)
Fig. 2. Seasonal dynamics of the numbers of small mammals in adjacent – indigenous and transformed biotopes. a – all species, b – ordinary shrew, in – red vole. 1 – ripe pine bilberry, 2 – clearing (1–10 years), 3 – pine-leafed young stand (10–20 years)
Флуктуации численности отдельных видов в значительной степени влияют на состав группировок и динамику всего населения мелких млекопитающих. Например, если в заповедных лесах темная полевка регистрируется не ежегодно и ее доля в уловах лишь однажды превысила 3 % (1987 г.), то в трансформированном рубками ландшафте этот вид практически ежегодно встречался в уловах мышевидных грызунов, а в отдельные годы (1981, 1982) уверенно доминировал.
Одой из наиболее надежных характеристик населения мелких млекопитающих может служить сезонная динамика их численности, которая также демонстрирует существенные различия между опытом и контролем (см. рис. 2), особенно для одного из фоновых видов – обыкновенной бурозубки. По данным учетов, численность этого вида в трансформированном рубками ландшафте наиболее быстро растет весной и в первой половине лета, в результате чего различия между опытом и контролем в июле достигают 10 раз и более, к осени соотношение выравнивается. В таком же направлении, но слабее варьирует численность рыжей полевки.
На базе сводных для региона данных (20 пунктов стационарных и экспедиционных наблюдений (см. табл. 2) можно в общем охарактеризовать биоценотические группировки млекопитающих основных сукцессионных стадий экосистем на вырубках. Из имеющихся материалов следует, что спелые (старше 100 лет) сосняки зеленомошные (контроль) близки по численности и видовому разнообразию мелких млекопитающих к средним значениям этих показателей по всем биотопам, но значительно уступают вырубкам с давностью рубки от 1 до 15 лет, причем эти различия (достигающие 30 раз) особенно четко проявляются в группе мышевидных грызунов, в первую очередь серых полевок. Уже эти данные свидетельствуют о довольно явных признаках смены видов (сукцессии) мышевидных грызунов, сильнее всего заметной в первые 15–20 лет вторичной сукцессии экосистемы на вырубках.
За весь период сукцессии экосистемы минимальные значения численности и видового разнообразия всех исследованных групп (землеройки-бурозубки и лесные полевки) зафиксированы в плотносомкнутых молодых древостоях (особенно хвойных) 25–45-летнего возраста. Во вторичных насаждениях 50–70-летнего возраста вновь наблюдается рост численности мелких млекопитающих. Для них отмечен существенный рост видового разнообразия, но уже не за счет мышевидных грызунов, а за счет землероек (рис. 3).
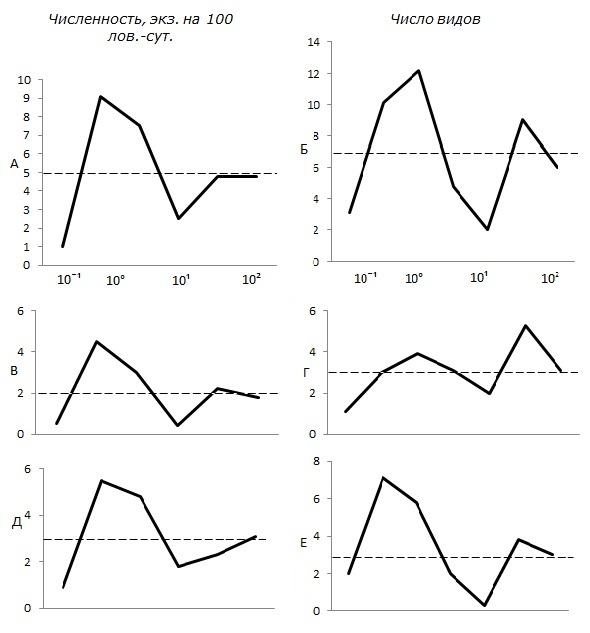
Рис. 3. Изменение численности и видового разнообразия мелких млекопитающих в процессе восстановления экосистем после рубки сосняков. а, б – все виды, в, г – землеройки, д, е – мышевидные грызуны. Давность рубки дана в системе десятичных логарифмов
Fig. 3. Changes in the number and species diversity of small mammals in the process of restoration of ecosystems after felling of pine forests. a, b – all species, c, d – shrews, d, e – mouse-shaped rodents. The age of felling is given in the system of decimal logarithms
Результаты исследований показывают, что непосредственно после рубки численность и число видов мелких млекопитающих резко сокращаются (на свежих вырубках отлавливались единичные рыжие полевки, но и те ближе к периферии лесосек и возле недорубов). Однако уже со второго года численность и видовое разнообразие зверьков существенно возрастали, достигая через 3–4 года после рубки значительно более высоких, чем до проведения рубок, значений. Иногда в качестве факторов, дополнительно влияющих на восстановление населения животных, упоминались также способ и степень очистки лесосек (Керзина, 1956; Курхинен и др., 2006; Gashwiller, 1970).
Как отмечено выше, после сплошной рубки на лесосеках практически не остается постоянного населения млекопитающих. Возможно лишь посещение и временная концентрация представителей растительноядных видов возле куч порубочных остатков.
Хотя в первые месяцы после рубки зверьки на открытых участках отсутствовали, однако их численность в небольшом по площади недорубе (1000 м2) составляла ~25 экз. на 100 лов./сут. В основном это были рыжие полевки, численность которых здесь оставалась высокой даже на второй год после рубки. Однако на третий год (к весне) ситуация резко изменилась: рыжая полевка исчезла, ее место заняли темная полевка и экономка, численность которых была меньше таковой на окружающих недорубы лесосеках.
На фоне недорубов особое значение приобретают семенные куртины хвойных пород. В ходе стационарных исследований зафиксировано четкое отличие структуры биоценотических группировок мелких млекопитающих в крупных семенных куртинах сосны от таковой на окружающих куртины вырубках. В частности, доля лесных видов (средняя бурозубка, рыжая полевка) оказалась здесь выше, чем на окружающих вырубках. Более того, на территории одной из крупных семенных куртин (объекта наших постоянных наблюдений) зафиксировано немногочисленное, но, по-видимому, постоянное поселение лесного лемминга – типично таежного, сибирского вида (Ивантер, 1975; Сиивонен, 1979).
Немаловажную роль играет скорость восстановления растительности, которая, в свою очередь, зависит от гидрологического режима, рельефа, плодородия почв. При глубоких нарушениях напочвенного покрова, огневой очистке лесосек, их заболачивании на фоне бедных непродуктивных почв повторное заселение вырубок может значительно замедляться.
Как следует из наблюдений на стационарах, основные источники иммиграции мелких млекопитающих – массивы невырубленных суходольных и пойменных (водоохранных) лесов. Мелкие же недорубы в основном имеют значение лишь как объекты первичного заселения, но отнюдь не как источники иммиграции.
На открытых (необлесившихся) вырубках, с давностью рубки от 1 до 7 лет, формируется травяно-кустарничковый ярус растительности, который может достигать значительного развития, особенно на злаковых вырубках. Часто наблюдается активное плодоношение ягодных кустарничков, особенно на микроповышениях и периферии вырубок. Открытые вырубки характеризуются специфичным температурным режимом и влажностью приземных слоев воздуха, спецификой снежного покрова. Создаются в целом благоприятные, хотя и менее стабильные, чем в хвойных лесах, условия среды обитания мелких млекопитающих, численность и видовое разнообразие которых здесь выше, чем под пологом спелых хвойных лесов (табл. 3).
Таблица 3. Численность и соотношение видов мелких млекопитающих на вырубках различного типа
| Показатель | Тип вырубки | ||
| вейниково-луговиковая | луговиково-кустарничковая | осоково-сфагновая | |
| Общая численность | 12.7/6.6 | 8.4/– | 4.3/2.0 |
| Обыкновенная бурозубка | 2.9 (23)/3.7(56) | 2.2(26)/– | 1.2(23)/0.4(20) |
| Средняя бурозубка | 0(0)/0.2(3) | 0(0)/– | 0.1(2)/0(0) |
| Малая бурозубка | 0.2(2)/0.4(7) | 0(0)/– | 0.2(5)/0.3(15) |
| Рыжая полевка | 1.6(12)/0.3(4) | 2.4(29)/– | 0.8(19)/0.2(10) |
| Темная полевка | 7.6(1)/1.9(29) | 3.7(44)/– | 2.0(46)/1.1(50) |
| Обыкновенная полевка | 0.07(1)/0(0) | 0(0)/– | 0(0)/0(0) |
| Лесная мышовка | 0.07(1)/0.1(1) | 0.08(1)/– | 0(0)/0(0) |
| Лесной лемминг | 0(0)/0(0) | 0(0)/– | 0(0)/0.1(1) |
Примечание. В числителе – экз. на 100 ловушко-суток, в знаменателе – экз. на 10 канавко-суток, в скобках – индекс доминирования, % от улова. Прочерк – учеты не проводились.
Численность мелких млекопитающих на открытых и зарастающих вырубках в целом существенно выше таковой в спелых хвойных насаждениях и превышает средние по биотопам значения. Наиболее сильные изменения численности в первые 10 лет после рубки характерны для популяций двух основных видов полевок: рыжей и темной, составляющих основу уловов мышевидных грызунов на злаковых вырубках. В итоге удалось проследить весь процесс трансформации биоценотических группировок мелких млекопитающих в течение первого десятилетия после рубки, т. е. в период наиболее сильных изменений. В течение первых 7–8 лет основной фон населения вырубки составляла темная полевка, которая наряду с обыкновенной бурозубкой определяла и характер волн флуктуации численности зверьков. В процессе восстановления лесной растительности происходила постепенная сдача позиций темной полевкой и их завоевание рыжей. В дальнейшем восстановление лесной среды в процессе вторичной сукцессии приводило к ухудшению экологических условий для темной полевки. В результате преимущество вновь получает рыжая полевка, которая лидировала по обилию и доле участия в уловах мышевидных грызунов.
Процессы аналогичного характера (на начальном этапе – рост численности серых полевок, а затем спад и завоевание доминирующих позиций лесными полевками рода Clethrionomys) были зафиксированы и в ряде других регионов таежного севера Европейской России (Ельшин, Каратаев, 1988; Турьева, 1956), а также в Скандинавии (Henttonen et al., 1977). Однако есть и особенности, в том числе связанные с различными отношениями рыжей и темной полевок к опушкам. В итоге создается впечатление, что первая из них активно мигрирует на вырубку, а часть населения второй вытесняется на периферию вырубки конкурентом, т. е. полевкой-экономкой, обилие которой на лесосеке в несколько раз выше среднего.
Выявленные различия в видовом составе и соотношении населяющих различные биотопы видов становятся еще очевиднее при сопоставлении средней бурозубки, рыжей и темной полевок. Для последних двух видов соотношение обилия в сравниваемых местообитаниях прямо противоположно. Так, рыжая полевка абсолютно доминирует в уловах мышевидных грызунов в сосняке (88 %), темная полевка – на вырубке (97 %). Установлено также, что структуру населения млекопитающих на ранних стадиях сукцессии леса определяет и тип вырубки.
В наших исследованиях наиболее полно представлены три типа вырубок, население которых сравнивали на протяжении 11 лет в ходе массовых учетов мелких млекопитающих, проводимых одновременно в течение трех сезонов (весна, лето, осень) и в равных объемах. В итоге установлено, что все три типа вырубок существенно различаются между собой по численности мелких млекопитающих, ее динамике и соотношению видов. Так, численность мелких млекопитающих на вейниково-луговиковой вырубке выше, чем на луговиково-кустарничковой и осоково-сфагновой соответственно в 1.5 и 3 раза. При этом во всех трех случаях доминирует темная полевка, однако доля участия этого вида в уловах, как и других видов, резко меняется в зависимости от типа вырубки.
Обращает на себя внимание тот факт, что на фоне сокращения спектра видов (по сравнению со злаковой вырубкой) в биоценотических группировках заболоченных лесосек в 1.5–2 раза выше доля участия малой бурозубки, причем в отдельные сырые годы этот вид здесь даже доминировал.
В итоге полученные нами данные свидетельствуют о специфике биоценотических группировок вырубок разных типов в период их активного формирования, в частности в первые 6–8 лет после рубки. Более того, формирование характерных фитоценозов на вырубках разных типов определяет специфику биоценотических группировок млекопитающих и на этих вырубках, и на участках формирующихся здесь молодняков. Иными словами, еще задолго до рубки за счет своеобразия типа вырубаемого леса задаются и особенности сукцессии биоценотических группировок мелких млекопитающих.
Отчетлива и смена доминантов: если в коренном сосновом лесу постоянно численно доминирует рыжая полевка, то на вырубке преобладают представители рода серых полевок (темная или экономка). Между тем все вырубки Южной Карелии, где в отловах грызунов доминировала рыжая полевка, появились на месте рубок ельников (или соседствовали с ними). Характерно, что все они были захламлены и слабо очищены от порубочных остатков, тогда как злаковые вырубки, где доминировала, например, темная полевка, были хорошо очищены. Именно эти факторы повлияли и на видовой состав полевок (рис. 4). Злаковые же вырубки нормально функционируют не в качестве временных, а как своего рода резервные стации мышевидных грызунов. Высокая численность последних (особенно представителей рода Microtus) обеспечивает дополнительную гарантию выживания популяций как хищников (миофаги), так и их жертв (полевки) в условиях нестабильного климата таежной зоны Восточной Фенноскандии. Последнее положение подтверждается и тем, что в отдельные нестандартные по погодным условиям годы мы фиксировали довольно высокую зимнюю численность полевок именно на вырубках (при почти нулевой численности в окружающих лесах).
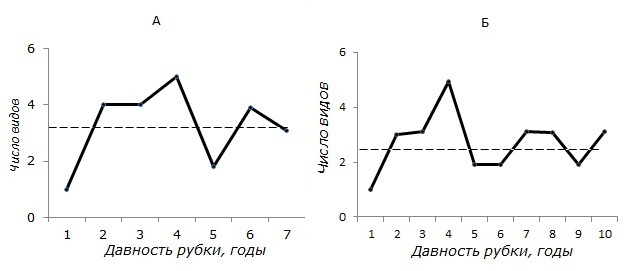
Рис. 4. Изменение числа видов мелких млекопитающих на злаковых вырубках с очисткой путем сбора и оставления остатков на месте (А) и путем вывоза и частичного сжигания порубочных остатков (Б). Штриховые линии – средние значения
Fig. 4. Change in the number of small mammal species on grass cutting with cleaning by collecting and leaving residues in place (a) and by exporting and partially decommissioning the felling residues (b). The dashed lines are the mean values
Есть и промежуточные варианты, например, сравнительно небольшие изменения, касающиеся конкретных соотношений видов при сохранении основных доминантов. Тем не менее в рамках этого широкого спектра можно выделить ряд общих закономерностей. При сплошной рубке высокопроизводительных сосновых лесов в средней и южной тайге Восточной Фенноскандии, особенно сосняков черничных и чернично-травяных, а также сосново-еловых лесов, сопровождающейся формированием злаковых вырубок с мощным травянистым ярусом, наблюдается отчетливая смена видов мышевидных грызунов. Она характеризуется сменой доминанта (рыжая полевка) в хвойном лесу на представителей рода Microtus на вырубке (темная полевка, полевка-экономка). Обычно доминирование нового вида сохраняется 6–8 лет после рубки, а в отдельных случаях при задержке лесовозобновления и дольше.
Мозаичность ландшафта, структура и состав соседствующих со свежей вырубкой биоценозов могут существенно влиять на последующие изменения в структуре группировок мелких млекопитающих. Как удалось установить в процессе стационарных исследований, наличие совершенно разных источников иммиграции полевок несомненно определяет как четкое доминирование темной полевки в одном случае, так и стойкое преобладание полевки-экономки в другом случае, когда расстояние между обоими объектами всего 4 км.
Лесохозяйственные мероприятия на вырубках существенно влияют на характер и скорость сукцессионных процессов, протекающих в биотопических группировках мелких млекопитающих. Оставление порубочных материалов на месте в кучах и валах, а также создание лесных культур хвойных пород обеспечило более заметное участие рыжей полевки в формировании многовидового населения и сократило период доминирования представителя рода Microtus. Формирование же мощного травяного покрова на хорошо очищенной злаковой вырубке (Восточная) определило полное и довольно длительное доминирование представителя серых полевок. В то же время в группе землероек после рубки леса в абсолютном большинстве случаев мы констатируем лишь некоторое перераспределение доли участия видов (например, некоторое увеличение этого показателя для малой бурозубки).
Таким образом, на сплошных открытых (необлесившихся) вырубках наблюдается общее увеличение численности мелких млекопитающих, а также перераспределение видов в пользу представителей открытых стаций (полевки рода Microtus). Вырубки очень динамичны по экологическим условиям, численности и видовому составу мелких млекопитающих. На структуру их населения влияют давность рубки, тип вырубки, конфигурация и площадь лесосек, породный состав вырубленного древостоя. Обычно вырубки характеризуются менее стабильным населением зверьков, демонстрирующих здесь более резкие колебания численности по годам и в течение сезона, по сравнению со спелыми сосняками-зеленомошниками, хотя средний показатель учета на вырубках выше. Это справедливо как для мелких млекопитающих в целом, так и для отдельных экологических групп (землеройки, мышевидные грызуны).
Заключение
Необходимо особо подчеркнуть, что неизбежно наступающий после промышленных рубок процесс трансформации и восстановления лесной биоты крайне сложен и неоднозначен. Он затрагивает все ее составляющие, в частности население мелких млекопитающих – землероек-бурозубок и лесных полевок. В результате сплошной рубки хвойных лесов происходят значительные изменения и в динамике численности, и в видовом разнообразии как всего населения мелких млекопитающих, так и отдельных их групп: землероек и мышевидных грызунов. Эти изменения особенно заметны впервые 5 лет после рубки. На лесосеках не только увеличивается общая численность мелких млекопитающих (прежде всего за счет мышевидных грызунов), но и меняется характер ее изменений: увеличиваются варьирование и частота колебаний в смежные годы, несбалансированность видовых группировок землероек и т. д. При некотором сокращении видового разнообразия землероек существенно и статистически достоверно возрастает видовое богатство мышевидных грызунов.
Библиография
Динесман Л. Г. Влияние диких млекопитающих на формирование древостоев . М.: Наука, 1961. 167 с.
Ельшин С. В., Каратаев А. Б. Сукцессии лесных млекопитающих на вырубках южной тайги // V съезд ВТО. М.: Наука, 1988. С. 275–276.
Ивантер Э. В. Популяционная экология мелких млекопитающих таежного Северо-Запада СССР . Л.: Наука, 1975. 244 с.
Калинин М. В. Экология охотничьих зверей и птиц сосновых вырубок: Автореф. дис. ... канд. биол. наук . Л.: Изд-во ЛГУ, 1973. 23 с.
Керзина М. Н. Влияние вырубок и гарей на формирование лесной фауны // Роль животных в жизни леса. М.: Изд-во МГУ, 1956. С. 21–30.
Курхинен Ю. П., Данилов Ю. П., Ивантер Э. В. Млекопитающие Восточной Фенноскандии в условиях антропогенной трансформации таежных экосистем . М.: Наука, 2006. 208 с.
Ларин Б. А. Влияние интенсивных рубок леса на продуктивность охотничьих угодий // Труды ВНИО. 1955. Вып. 14. С. 137–144.
Наумов С. П., Руковский Н. Н. Влияние рубки леса и его возобновления на полевок рода Clethrionomys и Microtus // Новые проблемы зоологической науки. Ставрополь, 1979. Ч. 2. С. 303–306.
Одум Ю. Экология . М.: Мир, 1986. 376 с.
Сиивонен Л. Млекопитающие Северной Европы . М.: Прогресс, 1979. 232 с.
Турьева В. В. Фауна мышевидных грызунов различных типов леса и ее изменения под влиянием вырубок // Труды Коми филиала АН СССР. 1956. Вып. 4. С. 112–115.
Численность и распределение мелких млекопитающих в Карелии в 1953–2010 гг.: свидетельство 2013621171 Российская Федерация / Э. В. Ивантер, Т. В. Ивантер, А. В. Кухарева, А. Е. Якимова; правообладатель ФГБУ науки ИБ КарНЦ РАН. № 2013620853; заявл. 19.07.13; дата рег. 17.09.13, опубл. Бюл. № 4 (86) (III ч.). 2013. С. 607–608.
Diffendorfer J. E., Gaunes M. S., Holt R. D. Habitat fragmentation and movements of three small mammals (Sigmo- don, Microtus and Peromyscus) // Ecology. 1978. Vol. 76. № 3. P. 827–829.
Gashwiller J. S. Plant and mammal changes on a clearcut in West-Central Oregon // 1970. Vol. 51. № 6. P. 1018–1026.
Hansson Z. Landscape ecology of boreal forest // Trend Ecol. Evol. 1992. Vol. 7. P. 299–302.
Henttonen H., Kaikusalo A., Tast J., Viitala J. Interspecific competition between small rodents in Subarctic and boreal ecosystems // Oikos. 1977. Vol. 29. P. 581–590.
Keith J. S., Smith D. J., Morris J. K. Dynamics of snowshoe hare population in fragmented habitat // J. Zool. 1993. Vol. 71. P. 1385–1392.
Lindzey A. Response of the white-footed mouse (Peromy- scus leocopus) to the transition between disturbed and undisturbed habitats // J. Zool. 1989. Vol. 67. № 2. P. 505–512.

 © 2011 - 2026
© 2011 - 2026