



Введение
Цикличность в популяциях живых организмов стала предметом изучения еще в начале прошлого века, и интенсивность исследований в этом направлении в последнее время быстро растет. Громадный эмпирический материал по оценке многолетней динамики численности животных теоретически обобщается; особенности цикличности у отдельных видов и таксономических групп прорабатываются все более детально (Максимов, 1984; Кривенко, 2005; Белецкий, 2007). Так, выяснено, что механизм многолетних колебаний численности популяций животных имеет существенную эндогенную компоненту (Кшнясев, Давыдова, 2005).
В работах, посвященных изучению динамики численности лесных полевок, предлагаются разнообразные математические подходы к анализу цикличности этих видов (Чернявский, Лазуткин, 2004; Жигальский, 2011; Ивантер, Моисеева, 2015; Ердаков, Моролдоев, 2017; Boonstra, Krebs, 2012). Среди видов лесных полевок, обитающих в России, наименее изучена многолетняя динамика численности красно-серой полевки (Myodes (=Craseomys) rufocanus). Этот вид имеет обширный ареал, западная граница которого располагается в Фенноскандии, а восточная – от побережья Охотского моря до Японских островов. Исследования многолетней цикличности M. rufocanus проводились на территории Фенноскандии (Hansson, Henttonen, 1985, 1988; Stenseth, 1985; Stenseth et al., 1996; Окулова, Катаев, 2003; Ивантер, Моисеева, 2015). В некоторых исследованиях цикличность связывают с воздействием глобальных климатических циклов, в частности с Североатлантическим колебанием, характерным для Скандинавии (Hornfeldt et al., 2005). Наиболее часто существование цикличности объясняется трофическими взаимодействиями видов (Hanski et al., 1993; Hanski, Korpimaki, 1995; Selås, 1997; Stenseth et al., 1996; Gilg et al., 2003). Нередки объяснения цикличности и динамикой местных климатических условий, суровостью зимы и глубиной снежного покрова (Hornfeldt et al., 2005; Bierman et al., 2006). При исследовании любой экологической характеристики необходимо предварительное накопление сведений о ее проявлениях. Для изучения цикличности динамики численности этот период начинается, появляются сведения об изменчивости цикличности у видов и популяций в различных участках ареалов.
Целью нашего исследования является описание цикличности динамики численности географических популяций красно-серой полевки (Myodes rufocanus) в различных частях ее ареала.
Задачи сводились к:
– построению спектров ритмов многолетней динамики численности различных географических популяций красно-серой полевки;
– определению параметров многолетних ритмов численности M. rufocanus;
– выявлению особенностей цикличности и возможных географических закономерностей ее изменений;
– описанию инвариантных частей спектра ритмов численности;
– поиску близких по значению природных местных циклов, к которым возможна подстройка популяционных ритмов численности M. rufocanus.
Материалы
Для описания многолетней цикличности популяций красно-серой полевки в различных частях ареала нами использованы опубликованные данные по следующим территориям: Северная Швеция, окр. г. Умео (Cornulier et al., 2013); Финляндия, национальный парк Паллас-Юллястунтури и исследовательская станция Хельсинкского университета Кильписъярви (Cornulier et al., 2013); Лапландский заповедник (Катаев, 2016); Средний Урал, Висимский заповедник (Лукьянова, 2013); Восточные Саяны, заповедник «Столбы» (Виноградов, Кельбешеков, 2009); Магаданский заповедник (Чернявский, Лазуткин, 2004); о-в Хоккайдо (Bjørnstad et al., 1998). В работе использованы наши данные о многолетней динамике численности M. rufocanus в Баргузинском заповеднике (Восточное Прибайкалье), а также неопубликованные сведения В. В. Панова (ИСиЭЖ СО РАН) по уловам красно-серой полевки в окрестностях Новосибирска (1980–2014 гг.).
Методы
В работе при проведении счетных операций применяли программы спектрального анализа, находящиеся в собственности ИСиЭЖ СО РАН. Оценка спектральной плотности мощности (СПМ) проводилась методом Уэлча (Марпл, 1990). Все расчеты выполнены с использованием свободной системы для математических вычислений GNU Octave (GNU Octave, 2018), которая представляет интерактивный командный интерфейс для решения различных линейных и нелинейных математических задач. Для оценивания СПМ методом Уэлча использовалась функция pwelch из пакета расширений Octave-Forge (Octave-Forge, 2018). Для трендов проверяли гипотезу об отличии коэффициентов от нуля, был рассчитан коэффициент детерминации R2. Полученные значения отражаются в тексте только в том случае, если нулевая гипотеза о равенстве обоих коэффициентов регрессии нулю отбрасывается. В противном случае линейная регрессия исключается из модели процесса. В модели для ограничения числа гармоник использовали тест Льюнга – Бокса для автокорреляции остатков с заданным критерием. Этот тест применяется к остаткам после включения каждой составляющей, и дальнейшее добавление прекращается, как только тест покажет некоррелированность остатков.
Для статистической обработки использован пакет программ Past.
Результаты
Некоторые статистические характеристики динамики численности
Во всех районах, где были проведены наблюдения за динамикой численности красно-серой полевки, численность вида была средней или высокой. Вариабельность ее за сроки наблюдений была относительно невелика, исключение представляет популяция в заповеднике «Столбы», где при высокой численности изменения ее достигали двух порядков (табл. 1). Самая стабильная численность зафиксирована в центральной гористой области о-ва Хоккайдо, а также на исследовательской станции Хельсинкского университета Кильписъярви. В обоих случаях наблюдения были очень продолжительными.
Таблица 1. Статистические показатели численности в популяциях красно-серой полевки
| Территория | n | Численность, M ± m | σ | CV | Тренд |
| Швеция, окр. г. Умео | 39 | 4.021 ± 0.56 | 3.51 | 87.29 | 7.037 - 0.159 * t; R2 = 0.266; α ≤ 0.01 |
| Финляндия, парк Паллас-Юллястунтури | 41 | 3.367 ± 0.289 | 1.855 | 55.09 | 4.078 - 0.036 * t |
| Финляндия, Кильписъярви | 43 | 3.99 ± 0.415 | 2.718 | 68.12 | 5.254 - 0.060 * t; α > 0.05 |
| Лапландский заповедник | 71 | 20.98 ± 2.607 | 21.88 | 104.29 | 20.489 + 0.014 * t; α > 0.05 |
| Урал, Висимский заповедник | 23 | 6.0 ± 1.39 | 6.64 | 3.864 + 0.194 * t; α > 0.05 | |
| Новосибирск, Академгородок | 35 | 2.51 ± 0.375 | 2.211 | 88.08 | 0.367 + 0.126 * t; R2 = 0.342; α ≤ 0.01 |
| Саяны, заповедник «Столбы» | 27 | 6.54 ± 2.225 | 11.56 | 176.76 | 10.320 - 0.430 * t; R2 = 0.145; α ≤ 0.01 |
| Баргузинский заповедник | 54 | 4.99 ± 0.61 | 4.49 | 89.98 | 7.992 - 0.113 * t; R2 = 0.158; α ≤ 0.01 |
| о-в Хоккайдо | 31 | 2.850 ± 0.267 | 1.486 | 52.14 | 1.771 - 0.010 * t; α > 0.05 |
| Магаданский заповедник | 21 | 9.78 ± 1.479 | 6.782 | 69.35 | 10.677 - 0.090 * t; α > 0.05 |
Примечание. n – число рассматриваемых лет, M – среднее, m – ошибка среднего, σ – квадратичное отклонение, CV – коэффициент вариации, R2 – коэффициент детерминации.
Близкое расположение районов наблюдений в Фенноскандии вынудило нас проверить, насколько синхронен здесь ход многолетней численности красно-серой полевки. Визуальное сравнение хронограмм не давало возможности предположить такую синхронность (рис. 1). Для сравнения были выбраны одинаковые отрезки ряда (1970–2010 гг.) и рассчитаны ранговые коэффициенты корреляции между ними. Статистических различий между многолетними средними численности в этих точках обнаружено не было. Однако на этом отрезке времени коррелятивная связь хода численности наблюдается между рядами из Швеции и Кольского полуострова (ƿ = 0.3703; α = 0.05).
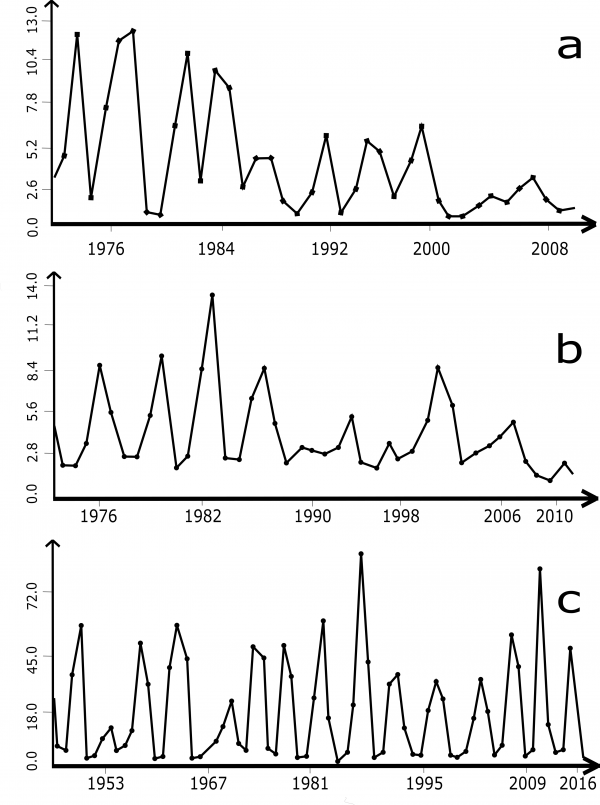
Рис. 1. Ход многолетней численности красно-серой полевки в Фенноскандии (а – Швеция, Умео; b - Финляндия, Pallasjärvi; с – Лапландский заповедник). По оси абсцисс указаны годы, по оси ординат – число особей на 100 ловушко-суток
Fig. 1. Long-term abundance of the grey red-backed vole in Fennoscandia (a – Sweden, Umea; b – Finland, Pallasjärvi; с – Lapland Reserve). The X-axis indicates years, the Y-axis indicates the number of individuals per 100 trap-days.
Спектральные оценки хода численности
Информацию о цикличности численности дают спектры ее ритмов, полученные при перенесении данных о ходе численности с временной шкалы на частотную. При этом мы получаем спектр циклов, скрытых в сложной кривой многолетних изменений численности, однако теряем сведения о самой динамике количества зверьков.
Представив цикличность многолетнего хода плотности популяции красно-серой полевки в Фенноскандии в виде спектров из трех районов, мы получили набор ритмов численности в каждом (рис. 2) и можем сравнить их между собой.
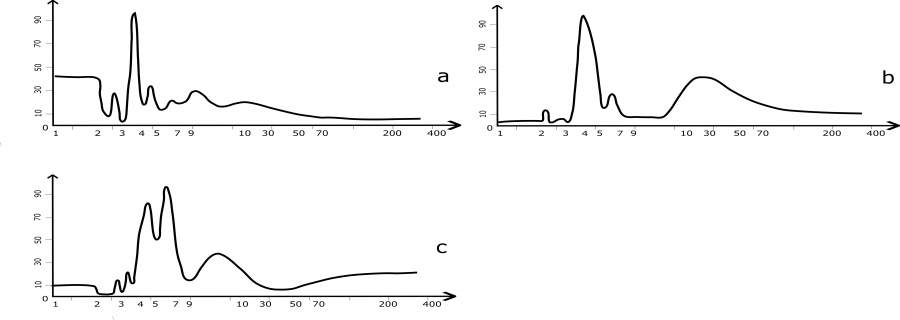
Рис. 2. Спектры многолетней цикличности численности красно-серой полевки в Фенноскандии (а – Швеция, Умео; b –Финляндия, Pallasjärvi; с – Лапландский заповедник). По оси абсцисс указаны периоды (лет), по оси ординат – мощность (в %)
Fig. 2. Spectra of long-term cyclicity of the grey red-backed vole in Fennoscandia (a – Sweden, Umea; b – Finland, Pallasjärvi; с – Lapland Reserve). The X-axis indicates the periods (years), the Y-axis shows the power (%)
В центральной части ареала длительные наблюдения за ходом численности M. rufocanus проведены на Урале, в Западной Сибири и в Красноярском крае. Рассчитанные спектры цикличности этих географических популяций представлены на рис. 3.
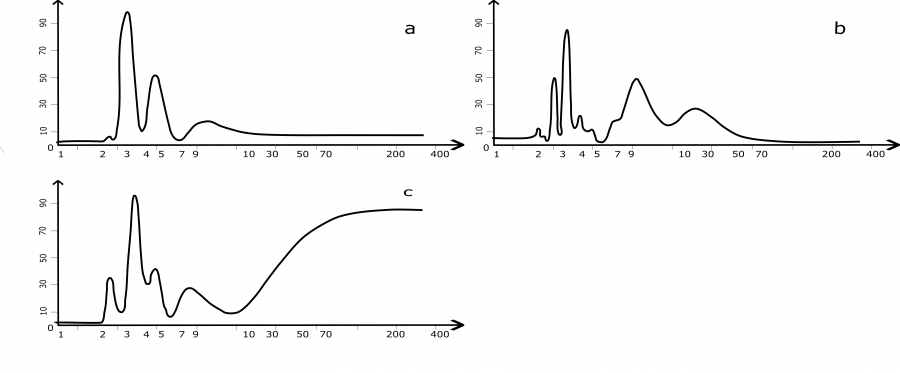
Рис. 3. Спектры многолетней цикличности численности красно-серой полевки в центральных частях ареала (а – Средний Урал; b – Новосибирск, Академгородок; с – Красноярский край, заповедник «Столбы»). По оси абсцисс указаны периоды (лет), по оси ординат – мощность (в %)
Fig. 3. Spectra of long-term cyclicity of the grey red-backed vole in the central parts of the area (a – Middle Urals; b – Novosibirsk, Akademgorodok; с – Krasnoyarsk Region, "Stolby" Reserve). The X-axis indicates the periods (years), the Y-axis shows the power (%)
В Восточном Прибайкалье ряд, использованный для поиска циклов в динамике плотности красно-серой полевки, был достаточно представительный (см. табл. 1).
На восточной границе ареала цикличность M. rufocanus изучалась как на севере, в Магаданском заповеднике, так и гораздо южнее, на о-ве Хоккайдо (Япония). В обоих случаях для выявления ритмов динамики численности применяли спектральный анализ. Мы для расчетов использовали временной ряд в 16 лет из Магаданского заповедника и самый длинный ряд, приведенный из центральной части о-ва Хоккайдо. Полученные спектры можно визуально сравнить между собой (рис. 4, b и c).
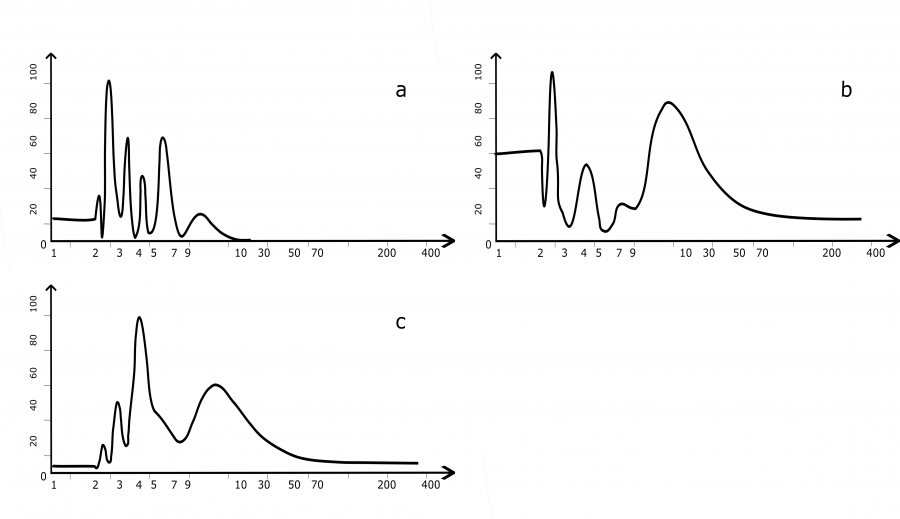
Рис. 4. Спектры многолетней цикличности численности красно-серой полевки в восточной части ареала (а – Баргузинский заповедник; b – Магаданский заповедник; с – о. Хоккайдо). По оси абсцисс указаны периоды (лет), по оси ординат – мощность (в %)
Fig. 4. Spectra of long-term cyclicity of the grey red-backed vole in the eastern parts of the range (a – Barguzinsky Reserve; b – Magadan Reserve; с – Hokkaido Island). The X-axis indicates the periods (years), the Y-axis shows the power (%)
Более детальное сравнение цикличностей динамики численности в разных частях ареала красно-серой полевки можно сделать, используя данные о параметрах ее циклов (табл. 2).
Таблица 2. Соотношение величин и мощностей гармонических составляющих на спектрах цикличности многолетней динамики численности красно-серой полевки в различных частях ареала
| Период, лет | 15–28 | 9.5–13 | 6.4–9 | 4.6–6.3 | 3.5–4.5 | 2.8–3.4 | 2.7–3.3 | 2.0–2.6 |
| Швеция, окр. г. Умео | 20.5 | 9.6 | 6.4 | 4.6 | 3.5 | – | 2.6 | 2.2 |
| 0.50 | 0.63 | 0.53 | 0.68 | 1.18 | 0.61 | 0.41 | ||
| Финляндия, парк Паллас-Юллястунтури | 26.3 | 11.3 | 8.4 | 6.2 | 4.0 | 2.9 | 2.5 | 2.2 |
| 0.63 | 0.27 | 0.30 | 0.51 | 0.95 | 0.24 | 0.11 | 0.36 | |
| Финляндия, Кильписъярви | 22.3 | – | – | 6.1 | 4.6 | 3.0 | – | – |
| 0.59 | 0.50 | 0.62 | 0.30 | |||||
| Лапландский заповедник | – | 14.0 | 6.4 | 4.7 | – | 3.4 | 2.9 | 2.3 |
| 27.5 | 44.30 | 40.8 | 20.93 | 17.0 | 6.16 | |||
| Североатлантическое колебание, весна | 23.2 | 11.4 | 6.7 | 5.2 | 4.2 | 3.5 | 2.7 | 2.3 |
| 114 | 48 | 104 | 179 | 175 | 151 | 282 | 169 | |
| Средний Урал | – | 9.1 | – | – | 3.8 | – | 2.8 | – |
| 1.44 | 1.58 | 1.68 | ||||||
| Академгородок, Новосибирск | – | 8.3 | – | 5.5 | 3.3 | – | – | – |
| 127 | 256 | 600 | ||||||
| Саяны, заповедник «Столбы» | – | 8.4 | – | 4.9 | 3.5 | – | – | 2.4 |
| 1.19 | 1.44 | 2.17 | 1.35 | |||||
| Баргузинский заповедник | – | 15.5 | 7.4 | – | 4.2 | – | – | 2.4 |
| 1.20 | 0.59 | 0.88 | 1.33 | |||||
| Магаданский заповедник | – | 10.5 | – | 5.9 | 4.2 | 3.3 | 2.5 | 2.1 |
| 1.24 | 2.40 | 1.94 | 2.43 | 2.99 | 1.60 | |||
| о. Хоккайдо | – | 13.5 | – | – | 4.1 | – | 2.9 | 2.3 |
| 0.32 | 0.43 | 0.28 | 0.17 | |||||
| Южное колебание,
весна |
24.4 | 10.5 | 6.5 | 5.0 | 3.9 | 3.4 | 2.9 | 2.3 |
| 0.06 | 0.08 | 0.11 | 0.09 | 0.15 | 0.13 | 0.19 | 0.06 |
Примечание. Верхняя цифра – период, лет; нижняя – мощность (амплитуда).
Обсуждение
Во всех изученных районах Фенноскандии численность имела тенденцию к постепенному снижению; заметнее всего такое снижение фиксируется в Швеции, где проявляется значимый отрицательный тренд (см. табл. 1). В целом постепенное изменение численности заметно в точках наблюдений из центральной части ареала вида. В окрестностях г. Новосибирска и в районе заповедника «Столбы» наблюдается достоверный ее тренд, в первом районе – положительный, во втором – отрицательный.
Хронограммы изменений в любой из наблюдаемых точек всегда представляют собой кривую неправильной формы. В горных местностях вариабельность увеличивается, крайнее ее значение отмечено на Саянах (см. табл. 1). Обычно же колебания невелики (в пределах 50–80 %). Приблизительные оценки цикличности по расстояниям между пиками можно привести, используя хронограммы из Фенноскандии (см. рис. 1). На всех хронограммах имеются 2–3-летние и 5–7-летние колебания. В Финляндии можно ожидать мощное ≈20-летнее колебание численности (см. рис. 1b).
Более адекватную информацию о популяционных колебаниях можно получить, познакомившись со всем их спектром в каждой географической популяции. Начнем рассмотрение с популяций в Фенноскандии (см. рис. 2). Визуально все спектры различаются. Хотя периоды циклов в основном совпадают на всех трех спектрах, мощности их распределяются по-разному. На спектрах из Финляндии и Швеции наблюдается схожее распределение мощностей гармоник. Это заметно как в высоких частотах, где они доминируют, так и в низких, где они невелики (см. рис. 2, a и b). Наиболее мощными ритмами в Финляндии и в Лапландском заповеднике являются ≈4.5-годовые, а вот в Швеции превалирует по мощности 3-летний цикл. В Лапландии доминирует по мощности ≈7-летний ритм (см. рис. 2с).
Важным отличием спектра из Финляндии от шведского и лапландского служит присутствие на нем мощной низкочастотной составляющей. Здесь есть многолетний ≈15–25-летний цикл. Об этом говорит наличие пиков в этой области, хотя широкие основания их не дают возможности точного определения периодов. На спектре ритмов из Швеции в этой области, возможно, имеется низкочастотная составляющая, большая по периоду, но незначительная по мощности (см. рис. 2a). На Кольском полуострове же, несмотря на длинный ряд наблюдений, низкочастотная компонента спектра не проявилась (см. рис. 2с).
Особенно длинный, 70-летний, ряд данных численности красно-серой полевки известен из Лапландского биосферного заповедника. Имеется словесное описание ряда, где упоминается только одна цикличность, ≈4–5-летняя. Ее определили как среднюю между пиками (Катаев, 2016). Эти данные взяты всего в 500 км от Кильписъярви, и авторы, сравнивая динамику полевок в этих точках, обнаружили, что все всплески численности в районе Кильписъярви совпадали с подъемами численности в Лапландском заповеднике (Кашулин и др., 2004). Расчет полного спектра этого ряда проясняет ситуацию с низкочастотными гармоническими составляющими хода численности красно-серой полевки. В Лапландии самый длинный ритм всего 15–17-летний (см. рис. 2с).
В центральных частях ареала красно-серой полевки изображения спектров колебаний численности близки к западным, но имеются и некоторые отличия (см. рис. 3). Даже при беглом их сравнении заметно, что доминирующие по мощности циклы одинаковы, различия в мощности у 8–10-летнего ритма могут характеризовать изменения в этих спектрах. При продвижении на восток мощность 9-летнего цикла постепенно снижается. Тем не менее в полосе высоких частот у всех популяций наблюдаются обычные для красно-серой полевки циклы в 3–4 года, а мощности гармонических составляющих в сибирских популяциях – самые большие. В средних частотах имеются 5–7-летние и заметны 9–12-летние циклы (см. рис. 3). В Висимском заповеднике (Средний Урал) с помощью спектрального анализа в динамике красно-серой полевки найдены мощные гармоники в 3-летней полосе частот и небольшие – в годичной (Кшнясев, Давыдова, 2005). Более длинный ряд, полученный из наблюдений за численностью красно-серой полевки этой же территории, приведен в работе Л. Е. Лукьяновой (2013). Его обработка позволила выявить больше гармонических составляющих в этом же заповеднике (см. рис. 3а). Долгосрочные наблюдения в районе Академгородка (Новосибирск) дали возможность уточнить низкочастотную составляющую спектра (≈25-летняя) (см. рис. 3b). Спектр ритмов из заповедника «Столбы», кроме обычных для динамики этих полевок пиков, может иметь мощную гармоническую составляющую в области низких частот (см. рис. 3c). Этот многолетний ритм пока не удается определить из-за небольшой длины наблюдений. По-видимому, он ≈50–60-летний, то есть длина всего ряда наблюдений включает только треть его синусоиды.
На востоке ареала вид спектров цикличности у красно-серой полевки имеет свои особенности (см. рис. 4). Так, в Восточном Прибайкалье на нем доминирует по мощности 2,5-летняя цикличность (см. рис. 4а). Кроме нее визуально выявляются два близких по мощности колебания: 3.5- и ≈7-летнее. Имеются и незначительные по мощности 4.5, 2.5 и ≈10-летние циклы.
В Магаданском заповеднике продолжительность наблюдений была невелика, однако позволила получить изображение спектра циклов численности с мощным ≈2.5-летним пиком, значительным по мощности ≈4-летним, а также средним по мощности, но с очень широким основанием ≈6.5-летним пиками (Чернявский, Лазуткин, 2004).
Особенно детально исследовали множество популяций Сr. rufocanus в Японии, на о-ве Хоккайдо, где были собраны сведения по долгосрочным (23–31-летним) наблюдениям в 225 точках. Данные эти были не только различной длины, но и разного качества, часть из них была получена в результате опросов. Это заставило авторов быть осторожными при описании и остановиться на уверенной фиксации у этого вида полевок только 2–2.5 и 3.5–4.5-летних циклов (Bjørnstad et al., 1998). По соотношению циклов дальневосточные популяции похожи, но различие им придает неодинаковость мощностей одних и тех же гармонических составляющих. Так, самой мощной в Магаданском заповеднике гармоникой является ≈3-летняя. Она присутствует и на спектре полевок из Японии, но значительно уступает доминирующей (≈4.5-летней). В свою очередь, 4.5-летняя цикличность имеется и на магаданском спектре, но по мощности она весьма незначительна, на Хоккайдо же она доминирует в спектре.
Заметное отличие спектров дают их среднечастотные области. Восьмилетний цикл, характерный для Баргузинского заповедника, и шестилетний цикл в Магаданском заповеднике совершенно не фиксируются в Японии. В низких частотах все спектры имеют сходство. На севере, под Магаданом, проявлен ≈10-летний пик, присутствующий и в Японии. Это же колебание, хотя и маломощное, наблюдается в Баргузинском заповеднике.
Рассмотрим цикличность по полосам частот (см. табл. 2). Наибольший по длительности цикл динамики численности фиксируется в Финляндии, однако точность его определения невелика, пик имеет широкое основание и сглаженную вершину. Для его уточнения нужен, по крайней мере, втрое длиннее ряд наблюдений. Низкочастотные циклы (20–25-летние) фиксируются только на Европейском Севере; даже в Лапландии, несмотря на долгосрочные наблюдения, такого колебания обнаружено не было. На Дальнем Востоке сходные колебания плотности красно-серой полевки обнаружить также пока не удалось. На хронограммах такие циклы умещаются не более одного раза, так что статистика здесь невелика, и более корректно говорить о существовании в многолетней динамике численности красно-серой полевки цикла в 9–15-летней полосе частот. Это обычный средний по мощности ритм, характерный для всех изучаемых популяций, и только в Баргузинском заповеднике у него оказалась значительная мощность (см. табл. 2).
В большинстве районов наблюдений у динамики численности красно-серой полевки фиксируются 9–12-летние циклы, их можно считать обычными для этого вида животных. Для всех же рассмотренных популяций этих полевок характерны (как и для большинства мышевидных грызунов) высокочастотные колебания многолетней численности в диапазоне от 2 до 9 лет. В этих полосах частот наблюдаются и самые высокие мощности гармонических составляющих, хотя в различных районах это могут быть разные по значению циклы. Так, и в североевропейских, и в японских популяциях доминируют по мощности 4–6-летние ритмы. У сибирских и дальневосточных популяций наиболее мощные циклы оказываются в высокочастотной области спектра, это 2–3.5-летние гармонические составляющие (см. табл. 2).
Устойчивость у популяционных ритмов обеспечивается синхронизацией к близким по значению природно-климатическим колебаниям в их местообитаниях. В столь географически удаленных районах, как Фенноскандия, Красноярский край и о. Хоккайдо, естественно ожидать различия в периодах ритмов именно по этим причинам.
Североевропейские популяции располагаются в зоне действия Североатлантического колебания (САК). Согласно многочисленным исследованиям, это колебание в значительной степени определяет погоду (интенсивность и траектории циклонов и штормов, аномалии осадков и приземной температуры воздуха) в большинстве стран Европы. Эта планетарная цикличность может быть внешним датчиком времени для близких по периоду ритмов в популяциях животных. При сопоставлении циклов САК и популяционных циклов в Фенноскандии можно заметить, что синхронизация возможна практически у любого популяционного ритма к соответствующей гармонической составляющей этого погодного индекса (см. табл. 2).
Для популяций на о. Хоккайдо датчиком времени для цикличности во всех полосах частот могут быть особенности местного климата, основой которому служит Южное колебание (SOI) (Bjørnstad et al., 1998). Однако с этой глобальной осцилляцией наблюдается значительное совпадение у популяционных ритмов только в высоких и средних частотах (см. табл. 2).
Повсеместно имеются природные циклы, соответствующие по периоду популяционным ритмам динамики численности красно-серой полевки. Так, в ходе температуры проявляются циклы продолжительностью 2–4, 5–6, 7–9 и 10–12 лет, а также полувековой и вековой (Коротина, 2002). Столь же широко распространены гидроклиматические циклы в 3–4 и 7–11 лет (Дружинин, 1987). Последние могут служить еще и датчиками времени для низкочастотных гармонических составляющих спектра ритмов полевок (см. табл. 2).
Заключение
У различных географических популяций красно-серой полевки на севере, западе, в центре и на востоке ареала динамика численности обладает определенной цикличностью.
Спектры многолетней цикличности динамики численности, построенные для десяти географических популяций этого грызуна, позволяют получить визуальное представление обо всем наборе популяционных циклов, а также провести сравнение периодов и мощностей гармонических составляющих.
Во всех рассмотренных популяциях проявлены циклы многолетней динамики в полосах высоких частот: 5–7, 3–4 и 2–3-летних. В средних частотах у многих популяций проявлены 9–14-летние циклы динамики численности, а на некоторых спектрах имеются и >20-летние ее периодические составляющие.
В центральных частях ареала у географических популяций проявлена тенденция роста мощности 9–10-летних гармонических составляющих численности с продвижением на восток. Кроме того, как и у большинства мелких грызунов, для этого вида характерны циклы динамики в диапазоне от двух до девяти лет.
Близкими по значению природными местными циклами, к которым возможна подстройка популяционных ритмов численности красно-серой полевки, являются циклические изменения погодных условий. Долгосрочное сохранение таких циклов в популяции дает возможность адаптации к изменениям местного климата. Кроме того, отмечена довольно точная подстройка у красно-серых полевок с о. Хоккайдо к высоким частотам глобальной южной осцилляции – планетарному циклу, который является основой погодных изменений региона.
Библиография
Белецкий Е. Н. Цикличность – фундаментальное свойство развития и функционирования природных систем // Вісник Харківського національного аграрного університету. Серія Біологія. 2007. Т. 3 (12). С. 100–116.
Виноградов В. Н., Кельбешеков Б. К. Структурно-временная организация сообщества грызунов среднегорной темнохвойной тайги Восточного Саяна (на примере заповедника «Столбы») // Вестник Томского государственного университета. Сер. Биология. 2009. Т. 4 (8). С. 5–14.
Дружинин И. П. Долгосрочный прогноз и информация . Новосибирск: Наука, 1987. 246 с.
Ердаков Л. Н., Моролдоев И. В. Изменчивость многолетней цикличности в динамике численности красной полевки (Myodes rutilus (Pallas, 1779)) // Принципы экологии. 2017. № 4. С. 26–36.
Жигальский О. А. Популяционные циклы рыжей полевки Myodes (Clethrionomys) glareolus: связь с репродуктивным процессом // Известия Самарского научного центра РАН. 2011. Т. 13. № 5. С. 185–191.
Ивантер Э. В., Моисеева Е. А. К экологии красной полевки (Clethrionomys rutilus Pall.) на юго-западной периферии ареала // Труды Карельского научного центра РАН. 2015. № 1. С. 37–47.
Катаев Г. Д. Долговременный (1936–2016 гг.) мониторинг видового состава и численности населения мелких млекопитающих северо-таежной Лапландии // Бюллетень Московского общества испытателей природы. Отд. Биол. 2016. Т. 121. Вып. 6. С. 3–17.
Кашулин П. А., Жиров В. К., Катаев Г. Д. Ритмы жизни на Кольском Севере: 70 лет наблюдений за флорой и фауной // Природопользование в Евро-Арктическом регионе: опыт XX века и перспективы. Апатиты: Изд-во Кольского НЦ, 2004. С. 175–183.
Коротина Е. Ф. Многолетние колебания температурного режима Южного Урала: Автореф. дис. ... канд. геогр. наук . Пермь: Перм. гос. ун-т, 2002. 23 с.
Кривенко В. Г. Концепция природной циклики и некоторые задачи хозяйственных стратегий России // Аграрная Россия. 2005. № 6. С. 41–47.
Кшнясев И. А., Давыдова Ю. А. Динамика плотности и структуры популяций лесных полевок в южной тайге // Вестник Нижегородского университета им. Н. И. Лобачевского. Сер. Биология. 2005. Т. 1 (9). С. 113–123.
Лукьянова Л. Е. Сопряженность симпатрических видов мелких млекопитающих в контрастных условиях среды // Экология. 2013. № 1. С. 65–72.
Максимов А. А. Многолетние колебания численности животных, их причины и прогноз . Новосибирск: Наука, 1984. 250 с.
Марпл С. Л. (мл). Цифровой спектральный анализ и его приложения . М.: Мир, 1990. 584 с.
Окулова Н. М., Катаев Г. Д. Многолетняя динамика численности красно-серой полевки (Clethrionomys rufocanus, Мicrotinae, Rodentia) в разных частях ареала // Зоологический журнал. 2003. Т. 82. № 9. С. 1095–1111.
Чернявский Ф. Б., Лазуткин А. Н. Циклы леммингов и полевок на Севере . Магадан: ИБПС ДВО РАН, 2004. 150 с.
Bierman S. M., Fairbairn J. P., Petty S. J., Elston D. A., Tidhar D., Lambin X. Changes over time in the spatiotemporal dynamics of cyclic populations of field voles (Microtus agrestis L.) // Am. Nat. 2006. Vol. 167. № 4. P. 583–590.
Bjørnstad O. N., Champely S., Stenseth N. C., Saitoh T. Cyclicity and Stability of Grey-Sided Voles, Clethrionomys rufocanus, of Hokkaido: Spectral and Principal Components Analyses // Philos. Trans. Biol. Sci. 1996. Vol. 351. P. 867–875.
Bjørnstad O. N., Stenseth N. C., Saitoh T., Lingjærde O. C. Mapping the regional transition to cyclicity in Clethrionomys rufocanus: Spectral densities and functional data analysis // Res. Popul. Ecol. (Kyoto). 1998. Vol. 40. № 1. P. 77–84.
Boonstra R., Krebs C. J. Population dynamics of red-backed voles (Myodes) in North America // Oecologia. 2012. Vol. 168. P. 601–620.
Cornulier T., Yoccoz N. G., Bretagnolle V., Brommer J. E., Butet A., Ecke F., Elston D. A., Framstad E., Henttonen H., Hörnfeldt B., Huitu O., Imholt C., Ims R. A., Jacob J., Jędrzejewska B., Millon A., Petty S. J., Pietiäinen H., Tkadlec E., Zub K., Lambin X. Europe-wide dampening of population cycles in keystone herbivores // Science. 2013. Vol. 340. № 6128. P. 63–66.
Gilg O., Hanski I., Sittler B. Cyclic dynamics in a simple vertebrate predator-prey community // Science. 2003. Vol. 302. № 5646. P. 866–868.
GNU Octave. URL: http://www.gnu.org/software/octave/ (дата обращения: 01.02.2018).
Hanski I., Turchin P., Korpimäki E., Henttonen H. Population oscillations of boreal rodents: regulation by mustelid predators leads to chaos // Nature. 1993. Vol. 364. № 6434. P. 232–235.
Hanski I., Korpimaki E. Microtine rodent dynamics in Northern Europe: Parameterized models for the predator-prey interaction // Ecology. 1995. Vol. 76. № 3. P. 840–850.
Hansson L., Henttonen H. Gradients in density variations of small rodents: the importance of latitude and snow cover // Oecologia. 1985. Vol. 67. № 3. P. 394–402.
Hansson L., Henttonen H. Rodent dynamics as community processes // Trends Ecol. Evol. 1988. Vol. 3. № 8. P. 195–200.
Hornfeldt B., Hipkiss T., Eklund U. Fading out of vole and predator cycles? // Proc. R. Soc. B Biol. Sci. 2005. Vol. 272. № 1576. P. 2045–2049.
Octave-Forge. URL: https://octave.sourceforge.io/ (дата обращения: 01.02.2018).
Selås V. Cyclic population fluctuations of herbivores as an effect of cyclic seed cropping of plants: The mast depression hypothesis // Oikos. 1997. Vol. 80. № 2. P. 257.
Stenseth N. C. Mathematical models of Microtine cycles: Models and the real world // Acta Zool. Fenn. 1985. Vol. 173. P. 7–12.
Stenseth N. C., Bjornstad O. N., Falck W. Is spacing behaviour coupled with predation causing the Microtine density cycle? A synthesis of current process-oriented and pattern-oriented studies // Proc. R. Soc. B Biol. Sci. 1996. Vol. 263. № 1376. P. 1423–1435.
Stenseth N. C., Bjørnstad O. N., Saitoh T. A gradient from stable to cyclic populations of Clethrionomys rufocanus in Hokkaido, Japan // Proceedings. Biol. Sci. 1996. Vol. 263. № 1374. P. 1117–1126.
Благодарности
Исследование поддержано РФФИ (грант № 17-04-00269), а также Программой фундаментальных научных исследований (ФНИ) государственных академий наук на 2013–2020 гг., проект № VI.51.1.9 (АААА-А16-116121410119-4).

 © 2011 - 2026
© 2011 - 2026