



Введение
Мелководная прибрежная зона формируется под влиянием водосбора и открытой части водного объекта. Она в наибольшей степени подвержена антропогенному воздействию и выполняет функцию барьера между водосборной территорией и пелагическими районами водоемов (Schlosser, Karr, 1981; Decamps et al., 2004; Чекрыжева, Калинкина, 2016). При исследовании прибрежных зон особое внимание отводится фитопланктону (Петрова, 1975; Вислянская, 1982; Протопопова, 2011; Дудакова и др., 2015; Чекрыжева, Калинкина, 2016; Nozaki, 2001; Nozaki et al., 2002; Taniguchi et al., 2005; Zębek, 2007; Turner et al., 2011; Asma' Jamal et al., 2014) как основному продуценту органического вещества и первичному звену в трофической цепи, наиболее чутко реагирующему на изменения условий обитания и объективно отражающему особенности структурно-функциональной организации гидробиоценозов (Трифонова, 1990). Участвуя в трансформации органических и минеральных веществ, поступающих с водосбора, водоросли выполняют основную их функцию в самоочищении вод, способствуя устойчивости водных экосистем (Соловьева, Корнева, 2012).
Данная работа проводится в рамках изучения организации литоральных сообществ в условиях изменяющегося климата и антропогенных воздействий, что определило цель проводимых исследований – изучение таксономической структуры, доминирующего комплекса, экологических характеристик и количественных показателей фитопланктона прибрежной и центральной частей оз. Арахлей.
Материалы
Озеро Арахлей входит в состав Ивано-Арахлейской территориально-аквальной системы, расположенной на юге Витимского плоскогорья (рис. 1а). Это самый крупный водоем системы. Площадь его зеркала 58.2 км2, объем водных масс 0.63 км3, максимальная глубина 17 м, средняя – 10.4 м (Ивано-Арахлейские озера…, 2013).
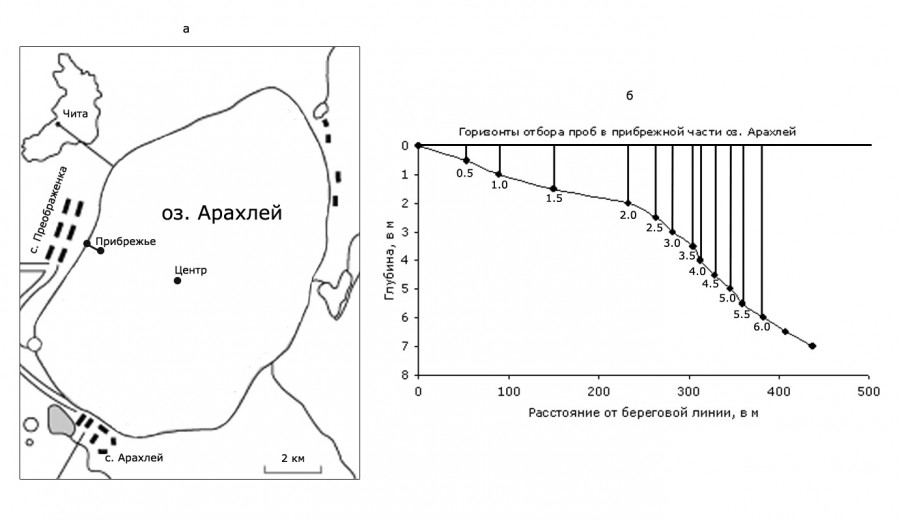
Рис. 1. Карта-схема отбора проб в оз. Арахлей: а – карта-схема оз. Арахлей, б – схема разреза и отбора проб в прибрежье озера
Fig. 1. Schematic map of sampling in Lake Arakhley: a – schematic map of Lake Arachley, b – scheme of cutting and sampling in the littoral part
Материалом для работы послужили результаты исследований планктонной флоры, проведенные в прибрежной и центральной частях оз. Арахлей в весенний (31.05–01.06) и летний (31.07–06.08) сезоны 2017 г.
Одновременно с отбором фитопланктонных проб проводилось исследование некоторых физико-химических параметров воды с использованием многопараметрического прибора контроля качества воды Aquareader. Результаты измерений приведены в табл. 1.
Таблица 1. Физико-химические параметры обследованных станций оз. Арахлей
| Глубина, м | T | EC | TDS | ORP | pH | Turb | Sal | |||||||
| 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | |
| ЗПВ* | 25.8 | 23.4 | 361 | 223 | 234 | 144 | 155 | 294 | 7.96 | - | 19.1 | 25.3 | 0.1 | 0.11 |
| 0.5* | 18.2 | 22.8 | 237 | 256 | 154 | 167 | 148 | 244.8 | 8.05 | - | 29.3 | 27 | 0.11 | 0.12 |
| 1* | 10.8 | 22.6 | 215 | 235 | 139 | 152 | 14.1 | 246 | 8.21 | - | 18.7 | - | 0.1 | 0.11 |
| 1.5* | 11 | 21.1 | 214 | - | 139 | - | 210 | - | 8.11 | - | 23.3 | - | 0.1 | - |
| 2* | 10.5 | 21.1 | 210 | - | 136 | - | 198 | - | 8.2 | - | 21.4 | - | 0.1 | - |
| 2.5* | 10 | 21.1 | 210 | 274 | 136 | 177 | 205 | 213.3 | 8.2 | - | 21.5 | - | 0.1 | 0.12 |
| 3* | 9.6 | 22.4 | 209 | 219 | 136 | 142 | 175 | - | 8.15 | - | 18.6 | - | 0.1 | 0.11 |
| 3.5* | 9.5 | - | 208 | - | 135 | - | 148 | - | 8.1 | - | 19.1 | - | 0.1 | - |
| 4* | 9.1 | 22 | 210 | - | 136 | - | 137 | - | 8.06 | - | 14.3 | - | 0.1 | - |
| 4.5* | 9.6 | 22.5 | 209 | - | 135 | - | 92.4 | - | 8.12 | - | 19.7 | - | 0.1 | - |
| 5* | 9.2 | 23 | 211 | - | 137 | - | 137 | - | 7.92 | - | 15.9 | - | 0.1 | - |
| 5.5* | 9.9 | 22.5 | 216 | - | 141 | - | 153 | - | 8.2 | - | 15.8 | - | 0.1 | - |
| 6* | 11.1 | 22.4 | 216 | - | 139 | - | 206 | - | 8.21 | - | 24 | - | 0.1 | - |
| 13.6* | 10.5 | 21.5 | - | - | 138 | - | 184 | - | 8.18 | - | 15.2 | - | 0.1 | - |
Примечание. ЗПВ – водоемы заплесковой зоны (глубина 0.05 м); Т – температура, ºС; EC – электропроводность, мкСм/см; TDS – общее количество растворенного вещества, мг/л; ORP – окислительно-восстановительный потенциал, минивольты, mV; pH – активный водородный показатель; Turb – мутность, ЕМФ; Sal – соленость, ppt; дата отбора: 1 – 31.05–01.06; 2 – 31.07–06.08; «-» – данные отсутствуют; «*» – физико-химические параметры приведены для поверхностного горизонта.
По степени минерализации воды оз. Арахлей относятся к группе пресных. Состав воды гидрокарбонатный магниево-кальциевый (Ивано-Арахлейские озера…, 2013). В период проводимых исследований общее количество растворенного вещества в озере не превышало 154 (май – июнь) и 177 (июль – август) мг/л, что соответствует ранее полученным данным 1993–2011 гг. (Ивано-Арахлейские озера…, 2013). В соответствии со значениями минерализации изменялись и значения электропроводности (214.09 ± 8.15 (весна) и 246.0 ± 24.04 (лето) мкСм/см). Значение рН воды в прибрежье варьировало от 7.92 до 8.21. Диапазон измеренных величин мутности весной составил от 14.3 до 29.3 ЕМФ.
Формирование термического режима оз. Арахлей в период открытой воды обусловлено метеорологическими условиями (Обязов, 2011). Прогрев вод в озере начинается в середине апреля подо льдом и заканчивается в приповерхностном слое воды (0–6 м) к середине июля – началу августа (Ивано-Арахлейские озера…, 2013). В период исследования средняя температура воды в прибрежье весной составляла 9.65 ± 2.24 ºС, летом – 22.19 ± 0.86 ºС. В центральной части водоема – 7.48 ± 0.44 ºС и 17.5 ± 4.89 ºС соответственно.
Необходимо также указать, что в прибрежной части озера на глубинах 2.5; 3.0; 3.5; 4.0; 4.5 м (в мае – июне) и на глубинах 3.5; 4.0; 6.0 м (в июле – августе) отмечены сообщества растительности, представленные роголистником и рдестами. Такое распределение сообществ гидрофитов соответствует поясам растительности оз. Арахлей (Базарова, 2003).
Методы
Ввиду достаточно высокой прозрачности воды (3.50 м (май – июнь) и 5.76 м (июль – август)) отбор проб в прибрежной части водоема проводился с поверхностного горизонта. Схема разреза и сетка станций отбора проб представлены на рис. 1б. В центральной части озера (геометрический центр водоема, глубина 13.6 м) сбор материала вели в приповерхностном слое, на глубинах, равных половине прозрачности, прозрачности, полторы прозрачности и у дна. Пробы, отобранные с каждого горизонта, сливали в чистую емкость, тщательно перемешивали и отбирали одну интегрированную пробу.
Методика отбора и обработки материала представлена в табл. 2.
Таблица 2. Методы, использованные при изучении фитопланктона оз. Арахлей
| Параметры | Методы/приборы | Литература | |
| Отбор проб | Батометр Паталаса, объем пробы 1 литр | Садчиков, 2003 | |
| Консервирование | 4 % формальдегид | ||
| Пробоподготовка | Осадочный метод концентрирования | ||
| Определение / оценка | таксономического состава | Микроскоп Nicon Eclipse E200-F (1000´) (Япония) | Монографии, сводки, определители, приведенные в работе Н. А. Ташлыковой (2009); уточнение синонимов – система сайта AlgaeBase (Guiry, Guiry, 2017) |
| численности | Метод Гензена | Садчиков, 2003 | |
| биомассы | Метод «истинного объема» | ||
| значение отдельных видов | По частоте встречаемости | Кожова, 1970 | |
| эколого-географической характеристики | Географическая приуроченность, местообитание, соленость среды, кислотность среды | Баринова и др., 2006 | |
| ценотической структуры альгоценозов | Индекс разнообразия Шеннона | Shanon, Weaver, 1963 | |
| выравненность видов в сообществе | Индекс Пиелу | Одум, 1975 | |
| степень флористического сходства | Коэффициент Серенсена | Sorensen, 1948 | |
| Статистическая обработка данных | Кластеризация, построение диаграммы размаха, корреляционный анализ с использованием коэффициента ранговой корреляции Спирмена (ρ) | XLSTAT (2017) | |
При кластеризации использовалось расстояние Пирсона с применением алгоритма одиночной связи, что позволило сравнить флористическую и ценотическую структуру сообществ разных участков озера, выраженную как число видов и численность определенных таксономических групп фитопланктона.
Результаты
За обследованный период в фитопланктоне обнаружено 97 видов, разновидностей и форм водорослей (Cyanobacteria – 18, Bacillariophyta – 30, Chrysophyta – 10, Charophyta – 5, Chlorophyta – 26, Dinophyta – 4, Euglenophyta – 4).
Наиболее богаты видами диатомовые (30.9 %), зеленые (26.8 %), цианобактерии (18.6 %) и золотистые водоросли (10.3 %), они составляли 86.6 % от общего таксономического разнообразия. Альгофлора прибрежья оз. Арахлей представлена богаче, чем пелагиали. Количество таксонов, встреченных в береговой зоне, составило 79, против 39 в центральной.
К наиболее часто встречаемым видам прибрежья и открытой части озера (интервал 50–100 %) относились: из диатомовых – Lindavia comta (Kützing) Nakov, Gullory, Julius, Theriot & Alverson, Fragilaria crotonensis Kitton, F. radians (Kützing) D. M. Williams & Round, Ulnaria ulna (Nitzsch) Compère in Jahn et al., Cocconeis placentula Ehrenberg, Nitzschia graciliformis Lange-Bertalot & Simonsen, из золотистых – Chrysococcus rufescens Klebs, Dinobryon cylindricum O. E. Imhof, D. divergens O. E. Imhof, D. sertularia O. E. Imhof, Kephyrion spirale (Lackey) Conrad, из зеленых – Oocystis marssonii Lemmermann. Из них шесть (L. comta, F. crotonensis, C. placentula, D. cylindricum, D. divergens, K. spirale) были постоянными для весеннего и летнего планктона 2017 г.
Анализ сходства альгофлоры продемонстрировал близость планктонных сообществ берегового и центрального участков. Коэффициент сходства по Серенсену в мае – июне составил 61 %, в июле – августе – 45 %. Различие в таксономическом составе фитопланктона между отдельными станциями прибрежной части оз. Арахлей было незначительным. Коэффициент видового сходства варьировал от 40 до 81 %. Дендрограмма, полученная в результате анализа, позволила выделить четыре кластера, объединяющих альгоценозы, сходные по биотопу (рис. 2).
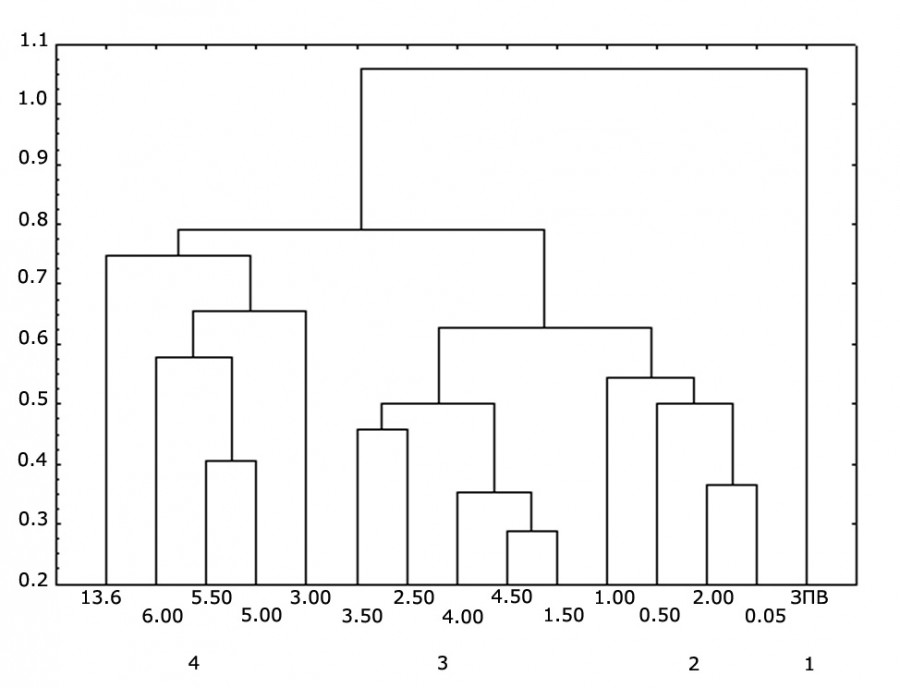
Рис. 2. Дендрограмма биоценотического сходства альгофлоры оз. Арахлей (весенний период) (расстояние Пирсона, одиночная связь): 1–4 – номера выделенных кластеров
Fig. 2. Dendrogram of the hierarchical cluster analysis of the algoflora of Lake Arakhley (spring) (Pearson distance, Single Linkage): 1–4 – numbers of selected clusters
Первый кластер позволил выявить специфичность состава водорослей станции «водоемы заплесковой зоны» (ЗПВ). Здесь обнаружено наибольшее количество представителей отдела Cyanobacteria, некоторые из них, например виды рода Oscillatoria Vaucher ex Gomont (O. geminata Schwabe ex Gomont, O. woronichinii Anissimova, O. tenuissima C. Agardh ex Forti), не были встречены на станциях других групп. Второй кластер объединил станции мелководной зоны прибрежного участка. Эта группа станций характеризовалась низким таксономическим разнообразием (19–23 вида (включая разновидности и формы)) и преобладанием бентосных форм диатомей (более 50 %). Коэффициент сходства водорослей в этом кластере изменялся от 35 до 78 %. В третий кластер вошли станции, характеризовавшиеся наличием высшей водной растительности. Они отличались от ассоциаций других станций высоким разнообразием золотистых водорослей (10 видов) и низким – хлорококковых водорослей (1–2 вида). Кроме того, присутствие растительности способствовало увеличению числа водорослей в группе за счет поступления в планктон факультативно планктонных видов. Коэффициент сходства для этой группы изменялся от 57 до 81 %. Четвертый кластер включил станции глубоководной зоны прибрежья и центральную станцию пелагиали. В этом объединении станций отмечено высокое разнообразие хлорококковых водорослей (10–13 видов).
Эколого-географический анализ показал, что в составе фитопланктона озера преобладали космополитные виды (89 %), преимущественно бентосно-планктонные и бентосные формы (63.6 %). При этом в прибрежье доля бентосных и планктонно-бентосных видов была в 2–2.5 раза выше, чем на центральной станции. По отношению к солености воды наиболее разнообразны индифференты, составляющие 86.8 % от общего числа видов. По отношению к кислотности среды преобладали алкалифилы (54.2 %) при значительной доле индифферентов (45.8 %).
Количественное развитие водорослей в мае – июне было невысоким и создавалось преимущественно мелкоразмерными золотистыми водорослями. В июле – августе их численность и биомасса возрастали на порядок, так как в толще воды отмечалось массовое развитие крупных форм диатомей (рис. 3, 4).
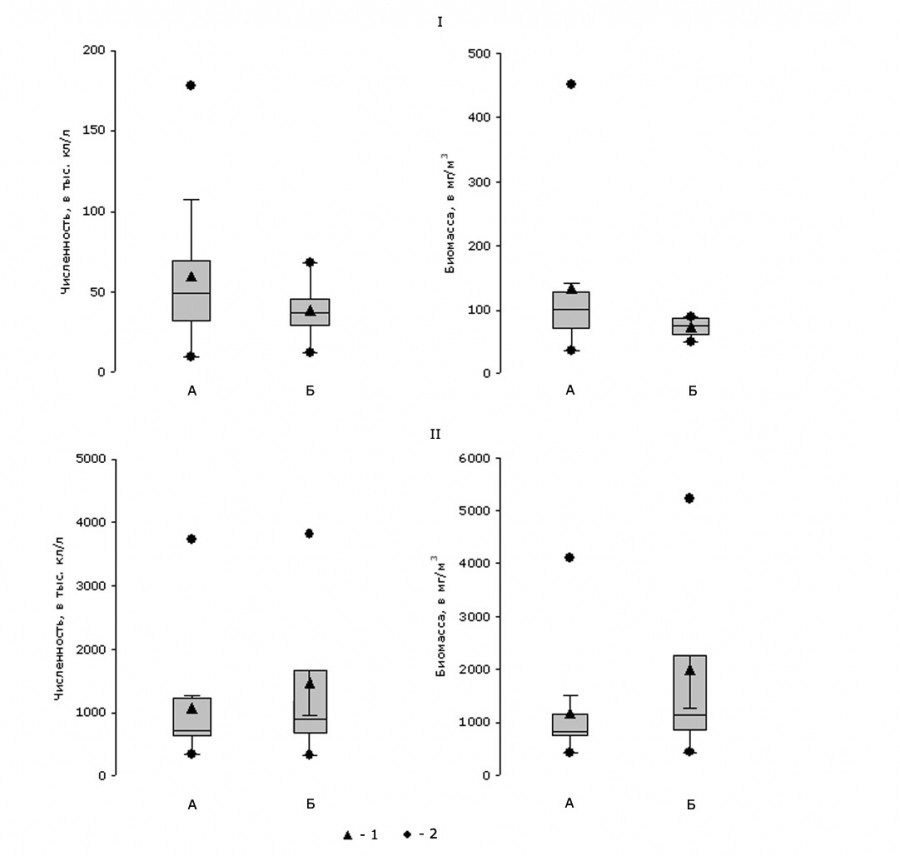
Рис. 3. Диаграмма размаха показателей количественного развития водорослей оз. Арахлей в исследуемые сезоны 2017 г.: I – весна, II – лето; А – прибрежье, Б – центр; 1 – среднее, 2 – минимум/максимум
Fig. 3. Diagram of the range of indicators of quantitative development of algae in Lake Arakhley in the studied seasons of 2017: I – spring, II – summer; A – coastal area, B – the center; 1 – mean, 2 – minimum/maximum
В оба сезона исследования наибольшие значения численности и биомассы фитопланктона выявлены на станциях прибрежья. Здесь отмечалась тенденция постепенного увеличения количественных показателей от станций мелководной зоны до станций, характеризовавшихся наличием высшей водной растительности (2.5–4.5 м). В мае – июне максимальные значения характеристик количественного развития водорослей (461.52 тыс. кл/л; 453.37 мг/м3) отмечались на станции с глубиной 3.0 м, в июле – августе (3733.4 тыс. кл/л, 4105.01 мг/м3) – на станции с глубиной 4.0 м. На следующей станции наблюдалось резкое снижение численности и биомассы. Так, в мае – июне на 3.5 м численность водорослей определялась в 37.51 тыс. кл/л, при биомассе 36.9 мг/м3, в июле – августе значения численности и биомассы на глубине 4.5 м соответствовали 341.3 тыс. кл/л и 433.52 мг/м3. Затем с увеличением глубины количественные показатели вновь возрастали. Особенно отчетливо это прослеживалось в июле – августе: 5.0 м (604.8 тыс. кл/л, 751.01 мг/м3) → 5.5 м (688 тыс. кл/л, 844 мг/м3) → 6.0 м (1184 тыс. кл/л, 1099.22 мг/м3). Фитопланктону мелких водоемов заплесковой зоны было свойственно обильное развитие водорослей (188.48 тыс. кл/л, 147.36 мг/м3) в весенний период, когда значения температуры доходили до 25.8 ºС. Летом численные показатели водорослей соответствовали средним значениям станций прибрежья.
Структуру альгоценозов, как в прибрежной, так и в пелагической части водоема, в весенний период исследований (31.05–01.06) определяли золотистые (10–75 % от общей численности и 30–92 % от общей биомассы) и диатомовые (20–85 % от общей численности и 10–60 % от общей биомассы) водоросли (рис. 4). Среди золотистых доминировали D. cylindricum, D. divergens, C. rufescens, среди диатомей – L. comta, F. crotonensis. Летний период (31.07–06.08) характеризовался преобладанием в фитопланктоне диатомово-золотистого комплекса. Представители Bacillariophyta создавали от 60 до 85 % от общей численности и от 50 до 95 % от общей биомассы. Основу их количественных характеристик определяла F. crotonensis, составляя 50–99 % от общей численности и 70–98 % от общей биомассы водорослей планктона. Вегетация золотистых водорослей значительно снизилась. Они составляли не более 25–30 % от общих количественных показателей.
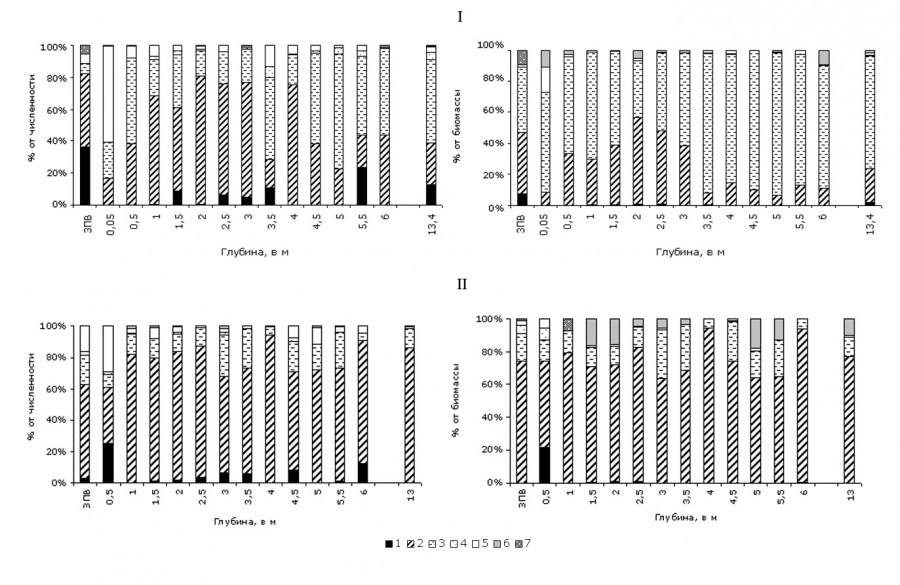
Рис. 4. Доля основных групп водорослей в фитопланктоне оз. Арахлей в 2017 г.: I – весна, II – лето; 1 – Cyanobacteria, 2 – Bacillariophyta, 3 – Chrysophyta, 4 – Charophyta, 5 – Chlorophyta, 6 – Dinophyta, 7 – Euglenophyta
Fig. 4. The share of the main algae groups in the phytoplankton of Lake Arakhley in 2017: I – spring, II – summer; 1 – Cyanobacteria, 2 – Bacillariophyta, 3 – Chrysophyta, 4 – Charophyta, 5 – Chlorophyta, 6 – Dinophyta, 7 – Euglenophyta
Оценка биоразнообразия в фитопланктоноценозах прибрежной и центральной частей оз. Арахлей проводилась с помощью индексов Шеннона и Пиелу. Фитопланктон озера характеризовался высокими значениями данных индексов, что указывает на сложность и высокое разнообразие сообщества. В весенний сезон в прибрежной части индекс Шеннона изменялся от 1.91 до 3.69, индекс Пиелу – в пределах 0.42–0.82; в центральной – 3.11 и 0.76 соответственно. Летом значение индекса Шеннона было несколько ниже и составляло в литорали 0.43–1.97; в пелагиали – 1.38. Индекс Пиелу в прибрежье варьировал от 0.13–0.84, на центральной станции определялся в 0.28. Такие значение показателей соответствуют сезонному распределению водорослей, так как известно, что максимальные значения индекса Шеннона фиксируются весной и осенью, минимальные – зимой и летом.
Проведенный анализ корреляционных связей количественных показателей фитопланктона (численность и биомасса фитопланктона, основных отделов, доминирующих и массовых видов, число таксонов) с некоторыми абиотическими характеристиками среды (глубина, температура, прозрачность, активный водородный показатель, мутность, общее количество растворенного вещества, электропроводность, окислительно-восстановительный потенциал, соленость) показал, что основными факторами, влияющими на развитие фитопланктона в пелагиали являются глубина, температура, прозрачность, активный водородный показатель, окислительно-восстановительный потенциал и электропроводность (табл. 3).
Таблица 3. Корреляционные связи количественных показателей фитопланктона с некоторыми абиотическими характеристиками среды в обследованной пелагической зоне оз. Арахлей (p < 0.05).
| Показатель | Значение | Показатель | Значение | ||
| Глубина | численность Chrysophyta | 0.706 | Температура | численность Chrysophyta | 0.721 |
| биомасса Bacillariophyta | 0.818 | численность Bacillariophyta | -0.756 | ||
| биомасса Chrysophyta | -0.870 | биомасса Chrysophyta | 0.710 | ||
| численность D. cylindricum | -0.854 | число таксонов | 0.751 | ||
| численность C. rufescens | -0.798 | численность K. spirale | 0.851 | ||
| биомасса L. comta | 0.952 | биомасса L. comta | -0.833 | ||
| биомасса D. cylindricum | -0.849 | Мутность | численность Euglenophyta | -0.819 | |
| биомасса C. rufescens | -0.799 | биомасса Euglenophyta | -0.787 | ||
| Прозрачность | численность Dinophyta | -0.873 | Активный водородный показатель | численность Dinophyta | -0.865 |
| численность Euglenophyta | -0.845 | численность Euglenophyta | -0.807 | ||
| биомасса Chlorophyta | -0.914 | биомасса Chlorophyta | -0.889 | ||
| биомасса Dinophyta | -0.928 | биомасса Dinophyta | -0.918 | ||
| биомасса Euglenophyta | -0.904 | биомасса Euglenophyta | -0.871 | ||
| биомасса C. rufescens | -0.711 | численность C. rufescens | -0.794 | ||
| численность C. rufescens | -0.795 | биомасса C. rufescens | -0.795 | ||
| биомасса K. spirale | -0.876 | биомасса K. spirale | -0.877 | ||
| Окислительно-восстановительный потенциал | численность Dinophyta | 0.894 | Электропроводность | численность Dinophyta | -0.885 |
| численность Euglenophyta | 0.748 | численность Euglenophyta | -0.812 | ||
| биомасса Bacillariophyta | -0.735 | биомасса Chlorophyta | -0.840 | ||
| биомасса Chlorophyta | 0.884 | биомасса Dinophyta | -0.825 | ||
| биомасса Dinophyta | 0.925 | биомасса Euglenophyta | -0.789 | ||
| биомасса Euglenophyta | 0.814 | численность C. rufescens | -0.822 | ||
| биомасса C. rufescens | 0.833 | биомасса C. rufescens | -0.822 | ||
| биомасса K. spirale | 0.904 | биомасса K. spirale | -0.893 | ||
| биомасса D. cylindricum | 0.763 | ||||
Для литорали оз. Арахлей выявлено низкое число высоких достоверных корреляционных связей биотических параметров фитопланктона с перечисленными выше морфометрическими, гидрофизическими и гидрохимическими характеристиками (табл. 4). Преимущественно корреляционные зависимости отмечены для численных характеристик второстепенных групп и видов фитопланктона, обуславливающих таксономическое разнообразие водорослей.
Таблица 4. Корреляционные связи количественных показателей фитопланктона с некоторыми абиотическими характеристиками среды в обследованной литоральной зоне оз. Арахлей (p < 0.05)
| Показатель | Значение | Показатель | Значение | ||
| Электропроводность | численность Chlorophyta | -0.8850 | Окислительно-восстановительный потенциал | биомасса Dinophyta | 0.9247 |
| биомасса K. spirale | -0.8927 | численность C. rufescens | 0.8827 | ||
| Прозрачность | биомасса Chlorophyta | -0.914 | численность Dinophyta | 0.8945 | |
| биомасса Dinophyta | -0.9281 | биомасса Chlorophyta | 0.8837 | ||
| биомасса Euglenophyta | -0.9037 | Общее количество растворенного вещества | численность Chlorophyta | 0.9938 | |
| pH | биомасса Chlorophyta | -0.8889 | Глубина | биомасса L. comta | 0.9524 |
| биомасса Dinophyta | -0.9180 | ||||
Обсуждение
Сравнение полученных результатов с материалами ранее проведенных исследований (1966–1976, 1975–1985, 2008–2009 гг.) показало, что количество обнаруженных таксонов несколько ниже, чем ранее. Так, в 2017 г. в фитопланктоне отмечено 97 видов, разновидностей и форм водорослей, в 2008–2009 гг. – 110, в 1966–1976 гг. – 144 (Морозова, 1975, 1981; Оглы, 1993, 1995; Ивано-Арахлейские озера…, 2013). Таксономическое разнообразие, как и при исследованиях 1960–1980-х (Морозова, 1975, 1981; Оглы, 1993, 1995) и 2000-х гг. (Ивано-Арахлейские озера…, 2013), определяли диатомовые, зеленые, цианобактерии, золотистые, харовые, динофитовые и эвгленовые водоросли. Аналогичное распределение водорослей по основным группам наблюдалось при изучении фитопланктона литоральной зоны водоемов Сибири и некоторых озер Европейской части России (Вислянская, 1982; Касперовичене, Каросене, 2005; Зарубина и др., 2005; Помазкина и др., 2010; Чекрыжева, Калинкина, 2016; Русановская и др., 2017 и др.).
Состав доминирующего комплекса в оз. Арахлей на протяжении более чем 50 лет стабилен (Морозова, 1975, 1981; Оглы, 1993, 1995; Ивано-Арахлейские озера…, 2013). Годовые изменения состава обусловлены колебаниями уровневого режима водоема. Для маловодных годов характерно доминирование Cyclotella comta Kützing (в настоящее время L. comta (Guiry, Guiry, 2017)), для многоводных – Asterionella formosa Hassall (Морозова, Шишкин, 1973). Так, в 1966–1969 гг. в альгоценозе озера преобладала C. comta (Морозова, 1975; Морозова, Шишкин, 1973), в 1970–1971 гг. – A. formosa (Морозова, Шишкин, 1973; Морозова, 1981), в 1990–1995 гг. – A. formosa (Оглы, 1993, 1995), в 2008–2009 гг. – Puncticulata radiosa (Kützing) H. Hakansson (в настоящее время L. comta (Guiry, Guiry, 2017)) (Ивано-Арахлейские озера…, 2013), в 2017 г. – L. comta. Такое распределение доминирующих видов соответствует чередующимся фазам гидрологического цикла в озере: в 1962 г. уровень достиг максимума и со следующего года начал снижаться; в 1967–1968 гг. падение уровня сменилось его подъемом, продолжавшимся до 1972 г. Следующая маловодная фаза продолжалась до 1980 г. Последующий рост уровня длился 4–6 лет. До 1990–1991 гг. его колебания происходили на достаточно высоких отметках. В начале 1990-х гг. уровень начал понижаться, достигнув к 2008 г. близких к абсолютному минимуму отметок (Обязов, 2011).
Несмотря на то, что изученные зоны озера являются биотопами, которые отличаются рядом абиотических и биотических факторов, состав фитопланктона был сходен, что подтверждается значениями коэффициента сходства (45–61 %). Как отмечают Зарубина и др. (2005), постоянные процесы водообмена между литоралью и пелагиалью способствуют обмену видами между открытыми участками литорали, зарослями и собственно пелагиалью.
Количественное развитие водорослей в центральной части оз. Арахлей весной и летом 2017 г. соответствовало данным, полученным в 1967–1971 (Морозова, Шишкин, 1973) и 2008–2009 гг. (Ивано-Арахлейские озера…, 2013). Однако соотношение основных групп фитопланктона было несколько иным. Так, в 1966–1971 гг. фитопланктон носил преимущественно диатомовый характер (до 80 % среднегодовой биомассы). В его составе эпизодически усиливали свою роль синезеленые водоросли Microcystis pulverea (H. C. Wood) Forti и Holopedia geminata Lagerheim. К примеру, в июле 1966 г. валовая продукция фитопланктона в момент обогащения H. geminata составляла 680 г/м2 С против 413 г/м2 С в июне того же года (Морозова, Шишкин, 1973). В 2008 г. лидирующее положение в весеннем фитопланктоне принадлежало золотистым и зеленым водорослям (50–95 % от общего количества), в 2009 г. – только золотистым (80 % от общей численности и биомассы). В летний период 2008–2009 гг. численность водорослей создавалась преимущественно диатомеями (75 %) (Ивано-Арахлейские озера…, 2013).
Анализ корреляционных связей показал, что в пелагиали основными для развития доминирующих групп и видов водорослей планктона из гидрофизических и гидрологических факторов являются глубина, температура и прозрачность. В прибрежной части водоема прозрачность воды, активный водородный показатель и окислительно-восстановительный потенциал обуславливали вегетацию преимущественно второстепенных групп водорослей с низкими количественными показателями, создающими таксономическое разнообразие фитопланктона озера. На определяющее действие освещенности, зависящее от изменения уровня воды и динамики водных масс, для фитопланктона оз. Арахлей указывалось в работе Б. А. Шишкина (1972). При исследованиях 1966–1971 и 2008–2009 гг. (Морозова, 1981; Ивано-Арахлейские озера…, 2013 ) отмечено существенное влияние температурного фактора, обуславливающего вертикальное распределение максимальных значений численности и биомассы фитопланктона в пелагиали озера. Подобные результаты, касающиеся значимости приведенных выше абиотических параметров, получены при проведении лабораторных и стационарных исследований для других водоемов (Трифонова, 1990; Абакумов, Израильский, 2012; Девяткин и др., 2012; Дудакова и др., 2015; Даценко и др., 2017; Smith, 1986; Zhu et al., 2018; Kremer et al., 2017).
Заключение
Проведенные исследования фитопланктона озера Арахлей показали, что таксономический состав водорослей в прибрежной части озера сравнительно богаче, чем в центральной. Количество таксонов фитопланктона, обнаруженных в прибрежной части оз. Арахлей, составило 79, в центральной – 39. Основу таксономического разнообразия прибрежного и пелагического фитопланктона составляли диатомовые, зеленые, золотистые водоросли и цианобактерии. К наиболее часто встречаемым видам прибрежья и открытой части озера отнесены 12 видов (L. comta, F. crotonensis, F. radians, U. ulna, C. placentula, N. graciliformis, C. rufescens, D. cylindricum, D. divergens, D. sertularia, K. spirale, O. marssonii), из которых 6 были постоянны для весеннего и летнего планктона 2017 г. Анализ сходства альгофлоры продемонстрировал близость планктонных сообществ берегового и центрального участков (коэффициент сходства по Серенсену изменялся от 45 до 61 %). Проведенная кластеризация позволила выявить четыре кластера, объединяющих альгоценозы, сходные по биотопу: водоемы заплесковой зоны, станции мелководной зоны прибрежья, станции с наличием высшей водной растительности и станции глубоководной зоны. Количественное развитие водорослей в весенний период было невысоким и создавалось преимущественно мелкоразмерными золотистыми водорослями. Летом их численность и биомасса возрастали на порядок, так как в толще воды отмечалось массовое развитие крупных форм диатомей. Проведенный корреляционный анализ показал, что в исследуемые периоды основным из гидрофизических и гидрологических факторов, определяющим развитие фитопланктона в прибрежной и центральной зонах озера, является освещенность воды. Высокие значения индексов Шеннона и Пиелу указывают на сложность структуры и высокое биоразнообразие фитопланктонного сообщества, а их сезонное распределение демонстрирует отклик фитопланктоценоза на природно-климатические изменения условий среды.
Библиография
Абакумов А. И., Израильский Ю. Г. Влияние условий среды на распределение фитопланктона в водоеме // Матем. биол. и биоинформ. 2012. Т. 7. № 1. С. 274–283.
Базарова Б. Б. Структура и продуктивность растительности водных экосистем Восточного Забайкалья (на примере озер бассейна реки Хилок): Автореф. дис. ... канд. биол. наук . Улан-Удэ, 2003. 18 с.
Баринова С. С., Медведева Л. А., Анисимова О. В. Биоразнообразие водорослей-индикаторов окружающей среды . Тель-Авив: Pilies Studio, 2006. 498 с.
Вислянская И. Г. Фитопланктон // Лимнологические исследования на заливе Онежского озера Большое Онего . Л.: Зоол. ин-т АН СССР, 1982. С. 70–81.
Даценко Ю. С., Пуклаков В. В., Эдельштейн К. К. Анализ влияния абиотических факторов на развитие фитопланктона в малопроточном стратифицированном водохранилище // Труды КарНЦ РАН. 2017. № 10. С. 73–85.
Девяткин В. Г., Метелева Н. Ю., Митропольская И. В. Адаптация фитопланктона к гидрофизическим факторам // Озерные экосистемы: биологические процессы, антропогенная трансформация, качество воды: Материалы IV междунар. науч. конф. Минск, 2011. С. 58.
Девяткин В. Г., Метелева Н. Ю., Вайновский П. А. О роли фитопланктона в формировании кислородного режима водоема в связи с климатическими вариациями // Вода: химия и экология. 2012. № 12 (54). С. 68–74.
Дудакова Д. С., Родионова Н. В., Протопопова Е. В., Капустина Л. Л., Бардинский Д. С. Состав и динамика гидробионтов литорали бухты Петрокрепость Ладожского озера // Труды КарНЦ РАН. 2015. № 2. С. 53–70.
Зарубина Е. Ю., Яныгина Л. В., Бурмистрова Щ. С., Митрофанова Е. Ю., Ким Г. В., Котовщиков А. В., Крылова Е. Н., Ковешников М. И. Литоральные биоценозы как один из факторов устойчивости экосистемы Телецкого озера // Ползуновский вестник. 2005. № 4. С. 201–207.
Ивано-Арахлейские озера на рубеже веков (состояние и динамика) / Под ред. Н. М. Пронина. Новосибирск: Изд-во СО РАН, 2013. 337 с.
Касперовичене Ю., Каросене Ю. Структурно-функциональная характеристика эпифитона и фитопланктона литоральной зоны оз. Сперва (Литва) // Вестник ТюмГУ. 2005. № 5. С. 70–77.
Кожова О. М. Формирование фитопланктона // Формирование природных условий и жизни Братского водохранилища. М.: Наука, 1970. С. 27–160.
Морозова Т. Н. Сезонные и годовые изменения фитопланктона Ивано-Арахлейских озер: Автореф. дис. ... канд. биол. наук . Иркутск, 1975. 29 с.
Морозова Т. Н. Фитопланктон озера Арахлей // Биологическая продуктивность. Новосибирск: Наука, 1981. С. 19–30.
Морозова Т. Н., Шишкин Б. А. Годовые изменения фитопланктона некоторых Ивано-Арахлейских озер // Лимнологические исследования в Забайкалье. Чита, 1973. С. 57–70.
Обязов В. А. Гидрологический режим озер Забайкалья в условиях меняющегося климата (на примере Ивано-Арахлейских озер) // Водное хозяйство России. 2011. № 3. С. 4–14.
Оглы З. П. Фитопланктон разнотипных озер Забайкалья: Автореф. дис. ... канд. биол. наук . СПб., 1993. 24 с.
Оглы З. П. Многолетние наблюдения за фитопланктоном озер Центрального Забайкалья // Проблемы экологии Прибайкалья: Материалы Междунар. конф. Новосибирск: Наука, 1995. С. 177–186.
Одум Ю. Основы экологии . М: Мир, 1975. 740 с.
Петрова Н. А. Фитопланктон литоральной зоны Онежского озера // Литоральная зона Онежского озера. Л.: Наука, 1975. С. 138–144.
Помазкина Г. В., Белых О. И., Домышева В. М., Сакирко М. В., Гнатовский Р. Ю. Структура и динамика фитопланктона в южном Байкале (Россия) // Альгология. 2010. Т. 20. № 1. С. 56–72.
Протопопова Е. В. Фитопланктон литоральной зоны Ладожского озера // Литоральная зона Ладожского озера. СПб.: Нестор-История, 2011. С. 102–128.
Русановская О. О., Кобанова Г. И., Шимараева С. В., Пислегина Е. В., Кращук Л. С., Зилов Е. А. Оценка качества вод Среднего и Северного Байкала по состоянию прибрежного фитопланктона // ХХI век. Техносферная безопасность. 2017. Т. 2. № 2. С. 30–37.
Садчиков А. П. Методы изучения пресноводного фитопланктона . М.: Университет и школа, 2003. 157 с.
Соловьева В. В., Корнева Л. Г. Характеристика сапробности мелководий и пелагиали Волжского плеса Рыбинского водохранилища по фитопланктону // Научные ведомости Белгородского государственного университета. Серия: Естественные науки. 2012. Т. 21. № 21 (140). С. 34–39.
Ташлыкова Н. А. Экологические особенности развития фитопланктона дельтовых проток реки Селенги и сора Черкалово (оз. Байкал): Дис. ... канд. биол. наук . Улан-Удэ, 2009. 207 с.
Трифонова И. С. Экология и сукцессия озерного фитопланктона . Л.: Наука. Ленингр. отд-ние, 1990. 182 с.
Чекрыжева Т. А., Калинкина Н. М. Структура и сезонная динамика фитопланктонных сообществ в открытой и закрытой литорали Онежского озера (Пиньгуба, Пухтинская бухта) // Труды КарНЦ РАН. 2016. № 12. С. 86–95.
Шишкин Б. А. Об основных закономерностях межгодовых изменений режима Ивано-Арахлейских озер // Зап. Заб. фил. геогр. об-ва СССР. Вып. 80. Чита, 1972. С. 151–162.
Asma’ Jamal, Fatimah Md. Yusoff, Sanjoy Banerjee, Shariff M. Littoral and limnetic phytoplankton distribution and biodiversity in a tropical Man-Made Lake, Malaysia // Advanced Studies in Biology. 2014. Vol. 6. No 4. P. 149–168. URL: http://ddx.doi.org/10.12988/asb.2014.4631.
Decamps H., Pinay G., Naiman R. J., Petts G. E., McClain M. E., Ilkowska H., Hanley T. A., Holmes R. M., Quinn J., Gibert J., Planty Tabacchi A.-M., Schiemer F., Tabacchi E., Zalewski M. Riparian zones: where biogeochemistry meets biodiversity in management practice // Polish Journal of Ecology. 2004. Vol. 52. No 1. P. 3–18.
Guiry M. D., Guiry G. M. AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. URL: http: //www.algaebase.org (дата обращения: 10.06.2017–10.09.2017).
Kremer C. T., Thomas M. K., Litchman E. Temperature and size scaling of phytoplankton population growth rates: Reconciling the Eppley curve and the metabolic theory of ecology // Limnology and oceanography. 2017. Vol. 62. No 4. P. 1658–1670.
Nozaki K. Abrupt change in primary productivity in a littoral zone of Lake Biwa with the development of a filamentous green‐algal community // Freshwater Biology. 2001. No 46 (5). P. 587–602. URL: http://ddx.doi.org/10.1046/j.1365-2427.2001.00696.x.
Nozaki K., Morino H., Munehara H., Sideleva V. G., Nakai K., Yamauchi M., Kozhova O. M., Nakanishi M. Composition, biomass, and photosynthetic activity of the benthic algal communities in a littoral zone of Lake Baikal in summer // Limnology. 2002. No 3. P. 175–180.
Schlosser I. J., Karr I. R. Water quality in agricultural watersheds: impact of riparian vegetation during baseflow // Wat. Res. Bull. 1981. No 17. P. 233–240.
Shanon C. E., Weaver W. The mathematical theory of communication. Urbana, 1963. 117 p.
Smith V. H. Light and nutrient effects on the relative biomass of blue-green algae in lake phytoplankton // Canadian journal of fisheries and aquatic sciences. 1986. No 43 (1). P. 148–153. URL: http://doi.org/10.1139/f86-016.
Sorensen T. A method of establishing groups of equal amplitude in plant ecology // Biol. Sci. 1948. Vol. 5. P. 25–39.
Taniguchi G. M., Bicudo D. C., Senna P. A. C. Gradiente litorâneo-limnético do fitoplâncton e ficoperifíton em uma lagoa da planície de inundação do Rio Mogi-Guaçu // Brazilian Journal of Botany. 2005. Vol. 28. No 1. P. 137–147. URL: http://dx.doi.org/10.1590/S0100-84042005000100011.
Turner M. A., Robinson G. O. C., Townsend B. E., Hann B. J., Amaral J. A. Ecological effects blooms filamentous green algae in the littoral zone of an acid lake // Canadian Journal of Fisheries and Aquatic Sciences. 2011. No 52 (10). P. 2264–2275. URL: http://ddx.doi.org/10.1139/f95-817.
Zhu R., Wang H., Chen J., Shen H., Deng X. Use the predictive models to explore the key factors affecting phytoplankton succession in Lake Erhai, China // Environmental Science and Pollution Research. 2018. Vol. 25. No 2. P. 1283–1293. URL: https://doi.org/10.1007/s11356-017-0512-2.
Zębek E. Qualitative and quantitative changes of diatoms with relation to physiochemical water parameters in the littoral zone of the urban Lake Jeziorak Mały // Oceanological and Hydrobiological Studies. 2007. Vol. XXXVI. No 4. P. 3–22.
Благодарности
Работа выполнена в рамках темы «Биоразнообразие природных и природно-техногенных экосистем Забайкалья (Центральной Азии) как индикатор динамики региональных изменений климата» подтемы «Выявление структурно-функциональной организации литоральных сообществ в условиях изменяющегося климата и антропогенных воздействий с использованием экспедиционных и стационарных исследований» проекта ФНИ № IX.137.1.1.
Выражаю свою искреннюю признательность и благодарность моим коллегам – сотрудникам лаборатории водных экосистем ИПРЭК СО РАН к. б. н. П. В. Матафонову и к. б. н. Г. Ц. Цыбекмитовой за помощь в отборе проб.

 © 2011 - 2026
© 2011 - 2026