



Введение
Сравнение однотипных природных сообществ различных регионов представляет значительный научный интерес, поскольку позволяет осмыслить зависимость структуры комплексов от факторов среды и исторических аспектов их формирования, но, к сожалению, работ такого плана относительно не много (Преображенская, 1982). Кроме того, такое сопоставление наглядно проявляет особенности функционирования экосистем в свете широтной, меридиональной и региональной зональности распределения организмов. За более чем 30 лет работы в заповедниках Урала и Приамурья был собран достаточно большой материал по населению птиц приречных участков пойм во время обследования рек с лодок. Представляется интересным сравнение имеющихся данных по столь удаленным регионам. Расстояние между Комсомольским и Норским заповедниками составляет около 500 км, между Вишерским и Норским – 4200 км, между Вишерским и Комсомольским – 4700 км. Комсомольский и Норский заповедники находятся на восточной и западной границах распространения представителей маньчжурской (амурской) фауны (Воробьев, 1954; Куренцов, 1965; Колбин, 2008). В Вишерском заповеднике, расположенном на Северном Урале, наблюдается преобладание представителей сибирской фауны (Колбин, 2011, 2016).
Материалы
Данные, послужившие основой для работы, собраны во время сплавов на лодках по рекам Комсомольского, Норского и Вишерского заповедников и сопредельных с ними территорий. В трех пойменных комплексах в общей сложности выявлено 195 видов птиц: из них на р. Горин – 106; на реках Норского заповедника – 132; в бассейне р. Вишеры – 112. Сравнительно низкое видовое разнообразие на р. Горин – в самом южном регионе, очевидно, соответствует наименьшей протяженности лодочных маршрутов.
Комсомольский заповедник расположен близ устья левого притока р. Амур – р. Горин (40 км ниже г. Комсомольск-на-Амуре Хабаровского края). Долина реки Горин (390 км) в пределах территории заповедника имеет юго-восточное направление. Ширина ее в северной части заповедника около 2 км, на юге, в устье реки долина расширяется до 4–5 км. В Комсомольском заповеднике сплавы по р. Горин в гнездовое время проводили в период с 1984 по 1991 г. Протяженность маршрутов в данном регионе составила 430 км.
Норский заповедник расположен в междуречье рек Селемджа (647 км) и Нора (305 км), занимает южные отроги хр. Джагды (бассейн р. Бурунда, 187 км) и прилегающую часть Амуро-Зейской равнины в Селемджинском районе Амурской области. Пойменные леса более чем на 90 % пройдены пожарами. В районе слияния рек Нора, Червинка и Селемджа образовалась своеобразная дельтоподобная система с сотнями проток и островов, меняющих свое расположение после крупных наводнений. Для рек характерен быстрый подъем и спад уровня воды в зависимости от дождей. В Норском заповеднике сплавы по рекам Нора, Бурунда и Селемджа проводили с 2000 по 2014 г. Протяженность маршрутов составила 920 км.
Вишерский заповедник расположен на северо-востоке Пермского края в верховьях р. Вишера (415 км). Для Вишеры, так же как и для рек Приамурья, характерны высокое половодье, дождевые паводки и низкая летняя межень. В Вишерском заповеднике и в Красновишерском районе сплавы по рекам Вишера, Велс и Язьва проходили с 2002 по 2016 г. Протяженность маршрутов составила 1220 км.
Методы
Сплавы проводили на весельных лодках в гнездовое время – с середины мая до начала июля. Во время таких маршрутов учитывали всех встреченных птиц, отмеченных визуально и по голосам.
Встречаемость птиц по результатам учетов с лодки рассчитывали в парах на 10 км реки. При этом нетерриториальные виды, которые не охраняют свою территорию, объединяли в пары условно. Для тех видов, которые многократно учитывались в разные годы, рассчитывали среднюю встречаемость, для оценки варьирования данных вычисляли стандартное отклонение.
Учеты птиц с лодки – удобный способ изучения фауны из-за легкости перемещения и возможности обследования значительных территорий, поэтому неудивительно, что этот подход применяли многие исследователи (Кистяковский, Смогоржевский, 1964, 1973; Пронкевич, 2013).
Вода затрудняет определение расстояния до слышимой птицы, поэтому целесообразно вычисление встречаемости, а не плотности населения. Объединение в одной выборке обитателей водной и наземной среды также вносит дополнительные сложности при интерпретации полученных данных. Обычно сплавы занимают продолжительное время и охватывают время как с высокой активностью птиц, так и с низкой, поэтому существенная часть малопоющих лесных птиц не учитывается, хотя околоводные виды выявляются полнее. Вследствие этого очевидно, что полученная информация не подходит для достоверной характеристики численности птиц пойменных лесов конкретного региона, но в качестве относительных показателей для сравнительного анализа она может использоваться. С другой стороны, большая протяженность водных маршрутов позволяет выявлять значительное количество видов за сравнительно короткое время.
Для характеристики сообществ рассчитывали индексы полидоминантности на основе индекса Симпсона (Песенко, 1982; Бигон и др., 1989):
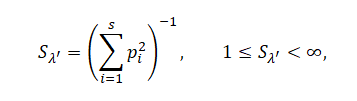
где pi – доля i-го вида в генеральной совокупности, включающей S видов, или вероятность его встречи в соответствующем населении. Согласно Ю. А. Песенко (1982), данные индексы наиболее пригодны для исследования биоразнообразия многовидовых сообществ.
Степень сходства различных сообществ оценивалась по индексу Съеренсена для качественных данных:
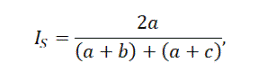
где a – число видов, входящих в оба списка, b и c – число видов, входящих только в 1-й или только во 2-й список.
Для количественных данных использовался индекс Съеренсена – Чекановского:
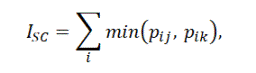
где pij – доля i-го вида в выборке J, pik – доля i-го вида в выборке K (Czekanowski, 1909; Sørensen, 1948; по Песенко, 1982). Данная формула рекомендована Ю. А. Песенко для сопоставления различных экологических выборок, поскольку напрямую измеряет мощность пересечения сравниваемых множеств.
Для анализа размещения гнезд использовались собственные наблюдения и литературные данные (Рябицев, 2014).
Достоверность различий между выборками оценивалась с помощью критерия Хи-квадрат. Анализ вклада разных систематических групп в наблюдаемые различия проводили методом главных компонент. Все расчеты выполняли средствами пакетов Microsoft Office Excel 2003–2007 и Statistica 6.
Результаты
На р. Горин и на реках Норского заповедника у 28 видов птиц доля в пойменном сообществе превысила 1 % (рис. 1, 2), на реках Вишера и Язьва такое превышение отмечено только у 20 видов (рис. 3). Индексы полидоминантности оказались в бассейне р. Вишеры также самыми низкими (табл. 1), что связано с уменьшением разнообразия таежной растительности по сравнению с широколиственными лесами, это подтверждают максимальные их значения на р. Горин.
Перекрывание рассматриваемых пойменных комплексов по индексам Съеренсена – Чекановского для количественных и качественных данных соответствует ожиданиям: сходство между реками Приамурья оказалось высоким, а между реками Урала и Приамурья низким. Причем перекрывание по видовому составу между тремя территориями имело более высокие значения, что определяется меньшей изменчивостью видовых списков без учета обилия.
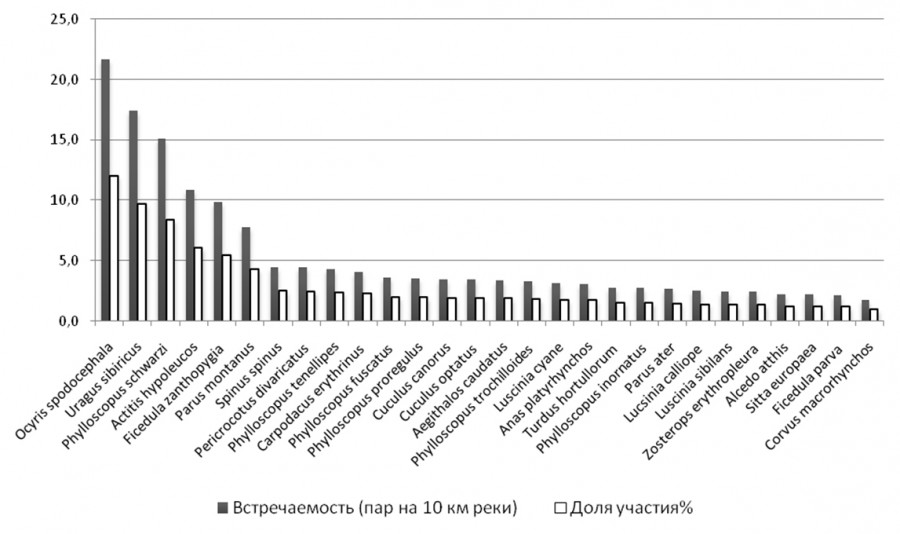
Рис. 1. Встречаемость птиц в пойменных сообществах рек Норского заповедника
Fig. 1. The occurence of birds in the floodplane communities in the rivers of Norsky nature reserve
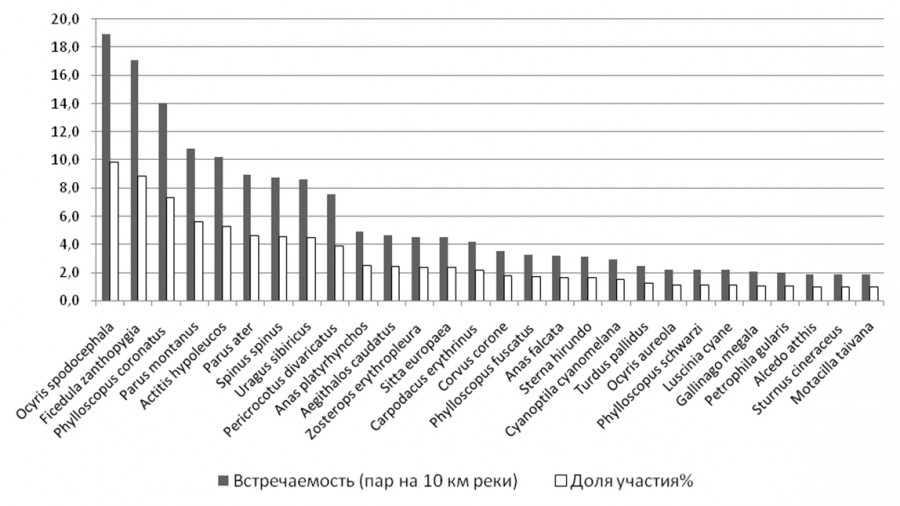
Рис. 2. Встречаемость птиц в пойменных сообществах р. Горин
Fig. 2. The occurence of birds in abundance in floodplane communities of the river Gorin
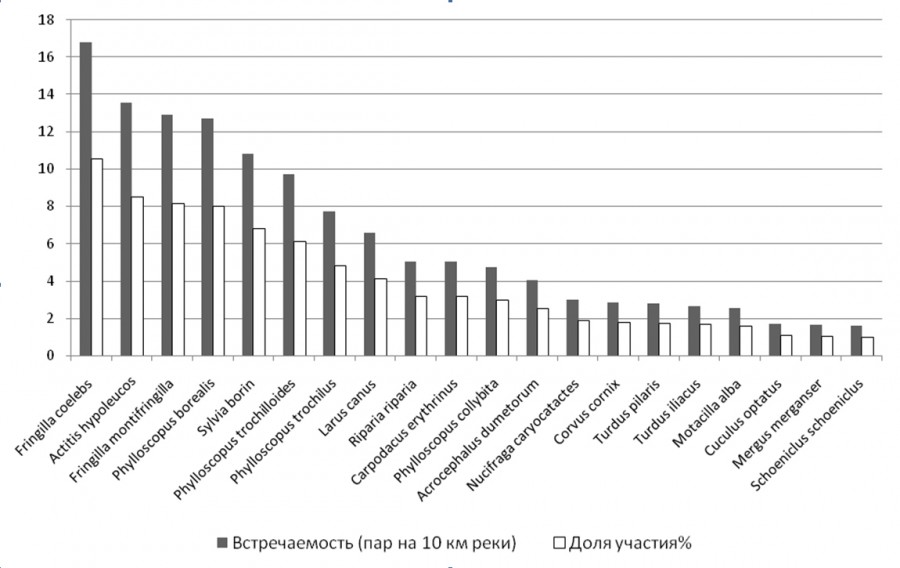
Рис. 3. Встречаемость птиц в пойменных сообществах р. Вишеры
Fig. 3. The occurence of birds in the floodplane communities of the river Vishera
Таблица 1. Показатели разнообразия и сходства пойменных орнитокомплексов
| Сравниваемые реки | Индекс полидоминантности | Показатели сходства | ||
| Вишера | Нора | Горин | ||
| Вишера, Велс, Язьва | 19.9 | – | 0.19 | 0.14 |
| Нора, Бурунда, Селемджа | 21.2 | 0.48 | – | 0.59 |
| Горин | 23.4 | 0.46 | 0.78 | – |
Примечание. Над главной диагональю – индекс сходства Съеренсена – Чекановского для количественных данных (обилие видов), под главной диагональю – индекс сходства Съеренсена для качественных данных (видовой состав).
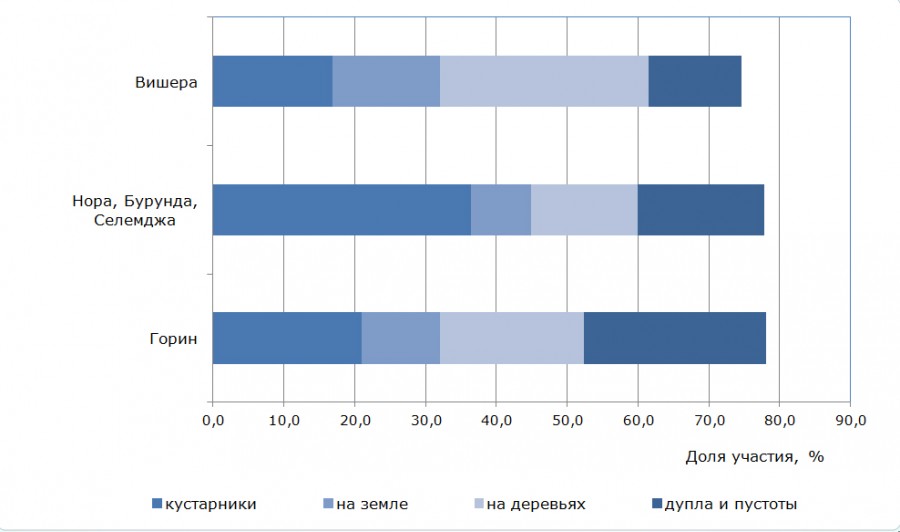
Рис. 4. Распределение воробьинообразных птиц по местам размещения гнезд
Fig. 4. The distribution of birds of the order of passerines by the locations of nests
Таблица 2. Доли отрядов и семейств птиц по встречаемости в %
| № | Отряды | Семейства | Вишера | Нора | Горин |
| 1 | Passeriformes | Sylviidae | 32.6 | 20.2 | 11.5 |
| 2 | Fringillidae | 23.9 | 15.4 | 12.1 | |
| 3 | Emberizidae | 1.7 | 12.2 | 12.2 | |
| 4 | Turdidae | 5.4 | 6.5 | 4.1 | |
| 5 | Paridae | 0.6 | 5.8 | 10.7 | |
| 6 | Sittidae | 0.1 | 1.2 | 2.4 | |
| 7 | Aegithalidae | 0.1 | 1.9 | 2.4 | |
| 8 | Motacillidae | 2.5 | 1.6 | 1.4 | |
| 9 | Corvidae | 4.1 | 1.5 | 3.7 | |
| 10 | Campephagidae | 0 | 2.5 | 3.9 | |
| 11 | Zosteropidae | 0 | 1.3 | 2.4 | |
| 12 | Muscicapidae | 0.1 | 7.5 | 9.4 | |
| 13 | Hirundinidae | 3.3 | 0.01 | 0 | |
| 14 | Прочие | 0.3 | 0.4 | 1.6 | |
| Сумма | 74.6 | 78.0 | 77.8 | ||
| 15 | Charadriiformes | 16.8 | 6.9 | 8.4 | |
| 16 | Anseriformes | 2.5 | 5.8 | 4.2 | |
| 17 | Cuculiformes | 2 | 4.1 | 1.5 | |
| 18 | Falconiformes | 1 | 1.3 | 1.2 | |
| 19 | Piciformes | 0.4 | 1.4 | 1.6 | |
| 20 | Columbiformes | 0.4 | 0.2 | 0.8 | |
| 21 | Coraciiformes | 0 | 1.2 | 1.4 | |
| 22 | Gruiformes | 1 | 0.1 | 0.5 | |
| 23 | Прочие | 1.4 | 1.0 | 2.6 |
По табл. 2 следует отметить, что семейства и даже отряды птиц представлены в достаточно высокой степени экологически близкими видами, и поэтому различия в систематической структуре сообществ отражают также и различия в их экологической структуре. Различия между выборками оценивались по абсолютным значениям встречаемости различных систематических групп птиц и оказались статистически значимы (Хи-квадрат = 5127.34, df = 44, p < 0.001). Для оценки вклада разных групп птиц в наблюдаемые различия использован метод главных компонент (рис. 5).
С первой компонентой связано 83.41 % дисперсии. Основной вклад в нее вносят доминирующие группы птиц: славковые, вьюрковые, ржанкообразные. Можно утверждать, что первая компонента – это характеристика доминирования. Однако следует отметить, что доля этих групп существенно выше на Вишере. Поэтому первая компонента, на наш взгляд, характеризует и различия в обилии доминирующих групп. Вторая компонента объясняет 10.87 % дисперсии. Основной вклад в нее вносят группы овсянок, синиц, мухоловок, дятлообразных. Она характеризует различия в обилии этих групп – их заметно меньше в первом сообществе (р. Вишера), чем в двух других. Остальные группы оказываются вблизи 0 и не вносят существенного вклада в наблюдаемые различия.
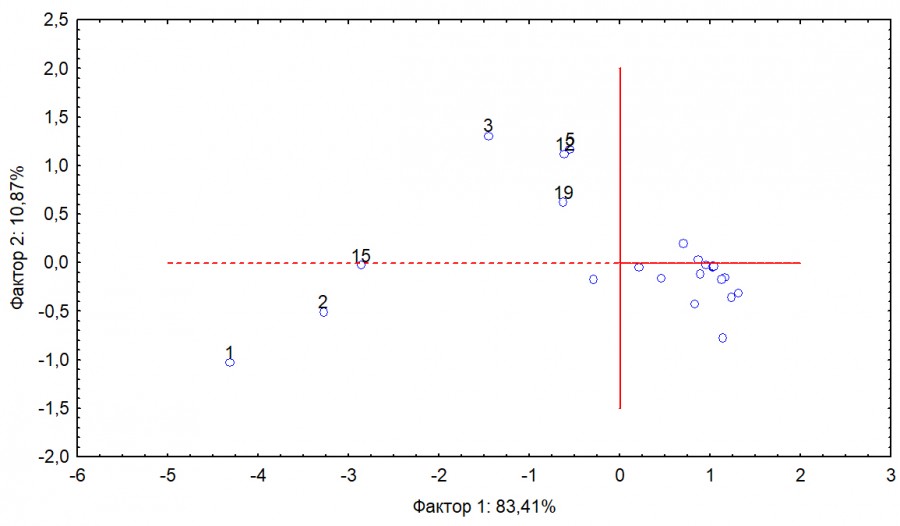
Рис. 5. Ординация систематических групп птиц (отрядов и семейств) в осях двух первых главных компонент. Цифры соответствуют номерам групп в табл. 2
Fig. 5. Ordination of systematic groups of birds (orders and families) in the axes of the first two main components. The numbers correspond to the group numbers in Table 2
Таким образом, с первым фактором связаны многочисленные группы, доля которых на Вишере примерно вдвое больше, чем на Дальнем Востоке. Со вторым фактором – группы, доля которых на Вишере существенно меньше, чем на Дальнем Востоке.
На Урале заметно выше доля Sylviidae, Fringillidae, Hirundinidae и отряда Charadriiformes. На Дальнем Востоке выше доля Emberizidae, Paridae, Sittidae, Aegithalidae, Muscicapidae и отряда Piciformes. Доля прочих групп достаточно низка, и вариации их присутствия в выборках могут все же оказаться случайными и незначимыми.
Доля представителей отряда воробьинообразных Passeriformes в целом оказалась очень близкой на всех трех сравниваемых участках: 74.6 % на р. Вишере, 78.0 % в бассейне р. Селемджи и 77.8 % на р. Горин. Естественно, что это наводит на мысль о высоком сходстве состава сообществ птиц исследуемых регионов при данном уровне усреднения видовых показателей.
Обсуждение
Самым массовым семейством на реках Вишера и Селемджа были славковые Sylviidae (см. рис. 5), на р. Горин они третьи по значимости. При этом необходимо отметить, что в Приамурье отсутствуют представители рода Sylvia, играющие заметную роль в пойменных комплексах Урала и имеющие европейское (средиземноморское) происхождение. На Дальнем Востоке, по всей видимости, их экологические ниши в значительной мере занимают пеночки Phylloscopus, которые имеют наибольший вес в рамках этого семейства в трех регионах, а также мухоловки Muscicapidae. Доля пеночек составила на р. Вишере 21.9 % (доминант пеночка-таловка Ph. borealis 8%); на р. Норе 18.5 % (доминант толстоклювая пеночка Ph. schwarzi 8.4 %); на р. Горин 10.6 % (доминант светлоголовая пеночка Ph. coronatus 7.3 %).
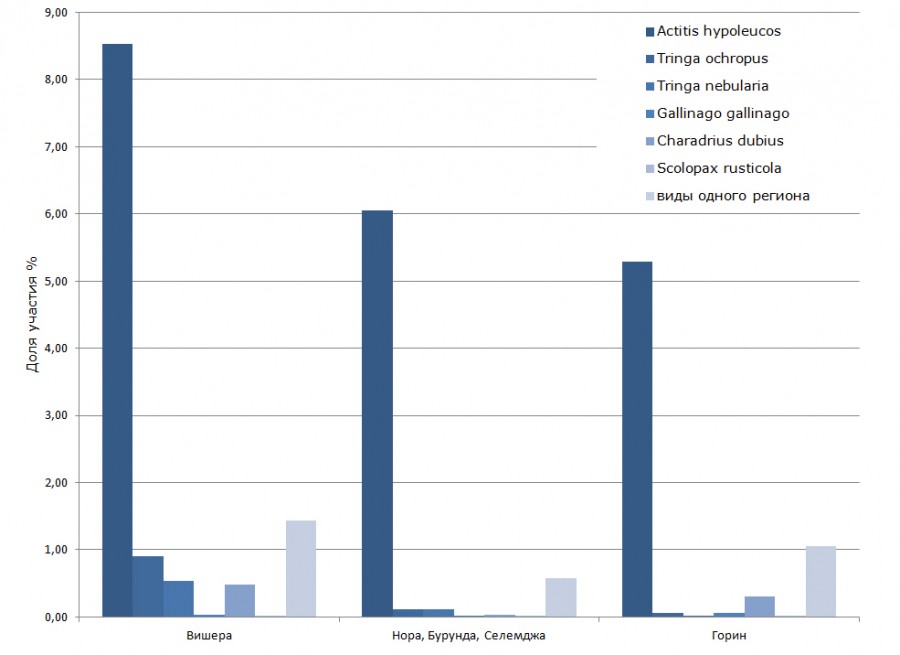
Рис. 6. Распределение птиц подотряда куликов по обилию
Fig. 6. Distribution of bird’s suborder of waders by their abundance
Наиболее массовым видом пойм Приамурья является седоголовая овсянка Ocyris spodocephala с долей участия 12 % в Норском и 9.9 % в Комсомольском заповедниках. По результатам учетов в пойменных комплексах, овсянковые Emberizidae в целом имеют высокую значимость для Приамурья и низкую для Вишерского Урала (см. табл. 2, рис. 5).
Еще значительнее были различия в рамках семейства мухоловковых. Вероятно, такая разница в значимости представителей этого таксона связана как с широтными, так и с меридиональными различиями регионов. Как известно, видовое богатство и численность птиц снижается к северу и югу от зоны широколиственных лесов (Гладков, 1958). В Приамурье желтоспинная мухоловка Ficedula zanthopygia – фоновый вид пойменных лесов: 5.5 % на р. Селемдже и 8.9 % на р. Горин. На р. Вишере же можно говорить только о присутствии 3 видов мухоловок. По всей видимости, в показателях обилия мухоловок проявляется прямая связь этой группы с широколиственными и смешанными лесами: на р. Вишере широколиственные породы отсутствуют как таковые, а в долине р. Горин они нередко доминируют в фитоценозах. Кроме того, возможно, высокая значимость мухоловок компенсирует отсутствие славок в Приамурье, экологические ниши которых отчасти заняли представители этой группы.
Доля семейства синиц Paridae на Урале также значительно ниже, чем в Приамурье (см. табл. 2), возможно, здесь нашло отражение снижение численности пухляка Parus montanus, проявившееся в Европе и значительной части Сибири в последние десятилетия (Преображенская, 2017). Вероятно, выявленные различия в значимости группы дендрофильных видов (синицы, поползни, длиннохвостые синицы) связаны как с широтными различиями, так и с большей нарушенностью лесов Приамурья: в Норском заповеднике свыше 90 % лесов пройдены пожарами, в Комсомольском – свыше 30 %.
Показатели обилия семейства дроздовых Turdidae (см. табл. 2) были весьма близки. Очевидно, что представители этого таксона выполняют в пойменных сообществах сравниваемых регионов одинаковую роль, при этом дрозды Приамурья, имеющие маньчжурское и сибирское происхождение, являются аналогами преимущественно европейских дроздов бассейна р. Вишеры. Единственным общим видом, который регистрировался в пойменных комплексах Урала и Приамурья, был пестрый дрозд Zoothera varia.
В рамках семейства трясогузок Motacillidae изучаемые регионы показали высокое сходство, причем 3 вида встречены как на Урале, так и в Приамурье: это белая трясогузка Motacilla alba, горная трясогузка M. cinerea и пятнистый конек Anthus hodgsoni, хотя последний вид на р. Вишере только присутствует (менее 0.1 %). По всей видимости, такая картина объясняется привязанностью трясогузок к околоводным стациям. А группировка околоводных птиц вследствие преобладания видов-транспалеарктов весьма сходна на востоке и на западе таежной зоны Евразии. Эта тенденция также заметна в рамках подотряда куликов и в других таксонах.
Отличия в рамках отряда наиболее наглядно проявляются в отсутствии на Урале представителей экзотических семейств личинкоедов Campephagidae и белоглазок Zosteropidae, которые обычны в поймах рек Селемджа и Горин. Причем в пойме р. Горин, в самом южном регионе, доля этих таксонов наиболее высока, что, видимо, коррелирует с высокой значимостью в местных долинных лесах широколиственных пород деревьев.
Анализ сообществ птиц в рамках отряда воробьинообразных по местам размещения гнезд (см. рис. 4) показал, что в наиболее страдающих от пожаров поймах Норского заповедника доминируют виды, гнездящиеся в кустарниках и дуплах. Наибольшая доля дуплогнездников выявлена в самом южном – Комсомольском – заповеднике. А в наиболее сохранившихся пойменных лесах Вишерского заповедника преобладают виды, гнездящиеся в кронах деревьев.
Доля представителей отряда ржанкообразных Charadriiformes на р. Вишере выше, чем на реках Селемджа и Горин (см. табл. 2). При этом необходимо отметить, что помимо подотряда куликов Limicoli и широко распространенных речных крачек Sterna hirundo (0.7 %) на р. Вишере обычны на гнездовании сизые чайки Larus canus (4.1 %), которых на реках Приамурья отмечали только на пролете. Очевидно, это связано с широтными различиями сравниваемых территорий. Поэтому в рамках данного отряда целесообразно сопоставление показателей обилия куликов, суммарная доля которых составила на р. Вишере 11.9 %, на р. Норе 6.9 %, на р. Горин 6.8 %. Везде доминировал перевозчик Actitis hypoleucos (рис. 6), встречаемость его на р. Вишере составила 13.6 пары на 10 км реки, на р. Норе – 10.9, на р. Горин – 10.2. Очевидно, что сообщества куликов имеют весьма сходный состав: кроме перевозчика в той или иной степени представлен еще один речной кулик – малый зуек Charadrius dubius, также повсеместно встречаются большой улит Tringa nebularia и черныш T. ochropus – виды пойменных лугов и болот, которые регулярно посещают реки. Достаточно значимы виды, характерные для конкретного региона: для р. Вишера это кулик-сорока Haematopus ostralegus 0.6 % и мородунка Xenus cinereus 0.5 %; для р. Нора дальневосточный кроншнеп Numenius madagascariensis 0.6 %, для р. Горин лесной дупель Gallinago megala 1.1 %.
На р. Вишере отсутствуют ракшеобразные Coraciiformes, которые в Приамурье представлены зимородком и широкоротом. Очевидно, что это различие имеет чисто широтный характер.
Заметной была разница в значимости представителей дятлообразных Piciformes: на р. Вишере их оказалось меньше, чем на реках Селемджа и Горин (см. табл. 2, рис. 5). Данные наземных учетов показывают по этой группе меньшую разницу.
Заключение
Сравнение орнитокомплексов пойм Вишерского Урала и Северного Приамурья показало, что неоднородность сообществ в первую очередь определяется разницей регионов в теплообеспеченности. В Вишерском заповеднике, расположенном в средней и северной тайге, преобладают вьюрковые, славковые и ржанкообразные. В Приамурье высока значимость овсянок, мухоловок и других дендрофильные видов, что связано с более южным расположением и распространенностью широколиственных лесов в регионе.
Широтные различия участков отображает индекс полидоминантности: в наиболее северном – Вишерском – заповеднике значения минимальные, в самом южном – Комсомольском – максимальные.
Меридиональные различия в степени континентальности, во влагообеспеченности проявляются в видовом составе сообществ, причем среди околоводных птиц эта разница минимальна. На уровне таксонов различие обозначилось через наличие в Приамурье двух экзотических семейств.
Библиография
Бигон М., Харпер Дж., Таунсенд К. Экология. Особи, популяции и сообщества . М.: Мир, 1989. Т. 2. 477 с.
Воробьев К. А. Птицы Уссурийского края . М., 1954. 360 с.
Гладков Н. А. О географической изменчивости численности видов и численности особей // Проблемы зоогеографии суши. Материалы совещания по зоогеографии суши. Львов, 1958. С. 57–63.
Кистяковский А. Б., Смогоржевский Л. А. Новые данные о границах китайского орнитофаунистического комплекса по р. Бурее // Научные доклады высшей школы. Биол. науки. М., 1964. № 3. С. 26–29.
Кистяковский А. Б. , Смогоржевский Л. А. Материалы по фауне птиц Нижнего Амура // Вопросы географии Дальнего Востока. Хабаровск, 1973. Сб. 11. С. 182–224.
Колбин В. А. Орнитогеографический обзор населения птиц Комсомольского и Норского заповедников // Сборник статей к 10-летию Норского заповедника. Благовещенск; Февральск: ОАО «ПКИ «Зея», 2008. С. 92–106.
Колбин В. А. Географический обзор населения птиц заповедника «Вишерский» // Особо охраняемые природные территории в жизни региона: Материалы межрегион. конф. Пермь, 2011.С. 75–83.
Колбин В. А. Сообщества птиц Вишерского заповедника и сопредельных территорий // Русский орнитологический журнал. 2016. Т. 25. Экспресс-выпуск 1376. С. 4767–4785.
Куренцов А. И. Зоогеография Приамурья . М.; Л.: Наука, 1965. 154 с.
Песенко Ю. А. Принципы и методы количественного анализа в фаунистических исследованиях . М.: Наука, 1982. 287 с.
Преображенская Е. С. Меридиональная изменчивость летнего населения птиц южной тайги // Размещение и численность позвоночных Сибири. Новосибирск: Наука, 1982. С. 48–69.
Преображенская Е. С. Динамика численности некоторых массовых зимующих птиц в лесах Европейской России с 1980-х по 2010-е годы // Динамика численности птиц в наземных ландшафтах. 30-летие программ мониторинга зимующих птиц России и сопредельных регионов. М.: Товарищество научных изданий КМК, 2017. С. 54–64.
Пронкевич В. В. Численность и распределение водоплавающих и околоводных птиц на реке Горин в поздневесенний период 2012 года // Амурский зоологический журнал. 2013. Т. V. № 2. С. 195–198.
Рябицев В. К. Птицы Сибири: Справочник-определитель: В 2 т. . М.; Екатеринбург: Кабинетный ученый, 2014. Т. 1. 438 с.
Czekanowski J. Zur differential Diagnose der Neandertalgruppe // Korrespbl. Dtsch. Ges. Anthropol. 1909. Bd. 40. S. 44–47.
Sørensen T. A new method of establishing groups of equal amplitude in plant sociology based on similarity of a species content and its application to analysis of the vegetation on Danich commons // Kgl. Dan. videnskab. selskab. biol. skr. 1948. Bd. 5. № 4. S. 1–34.

 © 2011 - 2026
© 2011 - 2026