



Введение
На весенний и весенне-летний периоды приходится период гнездования, успешность которого определяет уровень численности птиц в течение года. Поэтому выявление основных направлений изменчивости орнитокомплексов на разрезе от предгорий до высокогорий, описание по выявленным трендам неоднородности основных населенческих показателей представляет особый интерес. В анализ включены лидирующие виды птиц (по обилию и биомассе), плотность населения, суммарная биомасса, видовое богатство (общее и фоновое), а также фаунистический состав орнитокомплексов (с учетом обилия).
Материалы
Для полного цикла подобных работ проведены круглогодичные учеты птиц с 16 июня 1991 г. по 15 июня 1992 г. По результатам подсчета с помощью метода упорядоченной классификации отдельно для каждого уровня высот (высокогорья, среднегорья, предгорья) выявлены сезонные аспекты населения (Давранов, 2011). Границы их по уровням высот не всегда совпадают. Так, весенний аспект населения птиц в высокогорьях и среднегорьях начинается с начала апреля и заканчивается в конце того же месяца, в предгорьях он начинается во второй половине марта и завершается в конце апреля. Таким образом, весенний аспект в высокогорьях и среднегорьях длится один месяц, а в предгорьях – полтора. Весенне-летний период начинается во всех поясах с начала мая и длится в высокогорьях до середины июля, в среднегорьях – до конца июля, а в предгорьях – до конца июня. Короче всего этот период в предгорьях (два месяца), в среднегорьях – три месяца, а в высокогорьях – 2.5. Окончание периода определяет в предгорьях выгорание растительности, в высокогорьях – начало предосеннего похолодания. В среднегорьях максимальная продолжительность связана одновременным благополучием по двум факторам – тепло- и влагообеспеченности.
Методы
Весенние и весенне-летние учеты птиц проведены в 22 местообитаниях, в 18 из них – на маршрутах, в 4 – на обозреваемых участках (в малых населенных пунктах и кошарах). Всего весной с учетом пройдено 220 км, в весенне-летний период – 435 км. Учеты проведены без ограничения ширины трансекта с последующим пересчетом данных по среднегрупповым дальностям обнаружения (Равкин, Ливанов, 2008). В каждом местообитании с двухнедельной повторностью проходили по 5 км маршрута. Рассчитанные после этого показатели обилия усреднены в пределах выделенных сезонных аспектов населения. По этим данным на основе коэффициентов сходства Жаккара – Наумова, с помощью программы факторной классификации выявлены особенности среды, коррелирующие с неоднородностью населения птиц (Трофимов, 1976; Трофимов, Равкин, 1980), составлены классификации, по которым построены графы сходства. Для сравнения использованы результаты учетов птиц на Алтае (Цыбулин, 2009). Названия видов птиц даны по А. И. Иванову (1976), кроме чернозобого дрозда, седоголового щегла и маскированной трясогузки, которых, вслед за Л. С. Степаняном (2003), считали отдельными видами с соответствующими этому источнику названиями. По Б. К. Штегману (1938) даны названия типов фауны и их состав с некоторыми дополнениями.
В классификациях для каждого таксона указаны первые 5 лидирующих видов (по их доле в населении, %) и основные суммарные показатели сообществ – плотность населения (особей/км2 или 10 км береговой линии на водотоках), биомасса (кг/км2), видовое и фоновое богатство. Далее приведено соотношение преобладающих по числу особей типов фауны, представители которых составляют 10 % и более от общего обилия птиц. Затем эти показатели приведены перечислением в том же порядке без их наименования. Числа 1, 2...............11 означают номера типов, а 11.1; 12.1 и т. д. – номера подтипов.
Результаты
Неоднородность населения птиц в пространстве охарактеризована иерархической и структурной классификациями. Вторая из них выполнена на том или ином уровне разбиения, поэтому не является иерархической. Но она включает значимые связи выше выбранного порога и иллюстрирует основные тренды изменчивости орнитокомплексов в типологическом (факторном) пространстве.
Иерархическая классификация населения птиц в весенний период
Система населения незастроенной суши
Типы населения:
1 – высокогорный луговой (альпийских и субальпийских лугов; лидеры по обилию, % – гималайский вьюрок (Leucosticte nemoricola Hodgson, 1836) 32, горный конек (Anthus spinoletta Linnaeus, 1758) 15, бледная завирушка (Prunella fulvescens Severtzov, 1873) и красношапочный вьюрок (Serinus pusillus Pallas, 1811) по 9, клушица (Pyrrhocorax pyrrhocorax Linnaeus, 1758) 8 / лидеры по биомассе, %: клушица 33, темнобрюхий улар (Tetraogallus himalayensis G.R. Gray, 1843) 15, гималайский вьюрок(Leucosticte memoricolla Hodgson,1836) 11, кеклик (Alectoris kakelik J.E. Gray, 1830) 7, сизый голубь (Columba livia Linnaeus, 1789) 6; 276 / 18; 20 / 17; тибетского 59, монгольского 21, средиземноморского 11);
2 – высокогорный скально-стланиковый (арчовых стлаников, скал и осыпей; малая розовая чечевица (Carpodacus rhodochlamys J.F. Brandt, 1843) 25, черногрудая красношейка (Calliope pectoralis Gould,1837) 14, бледная завирушка 10, краснобрюхая горихвостка (Phoenicurus erythrogaster Guldenstadt, 1775) 8, красношапочный вьюрок 7 / кеклик 23, клушица 15, малая розовая чечевица (Corpodacus rodochlamus Brandt,1843) 13, черная ворона (Corvus corone Linnaeus, 1758) 10, сорока (Pica pica Linnaeus, 1758) 7; 435/34; 26/23; тибетского типа фауны 41, монгольского 27, китайского 15);
3 – среднегорный лесной (арчовых и еловых лесов; рыжешейная (Parus rufonuchalis Blyth,1849) и большая синицы (Parus major Linnaeus, 1758) 15 и 12, сорока 11, зарничка (Phylloscopus inornatus Blyth, 842) 9, желтогрудый князек (Parus flavipectus Severtzov, 1873) 8 / черная ворона 41, сорока 27, вяхирь (Columba palumbus Linnaeus, 1758) 9, черный дрозд (Turdus merula Linnaeus, 1758) 4, майна (Acridotheres tristis Linnaeus, 1766) 3; 159/14; 28/20; европейского типа фауны 42, китайского 22, средиземноморского 11, монгольского и сибирского по 10);
4 – среднегорный лугово-кустарниковый (лугостепей и кустарников со скалами и осыпями; черногорлая завирушка (Prunella atrogularis Brandt,1844) и черноголовый чекан (Soxicola toquata Linnaeus, 1766) по 11, горная овсянка (Emberiza cia Linnaeus, 1766) и теньковка (Phylloscopus collybita Vieillot,1817) по 9, лесной конек (Anthus trivialis Linnaeus, 1758) 8 / бородатая куропатка (Perdix daurica Pallas, 1811) 18, кеклик 14, черная ворона 9, сорока 7, фазан (Phasianus colchicus Linnaeus, 1758) 6; 752/46; 43/36; европейского типа фауны 24, монгольского 17, транспалеарктов 12, средиземноморского типа 11);
5 – предгорный садово-степной (степей и яблоневых садов; чернозобый дрозд 30, седоголовый щегол (Carduelis caniceps poronisi Vigors, 1831) 18, обыкновенная овсянка (Emberiza citrinella Linnaeus, 1758) 9, зяблик (Fringilla coelebs Linnaeus, 1758) 5, белошапочнaя овсянка (Emberiza leococephalos S.G. Gmelin, 1771) 3 / чернозобый дрозд (Turdus atrogularis Jarocki, 1819) 36, сизый голубь 10, грач (Corvus frugilegus Linnaeus, 1758) 8, бородатая куропатка 6, кеклик 5; 913/67; 42/34; европейского типа фауны 44, сибирского 35);
6 – предгорный пойменно-лесной (полевой воробей 29, майна 9, белошапочная овсянка 7, коноплянка (Cannabina cannabina Linnaeus, 1758) 5, горная овсянка 4 / фазан 25, майна 14, кеклик 11, сизый голубь и полевой воробей (Passer montanus Linnaeus, 1758) по 8; 1107/94; 37/34; транспалеарктов 32, европейского типа фауны 25, сибирского 11);
7 – предгорный полупустынный (каменка-плясунья 29, грач 23, обыкновенная овсянка 12, галка 11 и каменка-плешанка 8 / грач 67, галка 16, каменка-плясунья 6, обыкновенная овсянка 3 и полевой жаворонок 2; 172/24; 14/11; европейского и монгольского типов фауны по 36, транспалеарктов – 18);
8 – предгорный полевой (полей клевера и пшеницы; степной и полевой жаворонки 25 и 21, скворец 15, горный конек 11, белошапочная овсянка 8 / степной жаворонок 30, скворец 23, полевой жаворонок 15, черная ворона 6, горный конек 5; 357/18; 18/16; средиземноморского типа фауны 26, европейского 25, транспалеарктов 21, сибирского типа 14, тибетского 12).
Система населения застроенных местообитаний
Типы населения:
9 – высокогорных поселков (гималайский вьюрок 31, сорока 17, красношапочный вьюрок 16, сизый голубь 8, бледная завирушка 6 / сорока 33, черная ворона 24, сизый голубь 20, гималайский вьюрок 6,клушица 5; 939/105; 13/13; тибетского типа фауны 31, средиземноморского 25, европейского 21);
10 – среднегорных поселков (сизый голубь 38, майна 20, сорока 20, красношапочный вьюрок 10, черная ворона 8 / сизый голубь 48, черная ворона 21, сорока 19, майна 11, красношапочный вьюрок 0,4 ; 2000/460; 7/7; средиземноморского типа 50, европейского 27);
11 – предгорных поселков и кошар (домовый (Passer domesticus Linnaeus, 1758) и полевой воробьи 35 и 33, майна 7, скворец 6, малая горлица (Streptopelia senegalensis Linnaeus, 1766) 3 / домовый воробей 17, грач 16, майна 14, полевой воробей 12, сизый голубь 9; 7042/450; 40/38; транспалеарктов 70, европейского типа фауны 14);
Подтипы населения:
11.1 – крупных поселков (домовый и полевой воробьи 38 и 36, скворец и майна по 7, сизый голубь 2 / домовый и полевой воробьи 22 и 16, майна 16, сизый голубь 13, скворец 11; 8772/465; 37/36; транспалеарктов 76, европейского типа фауны 10);
11.2 – малых поселков и кошар (домовый и полевой воробьи по 20, грач 13, зяблик 10, майна 8 / грач 43, кольчатая горлица (Streptopelia decaocto Frivaldszky, 1838) 11, черная ворона 10, майна 9; 3583/420; 16/16; транспалеарктов 40, европейского типа фауны 37).
Система населения водно-околоводных сообществ
12 – Речной тип населения (горная и маскированная трясогузки 35 и 32, бурая оляпка (Cinclus pallasii Temminck, 1820) и оляпка (Cinclus cinclus Linnaeus, 1758) 18 и 13, синяя птица (Myophonus caeruleus Scopoli, 1786) 2 / бурая оляпка 36, маскированная трясогузка (Motacilla percenata Gould, 1861) 20, оляпка 19, горная трясогузка (Motacilla cinerea Tunster,1771) 16, синяя птица 9; 839/32; 5/5; транспалеарктов 48, средиземноморского типа фауны 32, китайского 20);
Подтипы населения:
12.1 – высокогорных и среднегорных рек (горная и маскированная трясогузки 44 и 19, бурая оляпка и оляпка 19 и 15, синяя птица 3 / бурая оляпка и оляпка 36 и 22, горная трясогузка 18, синяя птица 12, маскированная трясогузка 11; 854/35; 5/5; транспалеарктов 60, китайского типа фауны 22, средиземноморского 19);
12.2 – предгорных рек (маскированная и горная трясогузки 60 и 18, бурая оляпка и оляпка 15 и 7 / маскированная трясогузка 43, бурая оляпка и оляпка 36 и 12, горная трясогузка 9; 808/27; 4/4; средиземноморского типа фауны 60, транспалеарктов 25, китайского типа 15).
Иерархическая классификация населения птиц в весенне-летний период
Система населения незастроенной суши
Типы населения:
1 – высокогорный луговой (альпийских и субальпийских лугов; гималайский вьюрок 35, горный конек и коноплянка по 8, красношапочный вьюрок 7, каменка (Oenanthe oenanthe Linnaeus, 1758) 6 / клушица 23, темнобрюхий улар 22, гималайский вьюрок 13, альпийская галка (Graculus graculus Linnaeus, 1766) 6, черная ворона 5; 550/34 ; 50/27; тибетского типа фауны 58, монгольского 14, европейского 11);
2 – высокогорно-среднегорный лесово-кустарниковый (арчовых и еловых лесов, кустарников со скалами и осыпями; зарничка 16, обыкновенная чечевица (Carpodacus erytrinus Pallas, 1770) и серая славка (Silvia communis Latham,1787) по 8, зеленая пеночка (Phylloscopus trochiloides Sandr. 1837) 7, красношапочный вьюрок 6 / черная ворона 23, кеклик 15, сорока 8, майна 6, клушица 5; 717/48; 88/49; китайского типа фауны 24, европейского 23, сибирского 17, тибетского и монгольского по 10);
Подтипы:
2.1 – высокогорный скально-стланиковый (скал, осыпей и арчовых стлаников; черногрудая красношейка 12, малая розовая чечевица и красношапочный вьюрок по 11, зарничка 8 / кеклик 29, клушица 13, сорока 9, черная ворона 8, малая розовая чечевица 6; 630/47; 62/37; китайского типа фауны 27, тибетского 22, монгольского 18, средиземноморского и европейского по 11);
2.2 – среднегорный лесо-кустарниковый (лесов и кустарников; зарничка 21, серая славка 11, зеленая пеночка 9, чечевица 7, майна 4 / черная ворона 33, майна 9, фазан 8, сорока 7, кеклик 6; 775/48; 69/46; европейского типа фауны 30, китайского 22, сибирского 21).
Типы населения:
3 – среднегорный лугово-степной (серая славка 22, черноголовый чекан 13, желчная (Emberiza bruniceps Blyht, 1841) и горная овсянки 12 и 11, коноплянка 10 / кеклик 46, сорока 7, серая славка 6, черная ворона и желчная овсянка по 5; 548/33; 42/26; европейского типа фауны 38, транспалеарктов и средиземноморского типа по 32, монгольского 23);
4 – предгорный лесостепной (степей, пойменных лесов и полей пшеницы; полевой, домовый и черногрудый воробьи (Passer hispaniolensis Temminck, 1820) 14, 7 и 4, южный соловей (Luscinia megarhynchos Ch. L. Brehm, 1831) 6, желчная овсянка 5 / фазан 14, сизый голубь 11, майна 9, полевой воробей и скворец по 5; 1057/67; 71/57; европейского и средиземноморского типов фауны по 32, транспалеарктов 25);
Подтипы:
4.1 – лесостепной (лесов и степей; полевой воробей 11, южный соловей 8, черногрудый и индийский воробьи (Passer indicus Jardine et Selby, 1831) и седоголовый щегол по 6 / фазан 16, сизый голубь 11, майна 10, сорока 5, скворец 4; 1244/87; 68/57; европейского типа фауны 40, средиземноморского 32, транспалеарктов 16);
4.2 – полей пшеницы; полевой, домовый и черногрудый воробьи 29, 25 и 17, степной и хохлатый жаворонки (Galerida cristata Linnaeus, 1758) по 6 и 5 / домовый, полевой и черногрудый воробьи 20, 18 и 13, сизый голубь 11, степной жаворонок 10; 684/25; 22/19; транспалеарктов 56, средиземноморского типа фауны 34).
Типы населения:
5 – предгорный садовый (яблоневых садов; скворец 25, галка 18, чернолобый сорокопут (Lanius minor Gmelin, 1788) 15, желчная овсянка 12, сорока 8 / галка 31, скворец 17, сорока 15, черная ворона 12, чернолобый сорокопут 6; 368/41; 25/20; европейского типа фауны 58, транспалеарктов 23, средиземноморского типа 13);
6 – предгорный полупустынный (каменка-плясунья 36, хохлатый жаворонок и каменка по 10, степной конек (Anthus richardi Viellat, 1818) 9, маскированная трясогузка (Motacilla percenata Gould, 1861) 7 / черная ворона 26, каменка-плясунья 20, сизоворонка (Coracias garrulus Linnaeus, 1758) 10, хохлатый жаворонок 9, скворец 6; 129/6; 25/12; монгольского типа фауны 52, европейского типа и транспалеарктов – по 14, средиземноморского 12);
7 – предгорный полевой (полей клевера; желчная овсянка 25, полевой воробей 17, степной и полевой жаворонки 14 и 12, индийский воробей 8 / черная ворона 18, фазан и степной жаворонок по 15, желчная овсянка 11, скворец 8; 255/14; 22/16; средиземноморского типа фауны 48, транспалеарктов 34, европейского типа 14).
Система населения застроенных местообитаний
Типы населения:
8 – высокогорных поселков (майна 24, красношапочный вьюрок 22, сорока 19, черная ворона 10, гималайский вьюрок 6 / черная ворона 39, сорока 24, майна 18, сизый голубь 8, клушица 7; 408/69; 13/13; европейского типа фауны 32, средиземноморского 27);
9 – среднегорных поселков; (майна 31, сорока 21, маскированная трясогузка 9, сизый голубь 8, красношапочный вьюрок 7 / сорока 29, майна 23, черная ворона 20, сизый голубь 15, большая горлица (Streptopelia orientalis Latham, 1790) 6; 1943/317; 12/12; европейского типа фауны 26, средиземноморского 22);
10 – предгорных поселков и кошар (домовый и полевой воробьи 30 и 29, розовый скворец (Pastor roseus Linnaeus, 1758) 7, черногрудый воробей 6, грач 5 / грач 30, домовый и полевой воробьи 14 и 11, майна и розовый скворец по 8; 8480/554; 45/39; транспалеарктов 61, средиземноморского типа фауны 21 и европейского 12).
Система населения водно-околоводных сообществ
11 – речной тип (маскированная и горная трясогузки 32 и 30, бурая оляпка и оляпка 14 и 11, перевозчик (Aetitis hipoleucos Linnaeus, 1758) 8 / бурая оляпка 20, средний крохаль (Mergus serrator Linnaeus, 1758) 15, маскированная трясогузка 14, шилохвость (Anas acuta Linnaeus, 1758) 12, оляпка 11; 1227/67; 11/11; транспалеарктов 50, китайского типа фауны 15);
Подтипы населения:
11.1 – среднегорных и высокогорных рек (горная и маскированная трясогузки 33 и 26, бурая оляпка и оляпка 16 и 12, перевозчик 7 / бурая оляпка 21, средний крохаль 19, шилохвость 15, оляпка 12, маскированная трясогузка 10; 1333/82; 9/9; транспалеарктов 53, китайского типа фауны 19);
11.2 – предгорных рек (маскированная и горная трясогузки 49 и 22, перевозчик 13, оляпка и бурая оляпка по 7 /маскированная трясогузка 31, перевозчик 18, бурая оляпка 15, кряква 14, оляпка 11; 1015/38; 7/7; транспалеарктов 44).
Пространственно-типологическая структура орнитокомплексов в весенний и весенне-летний периоды
Классификации весеннего и весенне-летнего населения представлены тремя системами (надтипами), выделенными на графах в виде трех вертикальных рядов незастроенной и застроенной суши и рек. При этом первый ряд в весенний период имеет три отклонения, в весенне-летний – два. Они связаны с облесенностью и распашкой. Третий ряд в оба периода редуцирован до одного типа. Изменение населения птиц на незастроенной суше и в поселках четко совпадает с уровнями высот (рис. 1 и 2).
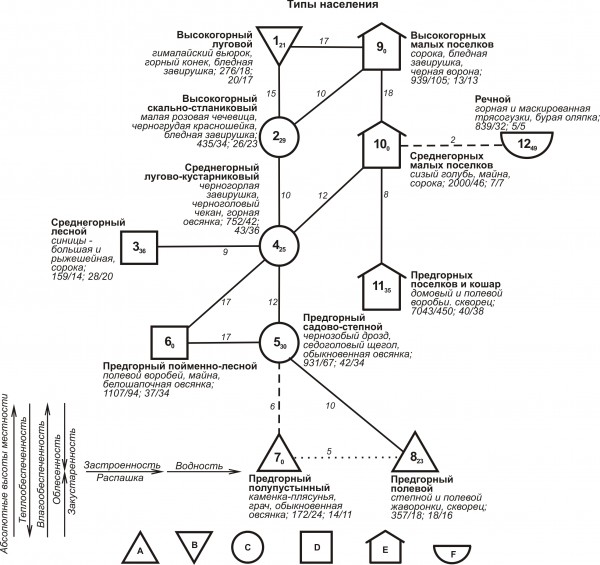
Рис. 1. Пространственно-типологическая структура весеннего населения птиц Киргизского хребта. A - обедненные открытые местообитания, B - богатые открытые, C - мозаичные местообитания, D – облесенные, E – поселки, F – водно-околоводные сообщества. Сплошной чертой показаны значимые связи, прерывистой – максимальные (при отсутствии значимых), пунктиром – дополнительные. Рядом со связями приведена их оценка. Цифры в значках – номера типов по классификации, рядом со значками – их названия, три лидирующих по обилию вида, плотность населения (особей/км2), биомасса (кг/км2), общее количество встреченных видов/ в том числе фоновых (обилие которых 1 и более особей/км2).
Fig. 1. Spatial-typological structure of the spring bird population of the Kyrgys Range
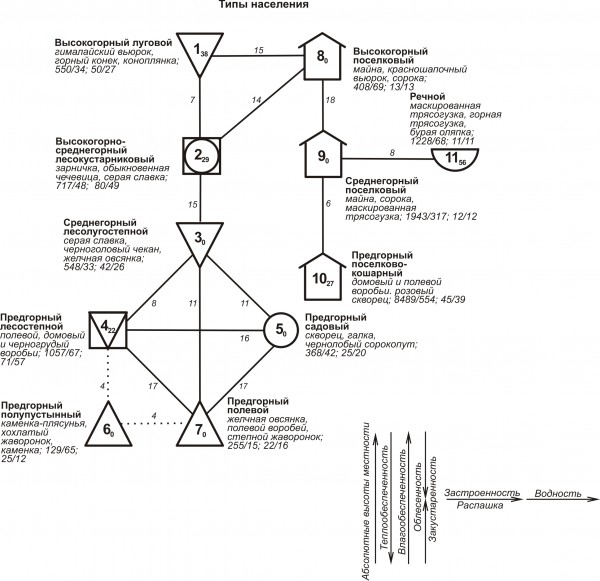
Рис. 2. Пространственно-типологическая структура населения птиц Киргизского хребта в весенне-летний период (условные обозначения - см. рис. 2)
Fig. 2. Spatial-typological structure of the spring-summer bird population of the Kyrgyz Range
Сила межклассовых связей обычно уменьшается к предгорьям, а внутриклассовых к высокогорьям. В поселках и на реках плотность и биомасса населения выше, чем на незастроенной суше, за исключением высокогорий весной, а видовое и фоновое богатство ниже.
Пространственно-типологическая организация населения птиц
Объяснение иерархической и структурной классификации позволяет выявить факторы среды и их неразделимые сочетания (природно-антропогенные режимы), по предметным соображениям определяющие изменчивость сообществ птиц. Для оценки силы и общности связи необходимо просчитать корреляцию этих факторов и режимов с изменчивостью орнитокомплексов, отраженных матрицей коэффициентов сходства. Эти оценки позволяют выявить иерархию влияния этих факторов и описать экологическую организацию сообществ.
Наиболее велика сила связи неоднородности населения в оба периода с водностью и несколько меньше с кормностью (таблица). С началом гнездования птиц в весенне-летний период возрастает значимость продуктивности, закустаренности и абсолютной высоты местности. Остальные факторы, указанные в таблице, корреллируют в существенно меньшей степени. Информативность иерархической классификации в оба периода равна 73 % дисперсии матрицы коэффициентов сходства, графов – 66 и 62 %, всех режимов и факторов вместе — 92 и 95 %.
Сила связи факторов среды и неоднородности весеннего и весенне-летнего населения птиц северного макросклона Киргизского хребта
| Фактор, режим | Учтенная дисперсия, период, % | |
| Весенний | Весенне-летний | |
| Водность | 31 | 32 |
| Кормность | 16 | 16 |
| Продуктивность | 8 | 14 |
| Абсолютные высоты местности | 7 | 12 |
| Застроенность | 6 | 5 |
| Закустаренность | 5 | 12 |
| Облесенность | 3 | 8 |
| Наличие скал и осыпей | 1 | 3 |
| Распашка | 1 | 3 |
| Все факторы | 80 | 81 |
| Режимы классификационные | 73 | 73 |
| структурные | 66 | 62 |
| Все режимы | 92 | 89 |
| Все факторы и режимы | 92 | 95 |
Обсуждение
Для первой половины лета по Алтаю выделено 7 типов населения: субнивальный, альпийско-тундровый, субальпийско-луговой, лесной, степной, синантропный и озерно-речной (Цыбулин, 2009). Кроме того, на подтиповом уровне разделены аридные и гумидные варианты высокогорных орнитокомплексов. На Киргизском хребте для весенне-летнего периода описано 11 типов населения: высокогорные луговой и скально-стланиковый, среднегорные – лесной, лесокустарниковый, лесолугово-степной и предгорные пойменно-лесной, лесостепной, садовый, полупустынный и полевой. Синантропный надтип выделен как на Алтае, так и на Киргизском хребте. Последний разделен на четыре типа по уровням высот. Лесной тип в обоих регионах номинально одинаков, а полупустынный присутствует только на Тянь-Шане. Из-за необследованности озер на Киргизском хребте по этой территории выделен только речной тип, а на Алтае – единый озерно-речной. Таким образом, по Киргизскому хребту полученное деление на типы более дробное, чем по Алтаю, причем не только из-за представленности полупустынного населения, но и из-за большей неоднородности характеристик орнитокомплексов.
Зимой население птиц Киргизского хребта делится на 10 типов (Давранов, 2016): два высокогорно-луговых (альпийский и субальпийский), два лесостепных (среднегорный и предгорный) и один полупустынный (предгорный). Население поселков разделено на четыре типа (высокогорный, среднегорный и два предгорных). Орнитокомплексы рек образуют единый тип. В предвесенний период выделено пять типов: высокогорный, высокогорно-среднегорный, среднегорно-предгорный, предгорный и речной. Уменьшение числа типов связано со значительным сходством в предвесеннее время ряда орнитокомплексов застроенных и незастроенных территорий и выделением по этой причине общих типов. Весной и в весенне-летний период деление на типы ближе к таковому в зимнее время, а не в предвесеннее. На Алтае наряду с влиянием абсолютных высот местности четко, хотя и на уровне подтипа, прослежено воздействие различий в увлажнении. Это приводит к парности сходных подтипов населения птиц – аридных и гумидных. В мозаичных и открытых местообитаниях на Алтае на уровне подтипа прослежены те же отличия, что и на Тянь-Шане. Синантропный тип сообществ на Алтае подразделен на два подтипа в соответствии с размерами населенных пунктов и степенью постоянства пребывания людей (стойбищный и поселковый). На Тянь-Шане на уровне типа прослежены отличия в зависимости от абсолютных высот местности. Суммарная информативность представлений о неоднородности весенне-летнего населения на Киргизском хребте выше, чем в первой половине лета на Алтае (81 и 66 %), так же как оценки для большинства общих факторов (обводненности, застроенности, распашки), кроме облесенности. Эти отличия связаны с объемом выборки, значительно большим по Алтаю, и соответственно с большей внутренней неоднородностью орнитокомплексов. Большую, чем на Тянь-Шане, оценку связи с облесенностью определяет наличие на Алтае настоящих высокополнотных лесов, в то время как на Тянь-Шане они сильно разрежены. Системы аппроксимирующих признаков среды по указанным режимам существенно неодинаковы из-за различий в размерах выборки, ее неоднородности и степени обобщения. С зимы до весны в основные структурообразующие факторы входят водность, застроенность и кормность. В весенне-летний период к ним добавляется продуктивность биоценозов – признак, сильно скоррелированный с кормностью и абсолютными высотами местности.
Состав преобладающих по типам населения видов на Алтае и Тянь-Шане существенно различен. Общими для обоих горных стран можно считать перевозчика, сизого голубя, горную и маскированную трясогузок, горного и степного коньков, черноголового чекана, каменку-плясунью, каменку, серую славку, зарничку, зеленую пеночку, гималайского вьюрка, чечевицу, домового и полевого воробьев, скворца, сороку и черную ворону. Для северного макросклона Киргизского хребта специфичны как лидеры в весенне-летний период: кеклик, чернолобый сорокопут, индийский и черногрудый воробьи, майна, клушица. Для Алтая это горбоносый турпан (Melanitta deglandi Bonaparte, 1850), хохлатая чернеть (Aythya fuligula Linnaeus, 1758), гоголь (Bucephala clangula Linnaeus, 1758), красноголовый нырок (Aythya ferina Linnaeus, 1758), полевой конек (Anthus campestris Linnaeus, 1758), варакушка (Cyanosylvia svecica Linnaeus, 1758), теньковка (Phylloscopus collybita Vieillot, 1817), пухляк (Parus montanus Bald, 1827) и бурая пеночка (Phylloscopus fuscatus Blyth, 1842). Отличия в составе лидеров связаны с большими абсолютными высотами Киргизского хребта и большей облесенностью Алтая, поэтому на Тянь-Шане в составе лидеров велико участие горных видов, а на Алтае – лесных и за счет обследованности озер – водоплавающих.
В весенне-летний период на Киргизском хребте плотность и биомасса населения выше, чем на Алтае (Давранов, 2014), а общее видовое богатство в 2–7 раза меньше на Киргизском хребте, но число фоновых видов почти одинаково. Количество встреченных видов, так же как фоновых, в весенне-летнее время в открытых бедных местообитаниях выше до 4 раз, чем зимних показателей. Для малых высокогорных и среднегорных поселков показатели почти одинаковы. В остальных местообитаниях общее число встреченных видов больше в 2–4 раза.
Плотность населения, биомасса, видовое и фоновое богатство, как правило, на Киргизском хребте возрастают с уменьшением абсолютных высот местности, и только в предгорьях в оба сезона значения закономерно меньше для полупустынь и полей. Показатели плотности зимнего населения по сравнению с весенне-летним периодом в бедных местообитаниях ниже в 7–14 раз, в закрытых – в 2–4 раза, а в предгорных малых поселках – в 6 раз. В высокогорных малых поселках и крупных поселках предгорий показатели почти одинаковые. На речках птиц в три раза больше. Показатели биомассы для многих местообитаний в 2–3 раза выше, чем зимой. Для высокогорных малых поселков, полей и полупустынь показатели почти не отличаются. В лесолуговых, степных и садовых местообитаниях суммарная биомасса птичьего населения меньше, чем зимой.
Число преобладающих по обилию фаунистических типов в населении обеих территорий почти одинаково: шесть на Алтае и семь на Тянь-Шане. В обоих горных регионах представители европейского типа фауны и транспалеаркты входят в число преобладающих во всех поясах. Птицы китайского типа фауны доминируют в среднегорьях и высокогорьях, а монгольского типа – в высокогорных и предгорных местообитаниях. Представителей сибирского типа фауны на Алтае много в среднегорьях и высокогорьях, а на Киргизском хребте – только в высокогорьях. Средиземноморские виды на Тянь-Шане доминируют во всех поясах, а на Алтае они не входят в число доминирующих. Представители тибетского типа в обоих регионах преобладают только в высокогорьях. Эти отличия связаны с большей облесенностью Алтая, меньшей степенью аридизации его и в целом меньшими абсолютными высотами местности. Весной на Киргизском хребте чаще встречаются представители европейского, средиземноморского, монгольского и китайского типов фауны, а также транспалеаркты. В первой половине лета на Алтае чаще отмечали птиц, отнесенных к транспалеарктам, китайскому, сибирскому и монгольскому типам фауны.
Межсезонные сравнения показывают, что транспалеаркты с зимы до весенне-летнего периода доминируют в среднегорных и предгорных местообитаниях. Доля их значительно меньше в высокогорьях. Тибетские, монгольские и китайские виды, напротив, чаще встречаются в высокогорьях и среднегорьях. Европейский тип фауны присутствует почти во всех местообитаниях, но больше их в среднегорных и предгорных местообитаниях. Средиземноморские формы в высокогорных местообитаниях встречаются часто, а в среднегорьях и предгорьях доминируют во многих местообитаниях.
Межгрупповое сходство населения птиц (в пересчете на одну значимую связь, отраженную на графе по Алтаю) втрое меньше, чем на Киргизском хребте, а внутригрупповое примерно такое же (4 и 18 %). Эти различия связаны, видимо, с большим объемом собранного по Алтаю материала и соответственно с большим разнообразием орнитокомплексов.
Заключение
Таким образом, границы распространения типов весеннего и весенне-летнего населения птиц совпадают с уровнем высот только для орнитокомплексов застроенной суши. Изменения внутри типов населения связаны с различиями в закустаренности, облесенности и распаханности, а также с составом агрокультур. Наибольшие показатели суммарного обилия и видового богатства свойственны среднегорно-предгорным местообитаниям. Водно-околоводное население птиц в весенний период с орнитокомплексами застроенных территорий имеет лишь слабую связь и значительную в весенне-летний период, зимой ее вообще нет. Население птиц сильно изменяется в предвесенний период, а в весенне-летний изменения незначительны. Зато увеличивается количество значимых структурообразующих факторов. На Алтае структура населения на типовом уровне почти не изменяется в течение года. И на Киргизском хребте (весенне-летний период), и на Алтае (в первой половине лета) транспалеаркты и представители европейского типа фауны преобладают во всех высотных поясах, а средиземноморского – только на Киргизском хребте. Тибетские птицы чаще встречаются на высокогорьях обоих регионов.
Библиография
Давранов Э. Границы сезонных аспектов населения птиц северного макросклона Киргизского хребта // Алтайский зоологический журнал. 2011. Вып. 5. С. 74–88.
Давранов Э. Сезоннная и высотно-поясная динамика населения птиц Киргизского хребта (Северный Тянь-Шань) // Вестник Бурятского государственного университета. Улан-Уде, 2014. Вып. 4. С. 36–43.
Давранов Э. Пространственно-типологическая структура и организация зимнего и предвесеннего населения птиц северного макросклона Киргизского хребта (Северный Тянь-Шань) // Сибирский экологический журнал. 2016. № 6. С. 900–910.
Иванов А. И. Каталог птиц СССР . Л.: Наука, 1976. 275 с.
Равкин Ю. С., Ливанов С. Г. Факторная зоогеография . Новосибирск: Наука, 2008. 184 с.
Степанян Л. С. Конспект орнитологической фауны России и сопредельных территорий (в границах СССР как исторической области) . М: ИКЦ «Академкнига», 2003. 808 с.
Трофимов В. А. Модели и методы качественного факторного анализа матрицы связи // Проблемы анализа дискретной информации. Новосибирск, 1976. Ч. 2. С. 24–36.
Трофимов В. А., Равкин Ю. С. Экспресс-метод оценки связи пространственной неоднородности животного населения и факторов среды // Количественные методы в экологии животных. Л., 1980. С. 113–115.
Штегман Б. К. Основы орнитогеографического деления Палеарктики. Фауна ССС. Птицы . Т. 1. Вып. 2. М.; Л.: Изд-во АН СССР, 1938. 157 с.
Цыбулин С. М. Птицы Алтая . Новосибирск: Наука, 2009. 236 с.
Благодарности
Все расчеты проведены с помощью пакета программ банка данных лаборатории зоологического мониторинга ИСиЭЖ СО РАН И. Н. Богомоловой и Л. В. Писаревской, рисунки выполнены Т. А. Кузнецовой. Автор искренне признателен им за помощь в работе, а также Ю. С. Равкину – за консультации и редактирование статьи.

 © 2011 - 2026
© 2011 - 2026