



Введение
Со времен Чарльза Элтона (Elton, 1924) проведено огромное количество исследований циклических процессов в популяциях различных видов животных, в том числе и птиц (см. обзор Lindström et al., 2001). У многих видов птиц хорошо известны межгодовые колебания численности и плодовитости (Klomp, 1970; Williams, 1985; Moss, Watson, 2001; Enemar et al., 2004; Nyholm, 2011). Как правило, они непосредственно или опосредованно (через погодные условия) связаны с флуктуациями кормовой базы и имеют адаптивный характер (Лэк, 1957; Klomp, 1970). В коррелятивной связи с начальной и конечной продукцией яиц находятся величина, линейные размеры и форма яйца, пигментация скорлупы (Мянд, 1988). Во многих исследованиях показана положительная связь качества и выживаемости птенцов с величиной яиц (Williams, 1994; Christians, 2002; Pinowska et al., 2004; Krist, 2011). Однако многолетняя динамика такого важного репродуктивного параметра птиц, как величина яиц, практически не изучена. Не ясна роль изменчивости репродуктивных показателей в динамике численности популяции. У воробьиных птиц популяционная регуляция за счет изменений величины кладки, по мнению В. А. Паевского (1985), маловероятна.
Целью нашего исследования было выявление характера многолетней цикличности величины кладки и ооморфологических показателей в популяции полевого воробья Passer montanus в окрестностях оз. Чаны (юг Западной Сибири).
Задачи исследования:
– оценка возможных связей между показателями;
– построение спектра ритмов динамики каждого показателя;
– расчеты периодов таких циклов и их мощности;
– оценка инвариантных периодик для хода каждого показателя;
– поиск близких по значению колебаний в природных местообитаниях для подстройки к ним популяционных ритмов полевого воробья.
Материалы
Материал собран в 1984–2005 гг. в Здвинском р-не Новосибирской области, на территории, прилегающей к юго-восточному побережью оз. Малые Чаны. Основная часть работы выполнена в двух ленточных осиново-березовых островных лесах-колках общей площадью 6 га, расположенных в районе устья р. Каргат.
Всего обследовано и находилось под наблюдением 1747 гнезд, промерено 7023 яйца.
На гнездах отловлена 671 взрослая особь, в том числе 135 брачных пар, окольцовано 4324 птенца.
Всего за годы исследований окольцовано свыше 9 тысяч особей. Большую часть из них отлавливали повторно, многих птиц неоднократно, из них 1035 особей зарегистрировано в последующие после кольцевания годы.
Методы
Размножение полевых воробьев изучали преимущественно с помощью развески искусственных гнездовий, в основном дощатых синичников с площадью дна от 100×100 до 120×120 мм и диаметром летка 32–40 мм, вывешенных на высоте 1.5 м. В разные годы под наблюдением было от 50 до 300 искусственных гнездовий. В период размножения птиц проводили регулярные осмотры синичников и систематические наблюдения за обнаруженными гнездами с целью выяснения сроков гнездования, величины кладки и успешности размножения. Дата начала откладки яиц в каждом гнезде определялась путем непосредственных наблюдений либо оценивалась в результате соответствующих расчетов, исходя из насиженности яиц, времени выклева или возрастных признаков птенцов.
Для оценки изменчивости величины яиц определяли с помощью штангенциркуля (точность деления 0.1 мм) их линейные размеры: длину (L) и максимальный диаметр (ширину) (B). Объем яиц вычисляли по формуле: V = 0.51 × L × B2 (Hoyt, 1979), а индекс формы (округленности) – по формуле: Sph = (B/L) × 100 (Мянд, 1988). При изучении изменчивости ооморфологических признаков за исходные данные приняты средние арифметические всех яиц в каждой кладке.
Гнездящихся птиц отлавливали руками на кладках или с помощью специальных опадных клапанов во время выкармливания птенцов. Отлов полевых воробьев местной популяции с целью кольцевания и контроля производился также с помощью переносных «паутинных» сетей.
Сведения о погодных условиях взяты на сайте World Weather (Climate ZDVINSK..., 2015). Статистическая обработка данных проведена с помощью программного пакета Statistica 6.1. Расчеты гармонических составляющих сделаны с использованием программ спектрального анализа, находящихся в собственности института. Кроме того, для этой же цели применялась авторская программа А. В. Тарновского «Harms». Временные ряды всех хронограмм исследовались на содержание скрытых гармонических составляющих. Были построены спектры ритмов динамики величины кладки и ооморфологических параметров, а также рассчитаны периоды и мощности гармонических составляющих каждого такого спектра. Оценка спектральной плотности (PSD) проводилась методом Уэлча (Марпл-мл., 1990). Отобраны наиболее устойчивые картины распределения спектральной плотности (мощности). При вычислении функции спектральной плотности с исследуемого ряда снимали тренд, чтобы обеспечить стационарность процесса. Все расчеты выполнены с помощью свободной программной системы GNU Octave (GNU Octave, 2013), которая использует совместимый с MATLAB язык высокого уровня и представляет интерактивный командный интерфейс для решения различных линейных и нелинейных математических задач. В частности, для оценивания PSD-методом Уэлча использовалась функция pwelch из пакета расширений Octave-Forge (Octave-Forge..., 2013). Для удобства работы был разработан интерфейс, позволяющий проводить обмен данными и всю обработку, включая выбор размера и количества окон, графическое представление данных и т. п., в диалоговом режиме.
Результаты
Величина кладки. Литературные сведения о межгодовой изменчивости величины кладки полевого воробья противоречивы. В Хорватии у полевого воробья существенные межгодовые различия в плодовитости не отмечены (Dolenec et al., 2011), тогда как в Ростовской области (Lebedeva, 1999) и в Испании (Garcia-Navas, Sanz, 2012) эти различия значимы. В районе наших исследований среднее число яиц в кладке полевого воробья варьирует, и в разные годы значения могут существенно различаться как между смежными годами, так и отдаленными друг от друга. По данным за 22 года, достоверные отклонения среднесезонной величины кладки от средней многолетней (5.50 ± 0.02 яица) отмечены 8 раз (Чернышов, 2011). Обусловленность таких различий не очевидна, и на хронограмме изменений величины кладки полевого воробья можно отметить только заметные колебания числа яиц (рис. 1). Некоторые изменения среднего числа яиц между смежными годами оказываются незначимыми. Чтобы выяснить, каковы периоды многолетних колебаний средней величины кладки, если они существуют, можно представить данные не на шкале времени, а на частотной шкале. Преобразовав хронограмму величины кладки у полевого воробья в спектр ритмов, находим, что на самом деле она имеет всего 4 значительные по мощности периодические составляющие, то есть это суперпозиция четырех циклов, один из которых обладает преобладающей мощностью, а три других незначительны и их мощность падает с увеличением частоты (см. рис. 1). Доминирующий цикл величины кладки у полевого воробья – 11-летний. Вероятно, он подстраивается к близким по периоду циклам динамики температуры воздуха во второй и третьей декадах мая (рис. 2а), когда к размножению приступает наибольшее количество пар. Поскольку полевые воробьи выкармливают птенцов преимущественно насекомыми, обилие которых зависит от температурных условий, такая подстройка обеспечивает особям и популяции в целом более высокую продуктивность в благоприятные годы. Менее выраженные 2–4-летние циклы величины кладки, скорее всего, обусловлены внутрипопуляционными причинами. У бицикличного полевого воробья среднесезонная плодовитость зависит и от соотношения первой и второй кладок. Доля вторых кладок изменяется с 3-летней и 12-летней (кратной 3-летней) периодичностью (рис. 2б). Возможно, это связано с трехлетней динамикой возрастного состава (таблица, рис. 2в), так как большинство годовалых особей полевого воробья моноцикличны, а первые кладки, как правило, крупнее вторых (Чернышов, 2011).
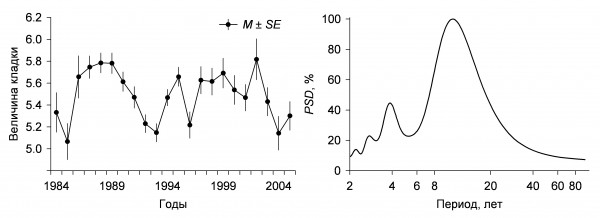
Рис. 1. Межгодовая динамика и спектр многолетних циклов величины кладки полевого воробья
Fig. 1. Interannual dynamics and spectrum of multi-year cycles of clutch size of the Tree Sparrow
Межгодовые различия в возрастной структуре (доли особей каждого возраста, %)
Interannual differences in age structure (percentage of individuals of each age)
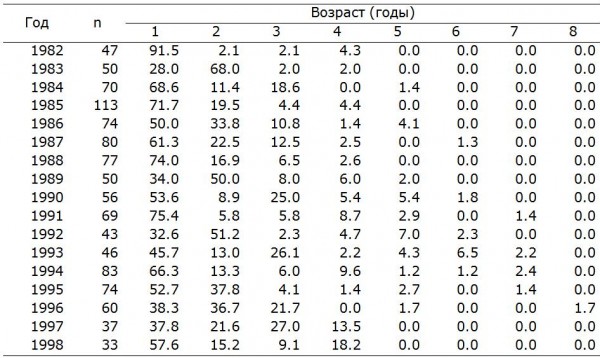
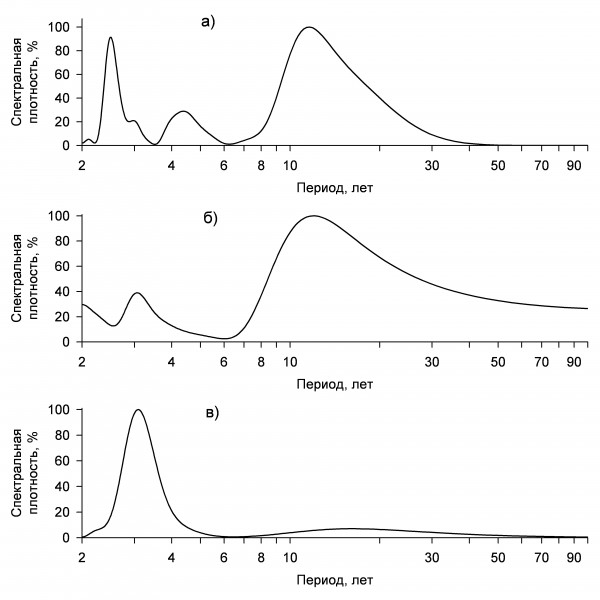
Рис. 2. Цикличность средней температуры воздуха во второй декаде мая (а), доли вторых кладок (б) и доли годовалых особей в популяции полевого воробья (в)
Fig. 2. Cyclicity of average air temperature in the second decade of May (а), the proportion of second clutches (б) and the proportion of one-year-old individuals in the population of the Tree Sparrow (в)
Морфология яиц. Ооморфологические показатели многих видов птиц в той или иной степени коррелируют с условиями сезона размножения, которые складываются в разные годы (Венгеров, 2001; Климов, 2003; Lebedeva, 1999). В Барабинской лесостепи достоверные отклонения среднесезонной длины яиц полевого воробья от средней величины за все годы зарегистрированы 5 раз, диаметра – 4 раза, объема – 8 раз. Существенных отклонений индекса округленности не отмечено. По мнению М. Громадского (Gromadzki, 1966), корреляции между длиной и шириной яиц не существует: ширина определяется величиной и возрастом птицы, а длина внешними условиями. П. Д. Венгеров (2001) также считает, что между признаками «длина» и «диаметр» яйца в процессе его формирования нет взаимозависимого роста и, следовательно, не может быть корреляции. Тот же ее обычно невысокий положительный уровень, регистрируемый при расчетах, есть результат опосредованной взаимосвязи через объем яйца. В чановской популяции полевого воробья, по данным анализа 1154 кладок, коэффициент корреляции между длиной и диаметром яиц составляет лишь 0.33, что соответствует данным, приводимым П. Д. Венгеровым по этому виду. Между линейными параметрами и объемом коэффициенты корреляции составляют 0.78 и 0.82 соответственно. Корреляция среднегодовых значений параметров яиц (n = 22) существенно выше: между длиной и диаметром r = 0.91, а между линейными показателями и объемом r = 0.97. Это свидетельствует о том, что все внутрипопуляционные ооморфологические параметры, по-видимому, регулируются одними и теми же факторами. Последняя из рассчитанных характеристик – динамика изменений индекса округленности яйца заметно отличается от изменений предыдущих показателей. Хронограмма ее имеет высокую, но отрицательную корреляцию с длиной яйца (r = -0.53).
Спектры ритмов многолетних изменений линейных размеров и объема яиц у полевого воробья во многом сходны (рис. 3). Здесь самая мощная примерно 7-летняя цикличность, имеются выраженные небольшие по мощности 3- и 2-летние периодические составляющие. В связи с тем что форма яйца – наиболее стабильный показатель, можно лишь предположить, что многолетние ритмы индекса округленности яиц близки по периодам к циклам величины кладки. Вероятно, цикличность морфологических параметров яиц, так же как и величины кладки, определяется совокупностью целого ряда внешних и внутрипопуляционных факторов (например, кормовой базы, возрастного состава популяции, сроков и продолжительности сезона размножения). Важнейшим из них, по-видимому, является погодный фактор, что подтверждается и литературными данными (Pinowska et al., 2002; Barkowska et al., 2003).
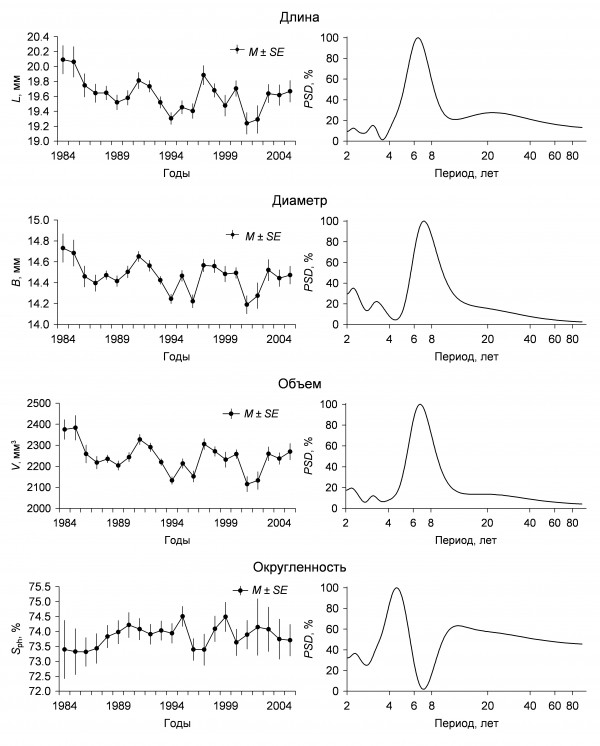
Рис. 3. Хронограммы и спектры многолетних циклов морфологических параметров яиц полевого воробья
Fig. 3. Chronograms and spectra of multi-year cycles of morphological parameters of Tree Sparrow eggs
Сравнивая фазовые характеристики ритмов исследуемых динамических процессов, мы обнаружили интересный артефакт. Циклы величины кладки находятся в противофазе с циклами ооморфологических характеристик. То есть зависимость конфигурации яиц от величины кладки не просматривается, и в то же время при совпадении циклов этих характеристик они всегда находятся в противофазе друг к другу.
Обсуждение
Итак, многолетняя динамика величины кладки и яиц полипериодична, и можно предполагать, что эти циклы имеют приспособительное значение. Популяционные циклы имеют эндогенное происхождение, но чтобы они не стали затухающими, должна быть постоянная связь с внешним устойчивым циклом (Ердаков, 1991). Они подстраиваются к определенным природным процессам, имеющим для них значение датчиков времени. Среди таких важных для всего живого природных ритмов описан цикл колебаний магнитного поля Земли с периодом в 7 лет (Мартынюк и др., 2006). К нему могут подстраиваться все параметры, связанные с объемом яиц. Кроме того, в структуре брикнеровского цикла выделяется 7-летняя, а также 20-летняя цикличности (Таранюк, 2000), которые также могут служить синхронизаторами цикличности значений ооморфологических показателей. Мощным синхронизирующим фактором для всего живого являются георитмы Земли, среди них описаны 7–12-летние, а также 19–22-летние (Якушев, 2002). Само озеро Чаны пульсирует с периодами 10–11 лет, 20–22 года и 45 лет, заметно изменяя климатический фон (Пульсирующее озеро Чаны, 1982). Естественно, к таким пульсациям адаптированы изменения многих характеристик у видов, живущих вблизи озера.
Заключение
- В динамике характеристик плодовитости полевого воробья имеются хорошо проявленные многолетние гармонические составляющие. Эти периодичности легко выявить, представив процессы динамики биологических характеристик на частотной шкале.
- Циклы характеристик не проявлены в хронограммах процесса, они замаскированы в сложной кривой динамики характеристики.
- Установление цикличности дает дополнительную биологическую информацию, если известны частотные характеристики различных популяционных изменений, а также циклы в местообитаниях рассматриваемой популяции.
Библиография
Венгеров П. Д. Экологические закономерности изменчивости и корреляции морфологических структур птиц . Воронеж: Изд-во Воронежского гос. ун-та, 2001. 248 с.
Ердаков Л. Н. Биологические ритмы и принципы синхронизации в экологических системах (хроноэкология) . Томск: Изд-во ТГУ, 1991. 216 с.
Климов С. М. Эколого-эволюционные аспекты изменчивости ооморфологических показателей птиц . Липецк: Изд-во Липецкого гос. пед. ун-та, 2003. 208 с.
Лэк Д. Численность животных и ее регуляция в природе . М.: Изд-во иностранной литературы, 1957. 404 с.
Марпл-мл. С. Л. Цифровой спектральный анализ и его приложения : Пер. с англ. М.: Мир, 1990. 584 с.
Мартынюк В. С., Владимирский Б. М., Темурьянц Н. А. Биологические ритмы и электромагнитные поля среды обитания // Геофизические процессы и биосфера. 2006. Т. 5. № 1. С. 5–23.
Мянд Р. Внутрипопуляционная изменчивость птичьих яиц . Таллин: Валгус, 1988. 192 с.
Паевский В. А. Демография птиц . Л.: Наука, 1985. 285 с.
Пульсирующее озеро Чаны . Л.: Наука, 1982. 304 с.
Таранюк М. И. Исследование цикличности климата и мониторинг атмосферных процессов на территории юго-востока Западной Сибири: Дис. … канд. геогр. наук . Томск: ТГУ, 2000. 116 с.
Чернышов В. М. Популяционная экология полевого воробья (Passer montanus) на юге Западной Сибири // Птицы Сибири: структура и динамика фауны, населения и популяций / Ред. Л. Г. Вартапетов. М.: Товарищество научных изданий КМК, 2011. (Труды Института систематики и экологии животных СО РАН. Вып. 47.) С. 197–223.
Якушев Д. И. Алгоритмы математического моделирования . СПб.: Поликом, 2002. 100 с.
Barkowska M., Pinowski J., Pinowska B. The effect of trends in ambient temperature on egg volume in the Tree Sparrow Passer montanus // Acta Ornithol. 2003. Vol. 38. № 1. P. 5–13.
Christians J. K. Avian egg size: variation within species and inflexibility within individuals // Biological Reviews. 2002. Vol. 77. P. 1–26.
Climate ZDVINSK. URL: http://en.tutiempo.net/climate/ws-297120.html (дата обращения: 27.08.2015).
Dolenec Z., Dolenec P., Møller A. P. Warmer springs, laying date and clutch size of tree sparrows Passer montanus in Croatia // Current Zoology. 2011. Vol. 57. Issue 3. P. 414−418.
Elton C. S. Periodic fluctuations in the numbers of animals: their causes and effects // British Journal of Experimental Biology. 1924. Vol. 2. P. 119–163.
Enemar A., Sjöstrand B., Andersson G., von Proschwitz T. The 37-year dynamics of a subalpine passerine bird community, with special emphasis on the influence of environmental temperature and Epirrita autumnata cycles // Ornis Svecica. 2004. Vol. 14. P. 63–106.
García-Navas V., Sanz J. J. Yearly and seasonal variation of breeding parameters in a declining multi-brooded passerine, the Tree Sparrow. // Ardea. 2012. Vol. 100. Issue 1. P. 79–88.
GNU Octave. URL: http://www.gnu.org/software/octave/ (дата обращения: 11.12.2013).
Gromadzki M. Variability of egg-size of some species of the forest birds // Ekol. Polska A. 1966. Vol. 14. P. 99–109.
Hoyt D. F. Practical methods of estimating volume and fresh weight of bird eggs // Auk. 1979. Vol. 96. № 1. P. 73–77.
Klomp H. The determination of clutch-size in birds. A review // Ardea. 1970. Vol. 58. Issue 1–2. P. 1–124.
Krist M. Egg size and offspring quality: a meta-analysis in birds // Biological Reviews. 2011. Vol. 86. P. 692–716.
Lebedeva N. V. Variability of clutch and egg sizes of the Tree Sparrow (Passer montanus) in South-Western Russia // Intern. Stud. Sparrows. 1999. Vol. 26. P. 48–57.
Lindström J., Ranta E., Kokko H., Lundberg P., Kaitala V. From arctic lemmings to adaptive dynamics: Charles Elton’s legacy in population ecology // Biological Reviews. 2001. Vol. 76. P. 129–158.
Moss R., Watson A. Population Cycles in Birds of the Grouse Family (Tetraonidae) // Advances in Ecological Research. 2001. Vol. 32. P. 53–111.
Nyholm N. E. I. Dynamics and reproduction of a nest-box breeding population of Pied Flycatcher Ficedula hypoleuca in a subalpine birch forest in Swedish Lapland during a period of 46 years // Ornis Svecica. 2011. Vol. 21. P. 133–156.
Octave-Forge – Extra packages for GNU Octave. URL: http://octave.sourceforge.net/signal/function/pwelch.html (дата обращения: 11.12.2013).
Pinowska B., Barkowska M., Pinowski J., Bartha A., Hahm K.-H., Lebedeva N. The effect of egg size on growth and survival of the Tree Sparrow Passer montanus nestlings // Acta Ornithol. 2004. Vol. 39. № 2. P. 121–135.
Pinowska B., Barkowska M., Pinowski J., Hahm K.-H., Lebedeva N. Influence of temperature on Tree Sparrow Passer montanus egg mass according to laying sequence // Intern. Stud. Sparrows. 2002. Vol. 29. P. 33–47.
Williams J. Statistical analysis of fluctuations in red grouse bag data // Oecologia. 1985. Vol. 65. P. 269–272.
Williams T. D. Intraspecific variation in egg size and egg composition in birds: effects on offspring fitness // 1994. Vol. 68. P. 35–59.
Благодарности
Работа поддержана программой ФНИ государственных академий наук на 2013–2020 гг. (проект № VI.51.1.6).

 © 2011 - 2026
© 2011 - 2026