



Введение
Ареал травяной лягушки (Rana temporaria L.), европейского вида, занимает центральную и северную Европу от горной системы Пиренеи в Испании и Франции до Урала. Европейско-сибирский вид лягушка остромордая (Rana arvalis Nilss.) распространен от севера Бельгии и, минуя Британские о-ва, горы центральной Европы и Фенноскандии, – на востоке достигает участка поймы р. Лена на 125° в. д. (Кузьмин, 2012; Glandt, 2014). R. temporaria встречается в шести природных областях: тундре, лесотундре, тайге, в смешанных и широколиственных лесах и в лесостепи. R. arvalis населяет те же ботанико-географические области, в лесостепи она обычна, заходит в степные районы Казахстана и горы Южной Сибири. Область, где можно встретить оба этих вида, простирается с запада на восток почти на 4 тыс. км, а в наиболее широкой части – по меридиану 33º в. д. – почти на 2 тыс. км. Наибольшей численности в пределах зоны симпатрии оба вида достигают в лесном поясе равнин Европы.
Единственная обстоятельная попытка сопоставить экологическую специфику этих двух видов предпринята почти 20 лет назад (Северцов и др., 1998). Проанализировав вероятные «точки пересечения интересов» на всех этапах годовых и жизненных циклов, авторы приходят к выводу, что «различия экологических ниш травяной и остромордой лягушек обусловлены различием путей их видообразования, а не конкурентным взаимодействием этих видов» (с. 295). За время, прошедшее с момента опубликования этой важной работы, с появлением значительного количества новых исследований, открылась возможность существенно дополнить и частично пересмотреть некоторые положения, выдвинутые А. С. Северцовым с коллегами.
Предварительное ознакомление с обширной литературой эколого-фаунистического плана, касающейся данных видов, давало основания судить о значительной степени их симбиотопии. Однако оставалось непонятным, почему, например, в пределах зоны симпатрии есть районы, где один вид отсутствует, тогда как другой успешно существует? Почему остромордая лягушка, избегающая гор Европы, обычна и местами обильна в горах Южной Сибири? Что позволяет популяциям этого вида населять как заболоченную Западно-Сибирскую равнину, так и засушливые степные районы от низовий Дона до предгорий Алтая? Почему границы ареалов двух видов, проходя по равнинам, не следуют видимыми физическими преградами? Наконец, чем вызван тот факт, что при суммарной протяженности ареалов, составляющей около 9000 км в субширотном направлении, их перекрывание не охватывает и половины этого пространства?
Целью настоящей работы является выяснение закономерностей размещения и ландшафтной приуроченности популяций двух видов лягушек в пределах их ареалов. Необходимо было выяснить, какие местообитания занимают травяные и остромордые лягушки по ходу трех основных этапов годичного цикла жизнедеятельности: зимовка, период икрометания, время нагула.
При сборе информации мы просмотрели большое количество доступных источников, включая интернет-ресурсы. В задачи входило обобщение и анализ разрозненных сведений по отдельным территориям зоны симпатрии двух видов, а также ключевым, на мой взгляд, участкам периферии их ареалов. Начиная с прошлого века (а где-то еще раньше) среда обитания «сухопутных» земноводных подверглась радикальным преобразованиям, что существенно повлияло на их распространение. В техногенных и современных агроландшафтах, на урбанизированных территориях приходится наблюдать лишь картину вынужденного размещения животных, отражающую скорее степень приспособленности популяций лягушек к экстремально изменившимся условиям. Поэтому при характеристике местообитаний и ландшафтной приуроченности лягушек в выбранных частях их ареалов мы старались максимально использовать сведения, содержащиеся в старых публикациях.
Основной объем собственного материала собран автором в Государственном природном заповеднике «Кивач» (Кондопожский район, Республика Карелия, средние координаты 62°17' с. ш., 33°55' в. д.) в 1980–2014 гг. В весенние сезоны разных лет предпринимали экспедиционные выезды в отдельные районы Карело-Кольского края. Это заповедник «Костомукшский» (64°30'–30°30'), участки Кандалакшского заповедника на побережье Белого (66°34'–33°09' и 66°45'–33°30') и Баренцева (69°00'–35°15' и 68°45'–37°15') морей, территория Лапландского заповедника (67°50'–32°00').
Аналитический обзор
Зимовка
Особи представителей Anura умеренной климатической зоны, в том числе R. temporaria, для которых подводная зимовка – видовой стереотип, не выносят длительного переохлаждения и мало устойчивы к замораживанию (Schmid, 1982; Pasanen, Karhaää, 1997 и др.). Иное – виды, способные зимовать вне водоемов. Исследования Rana (Lithobates) sylvatica, североамериканского аналога R. arvalis, показали, что этот вид в период спячки переносит охлаждение до –18 ºС. Это происходит благодаря, в частности, синтезу кроиопротекторов, препятствующих образованию летально травмирующих кристаллов льда в клетках и тканях организма (Storey, 1987; Costanzo et al., 2013 и др.). Важнейшее «сырье» для производства антифризов – накопленный за лето в печени гликоген. Выяснилось, что в южной Карелии относительный вес печени у остромордой лягушки к осени почти вдвое превышает таковой у травяной (Кутенков, 1991), что является следствием накопления значительно бόльших запасов гликогена первым видом. Наивысшие для R. arvalis значения индекса печени зафиксированы на равнинах полярного Зауралья и в Якутии (Шварц, Ищенко, 1971; Седалищев и др., 1981), где продолжительность зимовки особенно велика – до 9.5 мес.
Физиологические состояния, в которых пребывают зимующие под водой и промерзающие на суше лягушки, отличаются принципиально. Зимующие под водой амфибии реактивны на протяжении всего холодного периода, у них продолжается газообмен со средой. Соответствующие физиологические механизмы каждую осень перестраиваются на интенсивное выведение воды, поступающей в организм с постоянным осмотическим потоком. Эти животные, оказавшиеся в период осенней миграции к зимовальным водоемам в случайных ловушках (погреба, глубокие траншеи и т. п.), обычно гибнут в результате дегидратации в течение недели-двух (наши наблюдения). Существенные перестройки, происходящие в организме «сухопутных зимовщиков», к которым относится и R. arvalis, перед зимовкой имеют, по сути, противоположную направленность: значительно возрастает осмотическая концентрация плазмы крови (гипергликемия и уремия), в жидкостях организма увеличивается доля связанной воды, а свободная вода выводится в полости тела и замерзает. Наступает обратимое обезвоживание, организм существует в условиях жесткой гипоксии (Costanzo et al., 2013).
Таким образом, если подчеркивать видовую специфику, то зимующая травяная лягушка всего лишь снижает уровень метаболизма и настраивается на длительное пребывание в водной среде, тогда как остромордая переходит в состояние, близкое к анабиозу и исключающее длительный контакт с водой. В то же время известны популяции травяных лягушек, зимующих на суше, и популяции остромордых лягушек, особи которых зимуют под водой (см. ниже). Логично предположить, что во втором случае состояние гипобиоза поддерживается теми же механизмами, что и у «водных зимовщиков».
Доказанный и многократно подтвержденный эндогенный ритм жизнедеятельности представителей Anura умеренной зоны устанавливается у переживших первую зимовку особей после экспозиции годовым циклом местного климата и строго синхронизирован с ним (Wind-Larsen, Jørgensen, 1987). Последующая его «перенастройка» невозможна. Этот ритм включает в том числе и своевременный запуск всей суммы реакций, направленных на подготовку организма к зимовке. Так как физиологические и биохимические механизмы подготовки включаются заранее, то совершенно очевидно, что особи местных популяций не могут произвольно менять условия зимовки и то укрываться в воде, оставаясь реактивными, то пребывать на суше, запустив комплексный механизм адаптации к замораживанию.
Зимние стации травяной лягушки изучены весьма подробно (обзоры см.: Северцов и др., 1998; Кутенков, 2009). Это глубокие родники, проточные или имеющие родниковое питание озера и другие непромерзающие водоемы с достаточным содержанием кислорода. Наиболее удобны малые реки, в которых животные имеют возможность как выбора участков дна с подходящими условиями, так и их смены по ходу зимовки. В Онежском озере в Карелии травяные лягушки иногда попадались в рыбацкие снасти на глубине свыше 3 м (Коросов, Фомичев, 2005).
В западной части ареала R. temporaria, в зоне влияния атлантического климата, выбор убежищ зависит от местной гидрологической обстановки, и короткие по времени перезимовки на суше столь же успешны, как и в воде (Loman, 1980; Thiesmeier, 1992), а на побережье Бискайского залива к западу от Пиренеев годовой цикл не прерывается вообще (Bea et al., 1986). Однако в северной и восточной частях области распространения вида проведение зимы в подземных укрытиях весьма проблематично из-за частого глубокого промерзания почвы. И хотя отдельные примеры такого рода известны (Кузьмин, 1989 – г. Москва; Pasanen, Sorjonen, 1994 – юг Финляндии), успешная и постоянная зимовка на суше крупных группировок травяных лягушек или невозможна (Koskela, Pasanen, 1974), или ведет к их поголовной гибели в отдельные малоснежные и морозные зимы (Банников, 1940).
Сведений о зимних убежищах остромордой лягушки мало. Очевидно, это разного рода подземные полости или поверхностные (лесная подстилка, дернина, гниющая древесина и т. п.) укрытия. Лягушки обычно располагаются поблизости от репродуктивных водоемов, но могут оставаться и на расстояниях до километра от них. Так что сколько-нибудь выраженных весенних перемещений этого вида к местам икрометания может не быть, или, напротив, наблюдаются краткие и дружные миграции многих особей, зимовавших вдали от своих нерестилищ (Банников, Денисова, 1956; Ищенко, Леденцов, 1987; Juszczyk, 1987; Коросов, Фомичев, 2005; Elmberg, 2008; Остромордая…, 2014б; Glandt, 2014; В. Г. Ищенко, неопубл. данные).
В Европе водные зимовки остромордой лягушки являются исключением (Glandt, 2014). В редких случаях в ручьях и других небольших постоянных водоемах находили единичных взрослых R. arvalis, иногда в скоплениях зимующих R. temporaria (Красавцев, 1939; Juszczyk, 1987 и др.). Но, например, на востоке Скандинавского полуострова значительная часть населения остромордых лягушек на зиму уходит в озера, родники, канавы и другие водные стации, включая реки (Elmberg, 2008). В Якутии (зона вечной мерзлоты) вид зимует в основном на дне озер и рек (Белимов, Седалищев, 1979). В этих случаях амфибии зимой не оцепеневали, но сохраняли способность к перемещениям по дну, как это неоднократно отмечено у R. temporaria.
В отдельных частях зоны симпатрии рассматриваемых видов достаточно широко распространены болотные экосистемы. Поверхность, где чередуются сухие гряды с древесной и кустарниковой растительностью и заполненные водой мочажины (см., например, рис. 4 и 5), промерзающая на определенную глубину толща торфа и незамерзающие ключи, – эти биотопы объединяют черты и водоема, и суши. В таких районах оба вида могут использовать болота в качестве зимних убежищ. В Западной Украине в ямах проточных торфяных массивов находили скопления и травяных, и остромордых лягушек (Кушнирук, 1964). В Восточных Карпатах R. temporaria зимует в небольших долинных болотах, питаемых родниками (Demeter, Kelemen, 2011). В южной Карелии, несмотря на обилие постоянных водоемов, часть особей местных популяций травяной лягушки проводит зиму и в болотных стациях, забираясь в места, в которые поступают грунтовые воды или осуществляется подпитка из постоянных водоемов (Коросов, Фомичев, 2005; наши данные).
Таким образом, можно заключить, что различия и в физиологии гипобиоза рассматриваемых видов, и в характере их зимних убежищ принципиальны. Травяная лягушка, являясь видом с потенциально водной зимовкой, может оставаться в наземных укрытиях там, где это допускает местный климат. Остромордая лягушка, способная справляться с замерзанием организма, оказывается потенциально сухопутным зимовщиком, но в определенных ситуациях зимует и в воде.
Период размножения
Нерест
Наблюдения за самцами травяных лягушек, во множестве собирающимися на нерестилищах, неизменно дают возможность увидеть весьма оживленные и бурные события. Животные переплывают с места на место, расходятся по берегам, иногда дружно и большими группами переходят на соседние нерестилища; гораздо меньше самцов остаются на месте (Elmberg, 1986; Кутенков, 2009 и др.). Чем подвижнее самцы, тем они агрессивнее; подвижность и агрессивность прямо зависят от численности группировки (Elmberg, Lundberg, 1991). В разгар икрометания самцы, образуя своеобразную круговую зону активности вокруг скопления кладок икры, постоянно контактируют друг с другом, причем схваченная особь реагирует очень интенсивно и негативно. Подошедшие пары in amplexus привлекают внимание соседних особей, иногда образуются судорожные переплетения-клубки многих самцов вокруг полузадушенной самки.
На период нереста кожа самок травяной лягушки на спине и боках заметно краснеет. Показано, что примитивные модели (пластмассовые шарики), окрашенные в красный цвет, вызывают сильное возбуждение самцов: они обхватывают модель и друг друга, образуя цепочки, следуют за таким шариком на берег, в присутствии модели красного цвета их реакция схватывания вообще заметно активизируется (Кондрашов и др., 1976).
В классическом исследовании брачного поведения R. temporaria Р. Сэвидж описывает целую серию приемов борьбы самцов по удалению соперника и за правильную позицию спаривания, а также реакцию самки на правильный и неправильный ее захват (Savage, 1934). Однако успешная борьба за самку и выборочное спаривание осуществляются только при очень высокой плотности самцов, в такой ситуации выше и их общая агрессивность (Elmberg, 1991).
Весьма характерно и поведение самок: находясь в паре с самцом, они часто активно внедряются в уже образовавшуюся из кладок подушку, чтобы отложить там икру. Самкам R. arvalis подобное совершенно не свойственно. Поведение остромордых лягушек в разгар «тока» существенно отличается. Мы наблюдали, как на обводненном пространстве болота многочисленные «токующие» самцы R. arvalis подолгу оставались на одних и тех же местах порознь или, редко, по двое-трое вместе, перемещаясь лишь на расстояния до 10–50 см. Они почти не обращали внимания на соседей, попытки обхватить друг друга были вялыми и краткими. На проходящие мимо пары in amplexus и на пары, откладывающие по соседству икру, самцы реагировали спокойно. Животные очень пугливы.
В рассредоточенных скоплениях конкуренции среди самцов не наблюдается, перемещения их по нерестилищу заметно сокращаются с появлением самок (Аврамова и др., 1976). Борьба за самку происходит лишь изредка (Glandt, 2014). Обычные в разгар нереста судорожные клубки травяных лягушек у остромордых обнаруживали нечасто, они включали самку и двух-четырех самцов (Яковлев, 1979; Glandt, 2014; наши данные).
Самцы R. arvalis на время нереста приобретают специфическую окраску — от серебристо-голубой до синей, это единственный пример столь яркого проявления полового диморфизма у представителей рода Rana Палеарктики. Доказано, что голубая окраска самцов развивается, чтобы способствовать быстрому распознаванию особей своего пола (Sztatecsny et al., 2012). В определенных условиях (как правило, на мелководьях обширных и глубоких водоемов) остромордые лягушки иногда собираются большими скоплениями и выметывают сотни и тысячи кладок икры в одном месте (Панченко, 1980; Juszczyk, 1987; Sztatecsny et al., 2012 и др.). Но и в таких переуплотненных группировках никаких энергичных перемещений и серьезных попыток отбить самку обычно не наблюдают. То один, то другой самец совершает бросок к ближайшему соседу, касается его и сразу отплывает. Все скопление выглядит мельтешащим (Schweiger, 2014), но это не «свалка», как у травяных лягушек.
Крупные локальные репродуктивные группировки R. temporaria (сотни пар) редко откладывают всю икру в одном месте. Обычно формируются системы строго фиксированных и постоянных из года в год нерестилищ на ограниченном пространстве, когда самцы покидают те из них, где икрометание заканчивается, и скапливаются на новых местах (Кутенков, 2009). Напротив, многие самцы R. arvalis могут еще долго оставаться возле «своей» кладки, чутко реагируя на колебания воды (Аврамова и др., 1976; Гаранин, 1983). Авторы ошибочно считали такое поведение «охраной кладки»; на самом деле это реакция на вероятное появление очередной самки: икра лягушек несъедобна и в охране не нуждается.
Итогом столь разнящегося поведения двух видов являются и существенные различия в размещении кладок икры. У травяных лягушек почти всегда формируется «мат» из кладок, и чем больше животных собирается, тем он крупнее и плотнее (рис. 1). Иногда маты занимают до 10 м2 (Пикулик, 1985), а это более 1000 кладок. Рассеянное размещение кладок у остромордых лягушек не меняется с многократным увеличением количества нерестящихся в одном месте особей, всего лишь сокращается до минимума дистанция между ними. Даже в условиях относительного дефицита водного пространства, в случае соседства нескольких или многих икряных комков, они лишь касаются боками, не сливаясь в сплошную массу (Белимов, Седалищев, 1979; Пикулик, 1985; Северцов и др., 1998 и др.). И только в крайних ситуациях (например, бочаги на тростниковых сплавинах, ограниченные по площади мелководья и т. п.) многочисленные кладки остромордой лягушки образуют слитные массы (Яковлев, 1979; Фомичев, 2004; Loman, Andersson, 2007; В. Г. Ищенко, неопубл. данные; наши наблюдения).

Рис. 1. Репродуктивное скопление травяных лягушек на участке обводненного болота в заповеднике «Кивач». Здесь были отложены около 1000 кладок икры
Fig. 1. Breeding froggery of R. temporaria in the watered bog in the reserve «Kivach». Here, about 1000 spawn clumps were sheded
В случае совместного использования обширных мелководий рассеянные кладки R. arvalis могут находиться на плаву в местах глубиной до 50 см, тогда как «подушки» икры R. temporaria всегда будут касаться дна у берегов (Банников, Денисова, 1956; Пикулик, 1985; Коросов, Фомичев, 2007; наши наблюдения).
Репродуктивные водоемы в местах совместного обитания
Стации, используемые бурыми лягушками для икрометания, имеют принципиальное сходство. Это небольшие замкнутые или незамкнутые объемы стоячей воды, обычно с водной и полуводной растительностью. Приняв это во внимание, логично предположить, что в любой местности запас потенциальных репродуктивных стаций обоих видов будет одинаков. Существенными характеристиками мест размножения, помимо источников водного питания, свойств дна, берегов и т. п., оказываются положение на рельефе и ландшафтная приуроченность малых водоемов, используемых в ходе нереста.
В течение ряда лет мы исследовали размножение бурых лягушек в Карело-Кольском регионе. Его протяженность с юга на север составляет 1050 км между 60°40' и 69°57' с. ш. Характерно многообразие форм рельефа, обусловленное выходами кристаллических пород или близким их залеганием, деятельностью ледников и неравномерным покровом четвертичных отложений. Регион обладает избыточным увлажнением, густой гидросетью и включает морские побережья и крупнейшие озера Европы. Здесь, на широте полярного круга (66°30' с. ш.), по материку проходит северная граница зоны симпатрии исследуемых видов лягушек.
Проведена систематизация репродуктивных стаций по ландшафтному принципу, по приуроченности к тем или иным ландшафтным фациям (Кутенков и др., 1990; Кутенков, 2009). Упрощенная схема сводится к перечисленным ниже типам (в скобках – англоязычные названия, использованные при оформлении рисунков). Приведенные фотографии сделаны автором в заповеднике «Кивач».
Депрессии в скалах (Rock pools): ограниченные по площади непроточные и неглубокие естественные понижения по выходам кристаллических пород, расположенные в стороне от рек и озер (рис. 2).

Рис. 2. Обводненная скальная депрессия, площадь водного зеркала около 4500 м2, 38–363 кладки икры травяной и 0–40 – остромордой лягушек в разные годы
Fig. 2. Rock pool, water plane is about 4500 m2 . Preferable type of breeding ponds for R. temporaria. 38–363 spawn clumps of R. temporaria and 0–40 spawn clumps of R. arvalis in various years
Сырые леса (кóрбы, от финского korpi) (Forested swamps): слабопроточные протяженные ложбины или низины, заполненные торфом и занятые болотными лесами (рис. 3).

Рис. 3. Болотный лес (участок кóрбы) 0–9 кладок икры травяной лягушки в разные годы, остромордые лягушки в таких стациях не нерестятся
Fig. 3. Forested swamp (korpi) 0–9 spawn clumps of R. temporaria in various years; R. arvalis never spawns in such type of ponds
Облесенные болота (Forested mires): облесенные окрайки обширных болот (рис. 4), а также лужи и мочажины по небольшим массивам мезотрофных болотных комплексов с сосновым лесом.

Рис. 4. Облесенное болото, 124–953 кладки икры травяной и 0–29 – остромордой лягушек на контрольном участке в 20000 м2 в разные годы
Fig. 4. Forested mire, 124–953 spawn clumps of R. temporaria and 0–29 spawn clumps of R. arvalis in the reference area of 20000 m2 in various years
Открытые болота (Treeless mires): лужи и мочажины на болотных массивах с разреженным древостоем или на открытых болотах (рис. 5), а также водоемы по тростниковым сплавинам, торфяные мелководья остаточных озерков на болотах и т. п.

Рис. 5. Открытое болото Чечкино, 209–1745 кладок икры травяной и 60–1195 – остромордой лягушек на контрольном участке в 42000 м2 в разные годы
Fig. 5. Treeless mire Chechkino. Preferable type of breeding ponds for R. arvalis. 209–1745 spawn clumps of R. temporaria and 60–1195 spawn clumps of R. arvalis in the reference area of 42000 m2 in various years
Мелководья рек и озер (River and lake shallows): поросшие осокой мелководья и заливы крупных водоемов и постоянных ручьев (рис. 6).

Рис. 6. Мелководье р. Суна, 22–192 кладки икры травяной лягушки в разные годы; остромордая лягушка в этом месте не нерестится
Fig. 6. Shallow of the Suna river. The least important type of breeding ponds for both species. 22–192 spawn clumps of R. temporaria in various years; R. arvalis do not spawn in this place
Изолированные водоемы у берегов рек и озер (River and lake isolates): заполненные водой и не имеющие постоянной связи с материнскими водоемами понижения вблизи рек, озер (рис. 7) и крупных ручьев (старицы, отшнуровавшиеся заливы, затóки и пр.).

Рис. 7. Изолированный водоем у озера, 11–123 кладки икры травяной лягушки в разные годы; остромордая лягушка в этом месте не нерестится
Fig. 7. Isolated pond near the lake. 11–123 spawn clumps of R. temporaria in various years; R. arvalis do not spawn in this place
Антропогенные (Man-made ponds): заполненные водой придорожные кюветы, мелиоративные канавы, колеи транспорта по топким местам и другие искусственные углубления. Они невелики по размерам, могут быть расположены на любых формах рельефа, и представленность искусственных стаций размножения зависит от степени хозяйственной освоенности конкретного района. Изначально, как правило, располагаются на лишенных леса участках, но впоследствии по берегам часто вырастают заросли кустарника или молодой лес (рис. 8).

Рис. 8. Антропогенный водоем: канава заброшенной дренажной системы, устроенной на месте вырубленного болотного леса в 1950-е гг. В пределах всей дренажной системы на площади около 7500 м2 360–1714 кладок икры травяной и 8–75 – остромордой лягушек в разные годы
Fig. 8. Man-made pond: a ditch of the abandoned drainage system was built in 1950s in the site of the felled forest in a swamp. Within the entire drainage area of about 7500 m2, 360–1714 spawn clumps of R. temporaria and 8–75 spawn clumps of R. arvalis in various years
В строении поверхности заповедника «Кивач» (средние координаты 62°17' с. ш., 33°55' в. д.) сочетаются несколько генетических типов ледникового рельефа и развита гидросеть, включающая многочисленные постоянные и временные водоемы во всех ландшафтных фациях. По результатам обследования в 1986 г. мест икрометания лягушек в заповеднике на площади около 7000 га мы провели сравнение интенсивности использования водоемов разных типов. Всего были учтены 9410 кладок икры травяной и 2899 – остромордой лягушек. Результаты представлены на рис. 9. Оказалось, что травяная лягушка, нерестясь во всех вариантах водоемов, наиболее интенсивно использует заполненные водой скальные депрессии в лесу, облесенные окрайки болот и различные водоемы, возникшие в результате хозяйственной деятельности (на территории заповедника «Кивач» антропогенные водоемы составляют лишь 1 % всего запаса потенциальных репродуктивных стаций лягушек; см.: Кутенков, 1990). В сумме в этих угодьях в том году обнаружили 2/3 всех кладок икры R. temporaria. Остромордая лягушка нерестилась преимущественно по обводненным малооблесенным или открытым евтрофным участкам болот и тростниковым сплавинам по озерам.
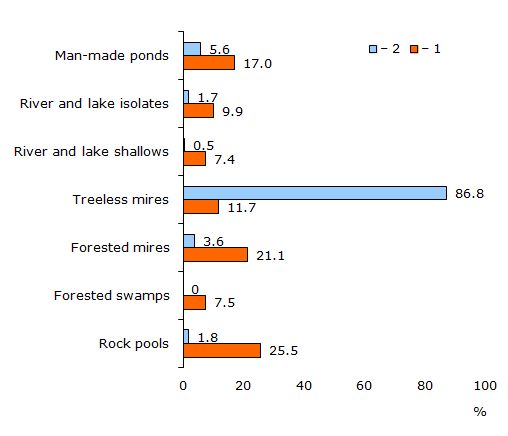
Рис. 9. Распределение кладок икры (%) травяной и остромордой лягушек по основным типам репродуктивных водоемов в заповеднике «Кивач» (62°17' с. ш., 33°55' в. д.) в 1986 г.: 1 – R. temporaria; 2 – R. arvalis
Fig. 9. Distribution (%) of brown frogs spawn clumps between different kinds of breeding ponds in «Kivach» State Reserve (62°17' N, 33°55' E) in 1986. 1 – R. temporaria; 2 – R. arvalis
На карельском о. Кижи в Онежском озере набор вариантов потенциальных репродуктивных водоемов бурых лягушек намного меньше, чем в «Киваче». Кроме обширных приозерных болот, занимающих 16 % площади острова и испытывающих крайне неравномерное затопление в разные годы, он включает также мочажины по берегам, озерки в термокарстовых воронках, лужи и канавы искусственного происхождения (Фомичев, 2004). Установлено, что в годы с низким (1998) и высоким (2001) весенним затоплением доля кладок травяной лягушки в пределах приозерных болот составляла соответственно 54 % и 49 %, а остромордой – 97 % и 98 % от общего числа учтенных на острове (рассчитано мною по данным: Коросов, 2010, с. 192).
Заповедник «Костомукшский» (средние координаты 64°30'–30°30') расположен на юго-восточном склоне водораздела Манселькя. Территория характеризуется выраженным мезорельефом, мощным чехлом водно-ледниковых отложений, обилием и разнообразием болот (преобладают «сухие» массивы мезотрофно-олиготрофного ряда). Мы обследовали около 4000 га территории, в том числе 12 км долины р. Каменной (левый приток р. Кеми). Здесь удалось обнаружить всего три разрозненных поселения R. arvalis. Места нереста (до 30 пар лягушек) были приурочены только к наклонным евтрофным грядово-мочажинным комплексам открытых болотных массивов. Ассортимент водоемов, используемых R. temporaria, оказался достаточно широким: мелководья рек и ручьев, лужи в их поймах, лужи при остаточных озерках на болотах, грядово-мочажинные болотные комплексы, обширные бессточные котловины меж высоких гряд и т. д. Большинство нерестилищ располагалось в долине р. Каменной и примыкающих урочищах. Здесь находились две трети мест икрометания и 70 % кладок икры от всего количества обнаруженных. По всей видимости, приходящие сюда на нерест травяные лягушки зимуют в реке. На удаленных от нее нерестилищах иногда встречались достаточно многочисленные группировки травяных лягушек – до нескольких десятков пар. Вместе с тем многие урочища, имеющие потенциальные репродуктивные стации, лягушек не привлекали.
На западном побережье Кандалакшской губы Белого моря на широте Северного полярного круга, где проходит граница ареала остромордой лягушки, мы в течение ряда лет (1986–1992) обследовали участок Кандалакшского заповедника, включающий о. Великий и п-ов Ковдский (Kutenkov, Panarin, 1995). Средние координаты района 66°34' с. ш., 33°09' в. д. Для него характерен крупногрядовый плавный рельеф с плащом рыхлых отложений. Здесь нет рек, сток проходит по сети ложбин. Равнинные участки невелики по площади и заняты преимущественно грядово-озерковыми аапа-болотами. Водоемы представлены средними и малыми озерами с заболоченными мелководьями. Имеется большое количество поросших осокой и тростником, заполненных торфом понижений с временными водоемами – бывшими заливчиками и лагунами, отделившимися от моря в результате неотектонического поднятия. На пологих берегах сформировались приморские луга с мелкими озерками и лужами. Нерестилища остромордой лягушки были приурочены к приморским лугам, к указанным заболоченным понижениям и, в меньшей мере, к сплавинам средних по величине озер. Все найденные места икрометания R. arvalis располагались по берегам или вблизи небольшого (около 40 км2) морского водоема, изолированного от Белого моря о. Великим и Ковдским п-овом. Набор репродуктивных стаций травяной лягушки включал дополнительно мелководья озер, озерки удаленных от моря аапа-болот и скальные депрессии в лесу, однако луж на приморских лугах вид избегал. В указанном для остромордой лягушки оазисе оказалась всего треть кладок икры травяной лягушки от общего числа обнаруженных. За пределами этого оазиса на открытом в Белое море берегу в точно таких же стациях остромордая лягушка отсутствовала, а травяная размножалась.
Опубликованные данные по некоторым другим частям зоны симпатрии таковы.
В северном Предуралье (Республика Коми) на равнине остромордая лягушка нерестится в лужах и озерах широких пойм, где пойм нет – в водоемах по болотам, а различные искусственные водоемы посещает гораздо реже. Травяная лягушка в этих условиях откладывает икру в те же водоемы в понижениях на пространстве пойм, очень часто нерестится в антропогенных водоемах, а озерки болот использует гораздо реже остромордой (Ануфриев, Бобрецов, 1996).
Низменность р. Онега, впадающей в Беле море, имеет холмистый рельеф. Он изобилует заболоченными западинами и озерными котловинами. Остромордая лягушка здесь нерестится в крупных мочажинах болот и по сплавинам зарастающих озер, травяная – в хорошо прогреваемых относительно глубоких водоемах преимущественно искусственного происхождения (Корнеева и др., 1984).
На Скандинавском полуострове в ландшафтах, где встречаются оба вида, R. arvalis в период нереста повсюду преобладает в болотных стациях, от зарастающих прудов в агроландшафте до тростниковых и сфагновых сплавин на озерах, а в дистрофных кислых мочажинах на олиготрофных болотах размножается только этот вид. R. temporaria нерестится в различных мелких водоемах, включая искусственные. На весенних разливах малых рек на луговые берега и полосы кустарника благоденствуют оба вида (Elmberg, 1978, 2008; Dolmen, 1986, 2008; Loman, Lardner, 2006).
В Московской области (р-н г. Звенигорода) исследован участок левобережья р. Москвы. На пространстве в 4 км от берега реки располагаются многочисленные (около 60) временные и постоянные водоемы в лесу и на открытых местах, включая старые торфяные карьеры на верховом болоте. Остромордые лягушки откладывали в карьерах тысячи комков икры, некоторая доля кладок оказывалась в ближайших лесных водоемах. Размещение травяных лягушек по репродуктивным стациям прямо противоположное: вид использовал прежде всего пруды и лесные водоемы надпойменной террасы и склона коренного берега реки. В торфяных карьерах число кладок травяной лягушки было на два порядка меньше, чем число кладок остромордой (Северцов и др., 1998; Ляпков, 2001).
В Белоруссии травяная лягушка нерестится в многочисленных лесных водоемах, во временных лужах, на разливах рек и озер, в старицах, часто – в искусственных водоемах. Остромордая лягушка использует почти тот же набор, обильна на разливах, но отличительной особенностью вида является икрометание на массивах верховых болот (Пикулик, 1985).
Для территории Нидерландов характерна относительная однородность равнинного ландшафта, поверхность сложена чередующимися песчаными и глинистыми отложениями разного генезиса и лёссом, а климат имеет черты океанического (География Нидерландов, 2014). Благодаря многообразию почвенных условий и обилию воды здесь сформировались различные болотные экосистемы, англоязычные названия которых не всегда имеют аналоги в русском языке: heathlands, raised bogs, fen meadows, polders, moorlands, peat moorlands, peat bogs, peatlands, marshes, marshes of the heathlands, etc. В олиготрофных «кислых» водоемах этих ландшафтных фаций R. arvalis преобладает или оказывается единственным размножающимся видом амфибий, однако в остальных случаях в постоянных и временных водоемах болот нерестятся оба вида. За пределами «болотной зоны» страны R. arvalis отсутствует, R. temporaria нерестится в искусственных водоемах (Strijbosch, 1979; Laan, Verboom, 1990; Vos, Chardon, 1998; Delft, Creemers, 2008).
В южной части Центральной Европы ландшафтные комплексы, населенные каждым видом, качественно различны и пространственно разделены (см. ниже). Поэтому случаи, когда в одной местности сосуществуют крупные популяции травяной и остромордой лягушек, весьма редки. Единственный хорошо документированный пример – это местность Csík Basin в румынских Восточных Карпатах. В разные годы популяции R. temporaria и R. arvalis насчитывали здесь до 2400–2800 половозрелых особей. Травяная лягушка распространена повсеместно, и в ассортимент ее репродуктивных водоемов входят горные ручьи, лужи-болотца в углублениях аллювиальных отложений, искусственные водоемы и мелководья небольшого водохранилища. Популяция остромордой лягушки занимает ограниченную территорию, и размножение ее наблюдали только в небольших болотцах по сырым лугам (Demeter, Kelemen, 2011).
Подытоживая, необходимо подчеркнуть принципиальные различия «стратегии и тактики» икрометания рассматриваемых видов.
Поведение травяных лягушек направлено на кучное икрометание, когда формируется икряной мат. Преимущества такого размещения кладок следующие. Во-первых, склеенные кладки мата, расположенного на мелководье ручья или речки, не сносит течением, а в глубоких старицах и озерках икра не погружается на дно. И чем ближе к центру плотного скопления, тем меньше вероятность смыва икры. Во-вторых, при снижении уровня водоема после невысокого паводка запасенная в оболочках множества икринок вода, высвобождаясь после вылупления личинок, дает им возможность нормально существовать на начальных стадиях развития. Единственной негативной стороной слипания многих кладок в «подушки» оказывается гипоксия: смертность эмбрионов достигает 30 % (Северцов и др., 1998), однако подобные «заморы» случаются редко (Пикулик, 1985).
Репродуктивное поведение остромордых лягушек явно демонстрирует тактику, обеспечивающую наибольший успех размножения в условиях обширных мелких водоемов. Это характерные для плоских форм рельефа заполненные талыми водами бессточные понижения, обводненные болота, разливы равнинных рек при половодьях и т. п. В стоячей воде потребность в фиксации кладок икры отсутствует. Диффузное же их размещение заметно увеличивает шансы на успешное завершение развития личинок в многочисленных лужах и «прудах», остающихся после обсыхания таких мелководий (см., например, Ищенко, 2008). При высокой плотности размножающихся R. arvalis образование скоплений кладок есть следствие простого сокращения до минимума дистанций между самцами, но не стремления животных к агрегации.
Специальные эксперименты убедительно продемонстрировали достаточно высокую генетически закрепленную устойчивость европейских популяций R. arvalis к кислотности репродуктивных водоемов (Andrén, Nilson, 1988; Hangartner et al., 2011). Она выше у эмбрионов и личинок из популяций, постоянно размножающихся в водоемах с низким рН (повышенной кислотностью), чем у потомства родителей, откладывающих икру в водоемы с нейтральной реакцией (значения рН соответственно 4.3 и 7.5). В аналогичных опытах на R. temporaria таких различий обнаружить не удалось: на потомство двух локальных группировок, откладывающих икру в водоемах с кислотностью 6.6 и 4.8, эффект низкого рН был одинаково негативен (Räsänen et al., 2002). Кислотность болотной воды обусловлена жизнедеятельностью сфагновых мхов и присутствием продуктов разложения органики.
Суммируя сказанное, учитывая поведенческие особенности и реальную ландшафтную приуроченность репродуктивных стаций, есть все основания полагать, что принципиальная схема, «стратегия» икрометания травяной лягушки сформировалась в условиях бассейнов небольших рек, не испытывающих высоких паводков, с сопутствующими ручьями-притоками, заливами, проточными озерками, старицами, рытвинами и пр. «Коренными» репродуктивными стациями остромордой лягушки, напротив, представляются плоские низинные пространства с непостоянным уровнем мелкой стоячей воды, часто закисленной. Следует, однако, подчеркнуть, что при совместном обитании видов бурых лягушек в благоприятных условиях спектры используемых репродуктивных стаций всегда в той или иной степени перекрываются.
Летние местообитания
Стации нагула – это летние прибежища, где амфибии растут, формируют половые продукты и запасы энергии на время зимовки и следующего за ней периода икрометания. Использование тех или иных летних местообитаний определяется их защитными свойствами, микроклиматом (температура и влажность приземного слоя воздуха) и влажностью субстрата: амфибии чрезвычайно чувствительны к физическому качеству местообитаний. Состав и структура растительного покрова конкретного ценоза, угол склона, экспозиция и т. д. имеют подчиненное значение. Отличия в использовании летних местообитаний близкими видами лягушек отражают различные их требования к качеству, к «специфическим ресурсам» нагульных стаций (Rittenhouse, Semlitsch, 2007). В следующем обзоре использованы прежде всего те источники, в которых содержатся сведения по биотопической приуроченности травяных и остромордых лягушек в районах, где они обитают совместно. В таких условиях предпочтения того или другого вида проявляются наиболее четко.
Фенноскандия
В лесном заповеднике «Кивач» R. temporaria встречается по всей территории, в минимальном количестве отмечена на болотах и в «сухих» сосняках. Встречи R. arvalis в лесах единичны, население вида сосредоточено в основном на болотах, включая сплавины на озерах (Ивантер, 1969; Кутенков, 1984). На протяжении ряда лет мы исследовали размещение особей двух видов на площадке, охватывающей участки влажного разнотравного луга и окружающего хвойно-лиственного леса, размеры площадки – 0.6 га (Кутенков, 2009). Всего зафиксированы точки 1042 встреч травяной и 114 – остромордой лягушек, из них 268 встреч травяной и всего 2 встречи остромордой лягушек пришлись на лесные части площадки. В процентном выражении это составляет соответственно 26 % и 1.8 %.
Сходное соотношение в численном распределении травяных и остромордых лягушек в соседствующих лесных и луговых стациях получено при исследовании в южной Швеции: в лесу отловили 25 % и 2 % особей этих видов. При этом влажные луговые ассоциации были заселены особями обоих видов с одинаковой плотностью (Loman, 1978, 1984).
На островах Кижского архипелага в Онежском озере ассортимент пригодных летних стаций бурых лягушек ограничен. Травяная встречалась повсюду, в том числе на обсыхающих летом болотах, а нагул остромордых лягушек проходил почти исключительно в обширных приозерных тростниковых болотах и ближайших окрестностях (Коросов, Фомичев, 2005).
На пределе распространения R. arvalis в регионе (побережье Белого моря) группировки вида приурочены к неширокой (до 1.5 км) полосе берега заливов или небольших изолированных морских водоемов. Амфибии держатся здесь по неглубоким прибрежным болотам, на сплавинах и приморских лугах, почти не встречаясь в глубине суши. R. temporaria обитает по берегам речек и ручьев, в заболоченных и поросших кустарником ложбинах временных водотоков (Kutenkov, Panarin, 1995; Кутенков, 2009).
В лесотундре северной Финляндии остромордая лягушка обитает только на болотах, тогда как травяная – повсеместно (Terhivuo, 1981). В южной Финляндии, при очевидной синтопии, R. arvalis более оседла и отдает предпочтение сырым околоводным местообитаниям (Haapanen, 1970).
На лесистых территориях центральной и северной Швеции травяная лягушка обнаружена во многих стациях, остромордая держится в основном на массивах сфагновых болот, в луговых ассоциациях, в лиственных и сырых хвойных лесах. В отдельных болотистых местообитаниях R. arvalis доминирует или оказывается единственным видом лягушек (Gislén, Kauri, 1959; Elmberg, 1995, 2008).
За пределами Фенноскандии, этого наиболее заболоченного региона Европы (Montanarella et al., 2006), прослеживаются сходные закономерности биотопической приуроченности данных видов в период нагула.
Северо-восток зоны симпатрии
В равнинной Большеземельской тундре остромордая лягушка летом держится по берегам озер, в долинах рек и обсыхающих летом руслах малых водотоков (Лобанов, 1977; С. А. Кутенков, устн. сообщение). На границе тундры и лесотундры за полярным кругом (Республика Коми) травяная лягушка была единственным видом в мохово-лишайниковых сообществах мелких кустарников в тундре, доминировала в островных лесах, а остромордая – по низменным, сильно заболоченным берегам р. Уса (Леонтьева, 1989). В северной и средней тайге Предуралья травяная лягушка заселяет весь спектр лесных местообитаний, предпочитает наиболее увлажненные их типы, а численность на болотах и в заливаемых поймах невысока. Напротив, «там, где поймы рек выражены слабо, а обширные болота отсутствуют, остромордую лягушку практически невозможно встретить» (Ануфриев, Бобрецов, 1996, с. 63). В восточных предгорьях Северного Урала в разнообразных северотаежных лесных и кустарниковых местообитаниях в долинах рек травяная лягушка распространена шире остромордой и в трети обследованных биотопов оказалась монодоминантом (Стариков, Матковский, 2011).
Средняя полоса России
В Предуралье и на севере Волжско-Камского края остромордая лягушка населяет сырые луга, болота, лесные поляны и редины, травяная – хвойные леса, поймы рек, выходы грунтовых вод как в лесах, так и в открытых ландшафтах (Гаранин, 1983).
В южнотаежных и смешанных лесах средней полосы (Калининская, Костромская и Ивановская обл.) травяная лягушка доминирует в лесных, остромордая – в пойменных и болотных стациях (Шапошников и др., 1959; Преображенская, Байкалова, 1984; Okulova, 1995 и др.).
Белоруссия и восточная Польша
В этом регионе соседствуют смешанные (72 %) и широколиственные леса и чередуются различные ландшафтные комплексы: от невысокой (до 345 м) Белорусской гряды до почти сплошь заболоченной Полесской низменности. Оба вида лягушек оказываются эвритопными. Остромордая обильна в лиственных лесах, в пойменных лугах и ольшаниках, она всюду и абсолютно доминирует на верховых болотах. Травяная лягушка, встречаясь в различных биотопах, в значительно большей степени приурочена к лесам в наиболее влажных их вариантах, к поросшим кустарником пойменным лугам, а в заболоченных местностях отсутствует (Пикулик, 1985; Рыжевич, 2001; Jędrzejewska et al., 2003).
Широколиственные и смешанные леса
На севере Среднерусской возвышенности во вторичных мелколиственных лесах с лугами и полянами отмечено пятикратное преобладание травяной лягушки над остромордой по численности (Леонтьева, Перешкольник, 1982).
В начале прошлого века в окрестностях Киева травяная лягушка населяла леса, рощи, кустарниковые заросли, а на лугах в пойме Днепра встречалась редко. Остромордая, напротив, была многочисленна на лугах и гораздо реже встречалась в лесных стациях (Шарлеман, 1917).
На холмистых равнинах юга Польши травяная лягушка встречается во многих местообитаниях, в том числе в лиственных и хвойных лесах, по берегам прудов, реже на лугах. Остромордая предпочитает открытые, заросшие кустарником пространства, лесные поляны, влажные торфяные осоковые луга, отмечена в лесных стациях. При этом вид редко заселял очень сырые заболоченные территории и не встречался у прудов и мелиоративных канав с водой (Juszczyk, 1987).
В широкой долине среднего течения Рейна (около 50° с. ш.) R. temporaria заселяет как лесные, так и открытые стации, тогда как R. arvalis – только сырые луга, заболоченные леса и поросшие тростником берега (Heimer, 1981).
В Венгерской низменности R. temporaria встречается в лиственных лесах в долинах и на склонах. R. arvalis населяет сырые луга, обильна и в лесных стациях, расположенных вокруг болот, стариц и в поймах рек (Puky et al., 2005).
На Трансильванском плато (Румыния) в бассейне р. Тырнава Маре располагается холмистая местность с узкими (100–150 м) долинами водотоков. В таких условиях болотные массивы сформироваться не могли. В результате R. arvalis, обильная в болотных местностях других частей страны, здесь очень редка, а R. temporaria – обычный вид лесных стаций (Hartel et al., 2014).
Побережье Северного моря
Пологохолмистые и равнинные ландшафты Нидерландов практически лишены леса: его остатки покрывают лишь около 8 % территории страны (География Нидерландов, 2014). В условиях избыточного увлажнения и повсеместного хозяйственного освоения летние местообитания бурых лягушек весьма сходны. Это луга, пастбища, кочковатые, с небольшими водоемами и кустарником верещатники и т. п. Расхождение видов в пространстве минимально: R. temporaria отмечена в лесополосах и других ландшафтных элементах с лесной растительностью, а R. arvalis доминирует на сырых пустошах (moorland), на верховых и низинных болотах (Laan, Verboom, 1990; Vos, Chardon, 1998; Vos et al., 2007; Delft, Creemers, 2008). Примечательно, что в этой части зоны симпатрии оба вида не заселяют как низменное морское побережье, где сосредоточены все озера страны, так и долины крупнейших рек Западной Европы (Рейн, Маас).
Зона лесостепи европейской России
В этой зоне в пределах Среднерусской возвышенности остромордая лягушка обычна в приречных лесах, по днищам логов. Травяная очень редка и встречалась в заливных ольшаниках и островных лесах по террасам, в оврагах, балках и на береговых склонах с выходами грунтовых вод (Птушенко, 1934; Елисеева, 1967; Лада, 1993; Кочетков, Шубина, 2011).
В северной части Приволжской возвышенности остромордая лягушка также населяет различные влажные местообитания, но предпочтение отдает поймам рек, увлажненным смешанным лесам, влажным балкам, заболоченным лугам, болотам. Травяная, как и на Среднерусской возвышенности, приурочена лишь к сырым местообитаниям с лесом или кустарником вдоль ручьев, речек, прудов. Повсюду здесь она тяготеет к местам выхода грунтовых вод в отрицательных формах рельефа (балки, овраги), которыми богата местность. В отдельных биотопах летом встречались оба вида (Барабаш, 1939; Гаранин, 1983; Лягушка травяная…, 2005; Шляхтин и др., 2005; Ручин, 2015 и др.).
Следует добавить, что в полосе возвышенностей Восточно-Европейской равнины (примерно от 50° с. ш., 7° в. д. до 56° с. ш., 44° в. д.) отмечено заметное участие внепойменных лесных ландшафтных комплексов, включая сухие сосняки, в составе летних местообитаний R. arvalis (Красавцев, 1939; Иноземцев, 1978; Juszczyk, 1987; Рыжевич, 2001; Glandt, 2014 и др.). Обычны встречи этого вида в лесных стациях на террасах и плакорах Среднерусской и Приволжской возвышенностей (Барабаш, 1939; Глазов, 1975; Леонтьева, Перешкольник, 1982; Рыжов, Ручин, 2007 и др.). Кроме того, остромордая лягушка встречена в смешанных, лиственничных и сосновых лесах восточных предгорий Южного Урала (Куранова, Каштанова, 2001). Это несколько искажает создающееся представление о повсеместном предпочтении видом «болотных» местообитаний. К сожалению, в цитированных работах авторы часто не перечисляют всего набора жилых стаций вида в исследованных районах. Возможно, ситуацию проясняет следующая цитата: «В меньшем числе она (остромордая лягушка на севере Приволжской возвышенности. — А. К.) встречается в лесах, расположенных на террасе. Обычно такие места (сосняки, березняки, дубравы) характеризуются пониженной влажностью, что предопределяет низкую численность остромордой лягушки в них. В то же время количество учитываемых лягушек увеличивается при наличии в подобных местах водоема, ручья, влажного оврага и т. п.» (Рыжов, Ручин, 2007, с. 35). Такие болотистые участки встречаются в заселенных этим видом сухих сосняках на песчаных почвах Северо-Германской низменности (Glandt, 2014).
За пределами зоны симпатрии в восточной части ареала R. arvalis демонстрирует те же биотопические предпочтения, что и в Европе.
Западно-Сибирская равнина
Повсюду от южных тундр до лесостепи на юге остромордая лягушка достигает максимального обилия в различных вариантах болотных экосистем в поймах и на надпойменных террасах. Это низинные, переходные и комплексные болота, займища, луга, реже приречные смешанные и березовые леса. В сходных условиях на обилии R. arvalis отрицательно сказывается степень удаленности от пойм крупных рек, а также увеличение залесенности – доли хвойных пород (Шварц, Ищенко, 1971; Равкин, 1976; Блинова, 1984; Вартапетов, Ануфриев, 1984; Куранова, 2001; Лягушка остромордая…, 2015 и мн. др.).
Алтае-Саянская горная область
В характерных обширных котловинах среднегорья (Турано-Уюкская, Тоджинская) R. arvalis обнаружена по заболоченным участкам долин с текущими реками (Коротков, Короткова, 1976; Заповедник Азас, 2014). К востоку, на Лено-Ангарском плато, обитает на заболоченных лугах, на сплавинах по озерам и прудам (Малеев, 2009).
На западе Алтайской части этой горной страны в условиях достаточного увлажнения заметную роль играют высокотравные луга (Катунский хребет), а при избыточном увлажнении (более 2000 мм осадков в год) получают широкое распространение и разнообразные болотные комплексы (Кузнецкий Алатау). В субальпийском поясе остромордая лягушка встречена только на указанных лугах и болотах. На Катунском хребте максимальное обилие этого вида зарегистрировано в березовых горно-долинных лесах. В Кузнецком Алатау, помимо болот, он обитает во влажных смешанных лесах и на пойменных лугах, практически не встречаясь в хвойных лесах. В крупнейшей на Алтае долине р. Чулышман, протянувшейся от засушливых степей до высокогорий с болотистыми кустарниковыми тундрами, изобилуют реки и мелкие и крупные озера, разбросанные по плоским поверхностям нагорий. R. arvalis обитает здесь в высокотравных лесах, на увлажненных лугах приозерных террас и пойм, в разреженных лиственничниках с элементами тундры (Брысова и др., 1961; Яковлев, 1977; Заповедник «Кузнецкий…, 1999, 2014; Вознийчук, Куранова, 2008; Эпова и др., 2013; Катунский…, 2015).
Локомоторные способности остромордой и травяной лягушек существенно различаются. Чтобы убедиться в этом, достаточно даже поверхностного взгляда на их внешний облик (Приложение I). Каплевидная форма тела R. arvalis, некрупные размеры (взрослые особи как минимум на треть короче, чем R. temporaria) и очевидная прыть (частые короткие прыжки, резкие смены направления движения) позволяют этой юркой лягушке мгновенно скрываться в мелких мочажинах, затаиваться в торфе или переплетениях травы. Напротив, имея массивное тело (взрослые самцы втрое тяжелее) и сильные задние конечности, травяная лягушка обладает мощным аллюром, способна проходить длинные дистанции. Иными словами, остромордая лягушка представляется существом достаточно «уютно-оседлым», тогда как в случае с травяной мы имеем дело с весьма мобильным животным.
Действительно, протяженные энергичные миграции травяных лягушек на нерестилища, затем к летним местообитаниям, достигающие порой 10 км, в определенной степени упорядоченные широкие перемещения по пространствам нагульных стаций, а также возвратные походы к местам зимовок хорошо известны (Кутенков, 2009). В то же время сколько-нибудь массовые перемещения, совершаемые взрослыми особями местных популяций, не описаны ни для каких периодов годичного цикла R. arvalis. Известна ситуация, когда эти лягушки держались на истоптанном коровами сыром лугу по одну сторону неширокого канала, но не появлялись на другом его берегу, не испытывавшем пастбищной нагрузки (Delft, Creemens, 2008). Лишь в районах, где условия позволяют или вынуждают R. arvalis разбредаться по различным местообитаниям, лягушки к осени могут собираться возле своих репродуктивных стаций, в которых (или поблизости) и проходит зимовка, а весной совершать краткие миграции к нерестилищам (Ищенко, Леденцов, 1987; Glandt, 2014; В. Г. Ищенко, неопубл. данные). Установлено, что в благоприятной обстановке перемещения взрослых особей «оседлых» видов лягушек обычно не превышают радиуса в 700 м от мест репродукции (Rittenhouse, Semlitsch, 2007).
С мобильностью травяной лягушки связана и широта спектра занимаемых ею летних местообитаний в северной части зоны симпатрии. Обычно это разнообразные лесные стации, заросли кустарника и близлежащие луговые участки. Эта лягушка способна преодолевать малопригодные территории на пути к знакомым местам нагула (Vos et al., 2007). В периферийных частях ареала за пределами лесного биома она вынужденно придерживается тех же «закрытых» стаций. Летние местообитания R. arvalis – это прежде всего открытые или полуоткрытые ландшафтные фации, часто заболоченные. Появление вида в «сухих» лесных биотопах связано, по-видимому, с находящимися поблизости болотистыми участками местности.
Обитание в горах
Скандинавские горы
В пределах этого гористого полуострова R. arvalis не идет выше 350 м над уровнем моря, единичные находки зафиксированы на 500 м. R. temporaria распространена всюду (икрометание отмечено на высоте свыше 1000 м), а до высоты 800 м н. у. м. является обычным видом (Dolmen, 1986, 2008; Elmberg, 1995, 2008).
Среднегорья и низкогорья Центральной Европы представлены полосой, протянувшейся между приатлантическими низменностями (Нидерланды, Северо-Германская и Польская низменности) и горными массивами Альп и Западных Карпат.
В Бескидах травяная лягушка обнаружена на максимальных высотах – до 2000 м н. у. м., тогда как остромордая – не выше 650 м н. у. м. (Berger, 1975).
В одном из давних исследований (Opatrný, 1978) приведены сведения о распространении земноводных в ряде районов бывшей Чехословакии. R. arvalis встречалась лишь до высоты 510 м, отсутствуя на невысоких Судетах и Чешско-Моравской возвышенности. При этом только средняя высота находок R. temporaria по указанным автором 80 пунктам составляет 520 м н. у. м. (рассчитано мною).
В среднем течении р. Рейн (массив Оденвальд) остромордая лягушка не поднимается из долин выше чем на 180 м н. у. м., тогда как травяная распространена повсеместно до предельных здесь высот около 500 м (Heimer, 1981).
В Альпах травяная лягушка оказывается единственным из рассматриваемой пары видов. В долинах северо-западных Альп она существует на высотах более 2000 м (Miaud et al., 1995), а в итальянских Альпах жизнеспособная популяция вида обнаружена почти на 2750 м н. у. м. (Vences et al., 2003).
Восточные Карпаты
Травяная лягушка населяет весь горный массив Карпат. Интерес представляют верхние пределы распространения здесь лягушки остромордой.
Наивысшие точки достоверных находок R. arvalis на западном макросклоне Восточных Карпат составляют 742 (центр) и 844 (юг) м н. у. м. В долине верховьев р. Олт, левого притока Дуная, обитают несколько популяций вида. Средняя высота по 18 пунктам встреч, включая указанные выше 844 м, составила 620 м н. у. м. (рассчитано мною по: Sas et al., 2008).
В наиболее высокой части украинских Карпат (Черногора) травяная лягушка встречается повсеместно до 2000 м н. у. м., тогда как остромордая обнаружена лишь однажды на высоте, по-видимому, около 900 м (Гуль, 2001). Есть сведения о нахождении R. arvalis в этом регионе на 987 м (Ishchenko, 1997).
В горах Балканского полуострова (массивы Стара Планина, Пирин и Родопы), где остромордая лягушка отсутствует, травяная оказывается обычным видом, но населяет только долины на высотах 1200–2400 м (Petrov, 2007).
В Венгерской низменности (Среднедунайская равнина), лежащей к востоку от Альп и к югу от Западных Карпат, почти все популяции R. arvalis обитают на высотах 100–200 м н. у. м. на равнине или на невысоких холмах. Напротив, крупные группировки R. temporaria обнаружены только начиная с высот более 600 м (Puky et al., 2005; Puky, Shad, 2008).
Уральский хребет является восточным рубежом зоны симпатрии рассматриваемых видов. На севере этой горной страны остромордая лягушка встречена только на равнине, уже в предгорьях (200–300 м) ее нет. Травяная обычна и в предгорьях, и в горах (Топоркова, Зубарева, 1965; Шварц, Ищенко, 1971; Ануфриев, Бобрецов, 1996). На Среднем Урале (западный макросклон, хребет Басеги) R. arvalis многочисленна на равнине, а по заболоченным долинам поднимается на высоты от 200 м и выше, доминируя над R. temporaria. На хребтах, наоборот, преобладает или оказывается единственным видом травяная лягушка (Топоркова, Варфоломеев, 1984). В восточных предгорьях Южного Урала травяная лягушка уже редка, а остромордая обнаружена всюду, кроме гольцов и степи, поднимаясь, видимо, до 500 м н. у. м. (Куранова, Каштанова, 2001).
Мы видим, что в горах Европы остромордая лягушка, по сравнению с вездесущей травяной, фактически избегает местностей, расположенных выше 500–600 м н. у. м. Предел для нее составляет здесь, по-видимому, около 900 м н. у. м. (обзор см.: Glandt, 2014), тогда как R. temporaria идет на втрое большие высоты. Ситуация меняется, когда мы обращаем внимание на гористую юго-восточную область ареала R. arvalis.
В горах Прибайкалья и Забайкалья остромордая лягушка населяет долины и нижние части хребтов, поймы, низовья и дельты рек. В глубине тайги и на высокогорных хребтах ее нет (Швецов, 1977; Байкало-Ленский…, 2014; Витимский…, 2014; Остромордая лягушка..., 2014). В Саянах в двух обширных котловинах: Турано-Уюкской (высота 700–1100 м н. у. м.) и Тоджинской (800–1800 м н. у. м.) вид оказался обычным (Коротков, Короткова, 1976; Заповедник Азас, 2014). В Кузнецком Алатау популяции R. arvalis найдены на высотах до 1600 м (Эпова и др., 2013). Наконец, в горах Алтая вид поднимается до 2140 м н. у. м. (Яковлев, 1980) и даже до 2400 м (Вознийчук, Куранова, 2008). Это близко к зафиксированному максимуму высот для R. temporaria в Европе (Альпы).
Основным фактором, лимитирующим высотное распространение лягушек в горных областях, является продолжительность безморозного периода. Так, в Пиренеях не обнаружено устойчивых популяций R. temporaria на высотах свыше 2500 м н. у. м., где в конце июля репродуктивные водоемы были еще покрыты льдом, а земля – снегом. Однако в Альпах долинные водоемы с тысячами головастиков этого вида были найдены на высоте почти 2750 м (Vences et al., 2003). Авторы полагают, что горные хребты Пиренеев недостаточно высоки, чтобы обеспечивать в долинах благоприятный температурный режим. Те же проблемы и у остромордой лягушки. На Алтае в высокогорном редколесье с элементами тундры на высотах от 1860 до 2140 м (самая высотная из изученных популяций вида) икрометание R. arvalis может начинаться не раньше конца мая – середины июня, когда озера еще покрыты льдом, а уже в конце июля там начинал идти снег (Яковлев, 1980, 1981).
Причины столь резких отличий в высотном распределении двух видов в зоне симпатрии и в то же время наличия свидетельств обитания R. arvalis на значительных высотах в горах Сибири заключаются в принципиальном различии свойств рельефа рассматриваемых горных областей Европы и Азии. Сведения о развитии гор, их рельефе и ландшафтах, использованные при написании следующего далее раздела, взяты из монографии Н. А. Гвоздецкого и Ю. Н. Голубчикова «Горы» (1987).
Все европейские горы (Пиренеи, Альпы, Карпаты, Скандинавские и горы Балканского п-ова), включая Урал, имеют преимущественно складчатую структуру, которая возникает в процессе деформации (складки и надвиги) земной коры при ее горизонтальных подвижках. Гораздо меньшую роль играют глыбовые и сводовые поднятия, которые существенно выражены только в рельефе среднегорий и низкогорий Центральной Европы (Рейнские Сланцевые горы, Шварцвальд, Чешский массив и др.), протянувшихся к югу от Среднеевропейской равнины. Эти горные системы линейновытянутые, расчленены слабо разработанными речными долинами, узкими и крутостенными. Среднегорья и низкогорья имеют характер рассеченных эрозией плоскогорий. В известняковых толщах Альп, Карпат и Балканских гор широко распространены карстовые явления (недалеко от своего истока под землей исчезает Дунай).
В европейских горах нет обширных котловин, ширина долин повсюду не превышает первых километров, а их днища почти не бывают плоскими. Тонкие частицы продуктов выветривания смываются со склонов, выносятся водными потоками и не скапливаются в отрицательных формах рельефа. Из-за сильно расчлененного рельефа и водопроницаемости рыхлых отложений (а то и самих кристаллических пород) болотные образования редки и занимают лишь первые проценты площади, а крупных массивов не существует вовсе (Кац, 1971). Потоки горных рек и ручьев не формируют пойм, долинные леса и луга произрастают на дренированных почвах. Озера имеют ледниковое происхождение, а их берега подвержены заболачиванию в минимальной степени.
Поселения травяной лягушки в горах приурочены к лесистым долинам с текущими по ним речками и ручьями, к берегам озер во всех поясах (Никитенко, 1959; Pascual, Montori, 1981; Heráň, 1982; Serra-Gobo et al., 1998; Vences et al., 1999; Куранова, Каштанова, 2001; Sztatecsny, Hödl, 2009 и мн. др.). В таких условиях остромордая лягушка лишена своего «коренного» ландшафта – плоских заболоченных пространств со стоячей водой. Есть лишь одно известное автору хорошо документированное исключение из этого правила. В румынских Восточных Карпатах на высоте 630–750 м н. у. м. расположена вытянутая долина, не типичная для Альпийского складчатого пояса. Этот водосборный бассейн (Csík Basin) в верховьях р. Олт, шириной около 10 км, имеет пологие склоны, плоское дно и характеризуется выраженной поймой, наносами, надпойменными террасами, ручьями и родниками. В этих условиях сформировались различные болотные экосистемы, сырые луга и временные стоячие водоемы (Demeter et al., 2011). Популяция остромордой лягушки занимала здесь ограниченную территорию площадью 4 км2, насчитывая при этом до 2800 половозрелых особей (Demeter, Kelemen, 2011).
В отличие от складчатых горных цепей Европы, в Сибири существуют объединенные в один пояс горные страны (Алтай, Саяны, Забайкалье, Алдан). При формировании этого пояса, протянувшегося почти на 3500 км, ведущее значение имели сводовые и сводово-блоковые поднятия. Обширные участки земной коры испытали воздымание с последующим разбиением на блоки. При этом вознесенные реликтовые поверхности выравнивания образуют участки всхолмленных равнин на плато, нагорьях и плоскогорьях, формируют днища межгорных котловин, выполненных продуктами выветривания. Многочисленные долины и котловины пояса гор Южной Сибири порой столь обширны (десятки километров в ширину и первые сотни в длину), что внутри них выделяют самостоятельные нагорья и хребты.
Такие возвышенные всхолмленные равнины и плоскогорья в силу климатических условий и воднофизических свойств рыхлых отложений обильно обводненные, благоприятны для широкого распространения болотных массивов. Ровные днища речных долин имеют заболоченные поймы, по плоским поверхностям разбросаны многочисленные мелкие и крупные озера. В высокогорьях (Алтай) в пологих долинах и по сглаженным плоскогорьям лежат сырые моховые редколесья и болотисто-кустарниковые тундры, занимающие обширные пространства (Брысова и др., 1961; Кац, 1971).
Жизнеспособные популяции остромордой лягушки населяют все высотные пояса перечисленных горных районов Сибири там, где существуют обширные выположенные поверхности: от заболоченных пойм рек (Витим) до высокогорных тундр (Алтай).
Ландшафтная приуроченность в ареалах и природные особенности их границ
На пространствах зоны симпатрии R. temporaria и R. arvalis обнаруживаются соседствующие обширные области природных ландшафтов, где один из пары видов успешно существует, а второй практически отсутствует. Остромордая лягушка абсолютно доминирует или оказывается единственным видом в монотонных равнинных болотистых местностях и подверженных половодьям речных бассейнах, а травяная – там, где местность становится возвышенной или гористой и пересеченной. Примеры тому мы находим на северо-востоке Европы (Топоркова, Зубарева, 1965; Лобанов, 1977), в Карело-Кольском регионе (Кутенков, 2009), на Скандинавском п-ове (Gislén, Kauri, 1959; Dolmen, 1986; Elmberg, 1995, 2008), на правобережье Среднего Поволжья (Ивановская обл.) (Okulova, 1995), в Белоруссии (Пикулик, 1985; Рыжевич, 2001).
Имеется хорошо документированный пример смены крупной популяции травяной лягушки популяцией лягушки остромордой в результате коренного преобразования среды обитания этих земноводных. Оно связано с устройством в 1941–1947 гг. Рыбинского водохранилища на р. Волге.
Междуречье левых притоков Волги – рек Мологи и Шексны – представляло собой незаливаемую древнюю озерную террасу, рассеченную речными долинами. На слабо расчлененной террасе росли хвойные леса, большие пространства занимали верховые болота. Ландшафт долин был разнообразен и включал многочисленные русла речек и ручьев, гривы, мелкие озера, низинные болота и заливные луга, участки смешанного и широколиственного леса. В сырых равнинных местообитаниях террасы была обычна остромордая лягушка, а в разнообразных стациях пойм травяная оставалась единственным из рассматриваемой пары видов. После заполнения чаши Рыбинского водохранилища большие площади Молого-Шекснинской низины, ставшие побережьем, были подтоплены. Берега начали испытывать значительные (до 5 м) и непостоянные сезонные колебания уровня воды. Со временем подтопленные территории превратились в болота, а пойменные луга, смешанные леса и травяно-кустарничковые заросли погибли. Из этих болотных пространств R. temporaria практически исчезла, уступив место значительно увеличившей численность R. arvalis, тогда как в сохранившихся по периферии исходных пойменных местообитаниях соотношение видов осталось прямо противоположным (Калецкая, 1953; Дарвинский…, 2014).
Наиболее яркие свидетельства пространственной дивергенции рассматриваемых видов, а именно разобщение на уровне соседствующих крупных природных территориальных комплексов, мы находим в периферийных областях зоны симпатрии.
На севере, в пределах Восточной Фенноскандии, граница распространения остромордой лягушки образует дугу вдоль южного подножия невысоких гор юга Мурманской области (восточная половина Кольского п-ова и, в частности, обширная заболоченная низменность в ее южной части остаются пока неисследованными). Далее граница оконтуривает по равнине северную часть меридионального водораздельного хребта Манселькя и, наконец, уходит на юг-юго-запад по восточным отрогам Скандинавских гор. Травяная же лягушка имеет здесь сплошное распространение до пределов материка, населяя как самые высокие горы северных частей Финляндии, Швеции, Норвегии и Мурманской области, так и скалистую тундру Кольского п-ова (Terhivuo, 1981; Elmberg, 1995; Kutenkov, Panarin, 1995; Кутенков, Коросов, 2001).
Крайняя западная часть ареала остромордой лягушки занимает субширотный выступ Восточно-Европейской равнины длиной около 500 км и шириной до 150 км (Glandt, 2014). Этот «угол» ареала с юга ограничен пологими волнистыми, сильно эродированными, подступами к плато Северогерманских гор и массиву Арденны (Бельгия – Горная..., 2015). Рельеф, а также механический и химический состав материнских пород не допускают развития на этих склонах болотных экосистем, а гидросеть представлена лишь небольшими реками и прудами. В таких условиях остромордая лягушка исчезает из состава фауны земноводных (Laan, Verboom, 1990; Delft, Creemers, 2008). Добавлю, что вся западная часть ареала R. arvalis, целиком занимающая Среднеевропейскую равнину (Польша, север Германии, Дания и Нидерланды), практически в точности повторяет контур сплошного распространения болотных (торфяных) почв в этой части Европы (Montanarella et al., 2006). Очевидно, что именно отсутствие заболоченных пространств со стоячей водой не пускает остромордую лягушку в Северо-Французскую низменность. Следует подчеркнуть, что в голоцене вид был распространен на запад несколько шире и заселял север Франции, а также юго-восток Великобритании, откуда исчез уже в историческое время (XVIII в.) в результате разрушения болотных местообитаний проведенной мелиорацией (Gleed-Owen, 2000; Roček, Šandera, 2008).
На юге Центральной Европы горные цепи формируют кольцо, образованное отрогами Восточных Альп, дугой Карпат и замкнутое с юга Балканскими горами. Внутри располагается Среднедунайская (Венгерская) равнина, переходящая к востоку в средневысотные (до 1800 м н. у. м.) Западные Румынские горы и сильно расчлененное холмистое Трансильванское плато (рис. 10). Горные цепи, поднимаясь на пути влажных воздушных течений с Атлантики, служат конденсаторами влаги. За год здесь выпадает 1200–2000 мм осадков, и равнина принимает сток с обращенных к ней макросклонов Альп и Карпат. Для рек Венгерской равнины характерны весенние половодья и летние (июнь) паводки. Годовая амплитуда колебаний уровня Дуная достигает 8 м. Расход воды в устье р. Сава в период половодья превышает среднегодовой в 3 раза, в р. Тиса – в 4.7 раза.
В очерченном регионе находят место многие природно-географические комплексы. Это горные массивы с выраженной высотной поясностью, лесистые холмы и расчлененные эрозией среднегорья с широколиственным лесом и зарослями кустарника, плоские равнины со степными участками. В отдельных районах распространены обширные водно-болотные угодья. Если широкие поймы больших рек (Сава, Драва) болотисты, то долины водотоков в восточной среднегорной части узкие, часто крутосклонные и лесистые. При выходе на северо-восточную часть Среднедунайской низменности р. Тиса извилисто течет по широкой пойменной долине в низких берегах, образуя меандры и старицы, разливы. Здесь распространены заболоченные и пойменные луга, болота (География Венгрии, 2014; География Румынии, 2014; География Словении, 2015; География Хорватии, 2015; Дунай, 2015; Украинские…, 2015; Тиса, 2015).
Размещение R. temporaria и R. arvalis в южно-европейской части зоны симпатрии показано на рис. 10. В этом регионе продвижение остромордой лягушки на юг останавливают сухие степи и агроландшафт. Травяная лягушка не спускается ниже предгорий, где в лесистых низинах обитает совместно с остромордой. Наибольшей численности популяции R. arvalis достигают в северо-восточной части этой низменности – по р. Тиса и к востоку от нее (Sas et al., 2006; Puky, Shad, 2008). Вид был обычен и на западе региона у предгорий Альп (выход на равнину рек Сава и Драва), однако в 60–90-е гг. прошлого века в результате интенсивной мелиорации и гидростроительства, которые уничтожили сырые земли и пойменные леса, вид отсюда почти исчез (Vogrin, 1997). В соседнюю Нижнедунайскую низменность оба вида не проникают, однако в охватывающем ее с юга горном массиве Стара Планина R. temporaria обычна (Petrov, 2007).
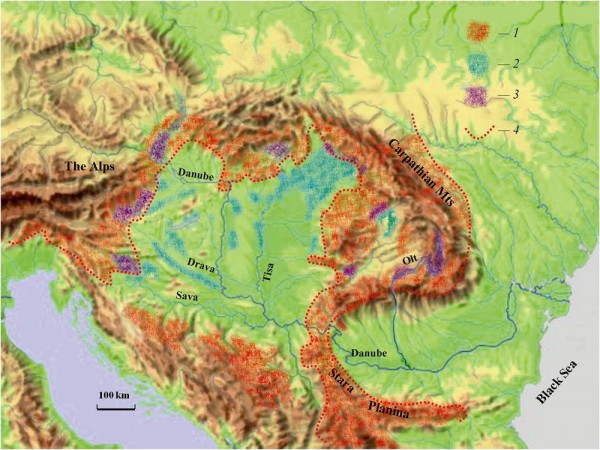
Рис. 10. Распространение травяной и остромордой лягушек на юге зоны симпатрии: 1 – R. temporaria, 2 – R. arvalis, 3 – районы совместного обитания, 4 – граница сплошного ареала R. temporaria. Источники информации: Кушнирук, 1970; Opatrný, 1978; Gasc et al., 1997; Vogrin, 1997; Puky et al., 2005; Petrov, 2007; Covaciu-Marcov et al., 2009a; Glasnović et al., 2009; Jablonski et al., 2012; Cogălniceanu et al., 2013; Vukov et al., 2013. Основа: © László Zentai. 1996: http://lemill.net/content/pieces/uppiece.2008-04-20.3183877257/image_large
Fig. 10. Distribution of the common frog and the moor frog in the south of sympatry zone; 1 – R. temporaria, 2 – R. arvalis, 3 – joint inhabited ranges, 4 – boundary of the continuous range of R. temporaria. Sources: Kushniruk, 1970; Opatrný, 1978; Gasc et al., 1997; Vogrin, 1997; Puky et al., 2005; Petrov, 2007; Covaciu-Marcov et al., 2009 a; Glasnović et al., 2009; Jablonski et al., 2012; Cogălniceanu et al., 2013; Vukov et al., 2013. Basic map: © László Zentai. 1996: http://lemill.net/content/pieces/uppiece.2008-04-20.3183877257/image_large
Другой регион периферии зоны симпатрии травяной и остромордой лягушек, который необходимо рассмотреть, охватывает области юга Русской равнины в секторе с географическими координатами 49°–56° с. ш. и 36°–49° в. д. Район почти полностью занимает лесостепь, он включает также широколиственные леса, а на северо-востоке и элементы южной тайги.
Протянувшиеся параллельно друг другу Среднерусская и Приволжская возвышенности похожи. Это местности, сильно рассеченные речной и овражно-балочной сетью, глубина эрозионного вреза достигает 100–120 м. Небольшие реки немноговодны. Более крупные имеют хорошо разработанные русла и развитые поймы. Многочисленные озера имеют разное происхождение, большинство из них пойменные. Отдельные районы обеих возвышенностей обладают значительными запасами подземных вод. Верхние водоносные горизонты иногда вскрыты оврагами и балками, и вода из них поступает в гидросеть, питает родники и образует заторфованные «потные места». Среднерусская возвышенность характеризуется умеренно-континентальным климатом, Приволжская – континентальным. Северная ее часть относится к зоне с недостаточным увлажнением, а южная – засушливая.
На Среднерусской возвышенности вплоть до южных ее склонов в растительном покрове участвуют, а иногда и преобладают по занимаемой площади, лесные экосистемы: нагорные и байрачные дубравы, пойменные ольшаники. Распространены участки лугово-степной и степной растительности. Для севера Приволжской возвышенности свойственно сочетание таежных и широколиственных лесов (лесистые балки) и степных участков, залесенность местами достигает 50 % территории. Однако уже за 53° с. ш. доля лесного компонента в составе лесостепи не превышает 5 %. Из-за глубокого вреза речных долин, густой сети оврагов и балок и дефицита влажности заболоченность охватывает десятые доли процента территории обеих возвышенностей. Основу составляют пойменные болотные массивы. На Среднерусской возвышенности существуют сфагновые болота, имеющие растительность северного, бореального типа (Кац, 1971; Республика…, 2014; Центрально-Черноземный…, 2014; Заповедник «Белогорье», 2016; Ульяновская…, 2016).
Расположенная между этими возвышенностями Окско-Донская низменность представляет собой плоскую равнину. Северная часть Окско-Донской равнины, ограниченная на юге рекой Окой, получила название Мещерской низменности, а ее центральная и южная части именуются Тамбовской равниной.
В плоской Мещерской низине озера имеют в основном ледниковое происхождение и постепенно заболачиваются. Много и пойменных озер, которые являются остатками старых русел рек. Озера связаны с окружающими болотами, от которых и получают питание. Нередки обширные заболоченные бессточные понижения. У всех рек низменности интенсивно развита пойменная терраса, ширина которой колеблется от десятков метров до 20 км. Крупные реки (Ока, Мокша, Цна, Пра) весной поднимаются на 5–8 м (иногда до 10 м) над меженным уровнем. Продолжительность половодья составляет 25–65 дней. Затапливает не только луговые участки пойм и болота, но и значительные площади лесов.
Пространства Тамбовской равнины эродированы достаточно слабо. Оврагами пересечены, за редким исключением, только крутые и высокие участки склонов долин крупных рек. Наряду с небольшой густотой расчленения невелика и глубина врезания эрозионной сети. Междуречья и межбалочные пространства представляют собой плоскую слабо дренированную равнину. Озерность невысока, это почти исключительно пойменные озера в долинах крупных рек (Дон, Хопер, Воронеж).
По всей Окско-Донской равнине произрастают хвойные, хвойно-широколиственные, вторичные березовые леса. Черноольховые болотные леса распространены в Мещерской низине, на Тамбовской равнине они редко встречаются по заболоченным участкам пойм. В Мещерской низине очень высока заболоченность территории, преобладают переходные и низинные массивы. Ныне почти вся Тамбовская равнина – это агрокультурные ландшафты, леса занимают около 8–10 % территории (Окский…, 2014; Республика…, 2014; Воронежский…, 2016; География Воронежской…, 2016; Мещерская низменность, 2016; Окско-Донская…, 2016).
Травяная лягушка обычна в лесных стациях севера Среднерусской возвышенности. Раньше, хоть и была редка, но встречалась почти до южной ее оконечности. Ныне популяция этой лягушки сохранилась, видимо, лишь по Дону и его правым притокам в восточных отрогах возвышенности (до 50°30′ с. ш.). На Приволжской возвышенности R. temporaria занимает ее северную часть до 52°40' с. ш. На северо-западе (Мордовия) вид спускается в «язык» Окско-Донской равнины, где становится очень редким или считается исчезнувшим к настоящему времени. На всем остальном пространстве равнины теперь сохранились, по-видимому, редкие изолированные поселения на юго-востоке Мещерской низины и на востоке Тамбовской равнины у окраины Приволжской возвышенности. На всем очерченном пространстве юга Русской равнины вид встречается в лесных и кустарниковых стациях по берегам рек, озер. Особенно часто его связывают с выходами на поверхность грунтовых вод в оврагах, балках и прочих эрозионных формах рельефа. Открытых заболоченных долин рек и степных участков травяная лягушка избегает.
Что касается остромордой лягушки, то она здесь самый многочисленный вид Anura в наземных биотопах и встречена повсюду, кроме самых освоенных человеком районов. Пункты с высокой и очень высокой численностью вида приходятся на районы Мещерской низины и на пойменные местообитания в долинах рек. На Среднерусской и Приволжской возвышенностях лягушка встречается и в лесных внепойменных стациях. За пределами Окско-Донской равнины ареал R. arvalis спускается к югу до дельты Дона (Аммон, 1928; Птушенко, 1934; Елисеева, 1967; Глазов, 1975; Леонтьева, Перешкольник, 1982; Гаранин, 1983; Перешкольник, Леонтьева, 1989; Травяная лягушка…, 2001; Лягушка травяная…, 2004; Лягушка травяная…, 2005; Рыжов, Ручин, 2007; Белик, 2010; Кочетков, Шубина, 2011; Травяная лягушка..., 2012б; Центрально-Черноземный…, 2014; Заповедник «Белогорье», 2015; Ручин, 2015; Воронежский…, 2016; Травяная лягушка…, 2011).
Далее к востоку, уже на восточных склонах Приволжской возвышенности, травяная лягушка становится редка, а в степное Заволжье не заходит нигде. К середине ХХ в. вид исчез из среднего течения р. Урал в пределах Оренбургской области и теперь на юг за пределы лесостепи не выходит (Травяyая лягушка…, 1998; Лягушка травяная…, 2014).
Восточная и восток-юго-восточная граница ареала травяной лягушки огибает восточные предгорья Полярного и Приполярного Урала, затем прилегающие, приподнятые (50–200 м н. у. м.) эродированные и рассеченные речной сетью Северо-Сосьвинскую возвышенность и Туринскую равнину. Далее граница идет по отрогам Южного Урала (до 52° с. ш.). В этой протяженной полосе периферии ареала лягушка держится, в основном, по берегам и долинам небольших рек (Травяная лягушка…, 2012а; Амфибии…, 2014; Травяная лягушка…, 2014; Лягушка травяная, 2015). Совершенно очевидно, что продвижению R. temporaria на восток от обозначенной границы препятствуют гидрологические условия Западно-Сибирской равнины. Многочисленные реки испытывают непостоянные и колоссальные весенние половодья – до 11 м превышения над нулем графика. Они иногда полностью заливают поймы на срок до трех месяцев (Максимов, Мерзлякова, 1981). Низкие междуречья равнины заболочены. К югу от этого «угла» ареала преградой для вида оказываются степные пространства с дефицитом влаги, заливаемыми паводком стоячими водоемами и полным отсутствием лесных стаций.
Распространение R. arvalis на восток и юго-восток за пределы зоны симпатрии частично рассмотрено в предыдущих разделах. Восточной преградой оказывается тектонический уступ Среднесибирского плоскогорья вдоль правого берега Енисея и плато Путорана на крайнем северо-востоке ареала (Кузьмин, 2012). К югу от плоскогорья остромордая лягушка заселяет невысокое эродированное и заболоченное плато с Подкаменной Тунгуской. Далее ареал узкой полосой по долине р. Лены протягивается на восток примерно до 124° в. д. (Боркин и др., 1981; Заповедник «Тунгусский», 2014). В Северном Казахстане ареал R. arvalis доходит на юг до 50° с. ш. Для этих степных районов характерен засушливый континентальный климат. Гидрографическая сеть представлена небольшими реками, многочисленными мелкими речками и временными водотоками. Имеется масса (тысячи) озер, представляющих собой бессточные мелководные водоемы. Все водотоки разливаются весной, заполняя низкие места долин, сухие старицы, наполняя озера. С началом спада воды реки и озера превращаются в цепочки плесов. Повсюду по берегам здесь распространены тростниковые и осоковые болота, формируются сплавины (Кац, 1971; Природные…, 2014). Основными стациями остромордой лягушки на данном участке периферии ареала оказываются исключительно болотные местообитания в поймах средних и малых рек, по берегам озер, в долинах временных водотоков (Динесман, 1953; Искакова, 1959; Кривко, 1976; Природные…, 2014).
И несколько слов об обитаемых ландшафтах краевых частей ареалов двух видов. На лишенных лесной растительности равнинах Заполярья, в альпийском поясе европейских гор или горных тундрах Урала популяции R. temporaria приурочены к берегам озер, к заболоченным ложбинам стока, поросшим кустарником или криволесьем, к болотам, не образующим здесь крупных массивов (Топоркова, Зубарева, 1965; Vences et al., 1999, 2003; Гуль, 2001; Кутенков, 2009; Sztatecsny, Hödl, 2009 и др.). Населенные видом урочища что в скалистых тундрах арктического побережья, что в альпийских долинах Балканских гор имеют заметное внешнее сходство (Приложение II, рис. 1, 2). Места обитания R. arvalis на севере (тундра) и на юге (горы Южной Сибири) – это те же обширные заболоченные пространства, что и в других частях ареала (Приложение II, рис. 3, 4). В приведенных примерах ни высота над уровнем моря, ни главные характеристики рельефа, в который вписаны жилые стации лягушек, ни характер растительности не имеют значения для существования видов.
Приложение I
Внешний вид взрослых остромордой и травяной лягушек
Appendix I
The external appearance of R. arvalis and R. temporaria adults


Остромордая (А) и травяная (В) лягушки, помещенные в одинаковые стеклянные сосуды
R. arvalis (А) and R. temporaria (В). Frogs are placed in the similar glass vessels
Приложение II
Места обитания травяной и остромордой лягушек в краевых частях ареалов за пределами зоны симпатрии
Appendix II
Habitats of the common frog and the moor frog in marginal parts of their geographic ranges outside of sympatry zone
Rana temporaria

Рис. 1. Скалистая тундра Кольского п-ова. Координаты 69°00′ с. ш., 36°00′ в. д. Крайний север ареала R. temporaria (http://www.chinamobil.ru/travel/kolski2012/IMG_1534.jpg)
Fig. 1. Rocky tundra of the Kola Peninsula. Coordinates: 69°00′ N, 36°00′ E. Northernmost part of R. temporaria geographic range (http://www.chinamobil.ru/travel/kolski2012/IMG_1534.jpg)

Рис. 2. Вид в массиве Пирин, Балканские горы. Координаты: 41°40′ с. ш., 23°30′ в. д. Высота ок. 1500 м н. у. м. Крайний юг ареала R. temporaria (http://static.bnews.bg/16ce92e4e85c.jpg)
Fig. 2. A view in the Pirin massif, Balkan Mountains. Coordinates: 41°40′ N, 23°30′ E. Altitude cca. 1500 m a. s. l. Southernmost part of R. temporaria geographic range (http://static.bnews.bg/16ce92e4e85c.jpg)
Rana arvalis

Рис. 3. Большеземельская тундра. Координаты 68° с. ш., 63° в. д. Крайний север европейской части ареала R. arvalis (http://rilmark.ru/catalog/20121108/20121108:571-799-610/20121108:571-799-610-download.jpg)
Fig. 3. The Bolshezemel'skaya Tundra. Coordinates: 68° N, 63° E. Norternmost part of R. arvalis geographic range in Europe (http://rilmark.ru/catalog/20121108/20121108:571-799-610/20121108:571-799-610-download.jpg)

Рис. 4. Тоджинская среднегорная котловина в горной системе Саяны, Южная Сибирь. Координаты: 52°40′ с. ш., 97°30′ в. д. Высота около 1200 м н. у. м. Юго-восточная периферия ареала R. arvalis (http://muzmix.com/images/songs/101868/6.jpg)
Fig. 4. The mid-mountain Todzhinskaya Hollow in Sayany Mountains, South Siberia. Coordinates: 52°40′ N, 97°30′ E. Altitude cca. 1200 m a. s. l. The south-east periphery of R. arvalis geographic range (http://muzmix.com/images/songs/101868/6.jpg)
Заключение
Распространение травяной лягушки повсюду в ареале связано с ландшафтными урочищами, для которых характерен выраженный микро- или мезорельеф. Гидрографическая сеть может быть хорошо развитой или примитивной, но наличие небольших постоянных и временных водотоков и проточных озер обязательно: непромерзающие, насыщенные кислородом зимовальные водоемы определяют сам факт присутствия данного вида в той или иной местности. Обычно представлены и различные малые водоемы. Особенности рельефа не допускают существенных половодий и паводков на реках и в озерно-речных системах, которые сопровождались бы большими разливами. Часто присутствуют небольшие болота евтрофно-мезотрофного ряда, участки заболоченных земель. Распространению вида сопутствует лесная растительность, а в тундре и в альпийском ландшафте – кустарниковые заросли и криволесье. Нерест происходит в прибрежных, пойменных и сходных по гидрологическому режиму замкнутых относительно глубоких временных и постоянных объемах воды, порой далеко удаленных от мест зимовки. Коллективное кучное икрометание препятствует смыву кладок с мелководий. В условиях достаточного увлажнения и благодаря относительно высокой вагильности в период нагула вид может использовать практически любые местообитания указанных ландшафтных урочищ. Более того, при неблагоприятных условиях (засуха) эти амфибии способны, преодолевая значительные расстояния, отыскивать влажные стации и концентрироваться там (Гумилевский, 1941; Сергеев, Ветшева, 1942).
В противоположность травяной, для мест распространения остромордой лягушки характерна принципиально иная морфология ландшафта. Остромордая лягушка оказывается, в терминах ландшафтоведения, видом плоских и вогнутых мезоформ и гидроморфных участков равнинного рельефа с застойными водоемами. В любой местности вид населяет прежде всего обводненные болотные урочища, заливаемые половодьем участки долин, тростниковые сплавины, сырые вересковые пустоши и прочие «мокрые земли», чем вполне оправдывает свое название «болотная» или «вересковая» лягушка, принятое во многих европейских языках. В разных физико-географических условиях такие урочища то суживаются до оторочки по берегам речек, озер или морских заливов, то занимают господствующее положение в ландшафте, как, например, в Западной Сибири или на равнинных тундрах Севера. «Тундра – это в некотором смысле чудовищно разросшееся таежное моховое болото» (Еськов, 2008, с. 235). В населенных видом урочищах древесная или кустарниковая растительность часто не представлена вообще. Зимовка, нерест (рассеянное размещение кладок икры) и нагул R. arvalis осуществляются практически в одном и том же биотопе. Остромордые лягушки в норме оседлы, однако у сеголеток обнаружено весьма интенсивное расселение: расстояние в 700 м они проходят за неделю (Ищенко, 2008; см. также Glandt, 2014).
Только там, где происходит взаимопроникновение «коренных» ландшафтов каждого из видов – в обширных областях северной и западной частей Восточно-Европейской равнины и прилегающей к Уралу части Западно-Сибирской равнины, – удается наблюдать примеры действительной симпатрии R. arvalis и R. temporaria.
Очевидными форпостами распространения R. temporaria являются горы. Из 15 тыс. км границы ареала, если ее очертить по крайним точкам нахождения вида на материке, половина приходится на горные массивы и подобные природные обстановки. На востоке это Урал. Благодаря горам Скандинавии и низким скалистым тундрам Кольского п-ова она достигает рубежей континента в Северной Европе. На юго-западе вид заселяет Пиренеи, на юге – южные отроги Альп, Балканские горы и Карпаты. В горы Балканского п-ова глубоко проникают свойственные умеренным широтам лесные ландшафты (Гвоздецкий, Голубчиков, 1987), и по этим горам (массив Родопы) ареал вида доходит к югу до 41°20′ с. ш. (Petrov et al., 2006), до побережья Эгейского моря здесь остается всего 50 км.
Ареал R. arvalis простирается от западных (4° в. д., юго-запад Голландии) до восточных (124° в. д., средняя Лена) пределов на 7800 км, а по меридиану 70° в. д. – на 2400 км. Все горные цепи Европы и Урал, которые способствуют распространению R. temporaria, остромордой лягушке, наоборот, препятствуют. Здесь уместно вспомнить давнее заключение Б. Стугрена о том, что R. arvalis неспособна преодолевать горы (Stugren, 1966). Его необходимо дополнить одним существенным обстоятельством: она не поднимается лишь в европейские складчатые горы. Но выровненные эрозией и вознесенные остатки древних равнин в горах Южной Сибири – это те же населенные видом ландшафтные единицы, что и повсюду в ареале. Благодаря им вид не только может существовать в высокогорье Алтая, но и смог проникнуть в северный Китай (см. Glandt, 2014).
Недавно была предпринята попытка с помощью специальных программных средств осуществить количественную оценку отношения к физико-географическим условиям и установить потенциальные ареалы трех видов Anura (в том числе R. arvalis и R. temporaria) на территории бывшего СССР (Пузаченко и др., 2011). За основу взяты все известные географические точки находок лягушек. С использованием баз данных и информации со спутниковых карт в соответствие этим точкам поставлены значения многочисленных (108) климатических переменных, значения дифференцированных вегетационных индексов NDVI (или дифференцированного индекса растительности, или биологической продуктивности). Рельеф был представлен 6 характеристиками. Путем соответствующих процедур все климатические переменные сведены к четырем параметрам: континентальность, в основном зимой; тепло-влагообеспеченность с октября по март; сумма осадков и биологическая продукция за май – сентябрь; средние температуры за май – сентябрь. Переменные характеристики рельефа сведены к четырем компонентам: экспозиция поверхности; уклон поверхности; иерархическая организация – 4 уровня высоты; форма поверхности – выпуклые и вогнутые формы. Точки встреч вида, характеризуемые всеми переменными, будут соответствовать определенной подобласти общего экологического пространства, которую можно определить как видоспецифическую (= многомерная фундаментальная экологическая ниша). «Проекция этого многомерного пространства на территорию даст потенциальный ареал с внутренней структурой, в котором каждой “точке” земной поверхности ставится в соответствие вероятность обнаружения вида» (с. 341). Построенные ареалы интерполируют возможное размещение вида на существенно бóльшую территорию с подобным климатом и рельефом.
Не углубляясь в детали (например, весьма сомнительным критерием видится отношение видов к тепло- и влагообеспеченности зимой; «Очевидно, что R. temporaria в сравнении с R. arvalis предпочитает более теплую зиму…» (с. 347)), необходимо обратить внимание на два упущения. Во-первых, авторы оценивали отношение видов к переменным, соответствующим географическому пункту находки, а не местообитанию популяций конкретных видов, что они вполне осознают (см. с. 349 цитируемой работы). И показатели таких существенных, на мой взгляд, условий, как гидрологический режим территории, густота речной сети, заболоченность (процент площади болот и заболоченных земель от всей площади (Киреев, 1984), мозаичность или низменный характер ландшафта и т. п., в анализ не вошли. По-видимому (и к большому сожалению), полных баз данных с этими физико-географическими характеристиками просто не существует.
Во-вторых, в задачи Ю. Г. Пузаченко с коллегами входил анализ частей ареалов, расположенных лишь в границах бывшего СССР. Между тем, как показано выше, наиболее яркие черты пространственной дивергенции R. arvalis и R. temporaria мы находим именно за западными пределами анализируемого авторами пространства. Причем решающая роль в этом «расхождении» видов принадлежит рельефу при почти полном подобии большинства климатических составляющих. Выводу о том, что «рельеф вносит некоторую дифференциацию в размещение видов, создавая основу для разделения территории R. arvalis и R. temporaria. Первая тяготеет в целом к возвышенностям, вторая – к долинам» (с. 348), категорически противоречит реальная ситуация в Западной Европе.
В результате получилось, что потенциальный ареал травяной лягушки на востоке (см. рис. 6 цитируемой работы) занимает всю Западно-Сибирскую равнину (что в принципе невозможно ввиду специфики экологии данного вида) и не распространяется на возвышенности и в горы Сибири. Более того, согласно вероятностной модели, на Северном Урале R. temporaria вообще не должна бы существовать (рис. 6б) – вопреки имеющимся фактам. Остромордая лягушка, напротив, может, хотя и с небольшой вероятностью, не только обитать почти во всех горных системах азиатской части России, но и встречаться на Малом Кавказе и в Армянском нагорье. А Среднесибирскую низменность (среднее течение Оби) с большим числом точек находок вида, как и сильно заболоченную и испещренную озерами Яно-Индигирскую низменность, модель представляет как пустые области (см. рис. 5).
Библиография
Аврамова О. С., Булахов В. Л., Бобылев Ю. П. Брачное поведение и системы «спаривания» у озерной и остромордой лягушек // Групповое поведение животных: Доклады участников II Всесоюзн. конф. по поведению животных. М.: Наука, 1976. С. 6–8.
Аммон П. Л. Список амфибий и рептилий Тульской губернии // Журн. Тульский край. Тула, 1928. С. 44–52.
Амфибии Красной книги Башкортостана. Травяная лягушка / В. Ф. Хабибуллин. URL: http://redbook.ru/article80.html (дата обращения 22.03.2014).
Ануфриев В. М., Бобрецов А. В. Амфибии и рептилии. Фауна европейского Северо-Востока России . Т. IV. СПб.: Наука, 1996. 131 с.
Байкало-Ленский заповедник . URL: http://irkipedia.ru/content/baykalo_lenskiy_zapovednik (дата обращения 12.12.2014).
Банников А. Г. Экологические условия зимовок травяной лягушки (Rana temporaria L.) в Московской области // Сборник научных студенческих работ МГУ. Зоология. М., 1940. Вып. XVI. С. 41–64.
Банников А. Г., Денисова М. Н. Очерки по биологии земноводных . М.: Учпедгиз, 1956. 168 с.
Барабаш И. И. Обзор стационарного распределения позвоночных животных в Кададинском опытном лесничестве Пензенской области // Бюллетень Воронежского общества естествоиспытателей. Воронеж, 1939. Т. 3. Вып. 2. С. 21–29.
Белик В. П. Материалы к фауне и экологии земноводных степного Придонья // Современная герпетология. 2010. Т. 10. Вып. 3/4. С. 89–100.
Белимов Г. Т., Седалищев В. Т. К биологии остромордой лягушки, обитающей в Якутии // Экология. 1979. № 5. С. 92–95.
Бельгия — Горная энциклопедия . URL: http://www.mining-enc.ru/b/belgiya (дата обращения 19.04.2015).
Блинова Т. К. Земноводные северной лесостепи Зауралья // Вид и его продуктивность в ареале: Материалы 4-го Всесоюзного совещания. Ч. V. Вопросы герпетологии. Свердловск, 1984. С. 5–6.
Боркин Л. Я., Белимов Г. Т., Седалищев В. Т. О распространении лягушек рода Rana в Якутии // Герпетологические исследования в Сибири и на Дальнем Востоке. Л.: ЗИН АН СССР, 1981. С. 18–24.
Брысова Л. П., Кожевникова Р. К., Коротков И. А., Криницкий В. В. Физико-географические условия северо-восточного Алтая и задачи изучения его природы // Труды Алтайского государственного заповедника. Горно-Алтайск, 1961. Вып. 4. С. 3–32.
Вартапетов Л. Г., Ануфриев В. М. Остромордая лягушка в Притазовской северной тайге // Вид и его продуктивность в ареале: Материалы 4-го Всесоюзного совещания. Ч. V. Вопросы герпетологии. Свердловск, 1984. С. 7–8.
Витимский заповедник . URL: http://vitimskiy.ru/index.php/sovremennoe-sostoyanie-ekosistem (дата обращения 12.12.2014).
Вознийчук О. П., Куранова В. Н. Земноводные и пресмыкающиеся Катунского заповедника и сопредельной территории (Центральный Алтай) // Современная герпетология. 2008. Т. 8. Вып. 2. С. 101–117.
Воронежский заповедник . URL: http://zapovednik-vrn.ru/o-zapovednike1/priroda/ (дата обращения 17.02.2016).
Гаранин В. И. Земноводные и пресмыкающиеся Волжско-Камского края . М.: Наука, 1983. 175 с.
Гвоздецкий Н. А., Голубчиков Ю. Н. Горы . М.: Мысль, 1987. 399 с.
География Венгрии . http://www.gecont.ru/articles/geo/hungury.htm (дата обращения 22.03.2014).
География Воронежской области . URL: https://ru.wikipedia.org/wiki/География _Воронежской_области (дата обращения 17.02.2016).
География Нидерландов . URL: http://www.gecont.ru/articles/geo/holland.htm (дата обращения 22.03.2014).
География Румынии . URL: http://www.gecont.ru/articles/geo/romania.htm (дата обращения 22.03.2014).
География Словении . URL: http://www.gecont.ru/articles/geo/slovenia.htm (дата обращения 19.04.2015).
География Хорватии . URL: https://ru.wikipedia.org/wiki/География Хорватии (дата обращения 19.04.2015).
Глазов М. В. О роли остромордых лягушек в регуляции численности беспозвоночных в биоценозе дубравы // Бюллетень МОИП. Отд. биол. 1975. Т. 80. № 6. С. 59–66.
Гуль И. Р. Земноводные и пресмыкающиеся хребта Черногора (Украинские Карпаты) // Вопросы герпетологии: Материалы I съезда Герпетологического общества им. А. М. Никольского (Пущино). М., 2001. С. 74–75.
Гумилевский Б. А. О некоторых эколого-фаунистических исследованиях на Валдайской возвышенности // Известия Всесоюзного Географического общества. 1941. Т. 73. Вып. 1. С. 129–136.
Дарвинский заповедник . URL: http://www.darvinskiy.ru/ecosistem.php (дата обращения 21.12.2014).
Динесман Л. Г. Амфибии и рептилии юго-востока Тургайской столовой страны и Северного Приаралья // Труды ИГ АН СССР. Вып. 54. М., 1953. С. 383–422.
Дунай . URL: https://ru.wikipedia.org/wiki/Дунай (дата обращения 19.04.2015).
Елисеева В. И. Фауна низших наземных позвоночных Центрально-Черноземного заповедника // Труды Центрально-Черноземного заповедника. Вып. 10. М.: Лесная промышленность, 1967. С. 83–87.
Еськов К. Ю. Удивительная палеонтология . М.: ЭНАС, 2008. 312 с.
Заповедник Азас . URL: http://green-azas.ru/zapovednik-azas/fiziko-geograficheskaya-kharakteristika.html (дата обращения 12.12.2014).
Заповедник «Белогорье» . URL: http://www.zapovednik-belogorye.ru (дата обращения 09.02.2016).
Заповедник «Кузнецкий Алатау» . Кемерово, 1999. 255 с.
Заповедник «Кузнецкий Алатау». Фауна. URL: http://www.kuz-alatau.ru/priroda/fauna/#midl (дата обращения 12.12.2014).
Заповедник «Тунгусский» . URL: http://www.tunzap.ru (дата обращения 12.12.2014).
Ивантер Э. В. Материалы по экологии травяной лягушки на Севере // Труды заповедника «Кивач». Петрозаводск, 1969. Вып. 1. С. 136–145.
Иноземцев А. А. Роль насекомоядных птиц в лесных биоценозах . Л.: ЛГУ, 1978. 264 с.
Искакова К. Земноводные Казахстана . Алма-Ата, 1959. 92 с.
Ищенко В. Г. Долговременные исследования демографии популяций амфибий: современные проблемы и методы // Вопросы герпетологии: Материалы III съезда Герпетологического общества им. А. М. Никольского (Пущино-на-Оке, 9–13 октября 2006 г.). СПб., 2008. С. 151–169.
Ищенко В. Г., Леденцов А. В. Влияние условий среды на динамику возрастной структуры популяций остромордой лягушки // Влияние условий среды на динамику структуры и численности популяций животных. Свердловск: УНЦ АН СССР, 1987. С. 40–51.
Калецкая М. Л. Фауна земноводных и пресмыкающихся Дарвинского заповедника и ее изменения под влиянием Рыбинского водохранилища // Рыбинское водохранилище. Ч. 1. М., 1953. С. 171–186.
Катунский природный биосферный заповедник . http://www.turistka.ru/altai/info.php?ob=1202 (дата обращения 14.10.2015).
Кац Н. Я. Болота земного шара . М.: Наука, 1971. 296 с.
Киреев Д. М. Эколого-географические термины в лесоведении . Новосибирск: Наука, 1984. 182 с.
Кондрашев С. Л., Гнюбкин В. Ф., Диментман А. М., Орлов О. Ю. Роль зрительных стимулов в брачном поведении самцов травяной лягушки (Rana temporaria), серой жабы (Bufo bufo) и зеленой жабы (Bufo viridis) // Зоологический журнал. 1976. Т. 55. Вып. 7. С. 1027–1037.
Корнеева Т. М., Быков А. В., Речан С. П. Наземные позвоночные низовьев реки Онеги . М.: Наука, 1984. 89 с.
Коросов А. В. Экология обыкновенной гадюки (Vipera berus L.) на Севере (факты и модели) . Петрозаводск, 2010. 264 с.
Коросов А. В., Фомичев С. Н. Ведущие факторы размещения амфибий на островах Кижского архипелага // 10 лет экологическому мониторингу музея-заповедника «Кижи». Петрозаводск, 2005. С. 120–126.
Коросов А. В., Фомичев С. Н. Территориальное размещение бурых лягушек в период размножения // Экология. Экспериментальная генетика и физиология. Труды КНЦ РАН. Вып. 11. Петрозаводск, 2007. С. 85–92.
Коротков Ю. М., Короткова Е. Б. Некоторые данные об экологии остромордой лягушки Rana terrestris в Туве // Экология. 1976. Вып. 3. С. 102–103.
Кочетков С. Н., Шубина Ю. Э. Распространение и экология травяной лягушки в Липецкой области // Вопросы герпетологии: Материалы IV съезда Герпетологического общества им. А. М. Никольского (Казань, 12–17 октября 2009 г.). СПб., 2011. С. 113–118.
Красавцев Б. А. Материалы по экологии остромордой лягушки (Rana terrestris Andrz.) // Вопросы экологии и биоценологии. Л.: ЛГУ, 1939. Вып. 4. С. 253–267.
Кривко А. М. К распространению и экологии амфибий Северного Прикаспия // Биологические науки. Алма-Ата, 1976. Вып. 3. С. 46–48.
Кузьмин С. Л. Земноводные и пресмыкающиеся северо-запада Москвы // Земноводные и пресмыкающиеся Московской области. М.: МОИП, 1989. С. 48–60.
Кузьмин С. Л. Земноводные бывшего СССР . М.: Товарищество научных изданий КМК, 2012. 370 с.
Куранова В. Н. Динамика популяций бесхвостых земноводных на юго-востоке Западной Сибири // Вопросы герпетологии: Материалы I съезда Герпетологического общества им. А. М. Никольского (Пущино). М., 2001. С. 147–149.
Куранова В. Н., Каштанова М. В. Земноводные и пресмыкающиеся Ильменского заповедника и окрестностей г. Миасс (Челябинская обл.) // Вопросы герпетологии: Материалы I съезда Герпетологического общества им. А. М. Никольского (Пущино). М., 2001. С. 149–152.
Кутенков А. П. Размножение и некоторые черты экологии остромордой лягушки в средней Карелии // Вид и его продуктивность в ареале: Материалы 4-го Всесоюзного совещания. Ч. V. Вопросы герпетологии. Свердловск, 1984. С. 25.
Кутенков А. П. Размножение бесхвостых амфибий южной Карелии и влияние на него некоторых форм хозяйственной деятельности // Антропогенные воздействия на природу заповедников. М.: ЦНИЛ Главохоты РСФСР, 1990. С. 38–50.
Кутенков А. П. Динамика размеров печени, жировых тел и гонад у травяных (Rana temporaria) и остромордых (R. arvalis) лягушек // Экология наземных позвоночных. Петрозаводск, 1991. С. 14–24.
Кутенков А. П. Экология травяной лягушки (Rana temporaria L., 1758) на Северо-Западе России . Петрозаводск, 2009. 140 с.
Кутенков А. П., Коросов А. В. Материалы по биогеографии амфибий Карело-Кольского края // Биогеография Карелии. Труды КНЦ РАН. Сер. Б. Вып. 2. Петрозаводск, 2001. С. 103–109.
Кутенков А. П., Панарин А. Е., Шкляревич Ф. Н. Экология размножения бесхвостых амфибий Карелии и Кольского полуострова // Наземные позвоночные животные в заповедниках севера европейской части РСФСР. М.: ЦНИЛ Главохоты РСФСР, 1990. С. 54–70.
Кушнирук В. А. О зимовках некоторых земноводных в условиях западных областей Украины // Вопросы герпетологии: Автореф. докл. 3 Всесоюзной герпетол. конф. Л.: ЛГУ, 1964. С. 37–38.
Кушнирук В. А. К изучению биологии остромордой лягушки на западе Украины // Фауна Молдавии и ее охрана: Материалы докладов Первой республиканской конф. Кишинев, 1970. С. 141.
Лада Г. А. Эколого-фаунистический анализ амфибий центрального Черноземья: Автореф. дис. ... канд. биол. наук . СПб., 1993. 22 с.
Леонтьева О. А. Некоторые вопросы экологии бурых лягушек на северной границе их ареала // Взаимодействие организмов в тундровых экосистемах: Тез. докл. Всесоюзной совещ. Сыктывкар, 1989. С. 146.
Леонтьева О. А., Перешкольник С. Л. Индикаторная роль герпетофауны в биоценозах с различной антропогенной трансформацией // Животный мир центра лесной зоны европейской части СССР. Калинин, 1982. С. 49–62.
Лобанов В. А. Распространение остромордой лягушки в Большеземельской тундре // Вопросы герпетологии: Автореф. докл. 4 Всесоюзной герпетол. конф. Л.: Наука, 1977. С. 134–135.
Лягушка остромордая. Герпетофауна Тюменской области . URL: http://herptyumen.narod.ru/amphibia/rana_arvalis.html (дата обращения 26.10.2015).Лягушка травяная / В. А. Кривошеев // Красная книга Ульяновской области (грибы, животные). Т. 1. Ульяновск, 2004. URL: http://geohobby.ru/enc/02-3-1-002.html (дата обращения 24.09.2014).
Лягушка травяная / О. А. Ермаков // Красная книга Пензенской обл. Т. 2. Пенза, 2005. С. 105.
Лягушка травяная / А. И. Файзулин, А. Г. Бакиев // Красная книга Самарской обл. Т. II. Редкие виды животных. С. 237. URL: https://issuu.com/dd_nn/docs/krasnaq_kniga_samarskoj _oblasti._tom_2 (дата обращения 14.09.2014).
Лягушка травяная. Герпетофауна Тюменской области . URL: http://herptyumen.narod.ru/amphibia/rana_temporaria.html (дата обращения 26.10.2015).
Ляпков С. М. Многолетняя динамика численности популяций бурых лягушек Подмосковья: естественные флуктуации или результат усиления антропогенных воздействий? // Вопросы герпетологии: Материалы I съезда Герпетологического общества им. А. М. Никольского (Пущино). М., 2001. С. 171–173.
Максимов А. А., Мерзлякова Е. П. Характеристика разливов в пойме Оби за 1968–1977 гг. // Сукцессии животного населения в биоценозах поймы реки Оби. Новосибирск, 1981. С. 165–174.
Малеев В. Г. К распространению амфибий и рептилий на территории Усть-Ордынского Бурятского автономного округа (верхнее Приангарье) // Байкальский зоологический журнал. 2009. № 1. С. 48–49.
Мещерская низменность . URL: https://ru.wikipedia.org/wiki/Мещёрская_низменность (дата обращения 17.02.2016).
Никитенко М. Ф. Земноводные Советской Буковины // Животный мир Советской Буковины. Черновцы, 1959. С. 160–205.
Окский заповедник . URL: http://oksky-reserve.ru/index/geography/ (дата обращения 21.12.2014).
Окско-Донская равнина . URL: http://ru.wikipedia.org/wiki/Окско-Донская_равнина (дата обращения 17.02.2016).
Остромордая лягушка // Красная книга Республики Бурятия. URL: http://www.minpriroda-rb.ru/redbook/2004/animals/amphibious_index.php?ELEMENT_ID=2449 (дата обращения 24.09.2014а).
Остромордая лягушка / О. Н. Чернышова, Л. Н. Ердаков, В. Н. Куранова, М. В. Пестов) // Информационные материалы к герпетофауне Сибири. URL: http://www.balatsky.de/NSO/Amphibii/Rana_arvalis.htm (дата обращения 09.09.2014б).
Панченко И. М. Эффективность размножения остромордой лягушки юго-востока Мещеры // Экология. 1980. № 6. С. 95–98.
Перешкольник С. Л., Леонтьева О. А. Многолетние наблюдения за изменением герпетофауны Приокско-террасного государственного заповедника // Земноводные и пресмыкающиеся Московской области. М.: МОИП, 1989. С. 84–96.
Пикулик М. М. Земноводные Белоруссии . Минск, 1985. 192 с.
Преображенская Е. С., Байкалова А. С. Численность и биотопическое распределение земноводных вне водоемов // Животный мир южной тайги. Проблемы и методы исследования. М.: Наука, 1984. С. 83–90.
Природные объекты Актюбинской области . URL: http://kzgov.docdat.com/docs/76/index-3326806.html (дата обращения 05.09.2014).
Птушенко Е. С. Наземные позвоночные Курского края. 1. Амфибии и рептилии // Бюллетень МОИП. Отд. биол. 1934. Т. 33. Вып. 1. С. 35–51.
Пузаченко Ю. Г., Кузьмин С. Л., Сандлерский Р. Б. Количественная оценка ареалов (на примере представителей рода Rana) // Журнал общей биологии. 2011. Т. 72. № 5. С. 339–354.
Равкин Ю. С. Численность и распределение земноводных в лесной зоне Западной и Средней Сибири // Экология. 1976. № 5. С. 53–61.
Республика Мордовия. Географический обзор . URL: www.geografia.ru/mordovia.html (дата обращения 04.09.2014).
Ручин А. Б. Экология земноводных и пресмыкающихся Мордовии. Сообщение 2. Травяная лягушка, Rana temporaria Linnaeus, 1758 // Труды Мордовского государственного природного заповедника. Саранск, 2015. Вып. 14. С. 344–358.
Рыжевич К. К. Ландшафтно-типологическая и ландшафтно-географическая изменчивость соотношения обилия травяной (Rana temporaria) и остромордой (R. arvalis) лягушек в Беларуси // Вопросы герпетологии: Материалы I съезда Герпетологического общества им. А. М. Никольского (Пущино). М., 2001. С. 254–255.
Рыжов М. К., Ручин А. Б. Биология остромордой лягушки Rana arvalis в Мордовии. Сообщение 1. Распространение, численность и биотопы // Биологические науки Казахстана. 2007. № 3. С. 33–39.
Северцов А. С., Ляпков С. М., Сурова Г. С. Соотношение экологических ниш травяной (Rana temporaria) и остромордой (R. arvalis) лягушек (Anura, Amphibia) // Журнал общей биологии. 1998. Т. 59. № 3. С. 279–301.
Седалищев В. Т., Белимов Г. Т., Бекенева Г. Н. Некоторые морфофизиологические адаптации остромордой лягушки (Rana arvalis) в южной Якутии // Вопросы герпетологии: Автореф. докл. 5 Всесоюзной герпетол. конф. Л.: Наука, 1981. С. 122–123.
Сергеев А. М., Ветшева А. Г. К вопросу о влиянии засухи на динамику численности травяной лягушки в европейской части СССР // Зоологический журнал. 1942. Т. 21. Вып. 5. С. 202–206.
Стариков В. П., Матковский А. В. Распространение и некоторые стороны экологии травяной лягушки (Rana temporaria L., 1758) севера Западной Сибири // Вестник Красноярского государственного аграрного университета. 2011. № 12. С. 125–128.
Тиса . URL: http://dic.academic.ru/dic.nsf/enc_geo/4887/%D0%A2%D0%B8%D1%81%D0%B0 (дата обращения 19.04.2015).
Топоркова Л. Я., Варфоломеев В. В. К распределению амфибий и рептилий на хребте Басеги // Вид и его продуктивность в ареале: Материалы 4-го Всесоюз. совещ. Ч. V. Вопросы герпетологии. Свердловск, 1984. С. 44–45.
Топоркова Л. Я., Зубарева Э. Л. Материалы по экологии травяной лягушки на Полярном Урале // Труды Института биологии УФ АН СССР. 1965. Вып. 38. С. 189–193.
Травяная лягушка / А. В. Давыгора // Красная книга Оренбургской обл. Животные и растения. Оренбург, 1998. URL: http://oopt.aari.ru/rbdata/1100/bio/8480 (дата обращения 24.09.2014).
Травяная лягушка / М. В. Дидорчук // Красная книга Рязанской области. Редкие и находящиеся под угрозой исчезновения виды животных. Рязань, 2001. С. 138.
Травяная лягушка / С. В. Репитунов, А. И. Масалыкин // Красная книга Воронежской обл. Т. 2. Животные. Воронеж, 2011. С. 258.
Травяная лягушка / В. Г. Ищенко // Красная книга Курганской области. Земноводные. Курган, 2012а. С. 94.
Травяная лягушка / Г. А. Лада, А. С. Соколов // Красная книга Тамбовской области: Животные. Тамбов, 2012б. С. 208.
Травяная лягушка / В. Л. Вершинин, М. В. Чибиряк // Уральская экологическая энциклопедия. URL: http://ecoinf.uran.ru/00003437.html (дата обращения 31.03.2014).
Украинские Карпаты — география . URL: http://uchebnikionline.com/geografia/geografiya_-_oliynik_yab/ukrayinski_karpati.htm (дата обращения 17.10.2015).
Ульяновская область. Географический обзор . URL: http://www.geografia.ru/ulyan.html (дата обращения 17.02.2016).
Фомичев С. Н. Экология островных популяций бурых лягушек в Карелии: Дис. … канд. биол. наук . Петрозаводск, 2004. 188 с.
Центрально-Черноземный биосферный заповедник . URL: http://zapoved-kursk.ru (дата обращения 05.09.2014).
Шапошников Л. В., Головин О., Сорокин М., Тараканов А. Животный мир Калининской области . Калинин, 1959. 460 с.
Шарлеман Э. В. Заметка о фауне пресмыкающихся и земноводных окрестностей Киева // Материалы к познанию фауны юго-западной России. Киев, 1917. С. 1–17.
Шварц С. С., Ищенко В. Г. Пути приспособления наземных позвоночных к условиям существования в Субарктике. Т. 3. Земноводные . Свердловск, 1971. 60 с.
Швецов Ю. Г. Земноводные и пресмыкающиеся северного Прибайкалья // Вопросы герпетологии: Автореф. докл. 4 Всесоюзной герпетол. конф. Л.: Наука, 1977. С. 232–235.
Шляхтин Г. В., Табачишин В. Г., Завьялов Е. В., Табачишина И. Е. Животный мир Саратовской области. Кн. 4. Амфибии и рептилии . Саратов, 2005. 116 с.
Эпова Л. А., Куранова В. Н., Бабина С. Г. Видовое разнообразие, биотопическое распределение и численность земноводных и пресмыкающихся заповедника «Кузнецкий Алатау» в градиенте высотной поясности (юго-восток Западной Сибири) // Вестник Томского государственного университета. Биология. 2013. № 4 (24). С. 77–97.
Яковлев В. А. Распространение и биотопическое размещение амфибий и рептилий в Алтайском заповеднике // Вопросы герпетологии: Автореф. докл. 4 Всесоюзной герпетол. конф. Л.: Наука, 1977. С. 241–242.
Яковлев В. А. О размножении и развитии остромордой лягушки, Rana arvalis в Алтайском заповеднике // Экология и систематика амфибий и рептилий. Труды ЗИН АН СССР. 1979. Т. 89. С. 109–117.
Яковлев В. А. К вертикальному распространению и размножению остромордой лягушки в Алтайском заповеднике // Экология. 1980. № 4. С. 89–90.
Яковлев В. А. Материалы по размножению остромордой лягушки в условиях высокогорий // Экология. 1981. № 1. С. 97–101.
Andrén C., Nilson G. Effects of acidification on Swedish brown frogs // Memoranda soc. fauna et flora fennica. 1988. Vol. 64. № 3. P. 139–141.
Bea A., Rodriquez-Teijerio J. D., Jover L. L. Relations between meteorological variables and the initiation of the spawning period in populations of Rana temporaria L. in the Atlantic region of the Basque Country (northern Spain) // Amphibia – Reptilia. 1986. Vol. 7. № 1. P. 23–31.
Berger L. Gady I płazy (Amphibia et Reptilia) // Fauna słodkowodna Polski. № 4. Warszawa; Poznań, 1975. 109 s.
Cogălniceanu D., Szekely P., Samoilă C, Iosif R., Tudor M., Plăiaşu R., Stănescu F., Rozylowicz L. Diversity and distribution of amphibians in Romania // ZooKeys. 2013. № 296. P. 35–57. DOI: 10.3897/zookeys.296.4872.
Costanzo J., de Amaral C., Rosendale A., Lee R. Hibernation physiology, freezing adaptation and extreme freeze tolerance in a northern population of the wood frog. // Journ. of exp. boil. 2013. Vol. 216. P. 3461–3473.
Covaciu-Marcov S.-D., Cicort-Lucaciu A., Gaceu O., Sas I., Ferenţi S., Bogdan H. The herpetofauna of the south-western part of Mehedinţi County, Romania // North-western journ. of zool. (Romania). 2009a. Vol. 5. № 1. P. 142–164. URL: http://www.lacerta.de/AS/Bibliografie/BIB_4967.pdf (дата обращения 23.08. 2015).
Covaciu-Marcov S.-D., Dincă I., Dimancea N. The herpetofauna of the hydrographical basin of the Moca stream from Valea lui Mihai town, Bihor County, Romania // Biharean biologist. 2009b. Vol. 3. № 2. P. 125–131. URL: http://biozoojournals.ro/bihbiol/cont/v3n2/bb.031118.Covaciu.pdf (дата обращения 30.04. 2015).
Delft J. van, Creemers R. Distribution, status and conservation of the moor frog (Rana arvalis) in the Netherlands // Zeitschrift für feldherpetologie. 2008. Suppl. 13. P. 255–268. URL: http://www.ravon.nl/Portals/0/Pdf/21-van%20Delft%20%20Cremers%20DEF%20DEF.pdf (дата обращения 15.01.2013).
Demeter L., Csergő A-M., Sándor A. D., Imets I., Vizauer C. T. Natural treasures of the Csík Basin (Depresiunea Cicului) and Csík Mountains (Munţii Ciucului) // Mountain hay meadows: hotspots of biodiversity and traditional culture. 2011. URL: http://www.mountainhaymeadows.eu/online_publication/03-natural-treasures-of-the-csik-basin-and-csik-mountains.html (дата обращения 30.04. 2015).
Demeter L., Kelemen A. Data on the distribution and population size of amphibians in the Csík Mountains (Munţii Ciucului), Eastern Carpathians, Romania // Mountain hay meadows: hotspots of biodiversity and traditional culture. 2011. URL: http://www.mountainhaymeadows.eu/online_publication/06-data-on-the-distribution-and-population-size-of-amphibians-in-the-csik-mountains.html (дата обращения 30.04. 2015).
Dolmen D. Norwegian amphibians and reptiles; current situation 1985 // Studies in Herpetology (Roček Z. ed.). Prague, 1986. P. 743–746.
Dolmen D. Distribution, habitat, ecology and status of the moor frog (Rana arvalis) in Norway // Zeitschrift für feldherpetologie. 2008. Suppl. 13. P. 167–178.
Elmberg J. Ǻkergrodan. En artöversikt samt nya rön om dess utbredning i Nord-och Mellansverige // Fauna och flora. 1978. Vol. 73. № 2. Р. 69–78.
Elmberg J. Apparent lack of territoriality during the breeding season in a boreal population of common frogs Rana temporaria // Herpetol. journ. 1986. Vol. 1. № 2. P. 81–85.
Elmberg J. Factors affecting male yearly mating success in the common frogs Rana temporaria // Behav. ecol. sociobiol. 1991. Vol. 28. P. 125–131.
Elmberg J. Grod- och kräldjurens utbreding i Norrland // Natur i Norr (Umeá). 1995. Ǻrgáng 14, h. 2. P. 57–82.
Elmberg J. Ecology and life history of the moor frog (Rana arvalis) in boreal Sweeden // Zeitschrift für feldherpetologie. 2008. Suppl. 13. P. 179–194.
Elmberg J., Lundberg P. Intraspecific variation in calling, time allocation and energy reserves in breeding male common frogs Rana temporaria // Annales zool. fennici. 1991. Vol. 28. № 1. P. 23–29.
Gasc J.-P., Cabela A., Crnobrnja-Isailovic J, Dolmen D., Grossenbacher K., Haffner P., Lescure J., Martens H., Martinez R., Maurin H., Oliviera M., Sofianidou T., Veith M., Zuiderwijk A. (eds). Atlas of amphibians and reptiles in Europe. Paris, 1997. 516 p.
Gislén T., Kauri H. Zoogeography of the Swedish amphibians and reptiles, with notes on their growth and ecology // Acta vertebrat. 1959. Vol. 1. № 3. P. 191–397.
Glandt D. Rana arvalis Nilsson, 1842 — Moorfrosch // Handbuch der reptilian und amphibien Europas/ Band 5/III A: froschlurche (Anura) III A (Ranidae I). AULA–Verlag, 2014. S. 12–113.
Glasnovič P., Krystufek B., Sovinc A., Bojović M., Porej D. Protected area gap analysis (final report). 2009. 327 p. URL: http://www.discoverdinarides.com/files/file/gap-analysis-final-report-1363265933.pdf
Gleed-Owen C. P. Subfossil records of Rana cf. lessonae, Rana arvalis and Rana cf. dalmatina from Middle Saxon (c. 600-950 AD) deposits in eastern England: evidence for native status // Amphibia – Reptilia. 2000. Vol. 21. P. 57–65.
Haapanen A. Site tenacity of the common frog (Rana temporaria L.) and the moor frog (R. arvalis Nilss.) // Annales zool. fennici. 1970. Vol. 7. № 1. P. 61–66.
Hangartner S., Laurila A., Räsänen K. Adaptive divergence of the moor frog (Rana arvalis) along an acidification gradient // BMC Evolutionary Biology. 2011. Vol. 11. P. 336. URL: http://www.biomedcentral.com/1471-2148/11/366 (дата обращения 13.03.2015).
Hartel T., Öllerer K., Demeter L., Nemes Sz., Moga C., Cogălniceanu D. Using a habitat based approach in mapping amphibian distribution in Romania: study case of the Saxon cultural landscape in the Târnava Mare basin. URL: www.mihaieminescutrust.org/files/studies/Hartel.amphibian.distribution.pdf (дата обращения 23.09.2014).
Heimer W. Amphibienvorkommen im Ostteil des landkreises Darmstadt-Dieburg // Hess. faun. briefe. 1981. Vol. 1. № 2. S. 20–23.
Heráň I. Výsledky výzkumu prostorové aktivity skokana hnédégo, Rana temporaria L., 1758, v Krknosském národnim parku // Sbornik národneho muzea v Praze. 1982. T. 38 B. № 4. S. 239–263.
Ishchenko V. Rana arvalis Nilsson, 1842 // Atlas of amphibians and reptiles in Europe (Gasc J.-P. ed.). Paris, 1997. P. 128–129.
Jablonski D., Jandzik D., Gvoždik V. New records and zoogeographic classification of amphibians and reptiles from Bosnia and Herzegovina // North-western journ. of zool. (Romania). 2012. Vol. 8. № 2. P. 324–337. URL: http://biozoojournals.3x.ro/nwjz/index.html
Jędrzejewska B., Brzeziński M., Jędrzejewski W. Seasonal dynamics and breeding of amphibians in prestine forests (Białowieża National Park, E Poland) in dry years // Folia zool. 2003. Vol. 52. № 1. P. 77–86.
Juszczyk W. Płazy i gady kraiowe. Warszawa, Cześć 2, 1987. 384 s.
Koskela P., Pasanen S. The wintering of the common frog, Rana temporaria L., in northern Finland // Aquilo, ser. zool. 1974. Vol. 15. P. 1–17.
Kutenkov A. P., Panarin A. E. Ecology and status of populations of the common frog (Rana temporaria) and the moor frog (Rana arvalis) in northwestern Russia with notes on their distribution in Fennoscandia // Amphibian populations in the Commonwealth of Independence States: current status and declines. Moscow: Pensoft, 1995. P. 64–70.
Laan R., Verboom B. Effects of pool size and isolation on amphibian communities // Biol. conserv. 1990. Vol. 54. P. 251–262.
Loman J. Macro- and microhabitat distribution in Rana arvalis and R. temporaria (Amphibia, Anura, Ranidae) during summer // Journ. of herpetol. 1978. Vol. 12. № 1. P. 29–33.
Loman J. Annual and daily locomotor activity of the frogs Rana arvalis and R. temporaria // Brit. journ. of herpetol. 1980. Vol. 6. № 3. P. 83–85.
Loman J. Density and survival of Rana arvalis and Rana temporaria // Alytes. 1984. Vol. 3. № 4. P. 125–134.
Loman J., Andersson G. Monitoring brown frogs Rana arvalis and Rana temporaria in 120 south Swedish ponds 1989–2005. Mixed trends in different habitats // Biological conserv. 2007. Vol. 135. P. 46–56.
Loman J., Lardner B. Does pond quality limit frogs Rana arvalis and R. temporaria in agricultural landscapes? A field experiment // Journ. of applied ecol. 2006. P. 690–700.
Miaud C., Guyétant R., Humbert A. Age structure in montane population of common frog (Rana temporaria) // 8th Ordinary general meeting Societas Eur. Herpetol. Bonn, 1995. P. 85–86.
Montanarella L., Jones R. J. A., Hiederer R. The distribution of peatland in Europe // Mires and Peat. 2006. Vol. 1. Article 01, 10 pp. URL: http://www.mires-and-peat.net.
Okulova N. M. Amphibians in northern European Russia // Amphibian populations in the Commonwealth of Independent States: current status and declines. Moscow: Pensoft, 1995. P. 59–63.
Opatrný E. Beitrag zur erkenntnis der verbreitung der amphibienfauna in der Tschechoslowakei // Acta univer. palack. olom. 1978. T. 59. S. 205–220.
Pasanen S., Karhaää M. Can boreal common frog (Rana temporaria L.) survive in frost? // Annales zool. fennici. 1997. Vol. 34. № 4. P. 247–250.
Pasanen S., Sorjonen J. Partial terrestrial wintering in a northern common frog population (Rana temporaria L.) // Annales zool. fennici. 1994. Vol. 31. № 2. P. 275–278.
Pascual X., Montori A. Contribucion al estudo de Rana temporaria L. (Amphibia, Ranidae) en Santa Fe del Monsteny (Barcelona). I. Descripcion de la zona y estima de la poblacion // Miscel. zool. 1981. № 7. P. 109–115.
Petrov B. 4. Amphibians and reptiles of Bulgaria: fauna, vertical distribution, zoogeography, and conservation // Biogeography and ecology of Bulgaria (V. Fet and A. Popov eds.). Springer, 2007. P. 85–107. URL: http://www.nmnhs.com/downloads/pdfs/petrov-boyan/biogeograpgy-ecology-bulgaria-2007-85-107.pdf.
Petrov B., Tzankov N., Strijbosch H., Popgeorgiev G., Beshkov V. The herpetofauna (Amphibia and Reptilia) of the Western Rhodopes mountain (Bulgaria and Greece) // Beron P. (ed.). Biodiversity of Bulgaria. 3. Biodiversity of Western Rhodopes (Bulgaria and Greece). Sofia, 2006. P. 863–912. URL: http://www.lacerta.de/AS/Bibliografie/BIB_1439.pdf
Puky M., Shád P. Distribution and status of the moor frog (Rana arvalis) in Hungary // Zeitschrift für feldherpetologie. 2008. Suppl. 13. P. 309–316.
Puky M., Shád P., Szövéniyi G. Magyarország herpetológiai atlasza. Herpetological atlas of Hungary. Budapest, 2005. 207 p.
Räsänen K., Laurila A., Merilä J. Carry-over effects of embryonic acid conditions on development and growth of Rana temporaria tadpoles // Freshwater biol. 2002. Vol. 47. P. 19–30.
Rittenhouse T., Semlitsch R. Distribution of amphibians in terrestrial habitat surrounding wetlands // Wetlands, 2007. Vol. 27. № 1. P. 153–161. URL: http://www.researchgate.net/ publication/226216986_Distribution_of_amphibians_in_terrestrial_habitat_surrounding_wetlands.
Roček Z., Šandera M. Distribution of Rana arvalis in Europe: a historical perspective // Zeitschrift für feldherpetologie. 2008. Suppl. 13. P. 135–150.
Sas I., Covaciu-Marcov S.-D., Demeter L., Cicort-Lucaciu A.-S., Strugariu A. Distribution and status of the moor frog (Rana arvalis) in Romania // Zeitschrift für feldherpetologie. 2008. Suppl. 13. P. 337–354.
Sas I., Covaciu-Marcov S.-D., Kovács E.-H., Radu N.-R., Tóth A., Popa A. The populations of Rana arvalis Nills. 1842 from the Ier Valley (The Western Plain, Romania): present and future // North-western journ. of zool. (Romania). 2006. Vol. 2. № 1. P. 1–16. http://01.nwjz.2.1.Sas.pdf
Savage R. M. The breeding behaviour of the common frog Rana temporaria Linn., and Bufo bufo Linn. // Proc. zool. soc. London, 1934. Vol. 6. P. 55–70.
Serra-Cobo J., Lacroix G., White S. Comparison between the ecology of the new europaean frog Rana pyrenaica and that of four pyrenean amphibians // Journ. of zool. 1998. Vol. 246. № 2. P. 147–154.
Schmid W. D. Survival of frogs in low temperature // Science. 1982. Vol. 215. № 4533. P. 697–698.
Schweiger M. Rana arvalis near Vienna (видео). URL: http://www.youtube.com/watch?v=bXJMK3ocgFY (дата обращения 19.11.2014).
Strijbosch H. Habitat selection of amphibians during their aquatic phase // Oikos. 1979. Vol. 33. № 3. P. 363–372.
Storey K. B. Persistence of freeze tolerance in terrestrially hibernating frogs after spring emergence // Copeia. 1987. № 3. Р. 720–726.
Stugren B. Geographic variation and distribution of the moor frog, Rana arvalis Nills. // Annales zool. fennici. 1966. Vol. 3. № 1. P. 29–39.
Sztatecsny M., Hödl W. Can protected mountain areas serve as refuges for declining amphibians? Potential threats of climate change and amphibian chytridiomycosis in alpine amphibian population // Ecology of mountains. 2009. Vol. 1. № 2. P. 19–24.
Sztatecsny M., Preininger D., Freudmann A., Matthias-Claudio L., Maier F., Hödl W. Don’t get the blues: conspicuous nuptial coloration of male moor frogs (Rana arvalis) support visual mate recognition during scramble competition in large breeding aggregations // Behav. ecol. sociobiol. 2012. Vol. 66. P. 1587–1593. DOI: 10.1007/s00265-012-1412-6.
Terhivuo J. Provisional atlas and population status of the Finnish amphibian and reptile species with reference to their ranges in northern Europe // Annales zool. fennici. 1981. Vol. 18. № 3. P. 139–164.
Thiesmeier B. Untersuchungen zur Jahresaktivitat der erdkrote (Bufo bufo Linnaeus, 1758) und des grasfrosches (Rana temporaria Linnaeus, 1758) im Niederbergishen land // Decheniana. 1992. Vol. 145. S. 146–164.
Vences M., Grossenbacher K., Puente M., Palanca A., Vieites D. The Cambales fairy tale: elevational limits of Rana temporaria (Amphibia, Ranidae) and other Europaean amphibians revisited // Folia zool. Vol. 2003. 52. № 3. P. 189–202. URL: http://www.ivb.cz/folia/52/2/189-202.pdf.
Vences M., Pique N., Lopez A., Puente M., Miramontes C., Vieites D. R. Summer habitat population estimate and body size variation in a high altitude population of Rana temporaria // Amphibia – Reptilia. 1999. Vol. 20. № 4. P. 426–435.
Vogrin N. The status of Amphibians in Slovenia // Froglog. The newsletter of IUCN DAPTF. 1997. № 20. P. 1–2.
Vos C. C., Chardon J. P. Effects of habitat fragmentation and road density on the distribution pattern of the moor frog Rana arvalis // Journ. of applied ecol. 1998. Vol. 35. P. 44–56.
Vos C. C., Goedhart P., Lammertsma D., Spitzen-Van der Sluijs A. Matrix permeability of agricultural landscapes: an analysis of movements of the common frog (Rana temporaria) // Herpetol. j. 2007. Vol. 17. P. 174–182.
Vukov T., Kalezić M., Tomović L., Krizmanić I., Jović D., Džukić G. Amphibians in Serbia — distribution and diversity patterns // Bull. of the Natural Museum (Serbia). 2013. № 6. P. 90–112. DOI: 10.5937/bnhmb1306090V
Wind-Larsen H., Jørgensen C. B. Hormonal control of seasonal growth in a temperate zone toad Bufo bufo // Acta zool. 1987. Vol. 68. № 1. P. 49–56.
Благодарности
Я искренне благодарен коллегам В. Г. Ищенко, С. М. Ляпкову, D. Dolmen, J. Elmberg, I. Sas, M. Puky за предоставление собственных, а также некоторых малодоступных публикаций по экологии рассматриваемых видов лягушек. Я также весьма признателен рецензентам за замечания и критику в адрес представленной на их суд работы. Их помощь способствовала существенному улучшению текста обзора.

 © 2011 - 2026
© 2011 - 2026