



Введение
Данная работа посвящена некоторым особенностям конденсации воды на поверхности растений. Приводятся аргументы, подтверждающие гипотезу автора (Прохоров, 2012, с. 10), состоящую в том, что особое значение для повышения эффективности и продолжительности конденсации воды имеет тот факт, что температура транспирирующих побегов и листьев растений обычно ниже температуры окружающей среды (Gates, 1968) и, соответственно, зачастую ниже точки росы.
Результаты
Формирование тумана – конденсация водяных паров в воздухе – начинается при достижении точки росы (Td), зависящей от температуры (T) и относительной влажности (φ) воздуха (рис. 1). В условиях тумана, т. е. при T = Td, величина температуры поверхности листьев и побегов TL не существенна. При T > Td конденсация воды осуществляется на поверхностях, охлажденных до Td, в т. ч. на поверхности листьев и побегов растений.
Известно, что температура транспирирующего листа растений понижается (Gates, 1968). Предполагаю, что в случаях когда TL ≤ Td, конденсация воды осуществляется на поверхности растений.
В работе не рассматриваются ситуации, когда TL увеличивается в результате: нагрева поверхности листа на солнце, прекращения или уменьшения транспирации, повышения температуры воздуха, уменьшения относительной влажности воздуха или любого другого фактора. Эти явления, как и вопросы радиационного баланса в целом, не имеют прямого отношения к описываемой ситуации, т. к. активная конденсация воды в этих условиях отсутствует.
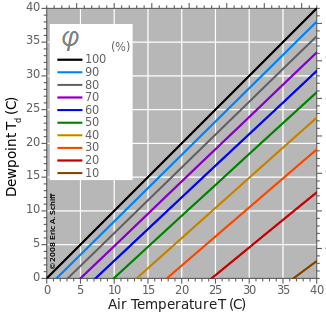
Рис. 1. Зависимость точки росы (Dewpoint, Td, °C) от температуры воздуха (Air Temperature, T, °C) и относительной влажности воздуха (φ, %), (Eric A. Schiff, 2008, цит. по Википедии, «Dewpoint»)
Fig. 1. Dependence of the dewpoint (Td, °C) on air temperature (T, °C) and relative humidity (φ, %), (Eric A. Schiff, 2008, cit. by Wikipedia, "Dewpoint")
Выбор экосистем для рассмотрения применимости предлагаемой гипотезы обусловлен двумя крайними экологическими ситуациями – высокими значениями φ и незначительным перепадом дневных и ночных температур в монтеверде; низкими значениями φ и высоким перепадом дневных и ночных температур в пустынях.
Эти условия создают оптимальные возможности для использования Td для конденсации воды. В первом случае, при незначительном повышении T, растения могут продолжить конденсировать воду из влажного воздуха за счет незначительного понижения TL в тени облаков. Во втором случае резкое снижение T ночью, увеличение φ и начало транспирации у суккулентов позволяет растениям сконденсировать доступную влагу.
Изолинии точки росы (Td), приведенные на рис. 1 для φ ≥ 80 %, характеризуют область муссонных экосистем и горных лесов, например, монтеверде Канарских островов при φ ≤ 20 % соответствуют условиям большинства пустынь в дневное время и при φ ≤ 60 % – в ночное время. Для выпадения росы на поверхности растений в таких условиях требуется снижение TL , ∆TL = ≤ 3 °C в условиях монтеверде и ∆TL ≥ 7 °C – в условиях пустынь в ночное время. Следует при этом учесть, что более общим лимитирующим условием является TL ≥ 0 °C, и ≤ Td. Как следует из приведенной диаграммы на рис. 1, в диапазоне T от 0 °C до 10 °C и при относительной влажности 50–90 % Td ≤ 0 °C, т. е. ниже физиологически оптимального диапазона температур. Следовательно, в арктических и высокогорных экосистемах данный механизм активной конденсации водяных паров может не работать.
Обсуждение
Основу растительных сообществ монтеверде на Канарских островах составляют лавровые леса («лаурисильва») с доминированием нескольких видов из семейства Lauraceae. Выше, в экотонной области монтеверде, произрастает Pinus canariensis C. Sm. Характерной особенностью этих экосистем, расположенных на высоте 800–1300 м над уровнем моря, является высокая влажность воздуха (φ ≥ 75 %) и относительно невысокая стабильная температура 13–16 °C. Данная экосистема расположена в облачном слое, высота которого меняется в зависимости от температуры воздуха (Marzol, 1993).
Конденсация осуществляется за счет механической сорбции микрокапель воды, что имеет особое значение в случае одного из лучших известных «конденсаторов тумана» – P. canariensis. Многочисленные, длинные (20–30 см), ниспадающие иглоподобные листья этой сосны, обитающей на горных склонах Канарских островов, способны сорбировать достаточное количество влаги. Это обеспечивает не только потребность самого растения, но и значительно повышает влажность почвы в монтеверде, что используется в лесном и сельском хозяйстве для выращивания растений, орошаемых с помощью сосны (Pines of Silvicultural Importance, 2002; Crawford, 2008).
В связи с данной темой следует также отметить снижение транспирации древесных растений в условиях тумана, зарегистрированное в условиях монтеверде (Ritter, Regalado, Aschan, 2009). При этом TL должна повышаться, что, впрочем, не препятствует конденсации микрокапель воды на листовой поверхности во время тумана.
Вышеприведенный пример пассивной механической сорбции воды на хвое P. canariensis дается, чтобы показать различия в принципах пассивной и активной конденсации водяных паров.
Объектом нашего внимания является значительно более интересная ситуация, когда температура воздуха несколько выше точки росы и туман не образуется, а следовательно, невозможна и его механическая сорбция на листьях растений. В условиях монтеверде это связано с дневным повышением температуры воздуха и перемещением тумана (облачного слоя) выше области, где произрастает P. canariensis. Следовательно, часть времени сосна проводит при φ менее 100 % (T > Td). В таких условиях транспирация увеличивается, и понижается температура растения. Одновременно, за счет значительной поверхности хвои, такое понижение температуры при TL ≤ Td приводит к активной и эффективной конденсации водяных паров. Разумеется, в условиях Канарских островов, при высокой инсоляции, период активной конденсации водяных паров непродолжителен. Он завершается при TL > Td.
При изучении травянистых растений и орошаемых культур в аридных зонах испанских степей (Lange, Lange, 1963) оказалось, что интенсивность их транспирации иногда достигает 300 г/м2 в час, что в 5–10 раз больше, чем у местных древесных растений. Из-за транспирационного охлаждения температура листьев этих трав по сравнению с температурой нетранспирирующих листьев (смазанных вазелином или отрезанных) оказывается ниже на 5,7–15,7 °С у разных видов, что превосходит величины ∆TL, требуемые для достижения Td на поверхности растения.
Из этих данных, многократно подтвержденных на других видах растений, следует, что за счет снижения TL конденсация воды (выпадение росы) может происходить непосредственно на их поверхности, при условии, что TL ≤ Td.
Косвенным доказательством эволюционного значения данного механизма конденсации воды является форма пустынных кактусов, молочаев и других суккулентов, позволяющая конденсату стекать прямо к корням растений. Однако и этот известный факт обычно связывается с пассивной конденсацией водяных паров – выпадением росы на охлажденной ночью поверхности почвы и растений – не учитывается возможность активной конденсации воды растением за счет снижения температуры поверхности стебля (TL ≤ Td < T) при достаточно низких (ночных) температурах, когда осуществляется транспирация, а снижение температуры ∆TL минимально в условиях пустыни. На значение данного явления может указывать увеличение охлаждаемой поверхности кактусов и молочаев за счет формирования ребер и мамилл, участки между которыми хорошо защищены от избыточной инсоляции.
Заключение
Таким образом, предположение о том, что за счет снижения температуры листьев и побегов растение активно конденсирует влагу из воздуха, увеличивая обычный период выпадения росы, подтверждается сопоставлением значений изменения точки росы в зависимости от влажности воздуха и его температуры со значениями снижения температуры транспирирующих растений. Вечернее выпадение росы на поверхности растений начинается раньше, чем формирование тумана. Утренний этап конденсации продолжается некоторое время после того, как температура воздуха превысит точку росы.
Значение же данного явления для большинства экосистем (если исключить Арктику) может быть крайне велико. В аридных экосистемах – это эволюционное приспособление, обеспечивающее выживание растений. В условиях монтеверде – экологический фактор, обеспечивающий защиту экосистем от опустынивания.
Библиография
Прохоров А. А. Большие Канары, или Амбиции как инструмент эволюции, 2012. 13 с. URL: http://alpro.hortusbotanicus.ru/photos/f4b.pdf.
C. A. B. International. Pines of Silvicultural Importance: Compiled from the Forestry Compendium, CAB International. CABI. 2002. P. 35. URL: http://books.google.com/books?id=DB8dCbmgQ74C&pg=PA35.
Crawford R. M. M. Plants at the Margin: Ecological Limits and Climate Change. Cambridge: Cambridge University Press, 2008, 478 p.
Dewpoint // Wikipedia. URL: http://en.wikipedia.org/wiki/Dew_point (дата обращения: 18.11.2013).
Gates D. M. Transpiration and Leaf Temperature // Annual Review of Plant Physiology. 1968. Vol. 19. P. 211–238.
Lange O. L., Lange R. Untersuchungen uber Blattemperaturen, Transpiration und Hitzeresistenz anPflanzen mediterraner Standorte (Costabrava, Spanien) // Flora. 1963. Vol. 153. P. 387–425.
Marzol M. V. Tipificacion de las tres situaciones atmosfericas mas importantes en las islas Canarias / Strenae Enmanuelae Marrero. Universidad de La Laguna, 1993. P. 80–95.
Ritter A., Regalado C. M., Aschan G. Fog reduces transpiration in tree species of the Canarian relict heath-laurel cloud forest (Garajonay National Park, Spain) // Tree Physiol. 2009. Vol. 29 (4). P. 517-28. Epub 2009 Jan 20.
Благодарности
Работа выполнена при поддержке Программы стратегического развития ПетрГУ и по заданию Министерства образования и науки РФ 4.1700.2011.

 © 2011 - 2026
© 2011 - 2026