



Введение
Общественные насекомые, типичным представителем которых является Apis mellifera L. (медоносная пчела), занимают важное положение в наземных экосистемах (Брайен, 1986; Кипятков, 1991). Ареал медоносной пчелы очень широк, что обусловило возникновение на всём его протяжении ряда подвидов (географических рас), адаптированных к местным условиям, которые с давних пор были окультурены человеком и подвергались им искусственному отбору (Ruttner, 1978; Ильясов, 2006; Руттнер, 2006; De la Rúa et al., 2009). Вследствие этого в пчеловодстве по отношению к расам пчёл устоялся термин «порода». Несмотря на то что пчёлы окультурены, они функционируют как элементы экосистем, опыляя не только монокультуры, но и массу цветковых растений естественных фитоценозов. Подвергаясь со стороны человека значительному влиянию, медоносная пчела остаётся важным природным звеном. Поэтому для неё, как и для других представителей общественных насекомых, актуальна проблема изучения популяционной структуры вида и механизмов ее становления и поддержания, являющаяся центральной как для теоретических исследований в области синтеза эволюционной теории и экологии, так и для практики сохранения биоразнообразия и организации управления хозяйственно значимыми видами (Биглова, 2013; Stamatopoulou et al., 2008).
Перемещение медоносных пчел (Apis mllifera L.) из одних биоклиматических зон в другие с целью повышения продуктивности провоцировало бессистемную гибридизацию и, как следствие, метизацию. В результате на месте естественно сложившихся местных популяций сформировались помеси неизвестного происхождения. Поэтому поиск методов изучения популяций медоносной пчелы, подвергшейся вмешательству хозяйственной деятельности человека, является необходимым для сохранения самовозобновляемой системы популяций этого вида и требует исследования потенциала медоносной пчелы на многих уровнях: внутривидовом, популяционном и внутрипопуляционном (Гайнутдинова, 2004; Биглова, 2013). Как известно, в отличие от организмов, ведущих одиночный образ жизни, структурным элементом (единицей) популяций общественных насекомых (муравьи, термиты, осы, шмели и пчелы) является семья, или колония (Алпатов, 1948; Русина и др., 2004). Этот специфический вид групповой изменчивости животных, образующих сообщества — семьи (произошедших от одной самки), В. В. Алпатов (1948) назвал семейной изменчивостью
Классический метод различения внутривидовых таксонов медоносной пчелы — морфометрический анализ, был развит В. В. Алпатовым (1948). Для характеристики внутритаксономических групп (рас или пород) A. mellifera обычно используют ширину и длину 3-го тергита, длину правого переднего крыла, ширину правого переднего крыла, кубитальный индекс, дискоидальное смещение, длину хоботка — показатели, в основном отражающие специфику развития хозяйственно-полезных признаков (Николаенко, 2005; Симанков и др., 2012; Shaibi et al., 2009; Radloff et al., 2010). На протяжении большого периода в рамках традиционного морфолого-географического подхода большинство исследователей применяли методы одномерной статистики, имеющие ограниченную различительную способность. Более впечатляющие результаты дало использование многомерного статистического анализа морфометрических признаков вида A. mellifera, позволившего подразделить вид на четыре эволюционные ветви (Ruttner, 1978, Ruttner, 1988), объединяющие 26 подвидов (Ruttner, 1992; Sheppard et al., 1997; Sheppard, Meixner, 2003; Meixner et al., 2010), а по некоторым данным (Engel, 1999), 28 подвидов.
Изучая межсемейные различия отдельных популяций медоносных пчёл, можно выявить признаки, наиболее и наименее подверженные изменчивости, и тем самым лучше понять механизм приспособительных реакций организма в условиях изменения окружающей среды.
Вопрос, касающийся внутрипородных межсемейных различий медоносных пчёл по комплексу морфологических показателей, недостаточно освещён в литературе. С учетом вышесказанного, в настоящей работе предпринята попытка с помощью непараметрического дискриминантного анализа, служащего для различения и распознавания биологических объектов и явлений, отличия между которыми не очевидны, в том числе для установления групповой принадлежности отдельных особей (Ивантер, Коросов, 2011), дать оценку вклада стандартного набора экстерьерных признаков рабочих особей медоносной пчелы, относящихся к породам Аpis mellifera carnica Pollmann, 1879 и Apis mellifera caucasica Gorbatchev, 1916, во внутрипородные межсемейные и межпородные различия вида Apis mellifera L.
Материалы
Пчёлы породы A. m. carnica были собраны в 2011 г. на племенных пасеках, принадлежащих ФГУ ПППХ «Майкопское». Пчелы породы A. m. caucasica были получены в этом же году на пасеках ГНУ «Краснополянская опытная станция пчеловодства» НИИ Пчеловодства РСХА. Обучающие выборки каждой породы были представлены 240 особями рабочих пчёл, относящихся к 8 семьям (по 30 особей рабочих пчел из каждой семьи). В выборку породы A. m. carnica входили пчелы из семей № 1—8. В выборку породы A. m. caucasica — из семей № 9—16.
Методы
Анализ экстерьерных признаков проводили по шести основным морфометрическим признакам рабочих пчёл: ширина 3-го тергита (Ш3Т); длина 3-го тергита (Д3Т), длина правого переднего крыла (ДППК), ширина правого переднего крыла (ШППК), кубитальный индекс (Ci), дискоидальное смещение: положительное дискоидальное смещение (ПДС), нейтральное дискоидальное смещение (НДС), отрицательное дискоидальное смещение (ОДС).
Все промеры экстерьерных признаков выполнялись по стандартной схеме, изображённой на рисунке. Измерения экстерьерных признаков производились с помощью стереоскопического микроскопа МБС-9, при этом пользовались окуляром 8x с диоптрийной наводкой со сменной шкалой. В ходе проверки на нормальность распределения исходных данных экстерьерных признаков, проведённой с помощью критериев Колмогорова – Смирнова, Лиллиефорса и Шапиро – Уилка, выяснилось, что среди большинства семей изучаемых пород нормальному распределению подчинялись только «кубитальный индекс» и «ширина 3-го тергита». Ввиду того что одни и те же нормально распределённые признаки встречались не во всех используемых в анализе выборках, мы воспользовались непараметрическим дискриминантным анализом. Для его проведения был применён модуль GDA (Общие модели дискриминантного анализа) программы STATISTICA 10.0.1, позволяющий, в том числе, использовать в анализе категориальные переменные (Халафян, 2007).
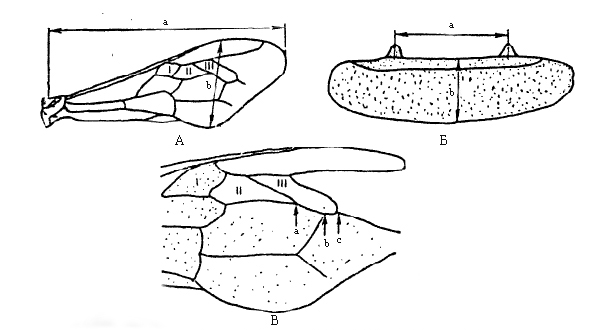
Рис. Экстерьерные признаки рабочих особей медоносной пчелы: А – промеры переднего правого крыла: a – длина, b – ширина; Б – промеры 3-го тергита: a – условная ширина, b – длина; В – переднее правое крыло с тремя кубитальными ячейками I, II, III (соотношение отрезков третьей кубитальной ячейки: bc/ab х 100% – даёт кубитальный индекс). По Николаенко (2005)
Fig. Exterior features of honeybee working individuals : A – measurements of right front wing: a – length, b – width; Б - measurements of third tergite: a – conditional width, b – length; В – right front wing with three cubital cells I, II, III (the ratio of the segments of third cubital cell: bc/ab х 100%. – gives cubital index)
Была проведена верификация полученных уравнений функции классификации (Клекка, 1989) на основе двух экзаменационных выборок из каждой породы (n = 20), состоящих из пчёл, ранее не использовавшихся для анализа в составе обучающих выборок. Результаты верификации оценивали по количеству верно классифицированных объектов по отношению к их общему числу, выраженному в процентах. Полученные результаты сравнивали с аналогичными значениями, выдаваемыми модулем GDA в виде таблицы «Матрица классификации для анализируемой выборки», составленной по обучающим выборкам.
Результаты
Внутрипородные межсемейные различия экстерьерных признаков рабочих пчел пород A. m. carnica и A. m. caucasica
Результаты анализа межсемейных различий по шести экстерьерным признакам рабочих пчел, относящихся к породам A. m. carnica и A. m. caucasica приведены в табл. 1. Анализ экстерьерных признаков рабочих пчёл породы A. m. carnica показал, что между семьями наблюдалась статистически значимая дискриминация по всем экстерьерным признакам. Семьи больше всего различались по параметрам «длина крыла» и «ширина крыла», при этом наибольшей разделительной силой между семьями обладал признак «длина крыла», т. к. ему соответствовало наименьшее значение частной лямбды.
Непараметрический дискриминантный анализ межсемейных различий пчел породы A. m. caucasica свидетельствует, что семьи этой породы также имели статистически значимые различия по исходному набору экстерьерных признаков. Но в отличие от A. m. carnica для семей породы A. m. caucasica наибольшей дискриминирующей силой обладал признак «дискоидальное смещение».
Таблица 1. Внутрипородные межсемейные различия по экстерьерным признакам рабочих пчел пород A. m. carnica и A. m. caucasica
Анализ квадратов расстояний Махаланобиса между центроидами классов объектов, образованных семьями породы A. m. carnica, показал, что статистически незначимые различия в расстояниях (p > 0.05) наблюдались между центроидами классов семей в следующих парах: № 1 и № 2; № 1 и № 6; № 4 и № 3. Следовательно, для этих пяти семей в пространстве 6-ти экстрерьерных признаков имеет место сильное перекрывание значений признаков, что приводит к слабой дискриминации, причём в наименьшей степени отличалась от остальных семья № 6. Для породы A. m. caucasica статистически незначимые различия (p > 0.05) в значениях квадратов расстояний Махаланобиса были обнаружены между семьями № 10 и № 9, а также № 14 и № 16.
Анализ beta-коэффициентов, свидетельствующих о вкладе каждого из имеющихся экстерьерных признаков в дискриминацию семей каждой породы, показал, что более половины семей породы A. m. carnica статистически значимо различались (p < 0.05) по признакам «длина крыла» (7 семей) и «длина 3-го тергита» (6 семей). Для породы A. m. caucasica ключевыми признаками, по которым статистически значимо различались семьи, были «ширина крыла» (5 семей) и «длина крыла» (4 семьи).
На следующем этапе работы был проведён дискриминантный анализ межпородных различий рабочих пчел в многомерном пространстве экстерьерных признаков.
Межпородные различия экстерьерных признаков пород A. m. carnica и A. m. caucasica
Чтобы выяснить силу различий между анализируемыми породами, в качестве группирующей переменной в модуле GDA была задана принадлежность к породе. По результатам анализа выяснилось, что породы не различались по признаку «длина 3-го тергита» (табл. 2), по всем остальным признакам наблюдались статистически значимые различия. Наиболее существенный вклад в разделение пород вносил «кубитальный индекс», что оправдывает его общеизвестную первостепенную важность в породной идентификации медоносных пчёл (Алпатов, 1948; Руттнер, 2006).
Таблица 2. Межпородные различия по экстерьерным признакам рабочих пчел пород А. m. carnica и A. m. caucasica
Оценка расстояния между центроидами классов объектов (пород) с помощью квадрата расстояния Махаланобиса показала их статистическую значимость (p < 0.05). Это свидетельствует о том, что в пространстве значений шести экстерьерных признаков их совокупное влияние дискриминирует центроиды изучаемых совокупностей на статистически значимое расстояние друг от друга.
Анализ знаков перед статистически значимыми betа-коэффициентами позволил сделать вывод о характере различий между породами по каждому из шести признаков. Так, порода А. m. carnica отличалась меньшими значениями признаков «длина крыла» и «кубитальный индекс» от соответствующих значений признаков породы A. m. caucasica. В свою очередь, последняя имела статистически значимо меньшие значения параметров «ширина 3-го тергита» и «ширина крыла» по сравнению с породой А. m. carnica.
С помощью модуля GDA для переменных (экстерьерных признаков) обеих пород были получены коэффициенты и соответствующие уравнения функции классификации (Y), способные идентифицировать породу пчелы по набору экстерьерных признаков. Градацию «отрицательное дискоидальное смещение» программа удалила из моделей (1) и (2) для минимизации доли ошибочной классификации. При построении уравнения функции классификации признак «длина 3-го тергита» был исключён вследствие того, что его вклад в дифференциацию пород оказался статистически незначим (табл. 2).
Уравнения имеют следующий вид:


Полученные уравнения позволили провести верификацию функций классификации, которая была проведена на основе двух экзаменационных выборок каждой породы (n = 20). Критерием отнесения наблюдения (т. е. пчелы) из экзаменационной выборки к той или иной породе является максимальное значение апостериорной вероятности классификации, рассчитанной в модуле GDA на основе квадратов расстояний Махаланобиса. В табл. 3 представлены результаты сравнения классификации наблюдений обучающих и экзаменационных выборок, полученных c помощью модуля GDA и уравнений функции классификации, свидетельствующие, что количество правильно классифицированных объектов в случае применения модуля GDA и уравнений функций классификации практически одинаково.
Таблица 3. Количество верно классифицированных объектов в обучающих и экзаменационных выборках рабочих пчел пород А. m. carnica и A. m. caucasica
Обсуждение
Увеличение числа регистрируемых признаков требует усложнения методов статистической обработки данных и перехода от одномерных методов (описательной статистики) к многомерному анализу, где главной характеристикой объектов становится расстояние между ними в многомерном пространстве изучаемых признаков. Так, в нашей предыдущей работе (Брагазин и др., 2012) с помощью критерия Крускала — Уоллиса были установлены статистически значимые внутрипородные межсемейные различия по каждому отдельному экстерьерному признаку у рабочих особей медоносной пчелы пород А. m. carnica и A. m. caucasica. Однако вопрос о совокупном вкладе комплекса экстрерьерных признаков во внутрипородные, межсемейные и межпородные различия оставался открытым. В настоящей работе применение дискриминантного анализа комплекса экстерьерных признаков позволило получить новые данные, которые дополнили и расширили имеющиеся представления об объекте — медоносной пчеле, а также явились приближением к целостному описанию объекта. Так, применяемый в данной работе для дискриминации разноуровневых внутривидовых группировок набор морфологических признаков демонстрирует различную эффективность, т. е. для внутрипородной дифференциации семей большее значение имеют одни признаки («длина крыла», «дискоидальное смещение»), а для межпородной — другие («кубитальный индекс»). Таким образом, рассматривая подвидовую структуру A. mellifera L., можно отметить следующие особенности. На уровне пород в большей, по сравнению с другими признаками, степени варьируют значения кубитального индекса, что применяется для породной идентификации пчёл. На внутрипородном, т. е. межсемейном, уровне кубитальный индекс отходит на второй план и наибольшей дискриминирующей способностью характеризуются размерные признаки крыла и форма дискоидальной ячейки. Но как показал анализ beta-коэффициентов, общим для обеих пород признаком, статистически значимо разделяющим, по крайней мере, половину семей, является «длина крыла», что имеет значение для анализа групповой (семейной) изменчивости экстерьерных признаков.
Заключение
Полученные результаты дают возможность сделать два существенных вывода. Во-первых, показано, что для обеих пород характерна внутрипородная морфологическая гетерогенность по комплексу экстерьерных признаков, но при этом надежная дискриминация не менее половины семей в каждой породе может быть осуществлена по признаку «длина крыла». Во-вторых, были успешно верифицированы уравнения функции классификации, способные на основе комплекса анализируемых признаков экстерьера идентифицировать породы А. m. carnica и A. m. caucasica. Следовательно, получен инструментарий анализа принадлежности рабочих особей медоносных пчёл к этим породам, который может быть востребован как в пчеловодстве, так и в демэкологических исследованиях общественных насекомых.
Библиография
Алпатов В. В. Породы медоносной пчелы. М.: Изд-во Московского общества испытателей природы, 1948. 183 с.
Биглова Л. Ф. Разнообразие морфотипов медоносной пчелы в популяции лесостепной природно-сельскохозяйственной зоны республики Башкортостан // Современные проблемы науки и образования. 2013. № 2. URL: www.science-education.ru/108-8908 (дата обращения: 23.07.2013).
Брагазин А. А., Радаев А. А., Нижегородцев А. А., Гелашвили Д. Б. Статистический анализ экстерьерных признаков рабочих особей пород медоносной пчелы Apis mellifera carnica Pollmann и Apis mellifera caucasica Gorbatchev // Вестн. Нижегород. ун-та. 2012. № 2 (3). С. 119–122.
Брайен М. Общественные насекомые: экология и поведение / Пер. с англ. М.: Мир, 1986. 400 с.
Гайнутдинова Л. М. Исследование экстерьерных признаков и популяционно-генетической структуры медоносной пчелы (Аpis mellifera L.) на Южном Урале. Автореф. дисс. … канд. биол. наук. Уфа, 2004. 24 с.
Ивантер Э. В., Коросов А. В. Введение в количественную биологию: Учеб. пособие. Петрозаводск: изд-во ПетрГУ, 2011. 302 с.
Ильясов Р. А. Полиморфизм Apis mellifera mellifera L. на Урале. Автореф. дисс. … к.б.н. Уфа, 2006. 22 с.
Кипятков В. Е. Мир общественных насекомых. Л.: Изд-во Ленинградского университета, 1991. 408 с.
Клекка У. Р. Дискpиминантный анализ // Фактоpный, дискpиминантный и кластеpный анализ / Пер. с англ. М.: Финансы и статистика, 1989. С. 78–138.
Николаенко В. П. Племенная работа с пчёлами. Ростов н/Д: Изд-во БАРО-ПРЕСС, 2005. 144 с.
Русина Л. Ю., Скороход О. В., Гилев А. В. Дискретные вариации окраски осы Polistes dominulus (Christ) (Hymenoptera: Vespidae) в Черноморском биосферном заповеднике // Труды Русского энтомологического общества. 2004. Т. 75. №1. С. 270–277.
Руттнер Ф. Техника разведения и селекционный отбор пчел: Практическое руководство… / Пер. с нем. 7-е изд., перераб. М.: ACT Астрель, 2006. 166 с.
Симанков М. К., Макаров В. Л., Симанков В. М., Ильясов Р. А., Поскряков А. В., Николенко А. Г. Морфогенетическая характеристика медоносной пчелы Пермского края // Материалы третьего международного форума пчеловодов «Медовый мир». Ярославль. 2012. С. 55–58.
Халафян А. А. STATISTICА 6. Статистический анализ данных: Учебник. 3–е изд. М: ООО «Бином-Пресс», 2007. 512 с.
De la Rúa P., Jaffé R., Dall’Olio R., Muñoz I., Serrano J. Biodiversity, conservation and current threats to European honeybees // Apidologie. 2009. V. 40. № 3. P. 263—284. DOI: 10.1051/apido/2009027.

 © 2011 - 2026
© 2011 - 2026