



Введение
Задачей данной работы будет обзор данных по биологии кумжи водоемов крайнего северо-востока Европы (эта территория административно относится к Архангельской области). К сожалению, в недавней зарубежной монографии (Jonsson, Jonsson, 2011) российские популяции кумжи не рассматриваются. Обзор (Махров, 1999) устарел, а разделы, посвященные кумже, в монографиях (Козьмин, 2011; Биологические ресурсы …, 2012) очень кратки.
Между тем описываемые популяции находятся на краю ареала вида и имеют ряд особенностей. Как показывают исследования, выполненные на других видах, окраинные популяции могут в значительной степени отличаться от других популяций и становиться родоначальниками новых форм (обзоры: Kawecki, 2008; Hardie, Hutchings, 2010; Ивантер, 2012). Кроме того, популяции с окраины ареала интересны как потенциальный источник заселения новых территорий.
Аналитический обзор
Ареал кумжи
Ареал кумжи очень обширен. Она населяет практически всю зарубежную Европу, включая Британские острова и Исландию, бассейны Черного и Каспийского морей, Малую Азию, часть бассейна Аральского моря, ее популяции есть в горах Северной Африки. По европейскому Северу России проходит северо-восточная граница ареала этого вида (обзор: MacCrimmon, Marshall, 1968).
Кумжа встречается в низовьях реки Онеги (Якобсон, 1913; Кудерский, Мельникова, 1983) и ее притоке Коже (Новоселов, 2006). Она есть в реках Палой (Алеев, 1913), Тамице, Вейге, Лямце (Новоселов, 1992) на восточном побережье Онежского залива, по опросным данным обитает в ряде других рек и ручьев этого побережья, в частности в реке Котовой (рис. 1).
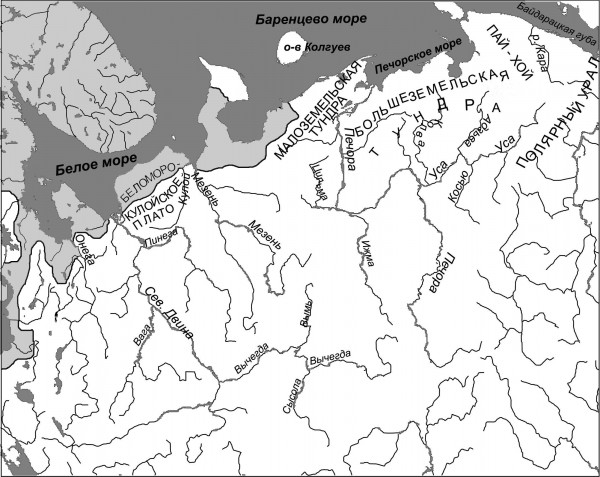
Рис. 1. Северо-восточная часть ареала кумжи
Fig. 1. Northeastern part of the area of brown trout
Встречается или встречалась кумжа и во многих водоемах Летнего берега Белого моря: реке Лопшеньге, ручьях Голодеиха 1-й и 2-й, речке Сярте, ручье Лавката, реках Яреньге, Большой и Малой Режме, Сюзьме; в последней реке к 1930 гг. исчезла (Плетцов, 1936). Кумжа есть и в нескольких реках, впадающих в Унскую губу, – Кумжевой, Вежме, Кинжуге, Луде, Бабьей (данные рыбинспектора Г. В. Севастьяновой).
Кумжа обитает в реках Мудьюге (Кулида, Тимофеев, 2005), Малой Торожме (Махров, Иешко, 2001), Зимней Золотице (Новоселов и др., 2001) и, по данным В. П. Дереца, в других реках и ручьях Зимнего берега Белого моря. Она есть и в реках Мезенского залива – Чиже, Мгле, встречается, хотя и редко, на западном побережье полуострова Канин – указана для реки Шойна (Житков, 1904; Андросова и др., 1934). На севере этого полуострова кумжа не обнаружена (Novikov et al., 2000), но есть восточнее – в реках Чешской губы Баренцева моря – Пеше, Прищатиннице, Волонге, Великой, Шубной, Черной и Индиге (Танфильев, 1896; Бурков, Гошева, 1968).
Восточную границу ареала кумжи определить сложно, поскольку ихтиофауна пресных водоемов восточного побережья Баренцева моря изучена недостаточно. Местное население часто называет кумжей арктического гольца, поэтому в данном случае опросные данные могут ввести в заблуждение (местные жители рассказывали мне о кумже в Коровинской губе в устье Печоры и даже на Таймыре).
Видимо, именно арктического гольца принимали за кумжу на Печоре (Якобсон, 1916; Кузнецов, 1951), Новой Земле (Вылко, 1981) и острове Колгуев (Путешествия …, 1805; Максимов, 1890). Однако два независимых правильных описания кумжи рыбаками, имеющиеся для Югорского шара (Пробатов, 1934) и впадающей в него реки Ой-яга (Друккер, 1927) позволяют считать, что в этом регионе обитает или обитала популяция кумжи.
Кумжа отсутствует в бассейнах крупных рек северо-востока Европы – Северной Двины, Мезени, Печоры (Мартынов, 2007). Отдельные экземпляры кумжи отлавливались в низовьях Северной Двины и Печоры (Кучина, 1941; Новиков, 1964; Корнилова, 1970) – возможно, это особи, заходившие в реки на зимовку. Известен случай поимки кумжи в Печорском море (Семушин и др., 2011). Данные о кумже в притоке Северной Двины, Пинеге (Смирнов, 1935), видимо, ошибочны.
Есть сведения, хотя недостаточно четкие, о вселении кумжи в пресные воды Большого Соловецкого острова монахами (Гульельми, 1888; Анухина, 1972). Эти сведения подтверждаются тем, что в обстоятельном описании островов, подготовленном в первой половине XIX века архимандритом Соловецкого монастыря Досифеем (1836), кумжа еще не упоминается. В XX и XXI веках кумжа на Соловках отмечена, хотя встречается она там редко (Захваткин, 1927; Анухина, 1972; Дворянкин, Новоселов, 2005; опросные данные 2012 года).
Систематическое положение и происхождение кумжи северо-восточного края ареала
Кумжа, живущая в море, и кумжа, живущая в реке (форель) настолько различаются по внешнему виду, что многие систематики, включая К. Линнея, рассматривали пресноводные формы кумжи как особые виды. Однако более 200 лет назад российский академик И. И. Лепехин, описывая кумжу Волонги, проницательно отметил (Путешествия …, 1805, с. 121): «я особливою от форели породою ее не считаю».
Многочисленные исследования, начало которым положила экспедиция И. И. Лепехина, полностью подтвердили его правоту. В частности, на водоемах Карельского берега Белого моря проведена серия работ К. В. Кузищина и Г. Г. Новикова. В ходе этих исследований отмечен совместный нерест жилых и проходных рыб (Кузищин, Новиков, 2001), прослежены изменения пластических признаков – размеров челюстей, головы, плавников, формы тела при смолтификации, то есть подготовке молоди к выходу в море (Кузищин, Новиков, 1994; Кузищин, 1997).
Кумжа образует целый ряд подвидов, популяции севера Европы объединяются в подвид Salmo trutta trutta. По ряду остеологических признаков кумжа бассейна Белого моря значительно отличается от балтийских популяций этого подвида и, возможно, заслуживает выделения в отдельный подвид (Salmanov, Dorofeyeva, 2001; Дорофеева, Салманов, 2007; Дорофеева, 2010).
Однако каких-либо качественных генетических отличий кумжи бассейна Белого моря от других северо-европейских популяций не отмечено, хотя популяции этого бассейна выделяются высоким уровнем разнообразия по генам, кодирующим белки (Осинов, 1984). Это позволило выдвинуть предположение, что регион заселялся кумжей по меньшей мере из двух ледниковых рефугиумов (Осинов, Берначе, 1996).
Поскольку в популяциях кумжи севера Европы обнаружены только гаплотипы митохондриальной ДНК (мтДНК), отсутствующие в популяциях бассейнов Черного и Каспийского морей (Bernatchez, Osinov, 1995; Осинов, Берначе, 1996; Bernatchez, 2001), очевидно, что оба рефугиума располагались севернее этих бассейнов.
Изучение распространения аллелей генов, кодирующих фермент аспартатаминотрансферазу (sAAT-1,2*), в популяциях кумжи севера Европы, позволило предположить, что один из рефугиумов находился в бассейне Балтики, откуда кумжа проникла в южную часть бассейна Белого моря. Другой рефугиум находился на западе Европы (возможно, на Британских островах). Кумжа из этого рефугиума, расселяясь вдоль побережья современной Норвегии и Кольского полуострова, достигла северной части бассейна Белого моря (Махров, Иешко, 2001; Makhrov et al., 2002).
Расселение из Норвегии подтверждается значительным сходством вариантов внутреннего транскрибируемого спейсера (ITS) генов, кодирующих рибосомальную РНК, у кумжи популяций Норвегии, севера Кольского полуострова и Кандалакшского залива Белого моря (Presa et al., 2002).
Высказанным выше представлениям о расселении кумжи не противоречат и данные о разнообразии митохондриальной ДНК популяций бассейнов Белого и Баренцева морей (Bernatchez, Osinov, 1995; Bernatchez, 2001; Lumme et al., 2005). Однако распределение гаплотипов митохондриальной ДНК в популяциях кумжи этого региона мозаично, что затрудняет использование этого маркера для изучения путей расселения.
Экологическая морфология кумжи
В литературе нет данных о морфологии кумжи рассматриваемого нами региона. В таблицах 1 и 2 мы приводим собственные данные по молоди кумжи рек Унской губы, выращиваемой на Онежском рыбоводном заводе (27 экз.). У этих рыб не определяли высоту спинного плавника и число лучей в этом плавнике, поскольку у многих особей он был поврежден из-за некроза. Кроме традиционных признаков (Правдин, 1966), измерялась толщина тела (хозяйственно-важный признак) и выемка хвостового плавника (признак, важный для систематики).
Как видно из табл. 2, форма тела молоди кумжи идеально соответствует ее образу жизни. Брусковатое, мощное тело с высоким хвостовым стеблем обеспечивает броски из убежища за добычей в быстрой воде (Никольский, 1891). Большие голова и рот позволяют захватывать относительно крупные пищевые объекты. Малое число жаберных тычинок также связано с относительно большими размерами добычи.
Показательны результаты сравнения «заводской» молоди кумжи и дикой молоди кумжи рек и ручьев Карельского и Терского берегов Белого моря (Евсин, 1980; Салманов, 1991; Кузищин и др., 1998). Отличий по большинству признаков не выявлено, но кумжа с Онежского рыбоводного завода имела меньшие значения длины и высоты головы, высоты анального плавника, длин грудного и брюшного плавников, наибольшей высоты тела. Видимо, это связано не с межпопуляционными различиями, а с меньшей подвижностью «заводской» рыбы; аналогичные особенности отмечены у «заводской» молоди черноморской кумжи (Артамонова и др., 2006).
Кроме того, у кумжи Онежского рыбоводного завода больше размер глаз, чем у дикой кумжи; это связано, видимо, с постоянным затенением в цеху, где содержалась эта рыба. Разница в размерах верхней челюсти между сравниваемыми выборками объясняется разной методикой измерения.
Таблица 1. Длина тела, масса и меристические признаки молоди кумжи
| Признак | Минимальное и максимальное значения | Среднее значение и ошибка средней |
| Длина всего тела (АВ), мм | 121–187 | 145.7±3.2 |
| Длина тела до конца средних лучей хвостового плавника (АС), мм | 119–184 | 143.0±3.1 |
| Длина тела до конца чешуйного покрова (АD), мм | 111–171 | 132.5±2.9 |
| Длина тушки (OD), мм | 84–136 | 103.2±2.5 |
| Масса, г | 15.6–52.9 | 28.4±1.8 |
| Число пятен на жаберной крышке | 2–5 | 3.2±0.1 |
| Число жаберных тычинок на первой жаберной дуге | 13–19 | 15.9±0.3 |
| Число мягких лучей в анальном плавнике | 7–9 | 8.0±0.1 |
| Число позвонков | 58–61 | 59.1±0.1 |
Таблица 2. Пластические признаки молоди кумжи (в % длины AС)
| Признак | Минимальное и максимальное значения | Среднее значение и ошибка средней |
| Антедорсальное расстояние | 40.0–43.6 | 42.18±0.17 |
| Постдорсальное расстояние | 39.1–42.0 | 40.84±0.14 |
| Антевентральное расстояние | 40.8–51.2 | 48.89±0.50 |
| Антеанальное расстояние | 66.4–68.5 | 67.29±0.10 |
| Пектровентральное расстояние | 27.5–31.0 | 29.63±0.17 |
| Вентроанальное расстояние | 17.8–21.1 | 19.31±0.16 |
| Длина хвостового стебля | 17.0–19.6 | 18.53±0.13 |
| Наибольшая высота тела | 17.0–21.6 | 18.80±0.23 |
| Наименьшая высота тела | 7.7–10.9 | 8.70±0.15 |
| Толщина тела | 10.3–13.2 | 11.44±0.14 |
| Длина головы | 18.9–22.9 | 20.81±0.19 |
| Длина основания спинного плавника | 9.1–12.3 | 11.23±0.15 |
| Длина основания анального плавника | 7.1–9.4 | 7.87±0.11 |
| Высота анального плавника | 9.0–11.2 | 10.23±0.11 |
| Длина грудного плавника | 8.2–12.8 | 10.28±0.22 |
| Длина брюшного плавника | 8.0–10.9 | 9.79±0.14 |
| Длина среднего луча хвостового плавника | 6.9–8.8 | 7.79±0.09 |
| Выемка хвостового плавника | 1.1–2.3 | 1.64±0.06 |
| Высота головы у затылка | 12.0–15.2 | 13.67±0.15 |
| Заглазничное расстояние | 9.9–14.3 | 11.31±0.21 |
| Длина рыла | 5.4–6.9 | 6.07±0.07 |
| Длина верхней челюсти | 10.0–12.5 | 11.30±0.12 |
| Длина нижней челюсти | 11.7–14.0 | 13.15±0.11 |
| Горизонтальный диаметр глаза | 5.2–7.1 | 6.02±0.09 |
Молодь кумжи отличается поразительным разнообразием окраски. Изменяются сразу несколько элементов, слагающих этот признак. Для кумжи характерно наличие небольших черных пятен на всем теле, но их число может быть различным; иногда пятна окружены светлым ободком. У молодых особей и созревающих жилых самцов есть крупные, овальные, располагающиеся через одинаковые промежутки по бокам тела темные пятна (так называемые «пестряточные»); эти пятна вытянуты сверху вниз и издали кажутся полосами. У диких особей часто имеются многочисленные мелкие красные пятна, но на рыбоводных заводах эти пятна почти всегда отсутствуют. В зависимости от освещенности и цвета дна фон окраски меняется от темного-коричневого до белого; наиболее обычны зеленоватый, стальной и оливковый фон.
В ходе смолтификации (то есть подготовки к выходу в море) кумжа приобретает серебристую окраску, более прогонистую форму тела и теряет «пестряточные» пятна и мелкие красные крапины. В дальнейшем, по мере роста, окраска меняется мало, но перед нерестом кожа производителей вновь темнеет, у крупных самцов на нижней челюсти появляется небольшой «крюк».
Ряд морфологических признаков отличает кумжу от близкого вида – семги, или атлантического лосося (Salmo salar L.). В частности, у кумжи больше темных пятен на жаберной крышке, выше хвостовой стебель, меньше выемка хвостового плавника (Jones, 1947). Однако все эти признаки достаточно пластичны, особенно у молоди. Например, у четырех изученных нами особей на жаберной крышке было два пятна, хотя обычно у кумжи три или более таких пятен. Видимо, этот признак в значительной степени зависит от условий среды обитания, как ранее показано для «заводской» молоди семги (Артамонова, Махров, 2000).
Структура популяций
Как упоминалось выше, кроме проходной (нагуливающейся в море) формы, кумжа образует эстуарную форму и ряд жилых (нагуливающихся в пресной воде) форм, часто называемых «форели». Среди жилой формы преобладают самцы, среди проходной – самки (Кулида, Тимофеев, 2005; данные автора), то есть эти две формы в каждой реке входят в состав единой популяции.
Как ранее отмечено при сравнении кумжи разных морских бассейнов В. Н. Евсиным (1977), возраст ската в море растет с запада на восток, по мере ухудшения условий обитания. Возраст мигрантов на Карельском берегу – 2–4 года (Мурза, Христофоров, 1984; Кузищин, 1997), в Унской губе и на Терском берегу Кольского полуострова – обычно 3–5 (Суслова, 1969; Евсин, 1977; Мурза, Христофоров, 1981; Ершов, 1985; Кулида, Тимофеев, 2005; наши данные), в реке Поной (Кольский полуостров) – 4–7 (Христофоров, Мурза, 1988).
В реке Вежма скатывающая молодь кумжи имела среднюю длину 19.5 см (двухгодовики), 23.6 см (трехгодовики) и 29.6 см (четырехгодовики). Средняя масса покатников разных возрастных групп была 72.5, 154.7 и 280 грамм, соответственно (Кулида, Тимофеев, 2005).
Особи проходной формы, идущие на нерест, в реке Волонге имеют длину 45–65 см и массу до 3 кг, особи, идущие на зимовку, – длину 30–35 см и массу 350–500 г (Даниленко, 1964). В Кумжевой длина идущих на зимовку самцов, проведших 3 года в реке и одно лето в море, – 27.0–30.5 (в среднем 28.4) см, самок такого же возраста – 25.5–29.0 (в среднем 27.3) см. Старшие рыбы крупнее – длина двух самцов, живших 3 года в реке и два лета в море, – 38.5 и 40.0 см, двух самок, пробывших 4 года в реке и одно лето в море, – 33.0 и 34.5 см (данные автора). В Вежме масса самок, мигрирующих на зимовку, в среднем 720 г, самцов – 680 г (Кулида, Тимофеев, 2005).
Максимальный возраст особей жилой формы из рек Зимнего берега (Мудьюги и Малой Торожмы) – 4+, в Мудьюге средняя длина рыб этой возрастной группы 31 см, масса – 272 г (Кулида, Тимофеев, 2005; данные автора). По нашим данным, полученным в 1990-е годы, жилые самцы кумжи в реках Малой Торожме и Кумжевой созревают в возрасте 4+.
В популяциях кумжи других участков побережья Белого моря жилые самцы созревают раньше, и возрастная структура упрощается. Так, в притоках Поноя самцы созревают в возрасте 3+ (Христофоров, Мурза, 1990), в ручьях Карельского берега – в возрасте 2+ (как исключение – 1+) (Мурза, Христофоров, 1984; Пономарева и др., 2006). Это может свидетельствовать о большем воздействии вылова на эти популяции, чем на кумжу Архангельской области. Генетическая структура различается в популяциях кумжи, обитающих в разных частях побережья Белого моря, что указывает на существование у этой рыбы хоминга. Особенно выделяется популяция реки Кумжевой, впадающей в Унскую губу; возможно, это связано со значительной обособленностью Унской губы от основной акватории Белого моря (Махров, Иешко, 2001; Makhrov et al., 2002).
Жизненный цикл
Молодь кумжи обитает на местах с быстрым течением и наличием укрытий – камней, коряг, ям, упавших в воду деревьев. «Особенно любит места, где стрежень переходит в омуток и образует встречные течения», – очень точно написал Н. Плетцов (1936). Часть молоди остается на всю жизнь в реках и ручьях, пополняя жилую форму, а часть уходит в море. Скат молоди кумжи в море происходит в мае-июне (Кулида, Тимофеев, 2005).
Нагул происходит в морских губах и на мелководьях. Уже в сентябре-октябре кумжа возвращается в реки и ручьи на зимовку (Даниленко, 1964; Кулида, Тимофеев, 2005; наблюдения автора). Эти особи имеют светлую окраску (их народное название – «светлячок»). Зимовка лососевых рыб в море невозможна. Зарубежные эксперименты на кумже выявили, что при 2 °C способность кумжи адаптироваться к морской воде понижена (Thomsen et al., 2007). Отечественные опыты на семге показали, что смертность в соленой воде при температуре ниже 1 °С резко увеличивается, а при температуре близкой к 0 °С все опытные рыбы погибают (Лега и др., 1991).
Судя по наблюдениям на Вежме (Кулида, Тимофеев, 2005), и Кумжевой (наши данные) кумжа созревает не раньше, чем после двух лет нагула в море. В Волонгу созревающая кумжа начинает подниматься со второй половины июля (Даниленко, 1964), в Вежму – во второй половине августа (Кулида, Тимофеев, 2005). Нерест кумжи происходит в сентябре-октябре, ее плодовитость составляет 2–3 тысячи икринок (Кулида, Тимофеев, 2005).
Роль кумжи в экосистемах
Исследование популяции кумжи реки Мудьюги (Кулида, Тимофеев, 2005) показало, что площадь нерестово-выростных угодий кумжи в этой реке – 30 га, а на 100 м2 приходится от 6 до 17 экземпляров молоди кумжи. Таким образом, в этой достаточно типичной для региона реке обитает 18–51 тысяч молодых рыб. К сожалению, это единственное подобное исследование, проведенное в Архангельской области.
Однако можно уверенно утверждать, что в целом ряде ручьев и небольших рек кумжа преобладает по численности и биомассе над другими видами рыб, то есть является «ключевым видом» их экосистем. Реки и ручьи, где доминирует кумжа, называют «кумжевыми» или «форелевыми».
Данные о питании кумжи в водоемах региона в литературе не обнаружены. Судя по данным, полученным при изучении популяций беломорской Карелии и Кольского полуострова, пищевые объекты кумжи очень разнообразны. «В ее пищевой рацион входит практически все живое население водоема, за исключением растительности» (Муравейко и др., 2007, с. 125).
Молодь кумжи, живущая в реках и ручьях, питается в основном беспозвоночными: личинками ручейников, веснянок, мошек, личинками и куколками хирономид, нимфами поденок, клопами, тендипедидами, моллюсками, пиявками, а в летний период - попавшими в воду наземными насекомыми (Солдатов, 1908; З.Н. Маслова, по: Евсин, 1970; Киреева и др., 1986; Задорина, 1988; Лососевые реки ..., 1991; Аверинцев, Прищепа, 1999; Крылова, 2003). По нашим наблюдениям, насекомыми питается и молодь кумжи притока Зимней Золотицы, ручья Бобриха.
Кроме того, среди пищевых объектов молоди кумжи отмечен гольян, Phoxinus phoxinus (Гринюк и др., 1977; Кузьмин, 1984). В небольшом ручье Черном на Карельском берегу пойман покатник кумжи, в желудке которого обнаружен сеголеток этого же вида (Кузищин, Махров, неопубл. данные). Разноразмерная кумжа может питаться икрой семги (Солдатов, 1908; Азбелев, 1960; Ксензов, 1969; Ершов, 1985). Очевидно, что это икра, которая не попала в нерестовые бугры и все равно обречена на гибель.
Крупная речная и озерно-речная кумжа в основном питается рыбой и может поэтому уничтожать молодь и смолтов семги (Бакштанский и др., 1980; Долотов, 1997; Кашин, 1997; Мартынов, 2007). Молодь семги отмечена в желудках рыб длиной более 16.5 см (Долотов, 1997). Кумжа может питаться молодью акклиматизированной горбуши Oncorhynchus gorbuscha (Бакштанский, 1964). Для кумжи Кольского полуострова отмечено питание мышевидными грызунами (Солдатов, 1908; Лососевые реки ..., 1991; Крылова, 2003; Аверинцев, 2005).
В эстуариях и в море кумжа питается бокоплавами, Gammarus oceanicus, молодью тресковых рыб, песчанки, Ammodytes marinus, трехиглой колюшки, Gasterosteus aculeatus, сельдью, Clupea pallasi, корюшкой, Osmerus mordax (Сент-Илер, 1913; Привольнев, 1934; Мельянцев, 1952; Кузьмин, 1984; Забрусков и др., 1990; Chernitsky et al., 1995). Рыбой питаются некоторые идущие на зимовку особи кумжи реки Кумжевой (данные автора).
Хищники, паразиты и конкуренты
Наблюдения на Кольском полуострове показывают, что злейший враг кумжи в северных водоемах – щука, Esox lucius. Она преследует даже крупных рыб: описан случай поедания щукой нерестовой кумжи массой более 1 килограмма (Аверинцев, Прищепа, 1999). Антагонизм этих видов выявлен статистически – при анализе структуры ихтиофауны малых озер Карельского берега Белого моря показано, что коэффициент ассоциации у кумжи и щуки отрицательный (Иванова и др., 2007). По наблюдениям Н. Плетцова (1936), кумжа не встречается в населенных щукой реках Летнего берега Белого моря.
В море кумжей питаются морской заяц, или лахтак, Erignathus barbatus (Потелов, 1998; Светочева, 2002), морская свинка, Phocaena phocaena, белуха, Delphinapterus leucas и, возможно, гренландский тюлень, Pagophilus groehlandicus (Сурков, 1966). Однако в настоящее время лососи не указываются в числе пищевых объектов двух последних видов (Назаренко, 1984; Матишов, Огнетов, 2006); возможно, это связано с падением численности этих рыб.
Молодь кумжи, как и молодь семги, служит хозяином личинок моллюска европейской жемчужницы, Margaritifera margaritifera. Эти личинки проводят зиму на жабрах лососевых рыб, а потом переходят к самостоятельной жизни. Жемчужница – исчезающий вид, занесена в Красные книги всех уровней – от международного до регионального. Кумжа – единственный хозяин жемчужницы в ряде водоемов Онежского полуострова (Беспалая и др., 2012).
Следует с тревогой отметить, что в регионе совершенно не изучены другие паразиты кумжи, а также ее бактерии и вирусы. Отсутствие паразитологического и микробиологического мониторинга в сочетании с ростом аквакультуры лососевых на Русском Севере и в сопредельных странах обеспечивает все условия для возникновения вспышек опаснейших заболеваний у рыб.
Кроме того, в регионе не изучены конкурентные взаимоотношения кумжи с другими видами рыб, в том числе с чужеродными лососевыми – радужной форелью, Parasalmo mykiss, и горбушей. Между тем нерест кумжи происходит вскоре после нереста горбуши – как раз в период, когда на нерестилищах лежат разлагающиеся рыбины.
Наши наблюдения в бассейнах рек Керети, Рынды и Зимней Золотицы показывают, что молодь кумжи обитает в основном в небольших протоках и ручьях, а молодь семги – в основном русле реки (Махров и др., 2010). Аналогичные наблюдения проведены в бассейнах рек Варзуги и Качковки (Веселов, Калюжин, 2001; Кузищин и др., 2001). Конкуренция молоди двух видов продемонстрирована в ряде зарубежных работ (Kennedy, Strange, 1986; Bremset, Heggenes, 2001; Harwood et al., 2002); показано также, что при падении численности атлантического лосося в реке растет численность кумжи (Hesthagen, 1986).
Атлантический лосось и кумжа, хотя и сходны морфологически, хорошо различаются по ряду генетических маркеров. С применением этих маркеров обнаружены в природе гибриды этих видов. Они выявлены, в частности, в реках Кольского полуострова и северной Карелии, но не в Архангельской области (обзор: Makhrov, 2008).
Можно предположить, что именно конкурентные отношения с атлантическим лососем мешают кумже заселить бассейны крупных рек крайнего северо-востока Европы – Северной Двины, Мезени, Печоры. Отдельные особи, пытающиеся заселить мелкие реки этих бассейнов, не находят друг друга и гибридизируют с атлантическим лососем.
К сожалению, не изучены взаимоотношения кумжи с арктическим гольцом (Salvelinus alpinus), хотя в скандинавских работах есть сведения о конкурентных отношениях этих видов (Svärdson, 1949). Между тем падение численности кумжи по мере продвижения на восток происходит параллельно с ростом численности арктического гольца, лучше адаптированного к условиям Арктики. Вполне вероятно, что именно конкуренция с арктическим гольцом ограничивает распространение кумжи в восточном направлении.
Хозяйственное значение
Большинство поселений на берегах Белого и Баренцева морей стоят в устьях семужьих рек. Однако популяции кумжи всегда были очень важны для небольших групп людей, временно или постоянно живущих вне сел: рыбаков, охотников, оленеводов, староверов, военных и геологов. Кумжа существенно пополняет их ограниченный рацион, а ловля ее удочкой разнообразит их суровую жизнь. Потребительский лов ведется до сих пор во всех населенных кумжей водоемах.
Промысел кумжи, видимо, никогда не был обильным. А. И. Фомин (1805) отмечает, что она привозится в Архангельск в небольшом количестве. Известно, что в 1909 г. в реке Палой было добыто с помощью забора 30 пудов, то есть около 491 кг кумжи (Алеев, 1913); в небольшом количестве кумжа добывалась и в Онеге (Якобсон, 1913, 1914); в 1910 году добыто 14 пудов, то есть 229 кг (Крысанов, 2000). В Чешской губе в этот период лов кумжи носил случайный характер, запасы ее недоиспользовались (Бибиков, 1912), хотя в этом регионе «ставили кумжу, по вкусу, даже выше семги» (Танфильев, 1896).
В течение XX века промысел кумжи постепенно сокращался. Насколько можно судить, это было связано не столько с уменьшением численности кумжи (хотя случаи исчезновения популяций в результате перелова известны), сколько с малой доходностью этого промысла. Рыбака-промысловика на кумяжьих реках и ручьях сменил браконьер. Иногда кумжу просто истребляли, считая конкурентом семги. Так, кумжа полностью изымалась промыслом на Волонге (Даниленко, 1967).
Уже к середине XX века в Чешской губе добывали только 1–2 ц кумжи в год, как прилов при промысле семги (Бурков, Гошева, 1968). В последние годы в Мезенском заливе добывается 0,9 т кумжи, в Двинском заливе – не более 0.8 т, в Онежском заливе – не более 0.4 т (Кулида, Тимофеев, 2005). В реке Онеге кумжа не имеет промыслового значения (Кудерский, Мельникова, 1983). Кумжа – прекрасный объект спортивного рыболовства, но в регионе оно мало развито (Киселев, 1967).
Разработана биотехника разведения беломорской кумжи, в том числе режимы выдерживания производителей и инкубации икры, выращивания молоди, успешно проведены эксперименты по выращиванию кумжи в садках в солоноватой воде (Кулида, Тимофеев, 2005, 2007 и ссылки в этих работах). В настоящее время кумжа воспроизводится на Солзенском и Онежском рыбоводных заводах (личн. сообщ. А. В. Лаврова).
Заключение
Популяции северо-восточного края ареала кумжи отличаются значительным экологическим, морфоэкологическим и генетическим разнообразием. Более широкому распространению кумжи по бассейну Северного Ледовитого океана препятствует, видимо, не ограниченность эволюционного потенциала популяций этого вида, а конкуренция с другими видами лососевых. Можно предположить, что потепление климата приведет к росту численности кумжи побережья Баренцева моря и расширению ее ареала. В то же время, следует отметить, что ситуацию, описанную для края ареала кумжи, нельзя считать типичной: кумжа – инвазионный вид, заселивший целый ряд регионов вне естественного ареала (Lowe et al., 2004).
Библиография
Полный список литературы по кумже бассейнов Белого и Баренцева морей доступен на сайте лаборатории экологии водных сообществ и инвазий Института проблем экологии и эволюции РАН: www.sevin.ru
Аверинцев В. Г. Лососевые речных систем // Ихтиофауна малых рек и озер Восточного Мурмана: биология, экология, ресурсы. Апатиты: Изд-во Кольского НЦ РАН, 2005. С. 47–122.
Аверинцев В. Г., Прищепа Б. Ф. Адаптивные особенности биологии кумжи, гольца и щуки в верховьях рек Восточная Лица и Варзина // Адаптация и эволюция живого населения полярных морей в условиях океанического перигляциала. Апатиты: Изд-во Кольского НЦ РАН, 1999. С. 120–125.
Азбелев В. В. Материалы по биологии семги Кольского полуострова и ее выживаемости // Труды ПИНРО. 1960. Вып. 12. С. 5–70.
Алеев В. Поездка на Летний и Онежский берега Белого моря в 1910 году и описание морских рыболовных угодий // Материалы к познанию русск. рыболовства. 1913. Т. 2. Вып. 2. С. 79–135.
Андросова В. П., Кальянов В. П., Савватимский И. П. Географический очерк западного побережья п-ва Канина по работам экспедиции Госуд. океаногр. института 1930 г. // Труды Гос. океаногр. ин-та. 1934. Т. 4. Вып. 2. С. 3–37.
Анухина А. М. Ихтиофауна Соловецких озер // Труды СевНИОРХ. 1972. Т. 6. С. 94–110.
Артамонова В. С., Махров А. А. Корреляция числа пятен на жаберной крышке с заболеваемостью некрозом плавников и дефектами жаберной крышки у заводской молоди семги (Salmo salar L.) // Марикультура северо-запада России: Тез. докл. научно-практич. конф. 25-27 октября 2000. Мурманск, 2000. С. 14–15.
Артамонова В. С., Махров А. А., Холод О. Н., Пашков А. Н., Решетников С. И., Сумароков В. С., Зубарев А. Н., Ганченко М. В., Сушков В. А. Морфологическая пластичность благородных лососей (Salmo) и ее значение для систематики // Проблемы эволюционной морфологии животных: Тез. межд. конф., посвящ. 100-летию со дня рождения академика А. В. Иванова. Санкт-Петербург, 30 октября – 2 ноября 2006 г. СПб., 2006. С. 12–14.
Бакштанский Э. Л. Воздействие хищников на молодь горбуши Oncorhynchus gorbuscha (Walb.) и кеты Oncorhynchus keta (Walb.) в Белом и Баренцевом морях // Вопросы ихтиологии. 1964. Т. 4. Вып. 1. С. 136–141.
Бакштанский Э. Л., Нестеров В. Д., Симуков Ю. А. О состоянии лососеводства в Мурманской и Архангельской областях // Лососевидные рыбы. Л. 1980. С. 192–206.
Беспалая Ю. В., Болотов И. Н., Махров А. А., Вихрев И. В. Историческая география промысла жемчуга в реках южного Беломорья (Архангельская область) // Известия РАН. Серия географическая. 2012. № 1. С. 96–105.
Бибиков С. Д. Архангельская губерния, ее богатства и нужды. Архангельск: Губернская типография, 1912. 199 с.
Биологические ресурсы Белого моря: изучение и использование (Исследования фауны морей. т. 69). СПб.: ЗИН РАН, 2012. 377 с.
Бурков А. И., Гошева Т. Д. Рыбы и морские млекопитающие Чешской губы и их промысел // Гидробиологические исследования в прибрежных районах Баренцева моря. Л.: Наука, 1968. С. 105–116.
Веселов Е. А., Калюжин С. М. Экология, поведение и распределение молоди атлантического лосося. Петрозаводск: Карелия, 2001. 160 с.
Вылко И. К. Ненецкие географические названия на Новой Земле // Природа и хозяйство Севера. 1981. Вып. 8. С. 102–106.
Гринюк И. Н., Задорина В. М., Исаева С. А. Экология пестряток и покатников семги реки Поноя // Труды ПИНРО. 1977. Вып. 32. С. 87–104.
Гульельми М. Озеро Гокча и производимое в нем рыболовство // Вестник рыбопром. 1888. № 4. С. 105–118.
Даниленко Л. А. Ихтиофауна р. Волонги Архангельской области // Вопросы ихтиологии. 1964. Т. 4. Вып. 2. С. 382–384.
Даниленко Л. А. О повторном нересте семги в реке Волонге // Изв. ГосНИОРХ. 1967. Т. 62. С. 70–78.
Дворянкин Г. А., Новоселов А. П. Современное состояние ихтиофауны и перспективы рыбохозяйственного использования озер Соловецкого архипелага // Рыбное хозяйство. 2005. № 2. С. 68–70.
Долотов С. И. Лососевые реки Кольского полуострова. Река Стрельна. Мурманск: Изд-во ПИНРО, 1997. 39 с.
Дорофеева Е. А. Беломорские форели в системе полиморфного вида Salmo trutta Linnaeus // Проблемы изучения, рац. использования и охраны прир. ресурсов Белого моря. XI Всеросс. конф. с междунар. участием. 9–11 ноября 2010 г. СПб., 2010. С. 46–48.
Дорофеева Е. А., Салманов А. В. Сравнительная остеология форелей рода Salmo и их таксономический статус // Сб. научн. тр. ГосНИОРХ. 2007. Вып. 337. С. 386–396.
Досифей. Географическое, историческое и статистическое описание ставропигиального первоклассного Соловецкого монастыря. Ч. 1. М.: Унив. тип., 1836. 446 с.
Друккер Г. Ф. Рыбный промысел Большеземельской тундры // Труды Ин-та по изучению Севера. 1927. Вып. 38. С. 5–21.
Евсин В. Н. Морфологическая и биологическая характеристика кумжи Salmo trutta Linne реки Малой Кумжевой // Материалы рыбохоз. исследований Северного бассейна. Мурманск, 1970. Вып. 16. Ч. 2. С. 169–182.
Евсин В. Н. Осенняя кумжа реки Варзуги // Труды ПИНРО. 1977. Вып. 32. С. 51–58.
Евсин В. Н. Морфометрические особенности ручьевой форели Salmo trutta L. morpha fario L. реки Пулоньга // Вопросы ихтиологии. 1980. Т. 20. Вып. 2. С. 211–219.
Ершов П. Н. К биологии кумжи рек Пила и Шогуй Кольского полуострова // Исследования популяционной биологии и экологии лососевых рыб водоемов Севера. Л., 1985. С. 94–119.
Житков Б. М. По Канинской тундре // Зап. Имп. Русск. геогр. об-ва по общей географии. 1904. Т. XLI. № 1. С. 1–170.
Забрусков Г. В., Забрускова М. М., Хлевная А. С., Черницкий А. Г., Шкурко Д. С. Кумжа эстуария р. Рында и условия ее существования // Экология и биологическая продуктивность Баренцева моря. М., 1990. С. 110–118.
Задорина В. М. Значение взрослых насекомых в питании молоди лососевых рыб // Вопросы ихтиологии. 1988. Т. 28. Вып. 2. С. 259–265.
Захваткин А. А. Соловецкие озера. Краткий гидробиологический очерк. Соловки: Бюро печати УСЛОН, 1927. 142 с.
Иванова Т. С., Иванов А. А., Ларикова А. В., Шатских Е. В. О структуре ихтиоценозов малых озер Карельского берега Белого моря // Экологические исследования беломорских организмов. Матер. 2-й межд. конф. 18-22 июля 2007 г. СПб., 2007. С. 49–50.
Ивантер Э. В. Периферические популяции политипического вида и их роль в эволюционном процессе // Принципы экологии. 2012. Т. 1. № 2. С. 71–75.
Кашин Е. С. Пищевые взаимоотношения молоди атлантического лосося Salmo salar L. и кумжи Salmo trutta L. р. Стрельна (Кольский полуостров) // Первый конгресс ихтиологов России: Тез. докл. (Астрахань, сентябрь 1997 г.). М., 1997. С. 153.
Киреева Е. А., Саматова Д. С., Черницкий А. Г. Кумжа озерно-речных систем восточного Мурмана // Экология и биологическая продуктивность Баренцева моря (тез. докл. Всесоюз. конф.). Мурманск, июль 1986 г. С. 207–208.
Киселев Я. У самого Белого моря // Рыбоводство и рыболовство. 1967. № 2. С 20.
Козьмин А. К. Рыбные ресурсы рек и озер Европейского Северо-Востока России: их сохранение и использование. Мурманск: Изд-во ПИНРО, 2011. 314 с.
Корнилова В. П. Ихтиофауна низовьев Печоры и Печорского залива Баренцева моря // Мат. рыбохоз. исслед. Сев. бассейна. 1970. Вып. 13. С. 5–44.
Крылова С. С. Кумжа (Salmo trutta L.) Кольского полуострова: Автореф. дисс. … канд. биол. наук. Петрозаводск: Изд-во ПетрГУ, 2003. 26 с.
Крысанов А. А. Поморские промыслы (Онежский уезд 1861-1916 гг.). Онега: Онежский историко-мемориальный музей, 2000. 106 с.
Ксензов Н. А. Некоторые данные по ихтиофауне бассейна р. Пурнач (приток Поноя) // Доклады отделений и комиссий Геогp. об-ва СССР. 1969. Вып. 9. С. 193–198.
Кудерский Л. А., Мельникова М. Н. Рыбы и рыбное хозяйство в бассейне реки Онеги // Современное состояние и качество вод реки Онеги и водоемов ее бассейна. Петрозаводск, 1983. С. 84–99.
Кузищин К. В. Особенности формирования внутривидовой разнокачественности у кумжи Salmo trutta L. Белого моря: Автореф. дисс. ... канд. биол. наук. М.: МГУ, 1997. 17 с.
Кузищин К. В., Махров А. А., Новиков Г. Г. Морфологические особенности кумжи Salmo trutta L. из водоемов бассейна пролива Великая Салма (Карельский берег Белого моря) // Проблемы лососевых на Европейском Севере. Петрозаводск. 1998. С. 31-51.
Кузищин К. В., Новиков Г. Г. Морфоэкологическая дифференциация молоди семги Salmo salar и кумжи S. trutta в небольших потоках (Северная Карелия) // Вопросы ихтиологии. 1994. Т. 34. № 4. С. 479-485.
Кузищин К. В., Новиков Г. Г. Топография нерестилищ и экология нереста кумжи Salmo trutta L. в водоемах бассейна Белого моря // Проблемы изучения, рац. использования и охраны природных ресурсов Белого моря: Тез. докл. VIII регион. научно-практ. конф., 16–18 апреля 2001 г., г. Беломорск, Республика Карелия. Архангельск, 2001. С. 146–148.
Кузнецов Н. М. Рыболовство в реках и озерах Архангельской области. : Архангельск. обл. гос. изд-во, 1951. 167 с.
Кузьмин О. Г. Экологическая характеристика и репродуктивное значение малых лососевых рек Кольского полуострова // Экология биол. ресурсов северного бассейна и их пром. использование. Мурманск, 1984. С. 36–48.
Кулида С. В., Тимофеев В. И. Кумжа рек юго-восточной части Белого моря, перспективы ее искусственного разведения // Биологические ресурсы Белого моря и внутренних водоемов Европейского Севера. Сборник материалов IV (XXVII) междунар. конф. 5-10 декабря 2005 г. Вологда, Россия. часть 1. Вологда. 2005. С. 233-235.
Кулида С. В., Тимофеев В. И. Опыт искусственного воспроизводства беломорской кумжи // Матер. отчетн. сессии Сев. фил. ПИНРО по итогам НИР 2003–2004 гг. Архангельск, 2007. С. 143–151.
Кучина Е. С. Изучение рыбных ресурсов р. Северной Двины // Отчет о научно-иссл. работах Северной базы АН СССР за 1939 год. Архангельск, 1941. С. 52–58.
Лега Ю. В., Черницкий А. Г., Белковский Н. М. Улучшение условий содержания лососей в морской воде в зимний период // Рыбное хозяйство. 1991. № 12. С. 21–23.
Лососевые реки Кольского полуострова. Р. Иоканьга. Мурманск: ПИНРО, 1991. 50 с.
Максимов С. В. Год на Севере. 4-е изд., доп. М.: Изд. П.К. Прянишникова, 1890. 698 с.
Мартынов В. Г. Атлантический лосось (Salmo salar L.) на Севере России. Екатеринбург: УрО РАН, 2007. 414 с.
Матишов Г. Г., Огнетов Г. Н. Белуха Delphinapterus leucas арктических морей России: биология, экология, охрана и использование ресурсов. Апатиты: Изд-во Кольского НЦ РАН, 2006. 295 с.
Махров А. А. Кумжа Salmo trutta L. бассейнов Белого и Баренцева морей // Адаптация и эволюция живого населения полярных морей в условиях океанического перигляциала. Апатиты: Изд-во КНЦ РАН, 1999. С. 110–120.
Махров А. А., Артамонова В. С., Александров Д. И., Широков В. А., Щуров И. Л. Экология кумжи (Salmo trutta Linnaeus) побережий Белого и Баренцева морей // Проблемы изучения, рац. использования и охраны прир. ресурсов Белого моря. XI Всеросс. конф. с междунар. участием. 9–11 ноября 2010 г. СПб., 2010. С. 113–114.
Махров А. А., Иешко Е. П. Генетическая дифференциация и послеледниковое расселение кумжи (Salmo trutta L.) бассейна Белого моря // Биогеография Карелии. Труды Карельского НЦ РАН. Серия Б. Биология. 2001. Вып. 2. С. 175-178.
Мельянцев В. Г. Форели водоемов Карело-Финской ССР. Петрозаводск: Госиздат КФССР, 1952. 88 с.
Муравейко В. М., Степанюк И. А., Емелина А. В. Биология и поведение лососевых рыб северной части Кольского полуострова // Современные исследования ихтиофауны арктических и южных морей европейской части России. Апатиты, 2007. С. 116–134.
Мурза И. Г., Христофоров О. Л. Гаметогенез у кумжи Salmo trutta m. lacustris L. и Salmo trutta trutta L. некоторых водоемов Терского побережья Белого моря // Сб. науч. тр. ГосНИОРХ. 1981. Вып. 174. С. 34–53.
Мурза И. Г., Христофоров О. Л. Динамика полового созревания и некоторые закономеpности фоpмиpования сложной стpуктуpы популяций кумжи Salmo trutta L. из водоемов побеpежья Кандалакшского залива Белого моpя // Сб. науч. тp. ГосНИОРХ, 1984. Вып. 220. С. 41–86.
Назаренко Ю. И. Биология и промысел беломорской популяции гренландского тюленя // Морские млекопитающие. М., 1984. С. 109–117.
Никольский А. М. О соотношении формы тела рыб с быстротой течения воды населяемого ими бассейна // Вестник естествознания. 1891. № 4. С. 137–139.
Новиков П. И. Рыбы водоемов Архангельской области и их промысловое значение. : Сев-Зап. кн. изд-во, 1964. 144 с.
Новоселов А. П. Структура рыбного населения малых рек, впадающих в Онежский залив Белого моря в пределах Архангельской области // Проблемы изучения, рац. использования и охраны природных ресурсов Белого моря (тез. докл. V регион. конф.). Петрозаводск, сентябрь 1992 г. Петрозаводск., 1992.С. 192–194.
Новоселов А. П. Биологическое разнообразие рыб и мониторинг ихтиофауны водоемов // Природа и историко-культурное наследие Кожозерья. Архангельск, 2006. С. 178–185.
Новоселов А. П., Студенов И. И., Дерец В. П. Состав ихтиофауны как показатель видового разнообразия рыб реки Зимняя Золотица (Зимний берег Белого моря) // Малые реки: Межд. научн. конф. Россия, г. Тольятти, 23–27 апреля 2001 г. Тольятти, 2001. С. 151.
Осинов А. Г. К вопросу о происхождении современного ареала кумжи Salmo trutta L. (Salmonidae): Данные по биохимическим маркерам генов // Вопросы ихтиологии. 1984. Т. 24. Вып. 1. С. 11–24.
Осинов А. Г., Берначе Л. «Атлантическая» и «дунайская» филогенетические группы кумжи Salmo trutta complex: генетическая дивергенция, эволюция, охрана // Вопросы ихтиологии. 1996. Т. 36. Вып. 6. С. 762–786.
Плетцов Н. Форелевые речки Летнего берега Белого моря // Архив областного краеведческого музея г. Архангельска. 1936. Т. 4. Оп. 51. л. 90–93.
Пономарева М. В., Пономарева Е. В., Кузищин К. В. Экологические особенности полового созревания кумжи (Salmo trutta L.) // Биоразнообразие и биоресурсы Урала и сопредельных территорий: Матер. III междунар. конф. Оренбург. 25–27 мая 2006 г. Оренбург, 2006. С. 255–257.
Потелов В. А. Отряд Pinnipeda, ластоногие // Фауна европейского Северо-Востока России. Млекопитающие. Т. 2. ч. 2. СПб., 1998. С. 186–242.
Правдин И.Ф. 1966. Руководство по изучению рыб. М.: Пищ. пpомышленность, 376 c.
Привольнев Т. И. К биологии Salmo trutta // Труды Ленингр. об-ва естествоиспытателей. 1934. Т. 63. Вып. 3. С. 311–323.
Пробатов А. Н. Материалы по научно-промысловому обследованию Карской губы и реки Кары // Материалы по научно-промысловому обследованию Карской губы и реки Кары. М., 1934. С. 1–140.
Путешествия академика Ивана Лепехина. Ч. 4. СПб.: При Имп. АН, 1805. 457 с.
Салманов А. В. Анализ изменчивости пластических признаков ручьевой форели (Salmo trutta L. morpha fario L.) р. Пулоньги // Тр. Зоол. ин-та АН СССР. 1991. Т. 235. С. 122–141.
Светочева О. Н. Питание нерпы (Pusa hispida) в Белом море с июня по ноябрь и пищевые взаимоотношения с другими настоящими тюленями // Материалы рыбохозяйственных исследований водоемов Европейского Севера. Архангельск, 2002. С. 404–427.
Семушин А. В., Шерстков В. С., Рухлова В. А. Видовой состав рыб траловых уловов в юго-восточной части Баренцева моря в 1980–2008 годах // Вопросы ихтиологии. 2011. Т. 51. № 6. С. 749–769.
Ихтиологические наблюдения экскуpсии студентов Импеpатоpского Юpьевского Унивеpситета, пpоизведенные летом 1911 года в Ковденском заливе Белого моpя // Вестник pыбопpом. 1913. № 4–6. С. 97–133.
Смирнов А. Г. Семга реки Пинеги, ее жизнь и промысел // Изв. ВНИОРХ. 1935. Т. 20. С. 231–263.
Солдатов В. К. Отчет по исследованию семужьего промысла в 1903 и 1904 гг. // Экспедиция для научно-промысловых исследований у берегов Мурмана. Отчет о работах в 1904 г. СПб. 1908. С. 1–88.
Сурков С. С. Морские млекопитающие, встречающиеся у берегов Кольского полуострова // Рыбы Мурманской области. Мурманск. 1966. С. 143–144.
Суслова Г. Н. Наблюдения над кумжей некоторых рек бассейна Белого моря // Изв. ГосНИОРХ. 1969. Т. 65. С. 135-140.
Танфильев Г. И. О рыбных и звериных промыслах в водах Мезенского и Печорскаго края // Вестник рыбопром. 1896. № 2–3. С. 53–64.
Фомин А. И. Опыт исторический о морских зверях и рыбах, промышляемых Архангельской губернии жителями в Белом море, Северном и Ледовитом океане, с описанием образа тех промыслов // Путешествия академика Ивана Лепехина. ч. 4. В 1772 году. СПб.: При Имп. АН, 1805. С. 304–370.
Христофоров О. Л., Мурза И. Г. Уровни эстрадиола и тестостерона в крови атлантического лосося Salmo salar L. и кумжи Salmo trutta trutta L. в периоды осуществления смолтификации и некоторых этапов гаметогенеза // Сб. науч. тр. ГосНИОРХ. 1988. Вып. 276. С. 54–72.
Христофоров О. Л., Мурза И. Г. Половое созревание и структура популяции кумжи реки Поной // Биол. ресурсы Белого моря и внутр. водоемов Европейского Севера: Тез. докл. Сыктывкар. 1990. С. 37.
Якобсон Р. П. Статистико-экономическое обследование морского побережья и рыболовных угодий на Онежской губе между г. Кемью и Онегой и Онежского закола в 1911 г. // Матер. к познанию русск. рыболовства. 1913. Т. 2. Вып. 5. 60 с.
Якобсон Р. П. Отчет по обследованию Онежского бассейна и по поездке на Мурман и на Новую Землю в навигацию 1912 г. // Матер. к познанию русск. рыболовства. 1914. Т. 3. Вып. 11. 57 с.
Якобсон Р. П. Рыболовство Печорского бассейна в пределах Пермской и Вологодской губерний в 1914 г. // Материалы к познанию русского рыболовства. 1916. Т. 5. Вып. 4. С. 1–44.
Bernatchez L. The evolutionary history of brown trout (Salmo trutta L.) inferred from phylogeographic, nested clade, and mismatch analyses of mitochondrial DNA variation // Evolution. 2001. V. 55. P. 351–379.
Bernatchez L., Osinov A. Genetic diversity of trout (genus Salmo) from its most eastern native range based on mitochondrial DNA and nuclear gene variation // Molecular Ecology. 1995. V. 4. P. 285–297.
Bremset G., Heggenes J. Competitive interactions in young Atlantic salmon (Salmo salar L.) and brown trout (Salmo trutta L.) in lotic environments // Nordic J. Freshw. Res. 2001. V. 75. P. 127–142.
Chernitsky A. G., Zabruskov G. V., Ermolaev V. V., Shkurko D. S. Life history of trout, Salmo trutta l., in the Varsina river estuary (the Barents Sea) // Nordic J. Freshw. Res. 1995. V. 71. P. 183–189.
Hardie D. C., Hutchings J. A. Evolutionary ecology at the extremes of species' ranges // Environ. Rev. 2010. V. 18. P. 1–20.
Harwood A. J., Metcalfe N. B., Griffiths S.W., Armstrong J. D. Intra- and inter-specific competition for winter concealment habitat in juvenile salmonids // Can. J. Fish. Aquat. Sci. 2002. V. 59. P. 1515–1523.
Hesthagen T. Fish kills of Atlantic salmon (Salmo salar) and brown trout (Salmo trutta) in an acidified river of SW Norway // Water, Air, and Soil Pollution. 1986. V. 30. P. 619–628.
Jones J. W. Salmon and trout hybrids // Proc. Zool. Soc. London. 1947. V. 117. P. 708–715.
Jonsson B., Jonsson N. Ecology of Atlantic Salmon and Brown Trout. Habitat as a Template for Life Histories. Dordrecht: Springer, 2011. 708 p.
Kawecki T. J. Adaptation to marginal habitats // Annu. Rev. Ecol. Evol. Syst. 2008. V. 39. P. 321–342.
Kennedy G.J.A., Strange C.D. The effects of intra- and inter-specific competition on the survival and growth of stocked juvenile Atlantic salmon, Salmo salar L., in relation to depth and gradient in an upland trout, Salmo trutta L., stream // J. Fish. Biology. 1986. V. 29. P. 199–214.
Lowe S., Browne M., Boudjelas S., De Poorter M. 100 of the world’s worst invasive alien species. A selection from the global invasive species database. Auckland: Invasive species specialist group, 2004. 12 p.
Lumme J., Asplund T., Kuusela J., Veselov A., Bakhmet I., Potutkin A., Primmer C. Endemic Karelian strains of brown trout, Salmo trutta L.: a preliminary analysis by mitochondrial DNA // Лососевидные рыбы Восточной Фенноскандии. Петрозаводск. 2005. С. 71–80. (www.krc.karelia.ru)
MacCrimmon H. R., Marshall T. L. World distribution of Brown Trout, Salmo trutta // J. Fish. Res. Bd. Canada. 1968. V. 25. No. 12. P. 2527–2548.
Makhrov A. A. Hybridization of the Atlantic salmon (Salmo salar L.) and brown trout (S. trutta L.) // Zoosystematica Rossica. 2008. V. 17. No. 2. P. 129–143.
Makhrov A. A., Skaala O., Altukhov Yu. P. Alleles of sAAT-1,2* isoloci in brown trout (Salmo trutta L.): potential diagnostic marker for tracking routes of post-glacial colonization in northern Europe // Journal of Fish Biology. 2002. V. 61. P. 842–846.
Novikov G. G., Politov D. V., Makhrov A. A., Malinina T. V., Afanasiev K. I., Fernholm B. Freshwater and estuarine fishes of the Russian Arctic coast (the Swedish-Russian Expedition 'Tundra Ecology - 94') // J. Fish Biology. 2000. V. 57. No. a. P. 158–162.
Presa P., Pardo B.G., Martinez P., Bernatchez L. Phylogeographic congruence between mtDNA and rDNA ITS markers in brown trout // Mol. Biol. Evol. 2002. V. 19. № 12. P. 2161–2175.
Salmanov A. V., Dorofeyeva E. A. Comparative osteology of river and lake trouts (genus Salmo) // Proc. Zool. Inst. 2001. V. 287. P. 149–195.
Svärdson G. Competition between trout and char (Salmo trutta and S. alpinus) // Report of the Institute of Freshwater Research, Drottningholm. 1949. No. 29. P. 107–111.
Thomsen D. S., Koed A., Nielsen C., Madsen S. S. Overwintering of sea trout (Salmo trutta) in freshwater: escaping salt and low temperature or an alternate life strategy? // Can. J. Fish. Aquat. Sci. 2007. V. 64. P. 793–802.
Благодарности
Пользуюсь случаем выразить искреннюю благодарность коллегам, помогавшим мне в ходе полевых работ и снабжавшим литературой: В. С. Артамоновой, Ю. В. Беспалой, И. Н. Болотову, Н. П. Бушуевой, В. П. Дерецу, Г. Ю. Жаркову, В. М. Зеленкову, А. К. Козьмину, А. Кононову, К. В. Кузищину, С. В. Кулиде, Д. Л. Лайусу, В. Г. Мартынову, А. П. Новоселову, Г. В. Севастьяновой, И. И. Студенову, О. Тюкову, З. Н. Юдиной. Подготовка работы осуществлялась при финансовой поддержке грантов РФФИ № 11-04-00697-а и 11-04-98815-р_север_а, а также программы «Живая природа: современное состояние и проблемы развития» (подпрограмма «Динамика и сохранение генофондов»).

 © 2011 - 2026
© 2011 - 2026