



Введение
Фауна паразитов представляет собой группу видовых популяций, совместно обитающих в популяции того или иного хозяина (или нескольких хозяев). Такое «компонентное сообщество» (Esch et. аl., 1990) следует рассматривать как некое организованное целое, отражающее взаимодействие между входящими в его состав видами и сложившимися эволюционными отношениями с хозяином (как местом обитания). Для паразитов характерно понятие специфичности или облигатности, т. е. есть целый ряд видов, которые способны существовать только на данном хозяине или близких видах. В то же время, в состав фауны могут входить и виды, способные паразитировать на многих видах рыб. Таким образом, паразитофауна отдельного вида хозяина включает (по аналогии со свободноживущими животными) стено- и эврибионтные виды. При этом в большинстве случаев оказывается, что специфичные виды доминируют по встречаемости в паразитофауне отдельного хозяина, тогда как из широко распространенных видов только единичных представителей можно отнести к массовым (Иешко и др., 1982).
Существуют определенные трудности при определении размеров природных сообществ, в связи с чем концепция сообщества часто рассматривается как некая абстракция, просто набор видов (Vandermeer, 1972). Однако существует метод объективного определения размеров сообщества, который учитывает зависимость видового богатства от площади обследованной территории (или числа собранных проб, что почти эквивалентно обследованной величине площади). Даже на простой диаграмме зависимости числа видов от увеличения обследованной площади можно определить минимальную площадь территории, содержащую репрезентативное количество видов данного сообщества, – как то значение, в котором оценки видового богатства выходят на плато (Джиллер, 1988). Этот метод можно взять на вооружение для исследования паразитарных сообществ.
Изучению закономерностей формирования видового богатства паразитофауны рыб посвящена практически единственная в отечественной литературе работа Г. К. Петрушевского и М. Г. Петрушевской (1960). На основе анализа большого фактического материала ими было показано, что видовое богатство паразитофауны имеет закономерную связь с числом исследованных рыб. Вместе с тем в работе отсутствовали какие-либо аналитические построения.
Создание модели зависимости паразитофауны или видового богатства компонентного сообщества от количества исследованных хозяев позволяет оценить репрезентативность параметров видового богатства, а также послужить основанием для сравнения видового разнообразия различных выборок.
Цель нашей публикации состоит в том, чтобы апробировать некоторые технологии изучения биоразнообразия наземных сообществ для оценки видового богатства паразитофауны рыб озер Карелии (на примере одного вида) и выявить существенные факторы, влияющие на характер и динамику показателей видового богатства паразитарного сообщества при росте выборки рыб.
Материалы
В качестве исходной информации нами были использованы данные по паразитофауне щуки (Esox lucius) системы р. Каменной за 1974–1977 гг. (озера Нюк, Лувозеро, 2 точки отлова в Кимасозере – западная и восточная части) (рис. 1) (Иешко и др., 1982). Озера Лувозеро и Кимасозеро облавливали по всей акватории, оз. Нюк – преимущественно в западной половине; состав рыбного населения был сходным.
В анализе использовано 103 щуки. Для каждой особи хозяина регистрировали факт встречи разных видов паразитов. Всего обнаружено 28 видов паразитов (из 133 видов паразитов, обнаруженных у всех исследованных видов рыб, включая сиговых и лососевых). Основную группу паразитов составляют представители бореально-равнинного комплекса. Это эвритермные, широко распространенные в Палеарктике паразиты щуки, ерша, плотвы, ельца, язя.
Для каждого из четырех изученных местообитаний были сформированы исходные таблицы данных, состоящие из s строк (по числу видов паразитов: j = 1, 2, … s) и n столбцов (по числу исследованных особей хозяина: i = 1, 2, … n). В ячейки были помещены оценки численности или отмечены факты регистрации j-го вида паразита для i-го хозяина.
Рис. 1. Места отбора проб в озерах системы р. Каменной (Карелия)
Fig. 1. Sampling locations in the lakes system of Kamennaya river (Karelia)
Методы
Лов рыбы на озерах проводился стандартным набором одностенных сетей с ячеей от 14 до 60 мм. Лабораторная обработка ихтиологического материала, определение размеров, пола и возраста, проводились по методикам И. Ф. Правдина (1966), Н. И. Чугуновой (1959), с учетом рекомендаций Ю. С. Решетникова (1980) и М. В. Мины (1981). Исследования зараженности рыб проводились методом полного паразитологического вскрытия, с изучением всех основных систематических групп паразитов, включая паразитов крови. Для видовой идентификации обнаруженных паразитов использовали Определитель … (1962).
Первичная обработка числовых данных для каждого местообитания состояла в том, чтобы получить ряды чисел, характеризующих постепенное накопление числа зарегистрированных видов паразитов при увеличении выборки изученных рыб. Так, если в первой особи случайной выборки было обнаружено 6 видов паразитов, то первое значения ряда накопления видового богатства равно 6. Если вторая случайно взятая особь имела 8 видов паразитов, из которых 3 было обнаружено в первой рыбе, а 5 встречено впервые, то второе число накопленного видового богатства становится равным s2 = 6 + 5 = 11. Составление ряда продолжается до исчерпания выборки (n). Так получали ряды (s1, s2, … si, … sn) роста видового богатства паразитов в каждом местообитании.
Процесс регистрации новых видов был автоматизирован в среде Excel. Для каждой таблицы исходных данных на листе Excel была построена дополнительная таблица, в которой с помощью операторов попарного сравнения смежных особей щуки выяснялось, был ли учтен данный вид паразита у предыдущих особей. Отсутствие паразита обозначалось числом 0. При первой встрече j-го паразита в i-м хозяине ячейке приписывалось число 1 и повторялось плоть до n. Сумма таких значений для всех паразитов si (для очередного i-го хозяина) представляет собой видовой список фауны паразитов, известных к моменту учета j-й особи хозяина. Таким образом, получали ряды чисел (s1, s2, … si, … sn), отображающие динамику пополнения видового списка паразитов по мере увеличении числа обследованных особей хозяина. Организация автоматического пересчета значений si позволяет получать новые ряды si при перестановке особей хозяина в матрице исходных данных.
Изменение порядка следования особей хозяина существенно влияет на результирующую кривую насыщения видового списка. Заметим, что включение в анализ рыб в порядке очередности их отлова в общем случае не будет оптимальным со статистической точки зрения, поскольку такая случайная траектория насыщения видового списка является всего лишь одно из большого числа возможных траекторий. Для конкретной выборки особей хозяина все многообразие кривых насыщения можно уложить между двумя крайними рядами si: в первом ряду особи хозяина расположены в порядке роста богатства индивидуальной паразитофауны, во втором ряду – в порядке снижения богатства инфрафауны. Первый ряд будет давать плавно возрастающую кривую пополнения видового списка паразитов (smin), второй – кривую, круто возрастающую и быстро достигающую плато (smax). Для характеристики каждой выборки мы взяли промежуточную, среднюю кривую: s = (smin+ smax)/2 (рис. 2).
Зависимости средней кривой видового богатства от числа изученных особей аппроксимировали с помощью логарифмических уравнений регрессии (Коросов, 2007). Поскольку для нулевой особи хозяина число видов паразитов также составляет 0, в ряды si для всех озер мы добавили одинаковое «нулевое» значение: при i = 0.1, s = 0.1 (значение 0 нельзя добавлять, поскольку логарифм 0 не определен). В результате все кривые прошли через начало осей координат. Уравнение регрессии было призвано выразить долю видового богатства паразитов, привносимого очередной особью относительно оценок общего видового богатства. Базы данных готовились в среде Excel. Комбинирование (ресамплинг), сортировка, фильтрация данных, расчет уравнений регрессии и их значимости выполнялись в среде R (Шитиков, Розенберг, 2012). Статистическое сравнение распределений выполняли по критерию Колмогорова – Смирнова с помощью пакета Statgraphics (Коросов, Горбач, 2010).
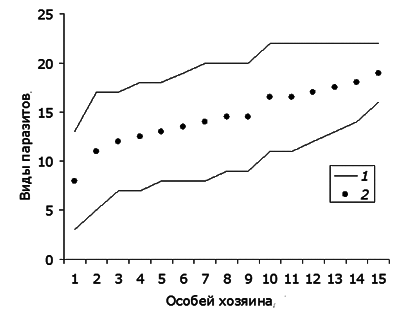
Рис. 2. Границы диапазона (1) возможных траекторий накопления видового богатства инфрасообществ паразитов и срединное значение между ними (s, 2) (фрагмент данных для оз. Нюк)
Fig. 2. The range (1) of possible trajectories of the accumulation of species richness in parasites infracommunity and their median value (s, 2) (data fragment for the lake Nook)
Результаты
Щука в озерах системы реки Каменной является широко распространенным и массовым видом. Фауна паразитов разнообразна и состоит из 11 видов простейших, 1 вида моногеней, 5 – цестод, 3 – трематод, 3 – нематод, и 1 вида ракообразных. Из 28 видов 10 являются массовыми и встречаются практически во всех исследованных озерах (табл. 1), формируя тем самым ядро фауны специфичных для щуки видов паразитов. Встречаемость оставшихся видов преимущественно состоит из неспецифичных видов, которых можно отнести к редким.
Таблица 1. Встречаемость (%) специфичных паразитов щуки в озерах системы реки Каменной
| Лувозеро | Кимасозеро1 | Кимасозеро2 | Нюк | |
| Myxidium liberkuhni | 78 | 88 | 84 | 73 |
| Chloromyxum isocinum | 26 | 14 | 52 | 33 |
| Myxosoma anurus | 35 | 12 | 32 | 70 |
| Tetraonchus monenteron | 91 | 64 | 84 | 80 |
| Triaenophorus crassus | 91 | 36 | 56 | 80 |
| Triaenophorus nodulosus | 65 | 48 | 72 | 87 |
| Azigia lucii | 35 | 56 | 52 | 27 |
| Cammallanus lacustris | 22 | 24 | 32 | 37 |
| Phylometra obturans | 52 | 8 | 20 | 27 |
| Raphidascaris acus | 78 | 16 | 64 | 87 |
| Всего видов паразитов | 17 | 11 | 17 | 23 |
Наибольшее видовое разнообразие паразитов щуки (23 вида) обнаружено в оз. Нюк, самом крупном из исследованных водоемов, самая бедная фауна (11) – в выборке из оз. Кимасозеро1 (табл. 2). Видовое богатство паразитов инфрасообщества (количество видов паразитов на одной особи хозяина) варьирует от 1 до 13. Минимальное значение обнаружено в оз. Кимасозеро1, максимальное – в оз. Нюк. Среднее богатство инфрсообществ имеет тот же порядок отличий (4.2 против 7 видов), что и показатели вариабельности (дисперсии состава инфрасообщества): наименьшее значение (2.5 ) отмечено для оз. Кимасозеро1, наибольшее (4.5) – оз. Нюк.
Таблица 2. Показатели видового богатства инфрасообществ и паразитофауны щуки из различных озер Карелии
| Нюк | Кимасозеро1 | Кимасозеро2 | Лувозеро | |
| Исследовано рыб | 30 | 25 | 25 | 23 |
| Число видов / особь, мин. | 3 | 1 | 3 | 4 |
| Число видов / особь, макс. | 13 | 8 | 11 | 11 |
| Число видов / особь, средн. | 6,8 | 4,2 | 6,7 | 6,5 |
| Ошибка средней | 0,4 | 0,3 | 0,4 | 0,3 |
| Дисперсия | 4,5 | 2,5 | 3,8 | 2,5 |
| Медиана | 7,0 | 4,0 | 6,0 | 6,0 |
| Паразитофауна в целом | 23 | 11 | 17 | 17 |
Средний уровень и вариабельность видового состава паразитов, которые регистрируются на конкретном хозяине, определяют характер динамики роста видового богатства паразитофауны при изучении большего числа особей хозяина. В разных озерах она имеет разный ход (рис. 3). При этом траектории кривых роста видового богатства щуки оз. Нюк и Кимасозеро1 визуально отличаются. Сообщества паразитов в выборках щук Кимасозеро2 и Лувозеро, представленные близким количеством видов (17 и 17 видов), имели сходные кривые роста пополнения видового списка.
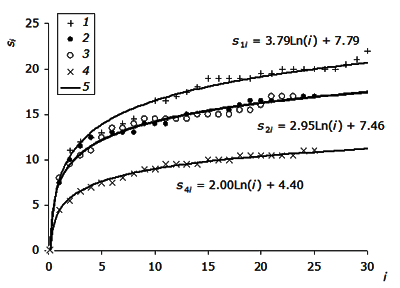
Рис. 3. Средняя динамика пополнения видового списка паразитов щуки в озерах Нюк (1), Кимасозеро1 (2), Лувозеро (3), Кимасозеро2 (4); 5 – логарифмические тренды
Fig. 3. Mean change in species replenish inventory of parasites of pike in the lake Nook (1), Kimasozero1 (2), Luvozero (3), Kimasozero2 (4) 5 – logarithmic trend
Подходя к сравнению выборок из разных озер, следует уяснить смысл коэффициентов логарифмической регрессионной модели si = a×ln(i) + b. Свободный член b представляет собой среднюю величину видового богатства паразитов рыб, которую дает изучение первой случайно взятой рыбы (s1 = a×ln(1) + b = a×0 + b = b); это среднее число паразитов, представленных в каждой рыбе данного вида (в данном местообитании). Коэффициент пропорциональности a стоит перед логарифмом номера особи хозяина. Он выражает средний прирост оценки видового богатства, полученный после исследования очередной особи, пропорциональный логарифму номера этой особи. У первой рыбы этот прирост равен нулю, но чем больше изучено рыб, тем больше видов паразитов уже учтены.
При сопоставлении свободных членов уравнений (рис. 3) видно, что в озерах Нюк, Лувозеро, Кимасозеро1 они примерно одинаковы и равны 7.5-7.8. Для оз. Кимасозеро2 это значение почти в два раза ниже – 4.4. Таким образом, общая зараженность щук первых трех выборок существенно выше, чем четвертой выборки. Коэффициенты пропорциональности a выше для выборки из оз. Нюк, существенно ниже для озер Кимасозеро1 и Лувозера и еще ниже – для выборки из оз. Кимасозера2. Это значит, что в оз. Нюк среди исследованных рыб доминируют щуки с большим количеством видов паразитов, в оз. Лувозеро и Кимасозеро1 видовое богатство инфрасообществ меньше, а в оз. Кимасозеро2 фауна паразитов самая бедная, и при этом большая часть рыб несет малое число видов паразитов.
Более детальную картину дает гистограмма распределения числа видов паразитов в особях хозяина в различных водоемах (рис. 4). Для оз. Нюк распределение оказалось широким и плосковершинным и смещенным вправо с медианой 7, для озер Кимасозеро2 и Лувозеро – более островершинным с медианами 6, для Кимасозеро1 – островершинным с медианой 4. По критерию Колмогорова – Смирнова значимо (p < 0.01) отличаются распределения видового богатства инфрасообществ паразитов щуки из озер Нюк и Кимасозеро1, Кимасозеро1 и Кимасозеро2.
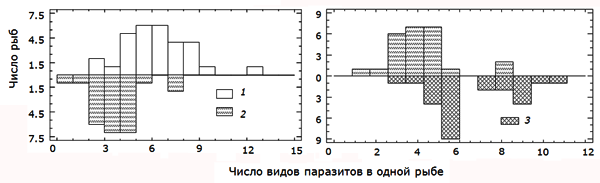
Рис. 4. Распределение значений видового богатства инфрасообществ паразитов щуки оз. Нюк (1) и Кимасозера1 (2), Кимасозера2 (3)
Fig. 4. Distribution of the values of species richness in the parasites infracommunity of pike in the lake Nook (1) and Kimasozera1 (2), Kimasozera2 (3)
Иными словами, распределение видового богатства исследованных рыб варьирует в зависимости от особенностей выборки: в популяции щуки есть группы с высоким и низким видовым богатством. Одним из возможных факторов, определяющих характер варьирования количества видов паразитов, является возраст исследуемых рыб.
В нашем случае отсутствует простая положительная зависимость числа паразитов в особи от возраста щуки. Как видно на диаграмме (рис. 5) наибольшая зараженность хозяина с возрастом вначале возрастает, потом стабилизируется около среднего уровня. Наибольшую зараженность имеют средневозрастные 4-6 летние особи, которые и играют основную роль в поддержании популяций паразитов.
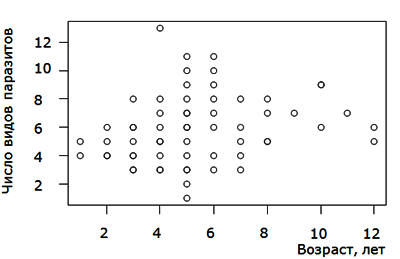
Рис. 5. Зависимость числа видов паразитов от возраста щуки в изученных озерах
Fig. 5. Dependence of the number of parasites on the age of pike in the studied lakes
Анализ возрастного состава исследованных выборок рыб показал следующее (рис. 6). В озерах Нюк, Лувозеро и Кимасозеро2 щуки имели близкие по своему характеру возрастные распределения (медиана составляет 5 лет). Высокая доля рыб младших возрастных групп среди щук из Кимасозера1 (медиана 4 года) вызвала достоверное (p < 0.05) отличие их возрастного распределения от прочих. С другой стороны, при определенном сходстве выборка из оз. Нюк отличалась большим числом особей старше 7 лет (и достоверно большей дисперсией), чем выборки из других озер.
В силу общего сходства возрастной структуры, у исследованных рыб были обнаружены практически все специфичные виды паразитов (табл. 1) и среднее количество паразитов у одной исследованной рыбы из оз. Нюк, Кимасозеро2 и Лувозеро имели близкие значения (табл. 2). Только щуки из Кимасозеро1, представленные в основном молодыми особями, имели минимальные значения видового богатства инфрасообществ паразитов (4,2 вида на рыбу).
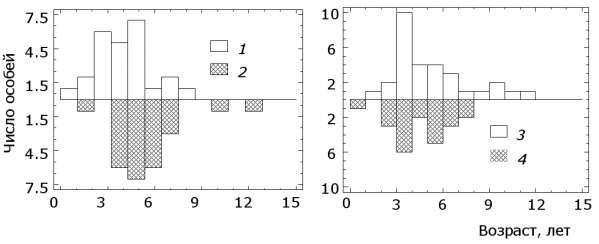
Рис. 6. Возрастной состав исследованных щук в оз. Кимасозеро1 (1) и Кимасозеро2 (2), Нюк (3) и Лувозеро (4)
Fig. 6. The age composition of the investigated pike in the lake Kimasozero1 (1) and Kimasozero2 (2) Nook (3) and Luvozero (4)
Эти материалы показывают превалирование наиболее зараженных возрастных классов именно в тех озерах, где выше и общая зараженность рыб (оз. Нюк).
Построенную нами регрессионную модель можно использовать для прогноза числа обнаруженных видов паразитов при исследовании определенного числа особей рыбы (например, для оз. Нюк). Так, 25 щук в среднем будет содержать 20 видов паразитов, 50 – 23 вида (реальный объем), 100 – 25 видов. Если использовать эти цифры для расчета относительной скорости прироста видового разнообразия, то окажется (рис. 7), что первые 3–4 особи содержат 50 % видов паразитов, 7 особей – 60–67 %, а 15 рыб несут до 80 % всех представителей фауны паразитов.
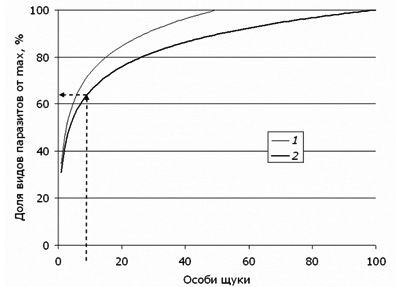
Рис. 7. Прогноз среднего видового богатства паразитофауны при росте числа обследованных особей щуки для оз. Нюк (с помощью уравнения для оз. Нюк y = 3.79Ln(x) + 7.79). 1 – за максимум приняты показатели видового богатства для 50 экз. рыбы, 2 – за максимум приняты показатели для 100 экз. рыбы
Fig. 7. Forecast of the average species richness of parasite fauna with the number of studied samples of pike in the lake Nook (using equations for the lake Nook y = 3.79Ln (x) + 7.79). 1 – indicators of species richness for 50 specimens of fish are taken for maximum, 2 – Indicators for 100 specimens of fish are taken for maximum
Сопоставление этих цифр с динамикой пополнения списка специфических для щуки паразитов (табл. 1), образующих половину фауны (11 из 23 видов), показывает, что первые 3–4 особи хозяина несут до 80 %, 7 особей – 95 %, а 15 особей – 100 % специфических паразитов (рис. 8). Сравнение рядов для всей фауны и для специфических видов говорит о том, что в среднем в выборке из 15 особей хозяина обнаруживаются все специфические виды паразитов и около 2/3 от списка широко распространенных видов паразитов (а в целом около 80 % ото всей паразитофауны).
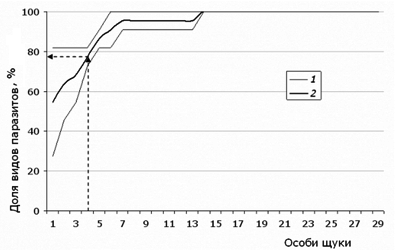
Рис. 8. Границы диапазона (1) возможных траекторий накопления видового богатства инфрасообществ специфических для щуки паразитов (всего 11 видов) и срединное значение между ними (2) (данные для оз. Нюк)
Fig. 8. The range (1) of possible trajectories of accumulation of species richness in infracommunity of specific pike parasites (11 species altogether), and the their median value (2) (data for the lake Nook)
В этом контексте становится актуальным вопрос, в какой мере небольшая случайная выборка (например, из 15 рыб) способна воспроизвести характеристики разнообразия паразитофауны. Для его предварительного решения был выполнен следующий анализ (для рыб из оз. Нюк). Из полной выборки в 30 особей случайным образом отобрали 5 выборок по 15 экз. Для них построили тренды срединных значений (рис. 9). Некоторые из этих трендов точно совпали с трендом, полученным для полной выборки (30 экз.), другие давали заниженный или завышенный прогноз. На уровне 15 экз. хозяина варьирование показателя видового богатства составило 3 вида (+1 и –2, или +7 % и –13 %).
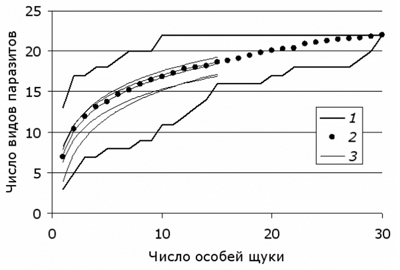
Рис. 9. Тренды для срединных значений пяти выборок по 15 экз. щуки (3) и для полной выборки (30 экз., 2) в границы диапазонов возможных траекторий накопления видового богатства паразитофауны оз. Нюк (1)
Fig. 9. Trends for the median values of five samples of 15 specimens of pike (3) and for the full sample (30 specimens., 2) in the ranges of possible trajectories of accumulation of parasite species richness in the lake Nook (1)
Обсуждение
Представленные материалы позволяют обсудить следующие три темы: насколько обосновано описание большого семейства рядов динамики накопления видового богатства единственной кривой? каковы причины различия динамики накопления видового богатства паразитов в изученных местообитаниях? позволяет ли полученная модель дать количественный прогноз репрезентативного объема паразитофауны данного местообитания?
Как показано в ряде работ (Джиллер, 1988, Poulin, 2007 и др.), одна из причин, затрудняющая сравнение паразитофаун (или компонентных сообществ) – это неравномерность выборочного усилия. Низкие показатели видового богатства могут действительно соответствовать бедной паразитофауне, а могут быть связаны с тем, что большая часть видов паразитов не была обнаружена, хотя паразитофауна составлена большим числом видов. При этом предлагаются различные варианты оценки видового богатства, при которых учитывается эффект неравномерности исследуемой выборки (Poulin, 2007). Следует отметить, что при этом мало уделяется внимания технике формирования рядов накопления видового богатства. Нельзя обсуждать характер этой динамики в отрыве от метода формирования ряда. Как было показано выше, простая (случайная) перестановка проб (рыб) приводит к появлению динамических рядов разной формы (соответственно, и разных моделей); значит, необходим унифицированный алгоритм их формирования. На наш взгляд, таким универсальным приемом может быть усреднение двух рядов с крайними скоростями накопления – максимально быстрого (особи ранжированы в прядке убывания богатства инфрасообщества) и самого медленного (особи ранжированы в прядке возрастания богатства инфрасообщества). Выполненный нами ресамплинг показал, что уже 5 рандомизированных выборок дают средний ряд накопления, практически совпадающий с рядом, построенным по двум крайним траекториям. Технология построения срединного ряда накопления видового богатства позволяет корректно выполнить описание биоразнообразия выборки с помощью логарифмической фукнции (si = a × ln(i) + b) и подойти к сравнению выборок с разными объемами.
Наши расчеты (которые не отражены в результатах) показали, что логарифмическая функция хорошо подходила только для описания динамики срединного ряда, но плохо соответствовала крайним рядам. Ряд самого медленного накопления видового богатства лучше всего описывался степенной функций. Ряд накопления с максимальной скоростью одинаково плохо описывался всеми видами простых функций, и высокую адекватность имела только динамическая модель, основанная на зависимости величины прироста биоразнообразия паразитов от добавления очередной новой особи хозяина. Эти факты означают, что те отмеченные в литературе случаи, когда логарифмическая функция плохо описывает кумуляту видового богатства, не следует относить к биологическим феноменам. По большей части – это артефакты, связанные с отсутствием адекватной методики формирования анализируемого ряда; чаще всего такие ряды соответствуют всего лишь последовательности получения данных, но не вариациям структуры изучаемых сообществ. С другой стороны, если логарифмическая функция подходит не к любой динамике накопления биоразнообразия, то ее нельзя считать специфическим «выразителем» процесса пополнения видового списка при росте объемов исследований. Это биологически «бессмысленный», но статистически адекватный инструмент для описания обобщенной динамики серии случайных процессов.
Возвращаясь к биологическому предмету исследований, следует отметить, что эффективнее было бы не только искать возможности нивелировать неоднородность выборочного усилия с применением математического аппарата, но и использовать корректную методологию сбора данных. Репрезентативность выборочного усилия обеспечивается в тех случаях, когда учитываются видовые и экологические особенности хозяина. Необходимо чтобы исследуемая выборка в полной мере характеризовала особенности популяционной структуры фауны паразитов. Это значит, что обязательно следует учитывать соотношение молодых и взрослых особей хозяина. Рыбы разного возраста физиологически и экологически различаются, что определяет их неодинаковую функциональную роль в поддержании численности и видового разнообразия паразитов. Благодаря особенностям экологии, поведения, питания, реактивности внутренней среды и пр. на разных фазах онтогенеза хозяин имеет неодинаковые шансы заразиться. Особи разного возраста получают разные дозы заражения, создают разные условия для выживания паразитов (Иешко, Голицына, 1984). Наблюдаемая на организменном уровне регуляция паразитом своих отношений с окружающей средой через хозяина (Шульман, Добровольский, 1977) прослеживается и при взаимодействии популяций паразита и хозяина. По этой причине при выполнении популяционных исследований важно знать, какая размерно-возрастная группа обеспечивает наиболее благоприятные условия для поддержания численности паразита, а какая играет подчиненную роль. В этих случаях различия в богатстве исследуемых паразитофаун будут связаны не с неравномерностью выборочного усилия, а с действием существенных для паразитов экологических факторов.
При сопоставлении представленных фактов складывается следующее объяснение разного хода кривых роста видового богатства в разных местообитаниях. В крупных озерах, каким является оз. Нюк, структура популяции щуки представлена многочисленными литоральными и пелагическими группировками (Первозванский, 1985), в силу этих причин в опытных уловах часто встречались рыбы старших возрастов. В малых озерах доля рыб старших возрастов, ведущих пелагический образ жизни, значительно меньше, и поэтому в уловах преобладают рыбы средних и младших возрастов. Отмеченные особенности в возрастной структуре исследованных в разные годы рыб Кимасозера связаны с различной долей возрастных групп щуки в опытных уловах. При доминировании младших возрастов рыб в уловах (Кимасозеро1) наблюдается обеднение видового богатства фауны паразитов, которая представлена преимущественно облигатными видами паразитов, а при увеличении доли старших возрастов (Кимасозеро 2), разнообразие и число обнаруженных видов возрастает.
Таким образом, можно полагать, что основным фактором, непосредственно определяющим богатство паразитарного сообщества щуки, является возрастной состав популяции, который, в свою очередь, определяется особенностями популяционной динамики в специфических условиях обитания (размером озера, разнообразием биотопом, антропогенной нагрузкой и пр.).
Для практики паразитологических исследований остается немаловажным вопрос: позволяют ли построенные кривые зависимости числа обнаруженных видов паразитов от количества обследованных хозяев определить то минимальное число особей, которое содержит репрезентативное число видов в паразитарном сообществе или в паразитофауне конкретного хозяина, его «конечный» репрезентативный объем? Как отмечается в литературе (Петрушевский, Петрушевская, 1960), исследование уже первых 15 экз. рыб позволяет учесть все специфичные виды паразитов, зараженность которыми превышает 20 %. Наши данные также показывают, что 15 экземпляров хозяина несут около 100% специфических и до 70 % широко распространенных видов паразитов. «Удача» или «неудача» при сборе полевого материала по паразитофауне отличаются весьма небольшой величиной: прирост объема выборки на 100 % (с 15 до 30 экз.) в лучшем случае повысит информационную насыщенность материалов на 10 %.
Заключение
Под экологическим подходом к изучению фауны паразитов рыб мы понимаем, во-первых, представление о том, что «совокупность паразитов, обитающих в одном каком-либо хозяине, представляет собой своеобразный биоценоз, имеющий свои закономерности развития и свою динамику» (Догель, 1962, с. 227). Следовательно, во-вторых, к этим объектам применимы экологические методы количественного описания биоразнообразия свободно живущих организмов. В‑третьих, для объяснения картины паразитарного разнообразия нами были привлечены экологические факторы. Особенность исследования фауны паразитов состоит в том, что в отличие от прочих организмов, для которых единицей оценки биоразнообразия является площадь местообитания, имеющая размер, зависящий от воли и возможностей исследователя, для сообществ паразитов минимальная единица биоразнообразия дискретна и четко определена – это организм одной особи хозяина (в нашем случае – одной рыбы). В этом контексте исследование причин изменчивости видового богатства паразитофауны определенного водоема следует начинать с анализа процесса формирования инфрасообщества (становления биоразнообразия «своеобразного биоценоза» паразитов одной особи хозяина). При таком подходе реализуется важный аспект изучения динамики видового богатства паразитов: распределение паразитофауны рассматривается на разных пространственных уровнях – не только на уровне популяции хозяина, но и на уровне особи хозяина, поскольку хозяин для паразита есть не просто среда обитания, но и территория, определяющая границы биоценоза (инфрасообщества), а значит, и паразитофауны (компонентного сообщества). Выполненное исследование показало, что высокое богатство паразитофауны разных местообитаний связано с более высоким количеством видов паразитов обнаруженных на отдельных особях хозяев (щука оз. Нюк). Кривые наполнения видового списка паразитов по мере роста числа исследованных особей будут существенно отличаться для выборок рыб с разным возрастным составом. При выраженном преобладании в популяции хозяина молодых особей с малым числом видов паразитов, кривая нарастания видового богатства имеет более пологий характер, тогда как при большой доле хозяев с высоким богатством инфрасообществ, темп насыщения паразитофауны происходит более динамично. При этом группы одновозрастных животных обладают относительно сходным разнообразием паразитов и кривые насыщения видового списка, построенные для разных унивозрастных выборок, будут сходными. Выборки же с разновозрастными особями хозяев будут характеризоваться широким размахом состава индивидуальных инфрасообществ и широким варьированием траекторий кривых биоразнообразия. Это значит, что для стабилизации показателей видового богатства паразитов следует контролировать факторы формирования выборок хозяина, в частности по сезону отлова и возрастному составу (средствами планирования исследований). Еще один путь стабилизации оценок разнообразия фауны паразитов состоит в унификации методов описания показателей биоразнообразия. Однородные результаты для выборок разного объема дает определение срединных значений между крайними траекториями кумуляты видового состава и их сглаживание с помощью логарифмической функции. В данной работе нами предложен общий подход к количественному описанию накопления видового богатства паразитов, в дальнейшем планируется использование предложенного метода для исследования паразитов других хозяев.
Ряд вопросов, рассмотренных в статье, может получить решение только после привлечения более длинных и объемных рядов данных. В числе таких перспективных тем – исследование зависимости типа кривой, описывающей динамику пополнения списка паразитов, от характера распределения числа паразитов в одном хозяине.
Библиография
Быховская-Павловская И. Е. Паразиты рыб. Руководство по изучению. Л.: Наука, 1985. 121 с.
Джиллер П. Структура и экологическая ниша. М.: Мир, 1988. 184 с.
Догель В. А. Общая паразитология. Л: ЛГУ, 1962. 463 с.
Иешко Е. П., Голицына Н. Б. Анализ пространственной структуры популяции трематоды Bunodera luciopercae из окуня Perca fluviatilis. // Паразитология. 1984. Т. 18, Вып. 5. С. 374–382.
Иешко Е. П., Малахова Р. П., Голицына Н. Б. Экологические особенности формирования фауны паразитов рыб озер системы р. Каменной. // Экология паразитических организмов в биогеноценозах Севера. Петрозаводск, 1982. С. 5–25.
Коросов А. В. Специальные методы биометрии. Петрозаводск: Изд-во ПетрГУ, 2007. 364 с.
Коросов А. В., Горбач В. В. Компьютерная обработка биологических данных. Петрозаводск: Изд-во ПетрГУ, 2010. 84 с.
Мина М.В. Задачи и методы изучения роста рыб в природных условиях. // Современные проблемы ихтиологии. М.: Наука. 1981. С. 177–195.
Определитель фауны пресноводных паразитов рыб водоемов СССР. Серия определителей по фауне СССР. М.-Л.: Изд. ЗИН АН СССР, 1962. Т. 80. 776 с.
Первозванский В. Я. Рыбы водоемов района Костомукшского железорудного месторождения(экология, воспроизводство, использование). Петрозаводск, «Карелия», 1986. 216 с.
Петрушевский Г. К., Петрушевская М. Г. Достоверность Количественных показателей при изучении паразитофауны рыб. // Паразитологический сборник ЗИН РАН, 1960. Вып. 19. С. 333–343.
Правдин И. Ф. Руководство по изучению рыб. М.: Пищ. пром-сть. 1966. 376 с.
Решетников Ю. С. Экология и систематика сиговых рыб. М.: Наука. 1980. 301 с.
Чугунова Н. И. Руководство по изучению возраста и роста рыб. М.: Изд-во АН СССР. 1959. 162 с.
Шитиков В. К., Розенберг Г. С. Рандомизация и бутстреп: статистический анализ в биологии с использованием R. Тольятти, 2012. 151 с. URL: http://www.ievbras.ru/ecostat/Kiril/Article/A32/Starb.pdf (дата обращения: 21.12.2012).
Шульман С. С., Добровольский А. А. Паразитизм и смежные с ним явления. // Паразитологический сборник ЗИН АН СССР. 1977. Т. 27. С. 230–263.
Esch G. W., Bush A. O., Aho J. M. Parasite Communities: Patterns and Processes.London: Chapman & Hall, 1990. P. 197–232.
Poulin R. Evolutionary Ecology of Parasite (second edition). New Jersey08540. Princeton University Press, 2007. 332 p.
Vandermeer J. H. Niche theory // Ann. Rev. Ecol. Syst. 1972. Vol. 3. P. 107–132.
Благодарности
Исследование выполнено при поддержке Министерства образования и науки Российской Федерации, соглашение 8101.

 © 2011 - 2026
© 2011 - 2026