



Введение
Целью проведенного в 2006–2007 гг. исследования был сбор данных о видовом составе эпифитных (растущих на коре деревьев и кустарников) лишайников территорий ряда административных округов Москвы в пределах Московской кольцевой автодороги (МКАД), для сравнения их с аналогичными материалами, полученными на тех же территориях в 1987–1991 гг. (Бязров, 1994, 1996). В Москве, как и во всей стране, в течение 1990-х произошло драматическое изменение структуры промышленности, в несколько раз возросло число автомобилей. Предполагалось, что эти изменения сказались и на видовом составе эпифитных лишайников, признанных индикаторов качества воздушной среды. В данной работе сообщаются результаты обработки материалов, собранных в Южном административном округе (ЮАО).
Материалы
Обследованная территория ЮАО на юге ограничена МКАД (рис. 1), на востоке, от МКАД до Нагатинского затона, осью русла реки Москвы, далее от реки осями 2-го Южнопортового проезда, Южнопортовой, Шарикоподшипниковской улиц, на севере осями улицы 2-я Машиностроения, Волгоградского проспекта, улиц Мельникова, 1-й Дубровской, 3-го Крутицкого переулка, улиц Симоновский вал, Арбатецкой, осями русла реки Москвы и Новоспасского моста, Кожевнической, Летниковской улиц, Жукова проезда, Дубининской улицы, 4-го и 1-го Щипковских переулков, улиц Павла Андреева, Хавской, Шухова, Шаболовки, Академика Петровского, на западе осью Ленинского проспекта до дома №12, далее юго-восточной и южной границами территории Нескучного сада, далее северо-восточным границам полос отвода Малого кольца Московской железной дороги (МЖД) и III транспортного кольца, осями полосы отвода Малого кольца МЖД, долиной реки Котловка и восточной границей Битцевского леса (Закон…). Округ включает 16 муниципальных районов: Бирюлево Восточное, Бирюлево Западное, Братеево, Даниловский, Донской, Зябликово, Москворечье-Сабурово, Нагатино-Садовники, Нагатинский Затон, Нагорный, Орехово-Борисово Северное, Орехово-Борисово Южное, Царицыно, Чертаново Северное, Чертаново Центральное, Чертаново Южное.
Рис. 1. Положение ЮАО на схематической карте Москвы в пределах МКАД
Fig. 1. The position of the Southern administrative district on the sketchy map of Moscow city within the ambit of Moscow ring road.
Площадь округа около 130 км2, что составляет 12 % территории Москвы в пределах МКАД. В округе проживает более 1.5 млн человек (Портал…).
Наряду с районами весьма плотной застройки в северной части округа по обоим берегам реки Москвы, на юге территории округа нередко можно встретить пустыри, овраги, долины небольших рек, таких как Чертановка, Горобня и др., пруды. Зеленые насаждения в ЮАО занимают 17.5 % его территории. На каждого жителя округа приходится лишь 7.2 м2 зеленых насаждений (по Москве в пределах МКАД в среднем 16 м2/человека) [Доклад…, 2000–2001]. Основными зелеными массивами на территории округа являются как земли лесного фонда – Бирюлевский лесопарк, небольшая часть природного парка «Битцевский лес» и 2 квартала Видновского лесопарка, так и 40 городских парков и садов, в том числе природно-исторический парк «Царицыно» с прудами, а также музей-заповедник «Коломенское». Деревья и кустарники – субстрат эпифитных лишайников – встречаются и по берегам водоcтоков и водоемов, на склонах оврагов, на кладбищах, на территориях лечебных учреждений, стадионов, остатках промышленного сада совхоза им. Ленина. Породный состав деревьев и кустарников округа включает несколько десятков видов, но чаще встречаются тополя, липа, клены, вязы, березы, дуб, сосна, ольха, ивы, рябина, лещина, черемуха, боярышник, яблони.
Генеральным планом развития Москвы, принятым в 1971 г., обозначено наличие и развитие в Москве (в пределах МКАД) более 60 промышленных зон (ПЗ), к которым относятся части городской территории, предназначенные для размещения промышленных и автотранспортных предприятий, научно-исследовательских учреждений, коммунально-складских и других объектов (Бязров и др., 1997; Битюкова, Слободской, 2002а). Значительная их часть сосредоточена в ЮАО. По степени экологической опасности имеющиеся ПЗ города были ранжированы на несколько категорий (Битюкова, Слободской, 2002б; Битюкова, 2003). К ПЗ с высоким и исключительно высоким уровнем опасности в ЮАО отнесены «ЗИЛ», «Бирюлево», к ПЗ с повышенным уровнем экологической опасности – «Южный порт», «Варшавское шоссе» и др., к ПЗ со средним уровнем экологической опасности – «Котляково», «Нагатино», «Верхние Котлы» и др., к ПЗ со слабым уровнем экологической опасности – «Каширское шоссе», «Царицыно» и др.
Методы
Для выявления особенностей распределения эпифитных лишайников в качестве картографической основы была использована карта Москвы масштаба 1:38 000. В 1987–1991 г. территория города (в пределах МКАД) на этой карте была поделена на квадраты 1х1 км. На долю ЮАО приходится 127 таких квадратов. В каждом квадрате было осмотрено от 100 до 400 стволов деревьев и кустарников. Деревья обследовались от основания до высоты 2.5 м, как пряморастущие, так и искривленные и наклоненные. Наличие лишайников фиксировалось на всей видимой поверхности ствола. Идентификация лишайников проведена в Лаборатории радиоэкологического мониторинга в регионах АЭС и биоиндикации Института проблем экологии и эволюции РАН. Номенклатура видов в тексте в основном соответствует обобщению Г. П. Урбанавичюса (2010).
Наряду с разнообразием видов на основе уже имеющихся обобщений (Инсарова, Инсаров, 1989; Wirth, 1991, 2010) и личного опыта (Бязров, 1994, 2002, 2009) по пятибалльной шкале были оценены экологические характеристики видов: отношение вида к освещённости местообитания – С; отношение вида к величине pH субстрата – К; отношение вида к богатству субстрата элементами питания – Э; отношение вида к степени загрязнения воздушной среды территории – токсифобность (Тф); отношение вида к степени окультуренности, или гемеробность (Гм) местообитания. Гемеробность оценена на основе модифицированной мною шкалы Х. Х. Трасса (Trass, 1973) с использованием сведений B. Litterski (1999). При этом следует помнить, что виды, принадлежащие к последующей группе названной шкалы, могут встречаться в местообитаниях, характерных для предшествующих групп. Фактически это шкала перехода от стенотопных к эвритопным видам по градиенту степени освоения местообитания человеком.
Результаты
Всего в 2006–2007 гг. на территории ЮАО на стволах деревьев и кустарников до высоты 2.5 м были обнаружены представители 54 видов лишайников (табл. 1). Из перечисленных в таблице 1 видов только Evernia prunastri, слоевища которого встречены на территории Покровского кладбища, включен в Красную книгу Москвы (2011). В начале 1990-х в обследованных 127 квадратах на деревьях и кустарниках были обнаружены представители 26 видов лишайников, причем 35 % квадратов территории ЮАО тогда формировали «лишайниковую пустыню», то есть эпифитные лишайники я в них не встретил (Бязров, 2002). Результаты учетов 2006–2007 гг. показали, что в принятом масштабе учетной единицы (1 х 1 км) в ЮАО «лишайниковая пустыня» отсутствует. Число видов эпифитных лишайников с начала 1990-х на территории округа возросло в 2 раза, а в квадрате варьирует от 3 до 24 (участок леса между Гурьевским проездом и МКАД). Но четыре вида [Arthonia atra (Pers.) A. Schneid. = (Opegrapha atra Pers.), Cladonia caespitica (Pers.) Flörke, C. ochrochlora Flörke, Lecidea erythrophaea Sommerf.], зафиксированные в конце 1980-х – начале 1990-х, при учете в 2006–2007 гг. я не обнаружил. Не встретил я и представителей Phaeophyscia ciliata (Hoffm.) Moberg, ранее указанных А. В. Пчелкиным для долины р. Котловки (Лишайники..., 2012). Два вида (Phaeophyscia orbicularis и Xanthoria parietina) при последнем учете встречены во всех квадратах (прежде 36 и 4 % соответственно). Более чем в 60 % квадратов представлены Caloplaca cerina, C. holocarpa, Candelariella vitellina, Lecanora hagenii, Parmelia sulcata, Phaeophyscia nigricans, Physcia adscendens, P. stellaris, Scoliciosporum chlorococcum. Практически все эти часто встречающиеся и нередко обильные виды считаются нитрофитами (Wirth, 1991, 2010; Davies, 2007). У 8 видов в 2006–2007 гг. встречаемость была от 10 до 60 %. Почти треть видов в 2006–2007 гг. были встречены всего в 1–2 квадратах. Эти участки, возможно, при благоприятных условиях станут для них отправными точками для освоения новых территорий. По данным учетов 1987–1991 гг. только у Physcia stellaris величина встречаемости на той же территории превышала 50 %. В целом же в оба срока более половины видов относились к классу встречаемости ≤ 5%
Таблица 1. Встречаемость (% от общего числа квадратов) эпифитных лишайников в пределах ЮАО Москвы в 1987–1991 гг. (1990) и в 2006–2007 гг.(2007) и экологическая характеристика видов
| Вид | Срок | Экологическая характеристика вида* | ||||||
| 1990 | 2007 | С | К | Э | Тф | Гм | ||
| Amandinea punctata | - | 1 | С4 | К3 | Э3 | Тф2 | плгм | |
| Arthonia atra | 6 | - | С2 | К3 | Э3 | Тф4 | эвгм | |
| Arthrosporum populorum | - | 11 | С4 | К3 | Э3 | Тф3 | эвгм | |
| Biatora helvola | - | 2 | С2 | К3 | Э1 | Тф4 | мзгм | |
| Caloplaca cerina | 1 | 81 | С4 | К4 | Э3 | Тф2 | мзгм | |
| C. citrina | 1 | - | С4 | К4 | Э4 | Тф2 | эвгм | |
| C. holocarpa | - | 62 | С4 | К5 | Э3 | Тф3 | плгм | |
| Candelariella vitellina | 3 | 75 | С4 | К3 | Э3 | Тф2 | плгм | |
| C. xanthostigma | - | 8 | С4 | К3 | Э3 | Тф3 | плгм | |
| Cetraria sepincola | 1 | 1 | С4 | К2 | Э2 | Тф4 | мзгм | |
| Chrysothrix candelaris | - | 19 | С2 | К2 | Э2 | Тф4 | эвгм | |
| Cladonia caespitica | 1 | - | С3 | К3 | Э2 | Тф4 | эвгм | |
| C. chlorophaea | 1 | 3 | С3 | К2 | Э2 | Тф4 | эвгм | |
| C. coniocraea | 7 | 13 | С3 | К2 | Э2 | Тф2 | эвгм | |
| C. fimbriata | 3 | 5 | С4 | К2 | Э1 | Тф3 | эвгм | |
| C. ochrochlora | 3 | - | С3 | К1 | Э2 | Тф4 | мзгм | |
| C. parasitica | - | 1 | С3 | К2 | Э1 | Тф4 | мзгм | |
| C. ramulosa | - | 1 | С4 | К1 | Э2 | Тф4 | мзгм | |
| C. squamosa | - | 1 | С3 | К1 | Э2 | Тф3 | эвгм | |
| Evernia prunastri | - | 1 | С4 | К2 | Э3 | Тф4 | плгм | |
| Hypocenomyce scalaris | - | 4 | С3 | К1 | Э2 | Тф3 | плгм | |
| Hypogymnia physodes | 11 | 21 | С4 | К2 | Э2 | Тф3 | плгм | |
| Lecania naegelii | - | 3 | С3 | К3 | Э3 | Тф3 | плгм | |
| Lecanora albellula | - | 5 | С4 | К1 | Э2 | Тф3 | плгм | |
| L. argentata | - | 5 | С3 | К3 | Э2 | Тф4 | плгм | |
| L. hagenii | 9 | 72 | С3 | К5 | Э4 | Тф1 | плгм | |
| L. populicola | - | 2 | С4 | К2 | Э2 | Тф2 | плгм | |
| L. pulicaris | - | 2 | С4 | К1 | Э2 | Тф3 | мзгм | |
| L. symmicta | - | 2 | С4 | К3 | Э3 | Тф4 | мзгм | |
| L. varia | 9 | 44 | С4 | К2 | Э2 | Тф2 | плгм | |
| Lecidea erythrophaea | 3 | - | С4 | К3 | Э1 | Тф4 | мзгм | |
| Lepraria cacuminum | - | 2 | С2 | К2 | Э2 | Тф4 | мзгм | |
| L. incana | 6 | 6 | С2 | К2 | Э2 | Тф1 | плгм | |
| L. lobificans | - | 2 | С2 | К2 | Э2 | Тф4 | эвгм | |
| Melanelia exasperatula | 1 | 3 | С4 | К3 | Э3 | Тф3 | плгм | |
| M. olivacea | - | 1 | С4 | К1 | Э2 | Тф3 | плгм | |
| M. subargentifera | - | 1 | С4 | К4 | Э4 | Тф4 | эвгм | |
| Micarea lignaria. | - | 2 | С3 | К2 | Э2 | Тф2 | эвгм | |
| Parmelia sulcata | 22 | 68 | С4 | К3 | Э3 | Тф2 | плгм | |
| Phaeophyscia nigricans | 2 | 72 | С4 | К5 | Э4 | Тф2 | мтгм | |
| P. orbicularis | 36 | 100 | С4 | К4 | Э4 | Тф1 | мтгм | |
| P. sciastra | - | 10 | С4 | К5 | Э4 | Тф2 | плгм | |
| Physcia adscendens | - | 86 | С4 | К4 | Э4 | Тф2 | плгм | |
| P. dubia | - | 7 | С4 | К3 | Э3 | Тф2 | плгм | |
| P. stellaris | 53 | 96 | С4 | К3 | Э3 | Тф1 | эвгм | |
| P. tenella | - | 17 | С4 | К3 | Э4 | Тф2 | плгм | |
| P. tribacia | 1 | 11 | С4 | К3 | Э3 | Тф2 | плгм | |
| ?P. vitii | - | 1 | С3 | К3 | Э3 | Тф3 | плгм | |
| Physconia detersa | - | 4 | С4 | К4 | Э4 | Тф3 | плгм | |
| P. distorta | - | 11 | С4 | К4 | Э4 | Тф3 | плгм | |
| P. grisea | 1 | 1 | С4 | К4 | Э4 | Тф2 | плгм | |
| Rinodina archaea | - | 1 | С4 | К4 | Э3 | Тф3 | плгм | |
| R. pyrina | - | 6 | С4 | К4 | Э3 | Тф3 | плгм | |
| Scoliciosporum chlorococcum | 35 | 98 | С3 | К2 | Э3 | Тф1 | плгм | |
| Strangospora pinicola | - | 2 | С4 | К2 | Э3 | Тф1 | плгм | |
| Trapeliopsis granulosa | - | 1 | С4 | К2 | Э2 | Тф1 | эвгм | |
| Vulpicida pinastri | 2 | - | С3 | К1 | Э1 | Тф3 | эвгм | |
| Xanthoria candelaria | - | 7 | С4 | К3 | Э4 | Тф3 | плгм | |
| X. parietina | 4 | 100 | С4 | К4 | Э4 | Тф1 | плгм | |
| X. polycarpa | 1 | 25 | С4 | К2 | Э4 | Тф2 | плгм | |
| Встречаемость всех видов | 65 | 100 | ||||||
| Число видов | 26 | 54 | ||||||
*- пояснение обозначений для табл. 1–5: С – отношение вида к освещённости местообитания (С2 – затененные местообитания, освещённость 5–10 % ; С3 – полузатененные местообитания, освещенность 10–20 %; С4 – полуосвещённые и освещённые местообитания, освещённость до 40 %); К – отношение вида к величине pH субстрата (К1 – субстрат экстремально кислый и очень кислый, pH6.5); Э – отношение вида к богатству субстрата элементами питания (Э1 – очень бедная минеральными элементами (Mg, Ca, K, Na, N) кора деревьев как у ели, лиственницы, березы; Э2 – умеренно богатая минеральными элементами кора деревьев с очень малой эвтрофикацией; Э3 – кора богата минеральными элементами или умеренно покрыта пылью; Э4 – богатая минеральными элементами кора, часто покрыта густам слоем пыли); Тф – отношение вида к степени загрязнения воздушной среды территории – токсифобность (Тф1 – наиболее устойчивые к загрязнению воздуха виды, Тф5 – наиболее чувствительные к загрязнению виды); Гм (гемеробность местообитания) – отношение вида к степени окультуренности (мезогемеротоп (мзгм) – вид, встречающийся на естественных субстратах также и в экосистемах, незначительно измененных человеком; эвгемеротоп (эвгм) – вид, встречающийся на естественных субстратах также и в экосистемах с умеренным или периодическим воздействием человека; полигемеротоп (плгм) – вид, встречающийся на естественных и искусственных субстратах также и в экосистемах со значительным воздействием человека; метагемеротоп (мтгм) – вид, также встречающийся на субстратах и в экосистемах, полностью созданных человеком).
Среди видов, встреченных на территории округа в оба сравниваемых срока, количественно преобладают лишайники, предпочитающие полуосвещённые и освещённые местообитания (табл. 2), а также предпочитающие довольно кислый (pH 4.1–4.8) и умеренно кислый (pH 4.9–5.6) субстрат (табл. 3). Однако виды, имеющие высокие показатели встречаемости (Candelariella vitellina, Lecanora hagenii, Parmelia sulcata, Phaeophyscia nigricans, P. orbicularis, Physcia adscendens, P. stellaris, Xanthoria parietina), чаще предпочитают полунейтральный или нейтральный субстраты.
Таблица 2. Распределение числа видов (%) эпифитных лишайников ЮАО по отношению к освещённости местообитания в 1987–1991 (1990) и 2006–2007 (2007) гг.
| Срок | Класс величины освещённости местообитания (С) | ||
| С2 | С3 | С4 | |
| 1990 | 7 | 30 | 63 |
| 2007 | 9 | 22 | 69 |
Таблица 3. Распределение числа видов (%) эпифитных лишайников ЮАО по отношению к величине pH субстрата в 1987–1991 (1990) и 2006–2007 (2007) гг.
| Срок | Класс величины pH субстрата (К) | ||||
| К1 | К2 | К3 | К4 | К5 | |
| 1990 | 7 | 33 | 30 | 19 | 11 |
| 2007 | 11 | 33 | 30 | 19 | 7 |
Среди эпифитных лишайников округа по числу видов преобладают такие, которые селятся на умеренно богатой минеральными элементами коре деревьев с очень малой эвтрофикацией или коре, богатой минеральными элементами или умеренно покрытой пылью (табл. 4). Однако виды с высокой частотой встречаемости, такие как Lecanora hagenii, Phaeophyscia nigricans, P. orbicularis, Physcia adscendens, Xanthoria parietina характеризуют как предпочитающие богатые элементами питания субстраты (Wirth, 1991, 2010).
Таблица 4. Распределение числа видов (%) эпифитных лишайников ЮАО по отношению к богатству субстрата элементами питания в 1987–1991 (1990) и 2006–2007 (2007) гг.
| Срок | Класс богатства субстрата (Э) | |||
| Э1 | Э2 | Э3 | Э4 | |
| 1990 | 11 | 29 | 30 | 30 |
| 2007 | 6 | 35 | 35 | 24 |
Сравнение видов по степени устойчивости к загрязнению (табл. 5) показывает, что за период с 1987–1991 гг. по 2006–2007 гг. произошло заметное увеличение доли видов, относительно более чувствительных к загрязнению воздуха (класс Тф3) и уменьшение доли относительно устойчивых и толерантных к загрязнению видов (классы Тф1 и Тф2).
Таблица 5. Распределение числа видов (%) эпифитных лишайников ЮАО по классам токсифобности (Тф) в 1987–1991 (1990) и 2006–2007 (2007) гг.
| Срок | Класс токсифобности (Тф) | |||
| Тф1 | Тф2 | Тф3 | Тф4 | |
| 1990 | 26 | 37 | 15 | 22 |
| 2007 | 15 | 30 | 33 | 22 |
Ранее по результатам учета особенностей распределения эпифитных лишайников в 1987–1991 гг. по числу видов в квадрате было проведено ранжирование территории города по степени загрязнения атмосферного воздуха. Это ранжирование опубликовано (Бязров, 1996, 2002). Аналогичный способ (по числу видов в квадрате) был применен для графического показа результатов, полученных в 2006–2007 гг. на территории ЮАО. Группирование квадратов было аналогично тому, которое было использовано в начале 1990-х. Прежним был и подход к заливке квадратов – от темного цвета к светлому, что соответствовало переходу от неблагоприятных условий жизни для лишайников к более благоприятным. Сравнение с вырезкой тех же участков из соответствующей картосхемы начала 1990-х (Бязров, 1996) показывает, что на территории округа в 2006–2007 гг. ситуация для лишайников кардинально улучшилась (рис. 2).
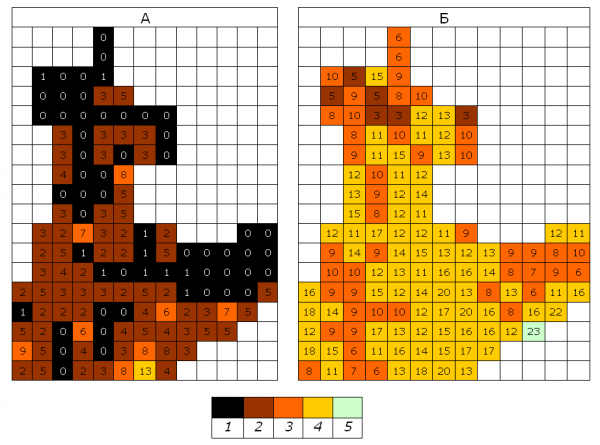
Рис. 2. Ранжирование территории ЮАО по числу видов лишайников в квадрате 1х1 км в 1987–1991 гг. (А) и в 2006–2007 гг. (Б). 1 – 0–1 видов в квадрате, 2 – 2–5 видов в квадрате, 3 – 6–10 видов в квадрате, 4 – 11–20 видов в квадрате, 5 – более 20 видов в квадрате
Fig. 2. Ranking the territory of Southern adminictrative district on the number of lichen species in a square 1x1 km in 1987–1991 yy (A) and in 2006–2007 yy (Б). 1 – 0–1 species in a square, 2 – 2–5 species in a square, 3 – 6–10 species in a square, 4 – 11–20 species in a square, 5 – more than 20 species in a square
Таблица 6. Количественные показатели выделов легенд картосхем для обследованной в 1987–1991 гг. (1990) и в 2006–2007 гг. (2007) территории ЮАО
| Выдел легенды по числу видов в квадрате | 1990 | 2007 | ||
| Число квадратов | % | Число квадратов | % | |
| 0–1 | 57 | 45 | 0 | 0 |
| 2–5 | 61 | 48 | 6 | 5 |
| 6–10 | 8 | 6 | 44 | 34 |
| 11–20 | 1 | 1 | 76 | 60 |
| >20 | 0 | 0 | 1 | 1 |
| Итого | 127 | 100 | 127 | 100 |
Исчез выдел легенды «лишайниковая пустыня», но появился выдел «>20 видов». На порядок уменьшилась доля выдела «2–5 видов», более чем в 5 раз увеличилась доля выдела «6–10 видов» и многократно – доля выдела «11–20 видов» (табл. 6). К 2006–2007 гг. число видов эпифитных лишайников увеличилось во всех обследованных квадратах как ближе к центральной части города, так и в периферийных частях округа. Это позволяет сделать вывод, что в сравнении с началом 1990-х условия для жизнедеятельности ряда видов лишайников улучшились. Это выражается не только в увеличении числа видов на единицу площади, но и в повышении частоты встречаемости на учетных площадях представителей Caloplaca cerina, Candelariella vitellina, Phaeophyscia orbicularis, Physcia stellaris, Scoliciosporum chlorococcum, Xanthoria parietina и некоторых других видов, причем слоевища Phaeophyscia orbicularis нередко покрывали значительные поверхности стволов деревьев.
Обсуждение
Установленные изменения видового состава лишайников и формируемых ими группировок, произошедшие за известный промежуток времени, несомненно, являются следствием как природных сукцессий, так и экзогенных смен (Работнов, 1983). И надо четко осознавать, что наблюдения за динамикой видового состава лишайников при мониторинге среды обеспечивают скорее корреляции, чем доказательства причинных связей, за исключением случаев, когда эти наблюдения являются частью контролируемых воспроизводимых экспериментов, при которых воссоздают те или иные параметры качества воздуха. Но опытов таких немного, и мы вынуждены делать выводы, основываясь на опубликованных сведениях о реакции разных видов лишайников на загрязнители воздуха и собственном опыте. Эти выводы должны помочь изолировать из общей суммы выявленных изменений те, которые связаны с действием загрязнителей. Арсенал способов такой изоляции невелик, и они основаны главным образом на знаниях экологии видов лишайников. Так, известно, что со временем происходит изменение свойств субстрата, например коры липы, в молодом возрасте гладкой, и бороздчатой – в зрелом. Соответственно, со временем меняется и состав лишайников, обитающих на этой породе.
Следует также учитывать, что городская среда в значительной степени естественной не является, она изменена, а во многих случаях создана человеком, особенно в мегаполисах, к которым принадлежит и Москва. Здесь зачастую трудно отделить природное от антропогенного. Например, деревья, используемые для озеленения. С одной стороны, это представители естественных видов древесных пород. С другой – во многих случаях их выращивали в питомниках, а затем высаживали в городе.
За промежуток времени между двумя сроками наблюдения молодые деревца, главным образом липы и клены, высаженные в конце 1980-х в только что застроенных жилых массивах на периферии города, превратились в деревья. Кора в нижних частях стволов на них из гладкой стала трещиновато-бороздчатой. Соответственно, вероятность закрепления на таком субстрате попадающих сюда диаспор лишайников увеличилась. И если в 1987–1991 гг. на ряде учетных площадей 1 х 1 км в этих районах лишайники отсутствовали или были представлены лишь одним видом, хотя здесь не было крупных источников загрязнения, то в 2006–2007 гг. в этих районах на тех же деревьях лишайники встречались нередко. Чаще других отмечали наличие слоевищ Phaeophyscia orbicularis, Physcia adscendens, P. stellaris, Scoliciosporum chlorococcum, Xanthoria parietina. Бедность лихенобиоты недавно застроенных городских территорий наблюдали и в других городах, например в Петербурге (Малышева, 2006).
В июне 2001 г. в результате шквалистых ветров на территории Москвы погибло несколько десятков тысяч деревьев, а значительно большее их число пострадало от ветролома. В основном это были 40–50-летние тополя, березы, клены. Вместе с ними могли исчезнуть и представители видов лишайников, некогда поселившихся на их стволах, такие как Cladonia caespitica, Phaeophyscia ciliata, которые не были встречены в 2006–2007 гг. (табл. 1). С другой стороны, в сомкнутых лесных массивах в результате этого урагана образовались обширные участки с разреженным древостоем, что привело к изменению светового режима и появлению слоевищ фотофильных видов лишайников на сохранившихся в таких местах деревьях. Или же виды, предпочитающие более светлые местообитания, стали встречаться здесь чаще. Это представители родов Caloplaca, Candelariella, Melanelia, Phaeophyscia, Physcia, Physconia, Xanthoria и др. Стволы некоторых деревьев в результате урагана изменили вертикальное положение на наклонное. Это привело к появлению на них слоевищ видов рода Cladonia и некоторых других.
Однако наиболее значительное влияние на современный видовой состав лихенобиоты в Москве оказали социально-экономические факторы. Результаты обследования лишайников, проведенного в конце 1980-х, отражают состояние лихенобиоты перед катастрофическим экономическим спадом начала 1990-х годов. Многие промышленные предприятия, относившиеся к категории стационарных источников загрязнения воздушного бассейна города, в середине 1990-х либо прекратили свою деятельность, либо значительно снизили объемы производства. Уже к 1993 г., в сравнении с 1990 г., объем валовых выбросов в Москве сократился на 4 %, в т. ч. твердых взвешенных веществ – на 19 %, а оксида углерода – на 10 % (Резер, Упелли, 1995). В течение 1990-х годов ряд предприятий был выведен за черту города, а на объектах теплоэнергетики структура используемого топлива изменилась в пользу природного газа, что привело к значительному снижению объема выбросов от стационарных источников – со 177 тыс. т в 1996 г. до 93.1 тыс. т в 2002 г. (Битюкова, Слободской, 2002а, 2002б; Василенко и др., 2004). В результате спада активности промышленных предприятий города и соседних регионов, а также улучшения качества воздуха в странах Западной Европы, откуда в основном поступают к нам воздушные массы вследствие трансграничного переноса, в Москве кислотность дождевой воды уменьшилась – если в 1987 г. среднегодовая величина pH осадков была 4.2, то в 2002 г. – 6.25, самая высокая за весь период измерения кислотности осадков (1980–2002 гг.) на Метеорологической обсерватории МГУ (Еремина, 2004). Более того, в 2002 г. кислые дожди (pHДо 1991–1992 гг. минерализация осадков возрастала, и в 1991 г. среднегодовая величина была 27.0 мг/л, а концентрация сульфатов в них - 10.9 мг/л. В 2000–2001 гг. среднее значение минерализации осадков составило 11.8 мг/л, а сульфатов – 2.6 мг/л (Еремина, 2004).
Одновременно в городе стало увеличиваться число автомобилей и в настоящее время оно превышает 3 млн. Соответственно, значительно изменилось соотношение между количеством выбросов от стационарных и передвижных источников загрязнения – доля первых неуклонно снижалась с 41% в 1986 г. до 6–8 % в 2001–2004 гг. (Москва в цифрах, 1987; Государственный доклад…, 2004) Изменилась и структура выбрасываемых загрязняющих веществ. Если в 1980-х среди загрязнителей преобладали окиси углерода (60 % от суммарных выбросов), оксиды азота (14%), углеводороды (13 %), двуокись серы (9 %), то в настоящее время атмосферный воздух города наиболее загрязнен оксидами азота, бенз(а)пиреном, аммиаком, формальдегидом. Средняя за год концентрация диоксида азота в целом по городу в 2003 г. составляла 1.6 ПДК, бенз(а)пирена – 2.8 ПДК, фенола – 1.3 ПДК, формальдегида – 2.3 ПДК (Государственный доклад…, 2004). Годовой ход примесей в воздухе характеризуется летним максимумом аммиака и формальдегида и весенне-осенним максимумом диоксида и оксида азота, при этом концентрация азота составляет 2.8–3.5 ПДК, а концентрация оксида углерода ПДК не превышает. Отмечена тенденция к заметному росту концентраций углеводородов, диоксида и оксида азота, аммиака и хлористого водорода. Рост концентраций по первым трем показателям связан с выбросами от автотранспорта. Таким образом, среди загрязнителей уменьшилось количество двуокиси серы, но значительно увеличилось количество выбрасываемых в атмосферу города соединений азота в форме аммиака и оксидов азота. Однако обсуждать азот и его соединения как загрязнители среды очень трудно, поскольку этот элемент является главным компонентом атмосферы (более ¾ ее объема). Он входит в состав всех органических веществ, участвует во всех жизненно важных процессах. Молекулярный азот атмосферы способны фиксировать и превращать его в форму, пригодную для использования, лишь лишайники, фотобионтом которых является цианобактерия. Такие виды на деревьях в Москве пока не зафиксированы. Однако известно, что оптимум фиксации азота у этих видов наблюдается при величине рН > 5, т. е. в нейтральной или щелочной среде (Шапиро, 1986, 1996).Один из источников азота аммоний NH4+ поглощается лишайниками пассивно (Dahlman et al., 2004), но в относительно больших количествах, поскольку этот катион притягивается отрицательно заряженными стенками клеток (Brown et al., 1994). Однако высокие концентрации аммония могут и подавлять развитие даже нитрофитов. Так, эксперимент с Xanthoria parietina, обычным в Москве нитрофитным лишайником, показал, что длительная обработка слоевищ этого вида хлористым аммонием NH4Cl в концентрации 0.69М приводила к необратимым отрицательным изменениям как микобионта, так и фотобионта. При концентрации 0.35М NH4Cl сначала происходило подавление активности обоих компонентов, но затем наблюдали восстановление до прежнего уровня деятельности как гриба, так и зеленой водоросли, причем восстановление активности водоросли происходило быстрее. Это дало основание авторам исследования сделать вывод, что фотобионт этого лишайника лучше адаптирован к высоким уровням азота в среде (Gaio-Oliveira et al., 2004).
Как производные азота в качестве первичных загрязнителей в первую очередь считают аммиак и окись азота. Аммиак нейтрализует SO2 в форме SO4 2- частично в NH4HSO4 или полностью в (NH4)2SO4, а окись азота легко окисляется озоном в NO2 (Krupa, 2003). Далее из продуктов окисления оксидов азота (NOx) атмосферы образуются вторичные загрязнители и химические соединения, в газообразном или аэрозольном состоянии. Аммиак NH3 образуется вследствие естественных процессов распада останков растений и животных, продуктов их жизнедеятельности. Его обмен с атмосферой аналогичен обмену углекислого газа CO2, в котором точка компенсации определяет концентрацию, при которой чистый обмен не происходит. Эта точка компенсации в условиях современного земледелия и животноводства превышена, и аммиак осаждается и накапливается (Krupa, 2003). Поскольку проводятся мероприятия по снижению выбросов SO2 и NOX в атмосферу, аммиак NH3 становится важным агентом эвтрофикации среды, а в действительности её гипертрофикации (Paoli et al., 2010). В настоящее время именно последний термин более адекватно описывает процессы обогащения местообитаний лишайников элементами питания (Seaward, 2004; Hauck, 2011). В Великобритании наблюдали, что величины покрытия субстрата нитрофитными видами положительно коррелируют с величинами концентрации NO2 в воздухе, тогда как корреляция между величинами покрытия всех видов с величинами концентрации NO2 в воздухе отрицательная, что свидетельствует о наличии в составе лишайниковых группировок нитрофобных видов (Gadsdon et al., 2010).
Увеличение трофности местообитаний лишайников в Москве происходит за счет местных источников (выбросы автотранспорта, дорожная и строительная пыль и др.). Так, двигатели современных автомобилей выбрасывают много органических соединений, таких как бензол, толуол, фенол. О локальности источников эвтрофикации среды свидетельствуют и данные об уменьшении таких концентраций соединений азота как аммоний NH4+или нитрат NO3- в дождевых осадках. Например, в 1992 г. концентрация аммония в осадках была 1.69 мг/л, а в 2001 г. – 0.45 мг/л (Еремина, 2004). В результате снижения порогового значения кислого загрязнения местообитаний и увеличения доли щелочного на территории города восстановились представители сравнительно большого числа видов, приспособленных к умеренно кислому субстрату, а также произошла экспансия по всей территории нитрофитных видов (Caloplaca cerina, C. holocarpa, Lecanora hagenii, Phaeophyscia orbicularis, Physcia adscendens, P. stellaris, Xanthoria parietina и др.). Экспансия отмечена и у Scoliciosporum chlorococcum, вида, считавшегося токситолерантным ацидофитом (Ahti, Vitikainen, 1974; Wirth, 1991), который и ранее встречалcя в городе нередко. Возможно, следует пересмотреть восприятие этого вида как ацидофитного, поскольку и в Великобритании его уже относят к нитрофитным (Wolseley, James, 2002; Davies et al., 2007). Увеличилась встречаемость и видов Hypogymnia physodes (К2), Lecanora varia (К2), Lepraria incana (К2) и др., обычных для довольно кислого субстрата. Вероятно, для этих видов ранее существовал пороговый уровень кислого загрязнения, который понизился, что и позволило им появиться в ряде ранее не заселенных ими участков города.
Выявленные в ЮАО изменения соответствуют тенденции поведения лишайников на всей территории города в пределах МКАД. Это показало как выборочное обследование с использованием нескольких трансект шириной 2 км, пересекающих город с севера на юг и с востока на запад (Бязров, 2009), так и обследование аналогичным с ЮАО методом всей территории Северного округа Москвы (Biazrov, 2010). Подобное наблюдается и в крупных городах стран Западной Европы с конца 1970-х – начала 1980-х (Kandler, Poelt, 1984; Hawksworth, McManus, 1989). Увеличение числа видов лишайников в мегаполисах ряда стран Европы стало следствием принятых в этих государствах в 1960-е годы законов о чистоте воздуха. Наблюдалось возвращение в состав местных лихенобиот некоторых чувствительных к диоксиду серы видов в связи с уменьшением загрязнения атмосферы этим соединением. Позднее обнаружили, что значительную роль в составе образующихся биот, причем не только городских, стали играть нитрофитные виды лишайников (van Dobben, 1993; van Herk, 1999; Purvis et al., 2003; Wolseley et al., 2006; Davies et al., 2007; Isocrono et al., 2007). Экспансию нитрофитных видов эпифитных лишайников связывают c уменьшением уровней двуокиси серы в приземном слое воздуха и с высокими значениями pH коры деревьев из-за загрязнения пылью различного происхождения (придорожная, строительная, угольная), золой и сажей, аммиаком, оксидами азота (van Dobben, de Bakker, 1996; van Herk et al., 2003; Larsen et al., 2007). Например, в двух городах земли Гессен (Германия) с 1985 по 2005 г. концентрация двуокиси серы в воздухе снизилась почти на порядок, а величина pH коры липы увеличилась с 2.9–3.1 в 1985 г. до 5.9–6.3 в 2005 г., у тополя – с 4.6–3.8 до 5.6–5.7. Число видов эпифитных лишайников в этих городах за тот же период увеличилось с 11–21 до 49–52 (Kirschbaum et al., 2006). В Лондоне (Великобритания) в 1960-х на всех деревьях было зафиксировано 32 вида лишайников, тогда как в 2002–2003 гг. только на ясене (Fraxinus excelsior) обнаружили представителей 74 видов (Davies et al., 2007). Широкое распространение и обилие лишайников-нитрофитов объясняют и глобальным потеплением (van Herk et al., 2002).
Экспансию ряда нитрофитных видов (Phaeophyscia orbicularis, Physcia adscendens и др.) можно объяснить и тем, что у этих видов превалирует вегетативное размножение. Наиболее часто встречающийся вид Phaeophyscia orbicularis, индикатор гипертрофикации среды в Западной Европе (Seaward, 2004), образует огромное число соредий, иногда покрывающих всю поверхность слоевища. Эти вегетативные пропагулы легко отделяются от слоевища и разносятся ветром или стекающей по стволу водой осадков. Попадая в неровности коры или приклеиваясь к слою пыли, они быстро превращаются в талломы. Нередко на одном стволе дерева можно насчитать десятки и сотни слоевищ этого вида. Часто они срастаются, и тогда практически невозможно отделить одно слоевище от другого. В начале 1990-х годов этот вид также показывал относительно высокую встречаемость на территории округа (36 %), что свидетельствовало о высокой трофности местообитаний. Но в те годы в пределах этих квадратов его слоевища встречались много реже, и покрытие ими стволов деревьев было небольшим. В 2006–2007 гг. встречаемость вида на учетной площади была высокой, а покрытие стволов деревьев до высоты 2 м иногда достигало 30 %. Увеличилось присутствие в городе и Parmelia sulcata, вида, отличающегося широкой экологической амплитудой, но также распространяющегося соредиями. Однако повысилась частота встречаемости и видов, у которых соредий и изидий нет, а распространяются они спорами. Это представители Caloplaca, Candelariella, Lecanora, Physcia stellaris, Xanthoria parietina и др. Возможно, это связано с тем, что их микобионты могут образовать слоевище с представителями разных видов зеленых водорослей. Например, гриб Xanthoria parietina для формирования слоевища использует представителей трех видов зеленой водоросли Trebouxia – T. arboricola, T. decolorans, T. irregularis (Голубкова, 1993). Также установлено, что этот вид адаптирован как к высоким, так и низким концентрациям соединений азота. В Португалии концентрация азота в слоевищах этого вида, собранных в 13 различных местообитаниях, варьировала от 11 до 43 мг/г сухого веса талломов (Gaio-Oliveira et al., 2005), тогда как обычно у лишайников с фотобионтной зеленой водорослью этот показатель составляет Caloplaca, Xanthoria содержат пигмент париетин, наличием которого объясняют яркий желто-оранжевый цвет их слоевищ. Однако это сложное соединение включает в себя антраквиноны, активные антиокислители, нейтрализующие свободные радикалы, образующиеся в слоевище при воздействии загрязнителей (Silberstein et al., 1996; Sochting, Lutzoni, 2003). Таким образом, в Москве наличие у лишайников пропагул для вегетативного размножения не дает особого преимущества для заселения новых участков над видами, у которых такие приспособления отсутствуют. Это наблюдали и в Лондоне (Larsen et al., 2007). Однако первые, как правило, в пределах учетных площадей встречаются чаще и покрывают более значительные поверхности субстрата. Возможно, это связано с особенностями распространения и закрепления диаспор лишайников. Они зависят как от количества и качества продуцируемых пропагул, так и пригодности для закрепления мест, куда эти пропагулы попадают. Для эпифитного вида Lobaria pulmonaria с вегетативными пропагулами (соредии) в лесах Швеции было установлено, что для его распространения на большие расстояния (до 75 м) имеет значение количество продуцируемых диаспор, а для закрепления вблизи материнского слоевища важно наличие подходящих условий на субстрате, куда они попали (Öckinger et al., 2005). Проецируя эти данные на результаты, полученные в Москве, можно предполагать, что повсеместное и нередко обильное присутствие слоевищ Phaeophyscia orbicularis на деревьях города связано с тем, что этот вид продуцирует громадное количество вегетативных пропагул, разносимых ветром и водой на многие десятки метров. Слоевища Xanthoria parietina, как правило, фертильные, т. е. с апотециями, из которых время от времени выбрасываются споры. Но их число значительно уступает количеству диаспор P. orbicularis. Кроме того, в местообитаниях, куда эти споры попали, должны находиться подходящие виды зеленой водоросли, чтобы сформировалось слоевище. Поэтому обилие X. parietina в Москве сравнительно невелико.
Ранее отмечалось увеличение встречаемости в городе за сравниваемый период представителей высокоустойчивого к загрязнению вида, считавшегося ацидофитом, Scoliciosporum chlorococcum – в 2006–2007 гг. он был отмечен практически во всех обследованных в округе квадратах 1 х 1 км (встречаемость 98 %), тогда как в 1987–1991 гг. встречаемость этого вида на той же территории была 35 % (табл. 1). В городах Западной Европы с уменьшением уровней кислого загрязнения и увеличением щелочного у аналогичного по устойчивости к загрязнению ацидофитного вида Lecanora conizaeoides отмечали его отступление (Bates et al., 2001; Purvis et al., 2003; Kirschbaum et al., 2006). В ЮАО, как и в Москве, Scoliciosporum chlorococcum значительно расширил свое присутствие, что ставит под сомнение принадлежность его к группе ацидофитов. Но, возможно, объемы кислого загрязнения все-таки ещё велики, да и прошло не так много времени, чтобы нитрофитные виды вытеснили этот лишайник.
Задают вопрос, могут ли лишайники в новых условиях выступать в качестве надежных индикаторов загрязнения атмосферы? Или по ним можно будет определять степень трофности местообитаний? Современные и будущие исследования (Hauck, 2010; Davies et al., 2011; Grandin, 2011; Pinho et al., 2011; Stevens et al., 2012 и др.), возможно, дадут ответы на эти вопросы. Автор в своих предшествующих публикациях (Бязров, 1996, 2002, 2005, 2009) призывал не переоценивать роль лишайников как биоиндикаторов состояния среды. Эмпирически установленные индексы и корреляции с концентрациями ряда ксенобиотиков имеют локальное значение, и закономерности, установленные в Англии или Альпах, не всегда можно экстраполировать на европейскую часть России. Мало еще известно о действии одних фитотоксикантов при наличии в среде других или о трансформации токсикантов под воздействием факторов среды. Например, выбрасываемые органические соединения могут образовать 16–20 продуктов трансформации, среди которых могут быть более токсичные и опасные, чем исходное вещество (Малышева, 1997). Скажем, двигатели современных автомобилей выбрасывают много органических соединений,таких как бензол, толуол, фенол, которые в результате активных химических взаимодействий могут быстро превратиться в вещества, более токсичные, чем исходные. Поэтому результаты экспериментов в контролируемых условиях лабораторий не всегда отражают процессы, происходящие в конкретных местообитаниях лишайников. В общем, при планировании, проведении и обобщении материалов по использованию лишайников как биоиндикаторов уместен умеренный скептицизм и реальная оценка их качеств
Заключение
Предшествующий период использования лишайников как индикаторов загрязнения воздуха в городах и промышленных областях проходил в условиях кислого загрязнения среды, главным образом двуокисью серы, на которую лишайники реагировали довольно чутко. Ныне доля SO2 в загрязнении снизилась. И результаты проведённого мною в разные годы на территории ЮАО сравнительного учёта видового состава эпифитных лишайников подтвердили значение лишайников как относительно чутких индикаторов загрязнения воздушного бассейна, ибо уменьшение доли кислого загрязнения привело к значительному увеличению как видового разнообразия эпифитов, так и частоты встречаемости ряда видов, в особенности нитрофитов. Последнее свидетельствует об увеличении количества поступающих в окружающую среду соединений азота, что и привело к существенным изменениям видового разнообразия и структуры лишайниковых группировок как в локальном или региональном, так и в глобальном масштабах (Bobbinik et al., 2010; Bell et al., 2011; De Schrijver, 2011; Pardo et al., 2011; Phoenix et al., 2012). Судя по результатам проведённого на территории ЮАО, а ранее и Северного округа (Biazrov, 2010), исследования, в Москве пока происходит как увеличение числа видов эпифитных лишайников, так и разрастание нескольких нитрофитных видов. Можно предполагать, что при наблюдаемых ныне в городе темпах и характере загрязнения воздушного бассейна , ведущего к гиперэвтрофикации местообитаний, через некоторый промежуток времени (5–10 лет) процесс увеличения числа видов эпифитных лишайников замедлится и прекратится, а затем начнется снижение числа видов из-за разрастания нескольких нитрофитных видов, главным образом размножающихся вегетативными пропагулами представителей семейства Physciaceae.
Библиография
Битюкова В. Р. Экологическая ситуация в Москве в конце XX в. // Экология и промышленность России. 2002б. № 11. С. 29–34.
Бязров Л. Г. Видовой состав и распределение эпифитных лишайников в лесных насаждениях Москвы. // Лесоведение. 1994. № 1. С. 45–54.
Бязров Л. Г. Исчезновение лишайников – сигнал опасности. // Наука в России. 1996. № 4. С. 64–68.
Бязров Л. Г. Лишайники в экологическом мониторинге. . М.: Научный мир, 2002. 336 с.
Бязров Л. Г. Лишайники – индикаторы радиоактивного загрязнения. . М.: КМК, 2005. 476 с.
Бязров Л. Г. Эпифитные лишайники г. Москвы: современная динамика видового разнообразия. . М.: КМК, 2009. 146 с.
Бязров Л. Г., Максимова В. Ф., Рухадзе Е. В. Использование эпифитных лишайников для индикации степени загрязнения воздущной среды Москвы. // Экология и промышленность России. 2004. № 8. С. 4–7.
Голубкова Н. С. К вопросу о происхождении и путях эволюции лишайникового симбиоза // Новости систематики низших растений. СПб.: Наука. 1993. Т. 29. С. 84–104.
Государственный доклад «О состоянии и об охране окружающей среды в Российской Федерации в 2005 году». М.: Мин.природн. ресурсов РФ. 2006. 499 с.
Доклад «о состоянии окружающей среды в г. Москве в 2000-2001 гг». // Сайт ГПБУ «Мосэкомониторинг URL: www.mosecom.ru/reports / (дата обращения 15.09.2012).
Еремина И. Д. Многолетние наблюдения за химическим составом атмосферных осадков. // Вестник Моск. университета. Сер. 5, География. 2004. № 2. С. 21–26.
«Закон о территориальном делении города Москвы (Редакция на 22.06.2005)». // Сайт «Референт-информер». URL: http://www.referent.ru/3/43748 (дата обращения 12.08.2012).
Инсарова И. Д., Инсаров Г. Э. Сравнительные оценки чувствительности эпифитных лишайников различных видов к загрязнению воздуха. // Проблемы экологического мониторинга и моделирования экосистем. Л.: Гидрометеоиздат, 1989. Т. 12. С. 113–175.
Красная книга города Москвы. . М., 2011. 928 с.
Лишайники природной территории по р. Котловке (между Нахимовским проспектом и ул. Ремизова). // Лихенологический сайт Алексея Пчёлкина URL: http://www.lichenhouse.narod.ru (дата обращения 10.12.2012).
Малышева А. Г. Закономерности трансформации органических соединений в окружающей среде. // Гигиена и санитария. 1997. № 3. С. 5–10.
Малышева Н. В. О субурбанофлорах лишайников городов. // Флора лишайников России: состояние и перспективы исследований. Труды международного совещания, посвященного 120-летию со дня рождения Всеволода Павловича Савича. Санкт-Петербург, 24-27 октября 2006. СПб: БИН РАН, 2006. С. 147–148.
Москва в цифрах. 1987: Статистический ежегодник. Мосгорстат. М.: Финансы и статистика., 1987. 208 с.
Портал префектуры ЮАО. . URL: http://www.uao.mos.ru (дата обращения 10.10.2012).
Пчелкин А. В. Лишайники природной территории по р. Котловке (между Нахимовским проспектом и ул. Ремизова). // Лихенологический сайт Алексея Пчёлкина URL: http://www.lichenhouse.narod.ru (дата обращения 10.12.2012).
Работнов Т. А. Фитоценология. . М.: МГУ. 1983. 296 с.
Резер С., Упелли Л. Московские проблемы экологической безопасности и энергетики. // Проблемы безопасности при чрезвычайных ситуациях. 1995. Вып. 7. С. 62–70.
Список лихенофлоры России / сост. Г. П. Урбанавичюс. . СПб: Наука. 2010. 194 с.
Шапиро И. А. Азотный обмен у лишайников и его регуляция. // Ботан. журн. 1986. Т. 71. № 7. С. 841–850.
Шапиро И. А. Физиолого-биохимические изменения у лишайников под влиянием атмосферного загрязнения. // Успехи современной биологии. 1996. Т. 116. № 2. С. 158–171.
Ahti T., Vitikainen O. Bacidia chlorococca, a common toxitolerant lichen inFinland // Memoranda Soc.Fauna Flora Fennica. 1974. V. 49. P. 95–100.
Bates J. W., Bell J. N. B., Massara A. C. Loss of Lecanora conizaeoides and other fluctuations of epiphytes on oak in SE England over 21 years with declining SO2 concentrations // Atmospheric Environment. 2001. V. 35. P. 2557–2568.
Bell J. N. B., Power S. A., Jarraud N., Agrawal M., Davies C. The effects of air pollution on urban ecosystems and agriculture // Int. J. Sustainable Development & Word Ecology. 2011. V. 18. Sp. Iss. SI. P. 226–235.
Biazrov L. G. Die Dynamik der Artendiversität epiphytischer Flechten im Nordbezirk von Moskau (Russland) // Archive for lichenology. 2010. V. 6. P. 1–8.
Bobbink R., Hicks K., Galloway J., Spranger T., Alkemade R., Ashmore M., Bustamante M., Cinderby S., Davidson E., Dentener F., Emmett B., Erisman J.-W., Fenn M., Gilliam F., Nordin A., Pardo L., de Vries W. Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis // Ecological Applications. 2010. V. 20. P. 30–59.
Brown D. H., Avalos A., Miller J. E., Bargagli R. Interactions of lichens with their mineral environment // Cryptogamic Botany. 1994. V. 4. P. 135–142.
Dahlman L., Persson J., Palmqvist K., Nasholm T. Organic and inorganic nitrogen uptake in lichens // Planta. 2004. V. 219. P. 459–467.
Davies L., Bates J. W., Bell J. N. B., James P. W., Purvis O. W. Diversity and sensitivity of epiphytes to oxides of nitrogen inLondon// Environmental Pollution. 2007. V. 146. P. 299–310.
Davies L., Bell J. N. B., Bone J., Head M., Hill L., Howard C., Hobbs S. J., Jones D. T., Power S. A., Rose N., Ryder C., Seed L., Stevens G., Toumi R., Voulvoulis N., White P. C. L. Open Air Laboratories (OPAL): a community-driven research programme // Environmental Pollution. 2011. V. 159. P. 2203–2210.
De Schrijver A., De Frenne P., Ampoorter E., van Nevel L., Demey A., Wuyts K., Verheyen K. Cumulative nitrogen input drives species loss in terrestrial ecosystems // Global Ecology & Biogeography. 2011. V. 20. P. 803–816.
van Dobben H. F. Vegetation as a monitor for deposition of nitrogen and acidity . Rijksuniversiteit te Utrecht, Utrecht., 1993. 214 p.
van Dobben H. F., de Bakker A. J. Re-mapping epiphytic lichen biodiversity in the Netherlands – effects of decreasing SO2 and increasing NH3 // Acta Botanica Neerlandica. 1996. V. 45. P. 55–71.
Gadsdon S. R., Dagley J. R., Wolseley .P. A., Power S. A. Relationships between lichen community composition and concentrations of NO2 and NH3 // Environmental Pollution. 2010. V. 158. P. 2553–2560.
Gaio-Oliveira G., Dahlman L., Palmqvist K., Maguas C. Ammonium uptake in the nitrophytic lichen Xanthoria parietina and its effects on vitality and balance between symbionts // Lichenologist. 2004. V. 36. P. 75–86.
Gaio-Oliveira G., Dahlman L., Palmqvist K., Maguas C. Responses of the lichen Xanthoria parietina (L.) Th. Fr. to varying thallus nitrogen concentrations // Lichenologist. 2005. V. 37. P. 171–179.
Grandin U. Epiphytic algae and lichen cover in boreal forests-a long-term study along a N and S deposition gradient inSweden// AMBIO. 2011. V. 40. P. 857–866.
Hauck M. Ammonium and nitrate tolerance in lichens // Environmental Pollution. 2010. V. 158. P. 1127–1133.
Hauck M. Eutrophication threatens the biochemical diversity in lichens // Lichenologist. 2011. V. 43. P. 147–154.
Hawksworth D. L., Mc Manus P. M. Lichen recolonization in London under conditions of rapidly falling sulphur dioxide levels, and the concept of zone skipping // Bot. J. Linnean Soc. 1989. V. 100. P. 99–109.
van Herk C. M. Mapping of ammonia pollution with epiphytic lichens in the Netherlands// Lichenologist. 1999. V. 31. P. 9–20.
van Herk C. M., Aptroot A., van Dobben H. F. Long-term monitoring in the Netherlands suggests that lichens respond to global warming // Lichenologist. 2002. V. 34. P. 141–154.
van Herk C. M., Mathijssen-Spiekman E. A. M., de Zwart D. Long distance nitrogen air pollution effects on lichens in Europe // Lichenologist. 2003. V 35. P. 347–359.
Isocrono D., Matteucci E., Ferrarese A., Pensi E., Piervittori R. Lichen colonization in the city of Turin (N Italy) based on current and historical data // Environmental Pollution. 2007. V. 145. P. 258–265.
Kandler O., Poelt J. Wiederbesiedlung der Innenstadt von München durch Flechten // Naturwissensch. Rundschau. 1984. Bd. 37. S. 90-95.
Kirschbaum U., Windisch U., Vorbeck A., Hanewald K. Mapping lichen diversity in Wetzlar and Giessen as an indicator of air quality - Comparison between the surveys of 1970, 1985, 1995 and 2005 // Gefahrstoffe Reinhaltung der Luft. 2006. Bd. 66. S. 272–280.
Krupa S. V. Effects of atmospheric ammonia (NH3) on terrestrial vegetation: a review // Environmental Pollution. 2003. V. 124. P. 179–221.
Larsen R. S., Bell J. N. B., James P. W., Chimonides P. J., Rumsey F. J., Tremper A., Purvis O. W. Lichen and bryophyte distribution on oak in London in relation to air pollution and bark acidity // Environmental Pollution. 2007. V. 146. P. 332–340.
Öckinger E., Niklasson M., Nilsson S. G. Is local distribution of the epiphytic lichen Lobaria pulmonaria limited by dispersal capacity or habitat quality? // Biodiversity & Conservation. 2005. V. 14. P. 759–773.
Palmqvist K., Dahlman L., Valladares F., Tehler A., Sancho L. G., Mattsson J.-E. CO2 exchange and thallus nitrogen across 75 contrasting lichen associations from different climate zones // Oecologia 2002. V. 133. P. 295–306.
Paoli L., Pirintsos S.A., Kotzabasis K., Pisani T., Navakoudis E., Loppi S., Effects of ammonia from livestock farming on lichen photosynthesis // Environmental Pollution. 2010. V. 158. P. 2258–2265.
Pardo L. H., Fenn M. E., Goodale C. L., Geiser L. H., Driscoll C. T., Allen E. B., Baron J. S., Bobbink R., Bowman W. D., Clark C. M., Emmett B., Gilliam F. S., Greaver T. L., Hall S. J., Lilleskov E. A., Liu L. L., Lynch J. A., Nadelhoffer K. J., Perakis S. S., Bobin-Abbott M. J., Stoddard J. L., Weathers K. C., Dennis R. L. Effects of nitrogen deposition and empirical nitrogen critical loads for ecoregions of the United States // Ecological Applications. 2011. V. 21. P. 3049–3082.
Phoenix G. K., Emmett B. A., Britton A., Caporn S. J. M., Dise N. B., Helliwell R., Jones L., Leake J. R., Leith I. D., Sheppard L. J., Sowerby A., Pilkington M. G., Rowe E. C., Ashmorek M. R., Power S. A. Impacts of atmospheric nitrogen deposition: responses of multiple plant and soil parameters across contrasting ecosystems in long-term field experiments // Global Change Biology. 2012. V. 18. P. 1197–1215.
Purvis O. W., Chimonides J., Din V., Erotokritou L., Jeffries T., Jones G. C., Louwhoff S., Read H., Spiro B. Which factors are responsible for the changing lichen floras of London? // Sci. Total Environ. 2003. V. 310. P. 179–189.
Seaward M. R. D. Lichens and hypertrophication // Lichens in a changing pollution environment / Eds. Lambley P., Wolseley P. English Nature Research Reports, English Nature, 2004. P. 9–12.
Silberstein L., Siegel B. Z., Siegel S. M., Mukhtar A., Galun M. Comparative studies on Xanthoria parietina, a pollution-resistant lichen, and Ramalina duriaei, a sensitive species. 2. Evaluation of possible air pollution-protection mechanisms // Lichenologist. 1996. V. 28. P. 367–383.
SochtingU., Lutzoni F. Molecular phylogenetic study at the generic boundary between the lichen-forming fungi Caloplaca and Xanthoria (Ascomycota, Teloschistaceae) // Mycological Research. 2003. V. 107. P. 1266-1276.
Stevens C. J., Smart S. M., Henrys P. A., Maskell L. C., Crowe A., Simkin J., Cheffings C. M., Whitfield C., Gowing D. J. G., Rowe E. C., Dore A. J., Emmett B. A. Terricolous lichens as indicators of nitrogen deposition: evidence from national records // Ecological Indicators. 2012. V. 20. P. 196–203.
Wirth V. Zeigewerte von Flechten // Scripta Geobotanica. 1991. Bd. 18. S. 215–237.
Wirth V. Ökologische Zeigerwerte von Flechten – erweiterte und aktualisierte Fassung.// Herzogia. 2010. Bd. 23. S. 229–248.
Wolseley P., James P. Assessing the role of biological monitoring using lichens to map excessive ammonia (NH3) deposition in the UK // Effects of NOx and NH3 on lichen communities and urban ecosystems. A Pilot Study: A report produced by Imperial College& The Natural History Museum, 2002. P. 68–87.
Wolseley P. A., James P. W., Theobald M. R., Sutton M. A. Detecting changes in epiphytic lichen communities at sites affected by atmospheric ammonia from agricultural sources // Lichenologist. 2006. V. 38. P. 161–176.
Благодарности
Работа выполнена по плану НИР Лаборатории радиоэкологического мониторинга в регионах АЭС и биоиндикации Института проблем экологии и эволюции РАН и частично финансировалась программой фундаментальных исследований президиума РАН «Живая природа: современное состояние и проблемы развития».

 © 2011 - 2026
© 2011 - 2026