



Введение
В последние десятилетия исследования видообразования очень часто сводятся к поиску механизмов формирования репродуктивной изоляции (Coyne, Orr, 2004). Между тем большинству биологов-«полевиков» очевидно, что вид имеет «экологическое измерение» – он занимает определенный ареал и определенную экологическую нишу.
Это противоречие находит выход в появлении работ, посвященных «экологическому видообразованию» (как будто видообразование может быть «неэкологическим»!). В то же время обзор по этой проблеме (Rundle, Nosil, 2005) содержит в основном описание экологических механизмов формирования репродуктивной изоляции.
В настоящей работе мы постараемся показать, что видообразование – это только конечный этап длинного процесса адаптации предкового вида к новым условиям обитания. В этой связи интересны переходные зоны между ареалами близкородственных видов, исследование которых может быть полезно для понимания механизмов образования этих видов. Мы обобщили данные об экологических и морфологических признаках, применяемых для диагностики двух видов ряпушек – европейской (C. albula) и сибирской (C. sardinella) – в переходной зоне между их ареалами, занимающей практически весь север европейской части России. Отметим, что ряпушка принадлежит к семейству сиговых (Coregonidae), которое уже много лет служит моделью для решения фундаментальных проблем теории эволюции (Попов, Сендек, 2003).
Для определения направления расселения ряпушек анализировались результаты исследований полиморфизма молекулярно-генетических маркеров, прежде всего фрагментов митохондриальной ДНК, что наилучшим образом подходит для решения этой задачи (Avise, 2004). Для анализа гибридизации двух форм использованы данные о ядерных генах (изолокусах), кодирующих фермент креатинкиназу (Перелыгин, 1988).
Аналитический обзор
Распространение ряпушки
В Европе ареал ряпушки охватывает бассейны Северного, Балтийского, Белого и Баренцева морей, в Сибири – простирается до Берингова моря. Южная граница обитания проходит по Псковско-Чудскому водоему и озерам Верхней Волги (рис. 1); в Сибири же ряпушка не поднимается выше среднего течения крупных рек (Обь, Лена, Енисей и др.), хотя отдельные ее популяции отмечены в озерах Камчатки, забайкальском озере Баунт. В районе р. Печора, согласно последней сводке (Атлас... , 2003), происходит перекрывание ареалов европейской и сибирской ряпушек.
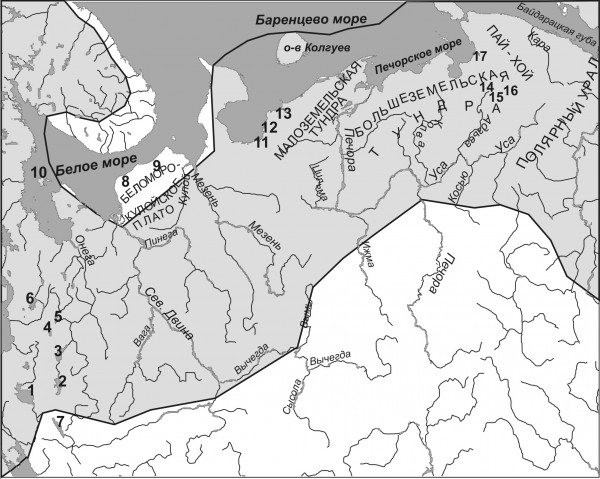
Рис. 1. Распространение ряпушки на Европейском Севере России. Цифрами обозначены: озера: 1 – Белое; 2 – Воже, 3 – Лача, 4 – Лекшмозеро, 5 – Кенозеро, 6 – Водлозеро, 7 – Кубенское, 14 – Вашуткины, 15 – Падимейские, 16 – Харбейские; устья рек: 8 – Зимняя Золотица, 9 – Мегра, 11 – Пежа, 12 – Волонга, 13 – Индига, 17 – Коротаиха; 10 – Соловецкие острова
(Fig. 1. The area of distribution of the vendace in the European North of Russia. Lakes are denoted by figures as well as the estuaries of the rivers.)
На территории европейского северо-востока России ряпушка обитает во многих реках и озерах. Озера предпочитает площадью не менее 10 га и встречается, по данным Л. Н. Соловкиной (1974), тем чаще, чем крупнее озеро. Наибольшее число самых крупных озер, населенных ряпушкой, сосредоточено в верхней и левобережной части бассейна реки Онега (например, озера Лекшмозеро, Кенозеро, Воже (Чарандское), Лаче). Во всех прочих случаях озер с ряпушкой больше в приморских районах, т. е. в бассейнах нижнего течения рек (Соловкина, 1974; Дворянкин и др., 2008). Так, ряпушка встречается в озерах бассейна р. Сев. Двина (но не выше г. Холмогоры) (Соловкина, 1974); отмечена она и в бассейнах рек Мегра и Кулой (озера Волчье, Бол. Восточное, Ернозеро, Зап. Мегорские озера) (Сендек и др., 2010). В бассейне реки Мезень обитает в озере Варша (Танфильев, 1896).
В пределах Малоземельской тундры ряпушка известна в озерах Хуновейто и Янгутейто к северу от дельты Печоры, озерах Голодная Губа и Матервисочное Нижней Печоры (Соловкина, 1974). Несколько южнее полярного круга она отмечена в Мыльских озерах, озере Пустозерское долины нижнего течения Печоры (Танфильев, 1896; Зверева и др., 1953), озере Лемваты бассейна одного из уральских притоков р. Усы (Кучина, 1962), а также в озерах Косьминском, Волочанском, Смольном (Зверева и др., 1955).
Распространение ряпушки в водоемах Большеземельской тундры мало изучено. Популяции ряпушки обнаружены в трех крупных озерно-речных системах восточной части региона: Вашуткинской, Харбейской и Падимейской, в озерах Варьяты, Коматы, Нямдоты и Серветы (Сидоров, 1974), а также в системе реки Пильня (Антрошенко и др., 1979).
Из крупных рек региона ряпушка указана для р. Кара (Пробатов, 1934; Соловкина, 1974); кроме того, Г. И. Танфильевым (1896) она указана для р. Индига. В составе ихтиофауны р. Вычегда ряпушка не отмечена; нахождение ее в реках Северная Двина и Мезень спорно (Зверева и др., 1955).
В реке Печора ряпушка широко распространена от устья до впадения одного из крупнейших притоков, реки Уса. В среднем течении Печоры ряпушка встречается уже в гораздо меньшем количестве, а выше реки Кожва – единично (Зверева и др., 1955; Соловкина, 1974). В реке Уса ряпушка довольно обычна лишь в нижнем течении; только отдельные особи поднимаются до с. Петрунь и иногда – до с. Абезь (Зверева и др., 1955; Соловкина, 1962). Указывается наличие ряпушки в ряде озер долины Усы, а также ее притоков: Лемва, Косью (левобережные), Адзьва, Колва и Макариха (правобережные). Необходимо заметить, что нахождение ряпушки в р. Колва – явление довольно редкое (Кучина, 1962; Соловкина, 1962); ряпушка р. Лемва – предполагаемый ледниковый реликт (Пономарев, 2008).
Кроме озер материка, ряпушка населяет также озера островов – Большого Соловецкого (Захваткин, 1927; Мухомедияров, 1963; Анухина, 1972; Дворянкин, 2005; Алексеева, Решетников, 1997) и Колгуева (Михайловский, 1903; Покровский, 1967).
Экологические особенности популяций переходной зоны между европейской и сибирской ряпушками
Значительная экологическая изменчивость ряпушки выражена в Печорском бассейне, что связано с наибольшим разнообразием мест нагула и нереста. Так, Л. Н. Соловкина (1974), кроме полупроходной зельди, которая нагуливается в опресненных участках океана, а на нерест поднимается вверх по течению Печоры, упоминает ямную (обитает в глубоких плесах реки Уса) и озерную (саурей) ряпушек. В среднем течении Печоры, реке Уса, а также усинских притоках – реках Колве и Косью – отмечено появление жилой (туводной) пойменно-речной ряпушки, нагуливающейся в пойменных озерах и заливах, а нерестующей, как и зельдь, в русле реки (Зверева и др., 1955; Кучина, 1962; Соловкина, 1974; Туманов, 2010).
Отметим, что даже в пределах Печоры и ее притоков разные популяции имеют свои особенности. Например, обитающая в оз. Голодная Губа ряпушка-саурей, а также ряпушка реки Уса отличаются от зельди большей продолжительностью жизни и поздним созреванием (Соловкина, 1962; Корнилова, 1967; Туманов, 2010). Более того, анализ возрастных спектров выборок из реки Уса позволил сделать ряду авторов предположение о формировании в ней местной, туводной, формы (Зверева и др., 1955; Соловкина, 1962, 1974; Туманов, 2010), хотя морфологически усинская ряпушка мало отличается от печорской, заходящей в Усу на нерест (Кучина, 1962; Соловкина, 1962). Возможность формирования местных форм ряпушки указывается для разных участков течения Печоры и ряда усинских притоков (Кучина, 1962; Соловкина, 1974).
Л. Н. Соловкина (1974) отмечает, что темп роста и полового созревания ряпушки из различных водоемов не совпадают. Так, саурей по темпу линейного роста близок зельди, но созревает в массе на 1–2 года позднее при длине 19–20 см. Половая зрелость у лекшмозерской ряпушки наступает при достижении длины тела всего 10–12 см (Дворянкин и др., 2007). У ряпушки Вашуткиных озер тугорослость сочетается с относительно ранней половозрелостью (Соловкина, 1974). Выделяется своими размерами, хорошим темпом роста и ряпушка ряда озер Большого Соловецкого острова. Однако в данном случае нельзя исключать последствий гибридизации с другими сиговыми, которых акклиматизировали в соловецких озерах (сиг, пелядь) (Захваткин, 1927; Мухомедияров, 1963; Дворянкин, 2005).
Подъем половозрелой ряпушки в реках Печора и Кара начинается в конце июля, пик хода приходится на август. Кроме августовского, есть еще сентябрьский ход, но он выражен гораздо слабее и, как правило, значительными миграциями не сопровождается (Пробатов, 1934; Зверева и др., 1953).
Дальность нерестовых миграций карской и печорской ряпушек различаются. Так, в Печоре зельдь поднимается для нереста достаточно высоко – граница ее подъема находится на участке верхней древнеозерной низины реки Уса у с. Абези. В то же время, хотя основные нерестилища находятся у местечка Адак, на участке гряды Чернышева и в районе устья реки Макариха (Зверева и др., 1955; Соловкина, 1974), нерест может происходить на разных участках миграционного пути – нерестилища зельди обнаружены даже в печорской дельте (Соловкина, 1974). Карская ряпушка нерестует в низовьях Кары и Сибирчи, в зоне действия морского прилива (Пробатов, 1934).
Основной период нереста в Печорском бассейне ограничивается 10 днями, предшествующими ледоставу или появлению шуги. На верхних нерестилищах зельди, в реке Уса, икрометание происходит раньше, чем в Печоре – в первой или во второй декаде октября, что связано с более суровыми температурными условиями (Соловкина, 1962, 1974). Нерест же ряпушки реки Кара сильно растянут, основная масса рыбы подходит уже подо льдом (Пробатов, 1934).
Условия и сроки нереста озерных популяций исследованы слабо. Предполагается, что ряпушка Харбейских (Сидоров, 1974) и Мыльских (Зверева и др., 1953) озер в основном нерестится подо льдом. Частично подо льдом происходит нерест более крупных особей в Лекшмозере, тогда как впервые нерестующие мелкие рыбы подходят к местам нереста еще по открытой воде (Дворянкин и др., 2007). Нерест соловецкой ряпушки, по-видимому, также подледный, поскольку нерестовые подходы начинаются лишь в начале ноября, а пик нереста приходится на вторую половину этого месяца (Мухомедияров, 1963).
Икра откладывается, как правило, на песчаный, галечно-песчаный и песчано-галечный грунты (Соловкина, 1962, 1974). Однако ряпушка, обитающая в тундровых озерах, по всей вероятности, способна откладывать икру на частично заиленный каменисто-галечный грунт, так как для большинства озер тундры грунт со значительной долей песка не характерен (Корнилова, 1967; Соловкина, 1974). Поскольку время нереста и нерестилища ряпушки и других сиговых, как в озерах, так и в реках, часто перекрываются, становится возможным образование межвидовых гибридов (Козьмин, 2011).
Абсолютная плодовитость ряпушки Печоры составляет в среднем 5.9 тыс. икринок и изменяется от 2.6 до 12.4 тыс. у формы «зельдь» и от 4.8 до 8.5 – у формы «саурей» (Соловкина, 1974). Значительно меньше плодовитость ряпушки, обитающей в озерах. Так, у ряпушки Кенозера и Лекшмозера значение этого показателя варьирует в среднем от 2.4 тыс. икринок у двухлеток до 6.1 тыс. икринок у рыб в возрасте 4+ (Дворянкин, 2010). Средняя индивидуальная плодовитость соловецкой ряпушки у трехлеток равна 1.8 тыс., у четырехлеток – 3.7 тыс. икринок (Мухомедияров, 1963).
Морфологическое разнообразие популяций ряпушки Европейского Севера России
В работах Л. С. Берга (1916, 1948) популяции региона были описаны как особый подвид сибирской ряпушки – беломорская, или печорская, ряпушка C. sardinella marisalbi (первоначальное написание названия maris-albi заменено на marisalbi в соответствии со статьей 32.5.2.3 Международного кодекса зоологической номенклатуры). Однако еще в конце XIX века исследователи указывали на ее неоднородность. Так, в ряде водоемов встречались особи, сходные по морфологическим критериям как с типичной европейской ряпушкой, так и с типичной сибирской, а часть особей имела промежуточные между двумя видами значения признаков (Танфильев, 1896).
Как правило, морфологически ближе к сибирскому виду оказывались речные и полупроходные формы ряпушки, а к европейскому – популяции, обитающие в озерах – как тундровых, так и пойменных и дельтовых (Иванова, Петрова, 1972; Соловкина, 1974; Новоселов, 1991; Туманов, 2010). Таким образом, сложилось представление о существовании зоны перекрывания ареалов двух видов ряпушки, где происходит (или происходила) их гибридизация и формируются смешанные (гибридные) популяции.
К подвиду C. s. marisalbi одно время относили также популяцию озера Водлозеро в восточной части бассейна Онежского озера (Покровский, 1967); эта популяция первоначально была описана как подвид европейской ряпушки, C. a. vodlosericus (Лукаш, 1939). Ряпушку озера Белого в верховьях Волги тоже считали сибирской, но рассматривали ее как другой подвид – C. s. vessicus (Дрягин, 1933). В то же время было показано, что в западной части беломорского бассейна обитает уже европейская ряпушка (Первозванский, 1986 и ссылки в этой работе).
Отсутствие четкого хиатуса по видовым признакам, биологические и морфологические особенности, отсутствие репродуктивной изоляции между европейской и сибирской ряпушками в зоне перекрывания ареалов привели к формированию в 1960–70‑х годах представления о едином виде ряпушки, включающем европейский и сибирский подвиды (Покровский, 1967; Дрягин и др., 1969; Соловкина, 1974).
Позже, однако, Ю. С. Решетников (1980) обобщил данные ряда исследователей и показал, что антедорсальное расстояние, которое использовали в качестве диагностического признака для ряпушек – критерий достаточно пластичный, и, следовательно, для целей систематики непригодный. В качестве диагностических признаков этим автором было предложено использовать число позвонков и отношение вентроанального расстояния к антедорсальному (в среднем 56 % у европейской и свыше 62 % – у сибирской ряпушек). По мнению Ю. С. Решетникова (1980), анализ этих характеристик подтверждает существование двух видов ряпушек.
Однако, как отмечал В. В. Кузнецов (1987), хиатуса по данным признакам между видами не наблюдается. Более того – при использовании морфологических критериев вида по (Решетников, 1980) некоторые озерные популяции ряпушки Сибири могут быть классифицированы как европейская ряпушка (Романов, 2000).
В работе (Атлас... , 2003), наряду с числом позвонков (54–59 у европейской, 57–64 – у сибирской ряпушек), в качестве диагностического признака для двух видов вновь использовано отношение антедорсального расстояния к длине тела (больше 42 % и «часто менее 42 %» соответственно). Тем не менее, согласно упомянутой публикации, сибирская ряпушка не встречается западнее Печоры.
В то же время, как в цитированных выше, так и в новых работах (Кузищин и др., 1999; Дворянкин, 2005; Петрова, Кудерский, 2006; Боровикова, 2009) отмечено, что в озерах Белом, Водлозере и озерах Соловецких островов обнаружены экземпляры, у которых значение предложенных Ю. С. Решетниковым (1980, 2003) диагностических признаков соответствует характерным для сибирской ряпушки. В озере Имандра на Кольском полуострове выявлена популяция, число позвонков у особей которой – промежуточное между значениями, характерными для двух видов – 59–61 (Марченко, 1981).
Кроме того, если рассматривать комплекс признаков, а не отдельно взятые характеристики, то ряпушка озера Водлозеро оказывается более «сибирской», чем печорская зельдь (Покровский, 1967). Ряпушка озер Воже и Белое в работах (Покровский, 1967; Коновалов, Болотова, 2004) по комплексу признаков оказалась ближе C. sardinella, чем C. albula. В работе Н. Л. Болотовой (2010) ряпушка озера Воже отнесена к виду C. sardinella.
Особо следует остановиться на статусе ряпушки Соловецких островов, откуда происходит типовой экземпляр C. s. marisalbi (Берг, 1916). По данным из работы, где приводится первое описание значительной выборки ряпушки из озера Красное Большого Соловецкого острова, число позвонков у рыб варьировало от 54 до 58 при среднем значении 55.5 (Мухомедияров, 1963). Отношение вентроанального расстояния к антедорсальному (VA к AD), рассчитанное нами на основании данных, приведенных в этой работе, составило 56.3 %. Таким образом, оба показателя соответствуют признакам европейской ряпушки.
Последующие исследования ихтиофауны озер Большого Соловецкого острова выявили популяции ряпушки, в которых число позвонков у некоторых особей достигало 61, а отношение VA к AD – 66.7 %. При этом средние значения не выходили, однако, за пределы признаков, характерных для типичной C. albula (Кузищин и др., 1999; Дворянкин, 2005; Боровикова и др., 2008).
Хотя для ряпушки, как и для всех сиговых, характерна значительная морфологическая пластичность, в пределах рассматриваемого региона ее популяции можно разделить на две группы (формы). Так, у рыб речных популяций имеется тенденция в пользу признаков сибирской ряпушки (C. sardinella) – в частности, спинной плавник их более сдвинут вперед; для озерных форм характерно преобладание и большая выраженность признаков европейской ряпушки (C. albula) (Покровский, 1967; Соловкина, 1974). Согласно проведенному нами сравнению (табл. 1–4), озерная и речная ряпушки отличаются, кроме того, положением грудных, брюшных и анального плавников (индексы: вентроанальное (VA) расстояние в % антедорсального (AD), вентроанальное и пектровентральное (PV) расстояния в % от длины тела по Смитту), высотой анального (Ha) и длиной брюшных плавников (Lv). Отметим, что положение плавников определяется подвижностью рыбы и способностью ее к длительным миграциям (Покровский, 1967).
Поскольку происхождение популяций ряпушки Соловецких островов не до конца ясно и нельзя исключить возможность гибридизации с пелядью, которая также вселялась в некоторые озера острова (Дворянкин, 2005), ряд морфологических признаков озерной и речной ряпушек мы сравнивали с учетом соловецких популяций и без них (группа озерных популяций, отмеченных звездочкой в таблице 1, столбец 5, не включает в себя популяции Большого Соловецкого острова; с учетом последних вычислены средние значения в столбце 3).
Среди меристических признаков, по которым различаются озерные и речные популяции – число неветвистых лучей в анальном плавнике и число ветвистых лучей в брюшных плавниках. Кроме того, при сравнении популяций озер бассейна реки Онеги с речными популяциями оказывается, что число позвонков у ряпушки первых – меньше. Таков же результат сравнения числа позвонков речной ряпушки и ряпушки из озер Соловецкого острова (табл. 1). Видимо, здесь сказывается влияние температурного фактора на эмбриогенез. Так, показано, что у эмбрионов сиговых рыб при низких температурах формируется большее число метамерных структур, дающих начало, среди прочих, и позвоночнику, чем при более высоких температурах, когда скорость развития эмбриона выше (Черняев, 2007).
Таблица 1. Достоверность различий разных групп популяций региона по ряду морфологических признаков (группы сравниваются попарно)
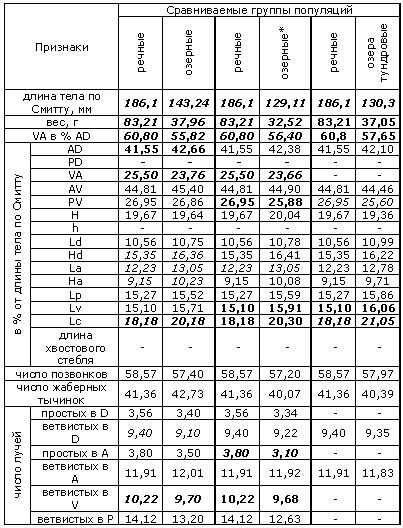
Таблица 1 (продолжение). Достоверность различий разных групп популяций региона по ряду морфологических признаков (группы сравниваются попарно)
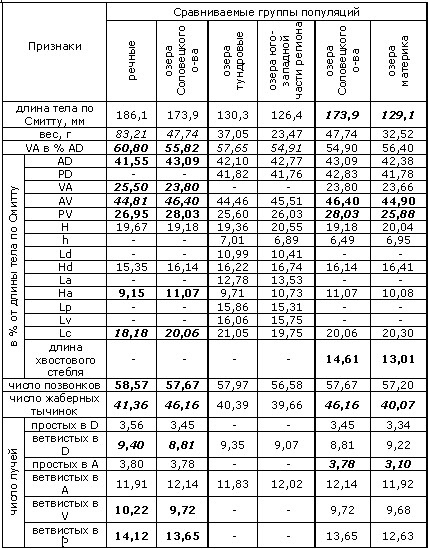
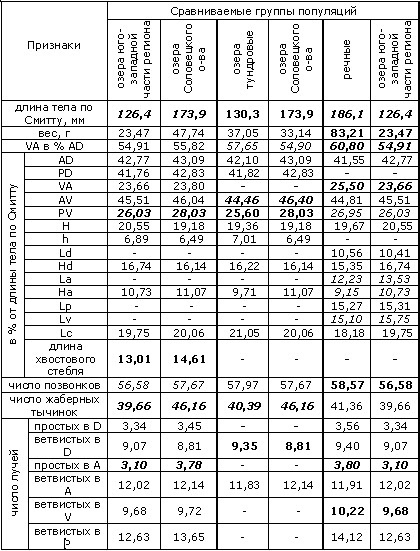
Пояснения к таблице 1. Для оценки достоверности различий использован критерий Стьюдента (t). Жирным курсивом отмечены признаки, различающиеся при уровне значимости 0.01; жирным шрифтом – различающиеся при уровне значимости 0.05, курсивом – 0.1. В таблице приняты обозначения признаков: расстояния: VA – вентроанальное; AD – антедорсальное; PD – постдорсальное; AV – антевентральное; PV – пектровентральное; H – максимальная высота тела; h – минимальная высота тела; Ld – длина основания спинного плавника; Hd – высота спинного плавника; La – длина основания анального плавника; Ha – высота анального плавника; Lp – длина грудного плавника; Lv – длина брюшного плавника; Lc – длина головы; плавники: D – спинной; A – анальный; Р – грудной; V – брюшной. Признаки, указанные в таблице, измерялись согласно схеме, предложенной в работе (Правдин, 1966). Информация о местах сбора выборок и ссылки на первоисточники приведены в таблицах 2–4.
Таблица 2. Речные популяции, для которых имеются данные по пластическим признакам
| Водоем | Источник | Объем выборки |
| Печора (у дер. Климовка) | Зверева и др., 1953 | 78 |
| Печора (Барашечное плесо) | 10 | |
| Печора | Покровский, 1967 | 11 |
| Уса, устье р. Макарихи | Соловкина, 1962 | 21 |
| Уса, тоня Артысьёль | 35 | |
| Уса, дер. Адак | 34 | |
| Уса, у с. Петруни | 19 | |
| Индига (бассейн Баренцева моря) | Покровский, 1967 | 1 |
| Кара | Пробатов, 1934 | 379 |
Таблица 3. Речные популяции, для которых имеются данные по меристическим признакам
| Название водоема | Источник | Объем выборки |
| Печора (у дер. Климовка) | Зверева и др., 1953 | 78 |
| Печора (Барашечное плесо) | 10 | |
| Печора | Покровский, 1967 | 11 |
| Печора, с. Усть-Цильма | Туманов, 2002 | 42 |
| Уса (туводная) | 86 | |
| Уса (полупроходная) | 131 | |
| Уса, устье р. Макарихи | Соловкина, 1962 | 21 |
| Уса, тоня Артысьёль | 35 | |
| Уса, дер. Адак | 34 | |
| Уса, у с. Петруни | 19 | |
| Колва (туводная) | Туманов, 2002 | 92 |
| Индига (бассейн Баренцева моря) | Покровский, 1967 | 1 |
Таблица 4. Озерные популяции, для которых имеются данные по пластическим признакам (отсутствовали данные по меристическим признакам для ряпушки из озер № 1, Большой Торавей, Науль-То)
| Группа | Регион, бассейн | Название водоема | Источник | Объем выборки |
| тундровые озера | Бассейн Нижней Печоры | Большое Мыльское | Зверева и др., 1953 | 100 |
| Малое Мыльское | 50 | |||
| Малоземельская тундра | Пустозерское | Покровский, 1967 | 2 | |
| Большеземельская тундра | Вашуткины озера | Соловкина, 1974 | 15 | |
| Есипов, 1941 | 94 | |||
| № 1 | Пономарев, 1996 | – | ||
| Большой Торавей | – | |||
| Науль-То | – | |||
| О-в Колгуев | Песчаное | Покровский, 1967 | 6 | |
| Басс. р. Мезень | Варша и Бормат | 12 | ||
| озера западной части региона | Система р. Онега, восточная часть бас. Белого моря | Лекшмозеро | Дворянкин и др., 2007 | 40 |
| Покровский, 1967 | 40 | |||
| Бас. Онежского озера | Водлозеро, 1956 | Петрова, Кудерский, 2006 | – | |
| Водлозеро, 2000 | 25 | |||
| Водлозеро | Боровикова и др., 2006 | – | ||
| Соловецкий архипелаг, Большой Соловецкий остров | Большое Остречье | Дворянкин, 2005 | 52 | |
| Большое Остречье (Горелое) | Боровикова и др., неопубл. данные | 20 | ||
| Средний Перт | Дворянкин, 2005 | 26 | ||
| Боровикова и др., неопубл. данные | 42–64 | |||
| Нижний Перт (Хуторское) | Боровикова и др., неопубл. данные | 99–154 | ||
| Красное | Мухомедияров, 1963 | – | ||
| Большое Красное | Боровикова и др., неопубл. данные | 31–93 | ||
| Большое Гремячье | 7 | |||
Молекулярно-генетические особенности популяций переходной зоны
Поскольку морфологические критерии, различающие C. albula и C. sardinella, нельзя назвать строгими, для уточнения видового статуса ряпушки ряда популяций рассматриваемого региона были привлечены результаты изоферментного (аллозимного) анализа (Сендек, 1998; Sendek, 2002; Махров и др., 2003; Боровикова и др., 2006, 2008; Боровикова, 2009; Borovikova et al., 2009; Сендек и др., 2010). В качестве маркерного использован дуплицированный локус креатинкиназы СК-A1,2*, по электрофоретическим спектрам которого ранее было показано различие между C. albula и C. sardinella (Перелыгин, 1988).
Применение метода изоферментного анализа позволило выявить бóльшую генетическую близость печорской ряпушки к C. sardinella, чем к C. albulа, причем сходство с тем или иным видом оказалось зависимым от гидрологических режимов, в которых обитает конкретная популяция (Сендек, 1998; Sendek, 2002). Так, ряпушка из участков основного русла Печоры по распределению частот аллелей креатинкиназы имеет бóльшее сродство с C. sardinella; ряпушка же из участков с невысокой проточностью и стоячих вод дельтовой части реки тяготеет более к C. albulа. Кроме того, в результате исследования были выявлены генотипы, свидетельствующие о гибридизации между двумя этими формами (Сендек, 1998; Sendek, 2002).
Популяции ряпушки озер Беломоро-Кулойского плато (озера Волчье, Большое Восточное, Западные Мегорские озера, Ернозеро) оказались отнесены по данным изоферментного анализа к типичной европейской ряпушке (Сендек и др., 2010), что, впрочем, соответствует их видовому диагнозу и по морфологическим критериям (Решетников, 1980; Атлас... , 2003; Новоселов, 1991).
Позднее аллель, характерный для сибирской ряпушки, был выявлен в популяциях из водоемов, расположенных намного западнее общепринятой зоны гибридизации, бассейна Печоры. Так, высокие частоты одного из «сибирских» аллелей обнаружены у ряпушки озера Белое (верховья Волги, бас. Каспийского моря) (Махров и др., 2003), озера Водлозеро (бас. Онежского озера) (Боровикова и др., 2006), в ряде озер западной части Архангельской области – Кенозере, Лекшмозере, Наглимозере, Масельгском (Гордеева и др., 2009) (табл. 5).
Таблица 5. Частоты аллелей локусов креатинкиназы СК-А1,2* в популяциях переходной зоны между европейской и сибирской ряпушками (аллели локусов обозначены условно согласно их повышающейся подвижности при электрофорезе; для сравнения приведены данные для ряда «типичных» европейских и сибирских популяций). Озера Большое Красное, Большое Остречье (Горелое), Нижний Перт, Гремячее расположены на Большом Соловецком острове.
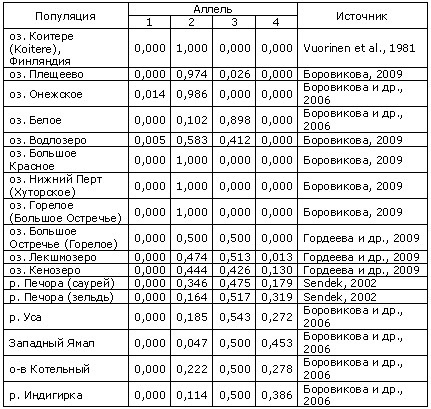
Кроме того, с использованием ПЦР-ПДРФ (полимеразная цепная реакция, полиморфизм длин рестриктных фрагментов) анализа ND-1 фрагмента митохондриальной ДНК (мтДНК) в популяции Водлозера показано присутствие гаплотипа, ранее отмеченного только у ряпушки Сибири (Боровикова и др., 2006; Боровикова, Махров, 2009).
Единственная особь из Вашуткиных озер (Большеземельская тундра), которую нам удалось изучить, согласно результатам ПЦР-ПДРФ анализа оказалась сибирской ряпушкой (Боровикова и др., неопубл. данные).
Ряпушка большинства популяций Большого Соловецкого острова, по данным о полиморфизме маркерного локуса креатинкиназы CK-A1,2*, является типичной европейской ряпушкой (Боровикова и др., 2008; Боровикова, 2009; Borovikova et al., 2009). В то же время, согласно работе Н. В. Гордеевой с соавт. (2009), в популяции озера Большое Остречье зарегистрированы высокие частоты одного из «сибирских» аллелей (табл. 5). Однако нами в ходе исследования выборки из этого озера «сибирских» аллелей CK-A1,2* не выявлено.
Кроме того, согласно данным ПЦР-ПДРФ анализа ND-1 фрагмента мтДНК в популяциях ряпушки исследованных соловецких озер, в том числе и озера Большое Остречье, преобладает гаплотип Е, широко распространенный у ряпушки европейских водоемов; «сибирский» гаплотип S и/или его производные в популяциях Соловков не выявлены (Боровикова и др., 2008; Borovikova et al., 2009).
Таким образом, генетические данные показывают, что на Европейском Севере России есть два региона, где обнаружены популяции ряпушки с вариантами генов, характерными для сибирских популяций: бассейн Печоры и водораздел Балтийского, Белого и Каспийского морей.
Анализ нуклеотидной последовательности участка ND-1 фрагмента мтДНК длиной 300 н. п. в популяциях ряпушки позволяет сделать вывод, что европейская ряпушка произошла от сибирской, причем по мере продвижения ряпушки на запад происходило формирование новых, все более отличающихся, гаплотипов (Боровикова, 2009).
С учетом генетических данных, имеющихся для других видов (Махров, Болотов, 2006), можно дать объяснение особенностям распределения генотипов в переходной зоне между двумя формами ряпушек. До последнего оледенения ряпушка постепенно расселялась из Сибири в Европу, при этом типично «сибирские» варианты генов заменялись «типично европейскими». В ходе последнего оледенения в районе современного водораздела Балтийского, Белого и Каспийского морей существовал рефугиум, где ряпушка сохранила «переходные» признаки. Но основная часть Европейского Севера заселялась из водоемов, расположенных западнее, где ряпушка имела «типично европейские» генотипы. В бассейне Печоры этот поток встретился с «типично сибирской» ряпушкой; только в этом бассейне возникли гибриды двух форм.
Существование «западного» и «восточного» рефугиумов, где переживали оледенение европейская и сибирская формы ряпушки, ранее обосновано Л. А. Кудерским (1977). Наше предположение о наличии отдельного, третьего, рефугиума в районе современного водораздела Балтийского, Белого и Каспийского морей опирается, кроме генетических данных, еще на три факта. Первый из них – существование мощной лопасти последнего ледника восточнее этого региона (Svendsen et al., 2004). Второй – отсутствие ряпушки в расположенном юго-восточнее Кубенском озере (рис. 1). Третий – отсутствие естественных популяций сига (С. lavaretus) в бассейне современной Волги (Кесслер, 1870), но наличие рефугиума сига в районе современного Кубенского озера (Боровикова, 2011): если бы связь между этими бассейнами существовала, высока была бы вероятность заселения сигом и водоемов бассейна Волги.
Заключение
Таким образом, европейская ряпушка возникла относительно недавно. В ходе вселения в Европу предковой формы – сибирской ряпушки – параллельно происходило изменение морфологических признаков и изменение генофонда; оба процесса, видимо, шли постепенно, и к началу последнего ледникового периода хиатуса между формами не было.
Изменение признаков, связанных с формой тела, было адаптивно и отражало переход от полупроходного к озерному образу жизни, имевший место в ходе заселения Европы. Изменение же меристических признаков, очевидно, неадаптивно и определялось повышением температуры воды по мере продвижения ряпушки на юг и запад.
Дифференциация по частотам аллелей креатинкиназы, как и по ряду морфологических признаков, может быть также связана с разным образом жизни речной и озерной ряпушек. Ведь креатинкиназа – один из основных ферментов, регулирующих метаболизм в мышцах, а сибирская и европейская ряпушки обитают в условиях, требующих разной мышечной активности. Прямых данных, которые свидетельствовали бы об отборе в пользу того или иного аллеля креатинкиназы в определенных условиях среды, пока нет, однако такую возможность необходимо учитывать при трактовке данных, касающихся распространения аллелей этого фермента. Отметим, что предположение об адаптивности распространения разных аллелей в популяциях в зависимости от условий обитания высказывалось в отношении локуса, кодирующего фермент лактатдегидрогеназу (LDH‑B2*) (Vuorinen et al., 1991).
Итак, адаптивная природа процесса формирования европейской ряпушки, по крайней мере для некоторых признаков, проявляется достаточно четко. Можно считать, что основная цель нашей работы достигнута. Однако нельзя не отметить, что факты, обнаруженные в ходе работы, заставляют усомниться в том, что европейская ряпушка полностью обособилась от сибирской и процесс видообразования завершен.
Так, хиатус между двумя изученными формами, как по морфологическим, так и по генетическим признакам, отсутствует; во многих популяциях встречаются особи с »промежуточными» признаками; в бассейне Печоры происходит гибридизация двух форм ряпушки. Особенно интересны »промежуточные» популяции из района водораздела Белого, Балтийского и Каспийского морей. Эти популяции, видимо, являются реликтом обширной доледниковой переходной зоны между европейской и сибирской формами ряпушек.
По нашему мнению, незавершенность процесса видообразования – в значительной степени неизбежная и даже необходимая характеристика »модельных» объектов, на которых оно изучается. Ведь именно наличие реликтовых переходных форм позволяет реконструировать процесс формирования новых видов. Ранее нами выявлена такая »незавершенность» у другого »модельного» объекта – атлантического лосося (Махров, 2005).
Библиография
Алексеева Я. И., Решетников Ю. С. Современная ихтиофауна Соловецких островов Белого моря // Первый конгресс ихтиологов России: Тез. докл. (Астрахань, сентябрь 1997 г.). М.: Изд-во ВНИРО, 1997. С. 7.
Антрошенко И. В., Буркова В. П., Ермакова О. Н., Козьмин А. К., Леженина Н. И., Шатова В. В. Лимнологическая характеристика и рыбохозяйственное использование озера Пильня // Тез. докл. респ. конф. по проблемам рыбохоз. исслед. внутр. водоемов Карелии, 14–16 нояб. 1979 г. Петрозаводск, 1979. С. 46–48.
Анухина А. М. Ихтиофауна Соловецких озер // Тр. СевНИОРХ. 1972. Т. 6. С. 94–110.
Атлас пресноводных рыб России / Под ред. Ю. С. Решетникова. М.: Наука, 2003. Т. 1. 379 с.
Берг Л. С. Рыбы пресных вод Российской империи. М.: Департамент земледелия, 1916. 563 с.
Берг Л. С. Рыбы пресных вод СССР и сопредельных стран. М.; Л.: Изд-во АН СССР, 1948. Ч. 1. 466 с.
Болотова Н. Л. Сибирская ряпушка Coregonus sardinella (Valenciennes, 1848) // Красная кн. Вологод. обл. Вологда: Вологод. гос. пед. ун-т, 2010. Т. 3. Животные. С. 102.
Боровикова Е. А. Особенности филогеографии сиговых рыб северо-запада европейской части России // Любищевские чтения – 2011: Сб. материалов междунар. конф. (Ульяновск, 5–7 апреля 2011 г.). Ульяновск: УлГПУ, 2011. с. 93-100.
Боровикова Е. А. Филогеография ряпушек Coregonus albula (L.) и C. sardinella (Valenciennes) Европейского Севера России: Автореф. дис. … канд. биол. наук. М., 2009. 24 с.
Боровикова Е. А., Алексеева Я. И., Артамонова В. С., Махров А. А., Шрейдер М. Ю. Систематическое положение ряпушки Соловецких озер (по морфологическим и генетическим данным) // Проблемы мониторинга природной среды Соловецкого архипелага: Материалы III Всерос. науч. конф. (Архангельск, 8–11 дек. 2008 г.). Архангельск, 2008. С. 20–22.
Боровикова Е. А., Балдина С. Н., Гордон Н. Ю., Махров А. А., Политов Д. В. Генетическое разнообразие, особенности морфологии и происхождения ряпушки оз. Водлозера // Водлозерские чтения: Естеств.-науч. и гуманитар. основы природоохран., науч. и просвет. деятельности на охраняемых природ. терр. Русского Севера: Материалы науч.-практ. конф., посвящ. 15-летию НП «Водлозерский». Петрозаводск: КарНЦ, 2006. С. 69–74.
Боровикова Е. А., Махров А. А. Обнаружение гаплотипа митохондриальной ДНК, характерного для сибирской ряпушки Coregonus sardinella (Valenciennes, 1848), в популяции европейской ряпушки C. albula (Linnaeus, 1758) Водлозера (бассейн Балтийского моря) // Изв. РАН. Сер. биологическая. 2009. № 1. С. 95–99.
Гордеева Н. В., Холод О. Н., Дворянкин Г. А., Сендек Д. С., Стерлигова О. П. О происхождении соловецкой ряпушки Coregonus albula и корюшки Osmerus eperlanus Сямозера // Вопр. ихтиологии. 2009. Т. 49. № 1. С. 28–36.
Дворянкин Г. А. Озера Соловецкого архипелага: особенности ихтиофауны и состояние промысла // Материалы отчет. сессии СевПИНРО по итогам НИР 2002–2003 гг. Архангельск: СевПИНРО, 2005. С. 239–246.
Дворянкин Г. А. Современное состояние ихтиофауны и пути управления рыбными ресурсами озер Кенозерского Национального парка: Автореф. дис. … канд. биол. наук. Калининград, 2010. 24 с.
Дворянкин Г. А., Козьмин А. К., Кулида С. В. Биология и промысел ряпушки оз. Лекшмозеро // Пробл. изучения, рацион. использования и охраны природ. ресурсов Белого моря: Материалы X Междунар. конф., 18-20 сент. 2007 г., Архангельск. Архангельск: Изд-во СГМУ, 2007. С. 113–116.
Дворянкин Г. А., Кулида С. В., Портнов В. В. Биология и рыбохозяйственное значение ряпушки Кенозерского Национального парка // Соврем. состояние вод. биоресурсов: Материалы науч. конф., посвящ. 70-летию С. М. Коновалова. Владивосток: ТИНРО-центр, 2008. С. 61–63.
Дрягин П. А. Белозерская ряпушка и вопрос акклиматизации сиговых в Белом озере // Изв. ВНИОРХ. 1933. Т. 16. С. 22–39.
Дрягин П. А., Пирожников П. Л., Покровский В. В. Полиморфизм сиговых рыб (Coregonidae) и его биологическое и рыбохозяйственное значение // Вопр. ихтиологии. 1969. Т. 9. Вып. 1. С. 14–25.
Есипов В. К. Ряпушка (Coregonus sardinella Valenciennes) северной части Обской губы и Гыданского залива // Тр. Науч.-исслед. ин-та поляр. земледелия, животноводства и промыслового хоз-ва. Сер. «Промысловое хоз-во». Л.: Изд-во Главсевморпути, 1941. Вып. 15. С. 7–36.
Захваткин А. А. Соловецкие озера. Соловки: Соловец. о-во краеведения, 1927. 142 с.
Зверева О. С., Кучина Е. С., Остроумов Н. А. Рыбы и рыбный промысел среднего и нижнего течения Печоры. М.; Л.: Изд-во АН СССР, 1953. 230 с.
Зверева О. С., Кучина Е. С., Соловкина Л. Н. Рыбные богатства Коми АССР и пути их освоения. Сыктывкар: Коми кн. изд-во, 1955. 106 с.
Иванова Н. В., Петрова Л. М. Некоторые данные о морфометрии ряпушки рек Печоры и Каратайки // Конф. молодых ученых ПИНРО по результатам исслед. 1971 г.: Тез. докл. Мурманск, 1972. С. 32–33.
Кесслеp К. Ф. Об ихтиологической фауне реки Волги // Тр. С.-Петерб. о-ва естествоиспытателей. 1870. Т. 1. Вып. 2. С. 236–310.
Козьмин А. К. Рыбные ресурсы рек и озер европейского северо-востока России: их сохранение и использование. Мурманск: Изд-во ПИНРО, 2011. 314 с.
Коновалов А. Ф., Болотова Н. Л. Жилые формы ряпушки на Европейском Севере России: Материалы Междунар. конф. Апатиты, 2004. С. 59–61.
Корнилова В. П. Результаты изучения сиговых рыб в Голодной губе дельты Печоры // Тр. Карел. отд-ния ГосНИОРХ. 1967. Т. V. Вып. 2. С. 102–111.
Кудерский Л. А. О путях проникновения сибирской ряпушки в водоемы Балтийского и Каспийского бассейнов // Изв. ГосНИОРХ. 1977. Т. 111. С. 54–62.
Кузищин К. В., Груздева М. А., Андреева А. П., Махров А. А., Голубев А. В., Новиков Г. Г. К вопросу о таксономическом статусе ряпушки (Coregonidae, Osteichthyes) Соловецких островов // Биол. основы изучения, освоения и охраны животного и раст. мира, почв. покрова Вост. Фенноскандии: Тез. докл. междунар. конф. и выезд. сес. Отд-ния общей биологии РАН, г. Петрозаводск, 6–10 сент. 1999 г. Петрозаводск, 1999. С. 135–136.
Кузнецов В. В. Определение видовой принадлежности и биометрическое исследование молоди сиговых рыб в дельте Печоры // Бюллетень МОИП. Отд. биол. 1987. Т. 92. Вып. 1. С. 50–58.
Кучина Е. С. Ихтиофауна притоков р. Усы // Рыбы бассейна р. Усы и их кормовые ресурсы. М.; Л.: Изд-во АН СССР, 1962. С. 176–211.
Лукаш Б. С. Рекогносцировочное рыбохозяйственное исследование Водлозера // Рыб. хоз-во Карелии. 1939. Вып. 5. С. 121–148.
Марченко Л. П. Ряпушка оз. Имандра // Биол. ресурсы Белого моря и внутр. водоемов Европейского Севера: Материалы семинара (Петрозаводск, 23–25 янв. 1980 г.). Петрозаводск, 1981. С. 74–75.
Махров А. А. «Диалектическое» видообразование: от кумжи (Salmo trutta L.) к атлантическому лососю (S. salar L.) // Эволюц. факторы формирования разнообразия живот. мира. М.: Т-во науч. изд. КМК, 2005. С. 248–256.
Махров А. А., Болотов И. Н. Пути расселения и видовая принадлежность пресноводных животных севера Европы (обзор молекулярно-генетических исследований) // Генетика. 2006. Т. 42. № 10. С. 1319–1334.
Махров А. А., Политов Д. В., Коновалов А. Ф., Болотова Н. Л., Думнич Н. В. Гибридная популяция европейской (Coregonus albula) и сибирской (C. sardinella) ряпушек в верховьях Волги // Биол. ресурсы Белого моря и внутр. водоемов Европ.о Севера: Тез. докл. междунар. конф. Сыктывкар, 11–15 февр. 2003 г. Сыктывкар, 2003. С. 56–57.
Михайловский М. Кое-что о промыслах на острове Колгуеве // Изв. Арханг. отд-ния Имп. о-ва судоходства. Год первый. 1903. С. 110–113.
Мухомедияров Ф. Б. Ряпушка соловецкая (бассейн Белого моря) // Пробл. использования промысловых ресурсов Белого моря и внутр. водоемов Карелии. 1963. Вып. 1. С. 206–210.
Новоселов А. П. Сиговые рыбы в озерах Европейского Северо-востока России // Биол. пробл. Севера. Современные проблемы сиговых рыб. Ч. 1. Материалы 4-го Всесоюз. совещания по биологии и биотехнике разведения сиговых рыб. Владивосток: ИБПС ДВО АН СССР, 1991. С. 23–37.
Первозванский В. Я. Рыбы водоемов района Костомукшского железорудного месторождения. Петрозаводск: Карелия, 1986. 216 с.
Перелыгин А. А. Популяционно-генетический анализ белков европейской (Coregonus albula) и сибирской (Coregonus sardinella) ряпушек: Автореф. дис. … канд. биол. наук. Новосибирск, 1988. 16 с.
Петрова Л. П., Кудерский Л. А. Водлозеро: природа, рыбы, рыбный промысел / НП «Водлозерский», Ин-т озероведения РАН. Петрозаводск: КарНЦ РАН, 2006. 196 с.
Покровский В. В. О морфологических особенностях, происхождении и географическом распространении беломорской ряпушки Coregonus sardinella maris-albi Berg // Изв. ГосНИОРХ. 1967. Т. 62. С. 100–114.
Пономарев В. И. Некоторые популяционные характеристики рыб разнотипных озер северной части Большеземельской тундры // Тр. Коми НЦ УрО РАН. 1996. № 147. С. 139–151.
Пономарев В. И. Рыбы предгорных озер водосбора реки Пага // Вестн. ОГУ. 2008. № 87. С. 96–100.
Попов И. Ю., Сендек Д. С. Квинтэссенция эволюции // Эволюц. биология: история и теория. СПб., 2003. Вып. 2. С. 172–189.
Правдин И. Ф. Руководство по изучению рыб. М.: Пищевая пром-сть, 1966. 376 с.
Пробатов А. Н. Материалы по научно-промысловому обследованию Карской губы и реки Кары. М.: Изд-во ВНИРО, 1934. С. 74–78.
Решетников Ю. С. Экология и систематика сиговых рыб. М.: Наука, 1980. 300 с.
Романов В. И. Морфоэкологическая характеристика ряпушки из оз. Томмот (бассейн р. Хатанги) и некоторые дискуссионные вопросы систематики евразийских ряпушек // Сибир. экол. журн. 2000. № 3. С. 293–303.
Сендек Д. С. О видовой принадлежности ряпушки, обитающей в реке Печоре // Сб. тр. ГосНИОРХ. 1998. Т. 323. С. 191–198.
Сендек Д. С., Новоселов А. П., Студенов И. И. Связана ли история становления популяций ряпушек восточного берега Белого моря с постледниковой гибридизацией европейской ряпушки (Coregonus albula) и сибирской ряпушки (Coregonus sardinella)? // Биология, биотехника разведения и состояние запасов сиговых рыб: Материалы седьмого междунар. науч.-производств. совещания / Под ред. А. И. Литвиненко, Ю. С. Решетникова. Тюмень: ФГУП Госрыбцентр, 2010. С. 59–64.
Сидоров Г. П. Рыбные ресурсы Большеземельской тундры. Л.: Наука, 1974. 164 с.
Соловкина Л. Н. Рыбы среднего и нижнего течения р. Усы // Рыбы бассейна р. Усы и их кормовые ресурсы. М.; Л.: Изд-во АН СССР, 1962. С. 88–135.
Соловкина Л. Н. Ряпушка Coregonus albula sardinella (Val.) бассейна Печоры // Вопр. ихтиологии. 1974. Т. 14. Вып. 5. С. 769–781.
Танфильев Г. И. О рыбных и звериных промыслах Мезенского и Печорского края // Вестн. рыбопром-сти. 1896. № 2–3. С. 53–64.
Туманов М. Д. Морфоэкологическая характеристика рыб нижнего течения р. Усы в условиях техногенного загрязнения (на примере сиговых, Coregonus): Автореф. дис. … канд. биол. наук. Томск, 2010. 24 с.
Туманов М. Д. Особенности морфологии, темпа роста и возрастной структуры полупроходной и речной форм ряпушки Coregonus albula (L.) бассейна реки Печора // Тр. КНЦ УрО РАН. 2002. № 170. С. 137–144.
Черняев Ж. А. Факторы и возможные механизмы, вызывающие изменения темпа эмбрионального развития костистых рыб (на примере сиговых Coregonidae) // Вопр. ихтиологии. 2007. Т. 47. № 4. С. 475–485.
Avise J. C. Molecular Markers, Natural History and Evolution. Second Edition. Sunderland, Massachusetts: Sinauer Associates, Inc. Publishers, 2004. 684 p.
Borovikova E. A., Schreider M. J., Makhrov A. A., Artamonova V. S. Populations of North-Eastern Europe with intermediate characteristics of vendace (Coregonus albula) and least cisco (C. sardinella) // 13th European Congress of Ichthyology. 6th–12th September. Klaipeda, Lithuania. Abstracts book. 2009. P. 17–18.
Coyne J. A., Orr H. A. Speciation. Sunderland, Massachusetts U. S. A.: Sinauer Associates, Inc., 2004. 545 p.
Rundle H. D., Nosil P. Ecological speciation // Ecology Letters. 2005. V. 8. P. 336–352.
Sendek D. S. Electrophoretic studies of Coregonid fishes from across Russia // Arch. Hydrobiol. Spec. Issues Advanc. Limnol. 2002. V. 57. P. 35–55.
Svendsen J. I., Alexanderson H., Astakhov V. I. et al. Late Quaternary ice sheet history of northern Eurasia // Quaternary Science Review. 2004. V. 23. P. 1229–1271.
Vuorinen J., Himberg M. K.-J., Lankinen P. Genetic differentiation in Coregonus albula (L.) (Salmonidae) populations in Finland // Hereditas. 1981. V. 94. P. 113–121.
Vuorinen J., Nasje T. F., Sandlund O. T. Genetic changes in a vendace Coregonus albula (L.) populations, 92 years after introduction // J. Fish Biology. 1991. V. 39. P. 193–201.
Благодарности
Авторы признательны Я. И. Алексеевой, В. С. Артамоновой, Ю. Ю. Дгебуадзе, С. П. Китаеву, А. К. Козьмину, Л. А. Кудерскому, К. В. Кузищину, М. В. Мине, А. П. Новоселову, Л. П. Петровой, В. Я. Первозванскому, Ю. С. Решетникову, В. И. Романову за обсуждение затронутых в работе тем и критические замечания. Подготовка работы осуществлялась при финансовой поддержке гранта РФФИ № 11-04-00697-а и программы «Живая природа: современное состояние и проблемы развития» (подпрограмма «Динамика и сохранение генофондов»).

 © 2011 - 2026
© 2011 - 2026