



Введение
На суточные и сезонные режимы активности ящериц большое влияние оказывают температуры воздуха и почвы. Но мы уже давно обращали внимание на то, что разные температуры окружающей среды могут функционально выступать для каких-то процессов или форм активности как оптимальные, как ограничивающие, как сигнальные, как опосредованные показатели тех или иных форм активности, физиологических состояний или циклов, как показатели сезонного состояния биотопа и фаз сезонных физиологических циклов у рептилий и т. п. При этом температура воздуха в приземном слое или на высоте 1–1.5 м над землей, температура поверхности почвы на горизонтальной поверхности или на склонах разной ориентации, на открытых участках или в тени, температура почвы на разных глубинах (5, 10, 20, 30, 50, 80, 100 см) несут разную функциональную нагрузку в экологии различных видов (Черлин, Целлариус, 1981; Черлин, Музыченко, 1983а, б; Черлин, 1988, 2021).
Кроме того, следует обратить внимание на то, что бархан Сарыкум – рефугиум среднеазиатской флоры и фауны на северо-востоке Кавказа, в течение десятков тысяч лет отделенный от исходной, «материнской» территории на сотни километров. Популяция ушастых круглоголовок, которая населяет бархан, длительное время изолирована от основного ареала и приобрела морфологические, физиологические, поведенческие особенности (Хонякина, 1965; Джамирзоев и др., 2011; Мазанаева, 2020). В связи с этими особенностями бархану был присвоен статус Особо охраняемой природной территории (ООПТ) и он был включен как отдельный участок в Дагестанский государственный заповедник.
Основной целью нашей работы было изучение значения разных участков биотопа ушастых круглоголовок с различными климатическими условиями в их экологии для реализации определенных физиологических процессов, сезонных циклов, поведенческих комплексов и состояний.
Материалы
Материалом для настоящей статьи явились два основных «блока»:
1) многолетние (более 55 лет) исследования одного из авторов (В. А. Черлина) по термобиологии и физиологической экологии разных групп и видов рептилий в естественных природных условиях в разных регионах СССР и Российской Федерации (Черлин, 1988, 2013а, б; Черлин, Музыченко, 1983а, б); в течение многих лет одним из важных объектов исследований были ушастые круглоголовки (Phrynocephalus mystaceus), физиологическую экологию которых изучали в пустынях Каракумы (Туркменистан) и Кызылкумы (Узбекистан) (Черлин, Музыченко, 1983а, 1988);
2) исследования физиологической экологии ушастых круглоголовок всеми авторами данной статьи на бархане Сарыкум в Республике Дагестан в течение 8 лет 9с 2018 по 2025 г.) (Окштейн и др., 2021; Окштейн, Черлин, 2021; Черлин и др., 2023; Черлин, Лабазанов (в печати)).
Методы
Методики полевых исследований, проводимых до 2022 г., подробно описаны нами в многочисленных публикациях и сведены воедино в отдельных книгах (Черлин, 2010, 2014). Основными элементами этих исследований были наблюдения за ящерицами на маршрутах, отлов и измерения температур их тела и окружающей среды, изучение динамики температур почвы и воздуха на метеорологических площадках непосредственно в местах работ. После чего полученные данные специальным образом обрабатывались, что также описано в предыдущих публикациях (Черлин, 2013в, 2014). Подробно рассматривать детали этой обработки в данной статье не имеет смысла, поскольку в ней такая обработка данных нами не применялась.
Исследования на бархане Сарыкум были начаты в 2018 г. с применением описанной выше методики, с отловами и измерениями температуры тела ящериц и внешней среды с помощью контактного двухканального измерителя температуры Mastech MS6512 фирмы Precision Mastech с крохотной термопарой типа К в качестве датчика. Но с 2022–2023 гг. мы начали дополнительно применять новые технические средства – логгеры температуры ДТН3-28 (производитель ООО ЭМБИ РЕСЕРЧ, Россия, г. Новосибирск) весом 0.8–0.9 г и объемом 0.6 см3 с продолжительностью автономной работы до 5 месяцев. А в 2024–2025 гг. мы начали использовать логгеры температуры ДТН-5-448 того же производителя. Эти логгеры имели те же размеры, что и предыдущие, но продолжительность их автономной работы была уже больше 6 месяцев (Petrovskii, Zavjalov, 2023). Кроме регистрации температуры тела, оба типа логгеров регистрировали и интенсивность двигательной активности ящериц. С соблюдением всех ветеринарных норм логгеры внедрялись интраперитонеально в ушастых круглоголовок на бархане Сарыкум (Окштейн, Черлин, 2021). Они давали возможность получать непрерывный график температуры тела и регистрировать динамику их активности вначале в течение 2–3 месяцев, а позже – в течение примерно полугода, включая период зимовки. Это уже были данные совершенно нового типа и качества.
В то же время на бархане, непосредственно в месте работ, нами устанавливалась автоматическая метеорологическая площадка. Она была составлена из нескольких логгеров температуры типа RC-4 фирмы Elitech, которые мы использовали как климатические приборы. Логгеры имели отдельные выносные датчики на мягком проводе длиной примерно 1 м. Они позволяли получать подробные графики температур непрерывно в течение более полугода. Логгеры регистрировали температуру приземного слоя воздуха на высоте 2 см, температуру поверхности почвы, температуру песка на разных глубинах, например 5, 30, 50 и 80 см. Один раз за каждый полевой сезон (весной или осенью) мы вырывали шурф глубиной 1 м и измеряли температуру на глубине 100 см. Чаще измерять температуру на больших глубинах не имело смысла, потому что колебаний температур в течение дней и недель там вообще не происходит (Черлин, Целлариус, 1981).
Кроме того, осенью 2025 года заместитель директора по науке Государственного природного заповедника «Дагестанский» Гаджибек Сефибекович Джамирзоев любезно предоставил нам во временное пользование тепловизионный бинокль ACCOLADE XP50 Фирмы «Pulsar». Этот прибор позволил не только инструментально зарегистрировать сам факт поздневечерней активности круглоголовок, но и изучить температурные условия этой активности, температурный баланс тела ящериц в это время, который в сумерках сильно отличается от дневного.
Результаты
Разные части биотопа, а следовательно, и их различные температурные условия могут иметь неодинаковое функциональное значение в экологии рептилий. В различных частях биотопа практически всегда имеется больша́я мозаичность температурных условий. Например, в верхней части бархана Сарыкум, где в основном живут ушастые круглоголовки, можно выделить: открытые песчаные пространства с горизонтальными поверхностями и со склонами разной ориентации по отношению к солнцу, с участками, в разной степени заросшими травянистой и кустарниковой растительностью, с открытыми в данный момент, т. е. освещенными солнцем или находящимися в тени, участками, с норами разной глубины, которые круглоголовки роют для разных целей, и т. п. Кроме того, все эти части биотопа существуют рано утром, днем в жару, вечером, в сумерках, в начале или середине ночи. То есть одни и те же микробиотопические участки в разное время суток могут иметь различные функции. Таким образом, внешняя среда предоставляет ящерицам огромное разнообразие температурных условий и возможных направлений их функционального использования. Более ранние наши исследования уже показали ряд вариантов функционального значения различных температурных факторов внешней среды (Черлин, Музыченко, 1983б).
Ограничения
1. Ряд внешних факторов ограничивают активность животных. Так, наши работы в Каракумах (Туркменистан) показали, что ушастые круглоголовки не могут быть активны на открытых пространствах с температурой поверхности песка выше 58–59 °С (в данной статье все температуры указываются в градусах Цельсия), поскольку пальцы этих ящериц не выдерживают более высокую температуру (Черлин, Музыченко, 1988).
2. Кроме того, в жаркое время круглоголовки также вынуждены уходить в убежища (в норы), когда температура воздуха в приземном слое и очень близкая к ней температура поверхности песка в тени начинают превышать 42–43°. Это объясняется тем, что максимальная температура тела у круглоголовок при полной активности как раз 42–44°. Чтобы остановить рост температуры тела в такое время, ящерицы в первую очередь уходят в тень, где она, в отсутствие прямого солнечного излучения, почти полностью соответствует температуре воздуха в приземном слое и температуре поверхности почвы в тени. Если же температура внешней среды в середине дня продолжает расти, то у ящериц нет никакой другой возможности не только снизить температуру тела или остановить ее рост, как только уйти с поверхности в норы (Черлин, Музыченко, 1983а). На бархане Сарыкум, по нашим наблюдениям, ушастые круглоголовки также не выходят на открытые пространства с температурой поверхности песка выше 58–59°.
Таким образом, оба указанных ограничения существенно влияют на режим активности ящериц.
Формирование циклов активности
Указанные в предыдущем абзаце ограничения функционально проявляются в том, что ушастые круглоголовки вынуждены либо уходить с открытых участков песка в тень, либо вообще покидать поверхность почвы, что, естественно, отражается на суточном режиме их активности.
На бархане Сарыкум максимум температуры поверхности почвы (65–70°) наступает примерно в 12–14 часов, минимум (25–28°) – около 4–6 часов утра. Амплитуда колебаний температуры поверхности почвы составляет примерно 40° или даже немного больше. Уходя в норы, круглоголовки оказываются на глубине около 20–30 см. Динамика температуры почвы в норах, где ящерицы проводят бо́льшую часть времени, соответствует температуре почвы на этих же глубинах. Амплитуда суточных колебаний температуры в этой зоне существенно меньше (всего 4–5°): максимум около 33–34°, минимум – примерно 29–30°. При этом экстремумы температуры наступают на этой глубине существенно позже, чем на поверхности, т. е. минимум – около 10–11 часов, а максимум – в 19–20 часов (рис. 1).
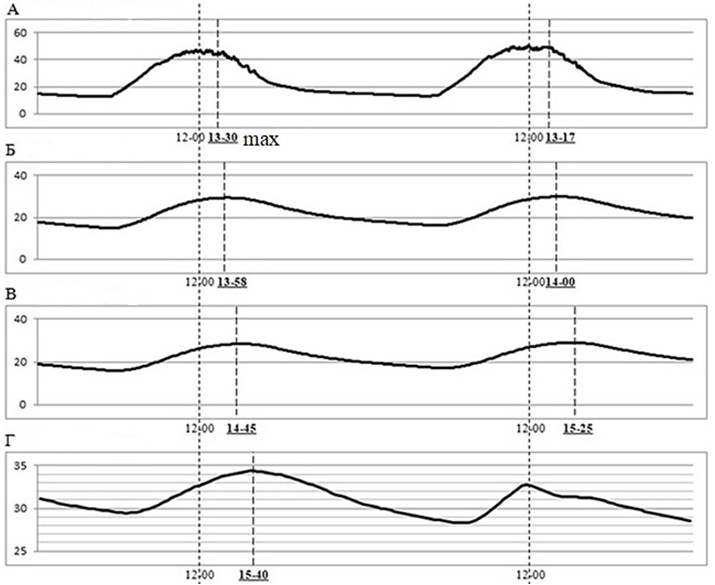
Рис. 1. Распределение и динамика температур 20–21.05.2024 на поверхности песка (А) и на разных глубинах – 5 (Б), 10 (В) и 20 см (Г) на бархане Сарыкум
Fig. 1. Distribution and dynamics of temperature on 20–21.05.2024 on the surface (A) and in the soil at different depths – 5 (Б), 10 (В) и 20 см (Г) in spring on the Sarykum dune
Еще более отчетливо эта закономерность видна в форме, представленной на рис. 2.
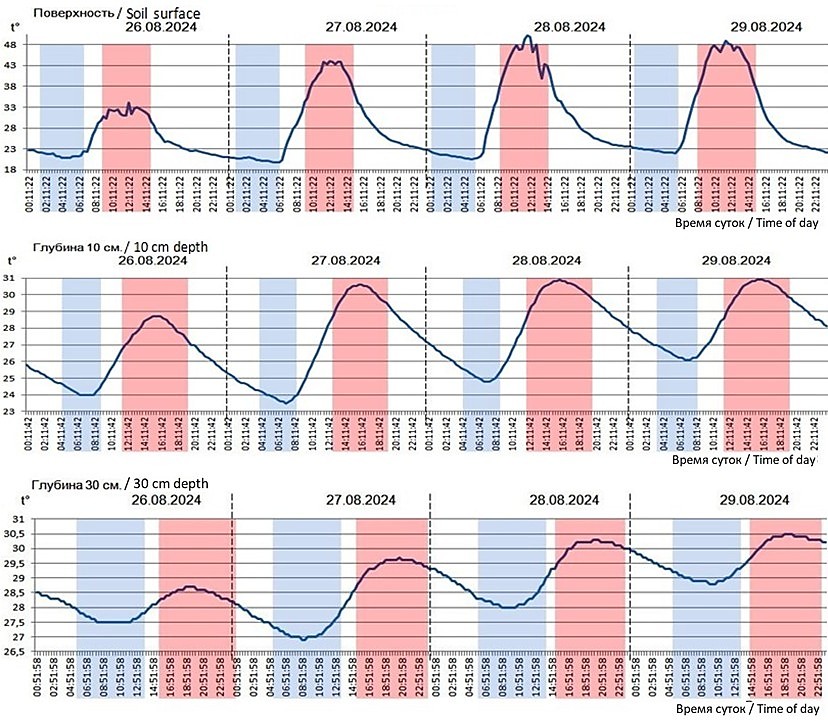
Рис. 2. Распределение и динамика температуры на поверхности и в почве на разных глубинах (10 и 30 см) осенью на бархане Сарыкум, полученные с помощью климатических логгеров. Розовыми полосами отмечены периоды, в которые наблюдались максимумы температуры на соответствующих глубинах, голубыми полосами – минимумы
Fig. 2.The distribution and dynamics of temperature on the surface and in the soil at different depths (10 and 30 cm) in autumn on the Sarykum dune, obtained using climate loggers. Pink bars indicate periods of maximum temperatures at the corresponding depths, and blue bars indicate minimum temperatures
На рис. 3 показано, как температурные факторы внешней среды достаточно жестко определяют возможные время и место нахождения ушастых круглоголовок (Phrynocephalus mystaceus) на бархане Сарыкум прохладной весной и жарким летом, т. е. как они формируют структуру их пространственно-временной активности, их экологические проявления.
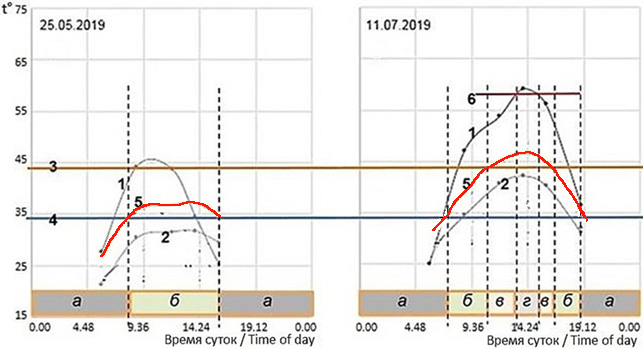
Рис. 3. Прогноз динамики режимов активности ушастых круглоголовок весной и летом на бархане Сарыкум. t° – температура; 1 – температура поверхности песка; 2 – температура воздуха на высоте 2 см; 3 – максимальная температура тела при полной активности у ушастых круглоголовок; 4 – минимальная температура тела при полной активности у ушастых круглоголовок; 5 – (красная линия) расчетная температура тела ушастых круглоголовок (расчетная температура тела = температура воздуха на высоте 2 см + 5°); 6 – критический уровень температуры поверхности почвы (58–59°), выше которого круглоголовки не могут находиться на поверхности. а – ночной покой, б – термостабилизирующее поведение (форма активности, при которой ящерицы реализуют комплекс поведенческих актов, направленных на удовлетворение своих индивидуальных потребностей, видоспецифичных потребностей жизнеобеспечения популяции и взаимодействие с биотическим окружением, т. е. выполняют активную функцию в биоценозе – питание, внутривидовые коммуникации, связанные с размножением, территориальностью, поддержанием социальной структуры популяции и т. п., межвидовые коммуникации – охрана территории и т. п.; температура их тела при этом активно удерживается или пассивно оказывается в определенном нешироком диапазоне температур полной активности, внутри которого стремится оказаться в еще более узких рамках – в диапазоне температур термостабилизации (Черлин, 2014)); в – активность преимущественно в тени, г – нахождение исключительно в тени или уход в норы
Fig. 3. Forecast of the dynamics of the modes of activity of toad-headed agamas (Phrynocephalus mystaceus) in spring and summer on the Sarykum dune. t° is the temperature; 1 – the sand surface temperature; 2 – the air temperature at a height of 2 cm; 3 – the maximum body temperature at full activity in toad-headed agamas; 4 – the minimum body temperature at full activity in toad-headed agamas; 5 – (red line) the calculated body temperature of toad-headed agamas (calculated body temperature = air temperature at a height of 2 cm + 5°); 6 – the critical level of soil surface temperature (58–59°), above which agamas cannot be on the surface. a – nocturnal rest, б – thermostabilizing behavior (a form of activity in which lizards implement a set of behavioral acts aimed to realize their individual needs, species-specific life support needs of the population and interaction with the biotic environment, i. e. they perform an active function in the biocenosis – nutrition, intraspecific communications related to reproduction, territoriality, maintenance of the social structure of the population, etc., interspecific communications – protection of the territory, etc.; at the same time, their body temperature is actively maintained or passively appears in a certain narrow temperature range of full activity, within which it tends to be in an even narrower range – in the temperature range of thermal stabilization (Cherlin, 2014)); в – activity mainly in the shade, d – activity exclusively in the shade or going into burrows
Данная статья и многие другие опубликованные материалы – пример того, как динамика температур внешней среды, с одной стороны, и физиологические ограничения, с другой стороны, организуют пространственно-временную структуру суточной активности отдельных видов рептилий (Черлин, Целлариус, 1981; Черлин, 1988, 2019, 2021). В целом мы можем выявить несколько основных направлений проявления таких взаимодействий.
Оптимизация
В теплый период в активности ушастых круглоголовок имеется ряд периодов, в которые они стараются реализовать определенные физиологические потребности. Так, днем, особенно когда температура окружающей среды повышается настолько, что начинает угрожать нормальной жизнедеятельности или даже жизни круглоголовок, ящерицы либо переходят в тень, либо закапываются в верхние слои песка, либо уходят в норы. Это период дневного отдыха. И в это время, в любом случае, они стараются удерживать температуру своего тела примерно на некотором оптимальном уровне. На рис. 4 коричневым овалом выделен такой период дневного отдыха, в течение которого круглоголовки, находясь в норе, удерживают температуру примерно на одном уровне – около 35°.
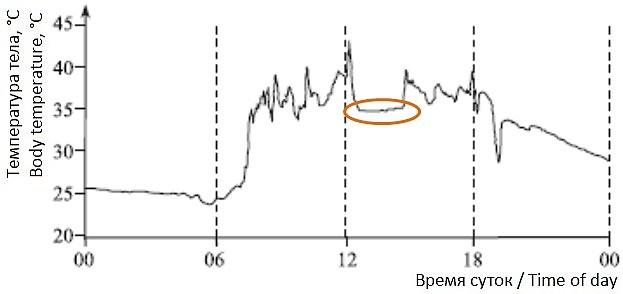
Рис. 4. Температура тела ушастой круглоголовки 31.08.2022, зафиксированная с помощью логгера
Fig. 4. Body temperature of the toad-headed agama in August 31, 2022, recorded by the logger
Регулирующие реакции
Эта регуляция возможна исключительно вследствие наличия тепловой мозаичности внешней среды, которая используется рептилиями для поведенческой регуляции температуры тела.
Длительные наблюдения над динамикой температуры тела ушастых круглоголовок и внешней среды, полученные с помощью логгеров на бархане Сарыкум (рис. 5), показывают, что, несмотря на изменчивость погодных условий и температурных показателей в разные дни, температура тела круглоголовок удерживается в определенном довольно узком диапазоне температур полной активности – примерно 35–42° (Черлин, 2025). Следовательно, мы имеем дело с хорошо развитыми и организованными регуляторными реакциями.
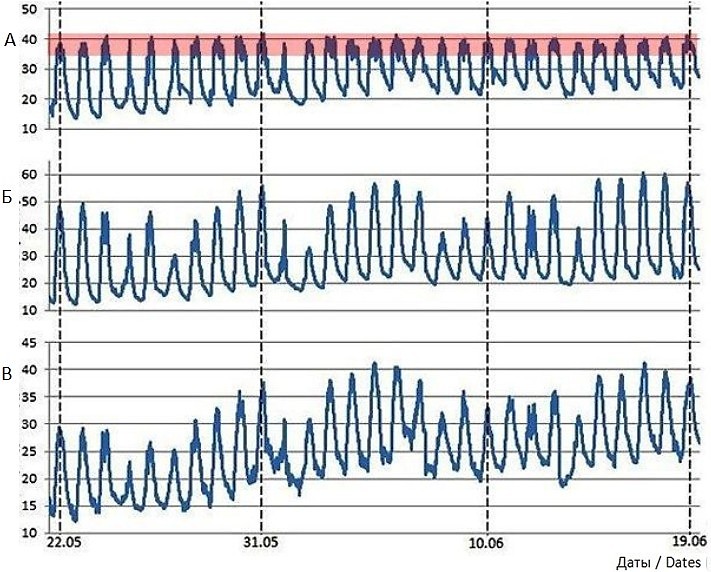
Рис. 5. Динамика температуры тела ушастой круглоголовки, Phrynocephalus mystaceus, полученная с помощью внедренного интраперитонеально логгера, и динамика температур внешней среды, полученных с помощью климатических логгеров, с 22 мая по 19 июня 2024 г. на бархане Сарыкум (Республика Дагестан, РФ) (по: Черлин, 2025). А – температура тела, Б – температура поверхности почвы (песка), В – температура воздуха на высоте 3 см от поверхности песка. Розовая полоса выделяет примерный температурный диапазон полной активности у круглоголовок
Fig. 5. Dynamics of the body temperature of the toad-headed agama, Phrynocephalus mystaceus, obtained by using an intraperitoneally embedded logger, and the dynamics of ambient temperatures, obtained by using climatic loggers from May 22 to June 19, 2024 on the Sarykum dune (Republic of Dagestan, Russian Federation) (according to Cherlin, 2025). A – the body temperature, Б – the soil surface temperature, В – the air temperature at a height of 3 cm above the surface of the sand. The pink stripe highlights the approximate temperature range of full activity in toad-headed agamas
Одни и те же участки (микробиотопы) могут использоваться ящерицами для нагрева и остывания в разных режимах. Так, весной и осенью, т. е. в более прохладные сезоны, круглоголовки нагреваются на открытых участках («режим грелки»). Летом, в самую жару, находиться на открытых участках для ящериц становится невозможно (об этом мы писали выше), и они вынуждены переходить в «режим противодействия нагреванию», для чего уходят в полутень, в ажурную тень, в плотную тень, забираются на пучки травы, закапываются в верхний слой песка, уходят в норы, т.е. уходят в любое место, где в данное время тепловая нагрузка меньше. В это жаркое время в тени, в верхних слоях песка и в норах прохладнее, чем на поверхности. Другими словами, «грелками» тогда являются открытые поверхности песка, а участки в тени, верхние слои почвы и норы служат местами для противодействия нагреванию.
Но летом и в начале осени, когда погода еще теплая, на бархане Сарыкум нами было описано удивительное явление – поздневечерняя и ранненочная активность ушастых круглоголовок (Окштейн, Черлин, 2021). В это время в сумерки, в плотные сумерки и уже практически в темноте эти вроде бы совершенно дневные ящерицы выходили из своих убежищ (выкапывались из верхних слоев песка или вылезали из нор) и некоторое время, иногда уже практически в темноте, вели активность, осуществляя все те же действия, что и днем, но только при температурах тела примерно на 5–10° ниже, чем днем. В этот теплый период соотношения температур тела и внешней среды менялись. Поздно вечером и в начале ночи температура на поверхности песка составляла вскоре после захода солнца примерно 26–30°, позже она постепенно опускалась и становилась на несколько градусов ниже. Но при этом температура в слое песка на глубине 5 см, куда в основном закапываются круглоголовки, была около 30–32°. Получалось, что круглоголовки, начиная свою вечернюю активность, оказывались на несколько градусов теплее, чем температуры среды на поверхности (рис. 6). Немного позже они постепенно остывали, но затем либо «прикапывались» в верхний слой песка, либо уходили в норы, где в это время было существенно теплее. То есть они использовали эти слои почвы как «грелки». Такие ситуации проиллюстрированы на рис. 6.
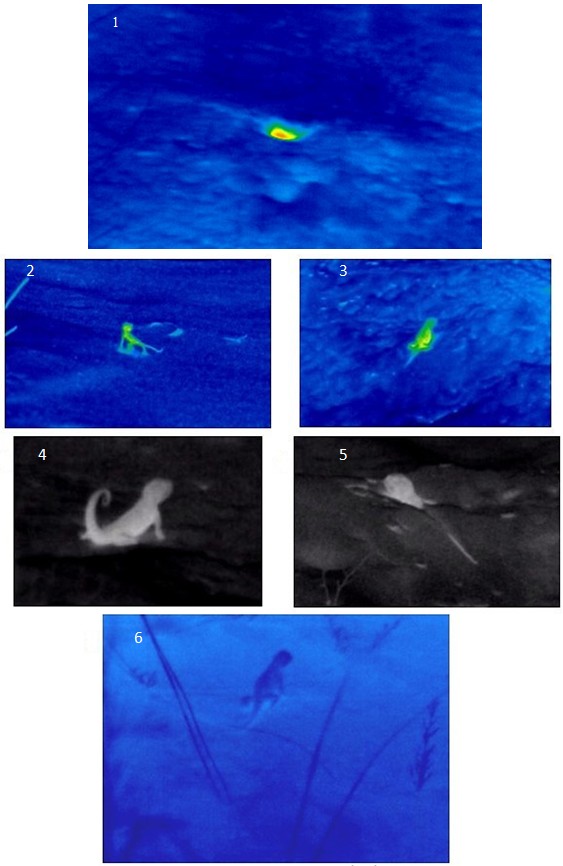
Рис. 6. Фотографии поздневечерней активности ушастых круглоголовок, выполненные И. Л. Окштейном с помощью тепловизионного бинокля. Комментарии к отдельным фотографиям приведены в тексте. 1 – теплая нора ярко выделяется на фоне прохладной поверхности песка; 2 – сеголеток ушастой круглоголовки имеет более нагретое ядро тела; 3 – круглоголовка имеет более нагретую нижнюю часть тела, поскольку она явно недавно сидела на более разогретом участке, который раскопала; 4 – нагретая взрослая круглоголовка сидит на прохладном песке; 5 – на фоне прохладной поверхности песка торчит передняя часть туловища круглоголовки, нагретая в верхних, еще теплых слоях песка; 6 – взрослая круглоголовка, которая, видимо, некоторое время была активной на поверхности, остыла в более прохладном воздухе и в данный момент оказалась прохладнее, чем поверхность песка
Fig. 6. Photographs of the late evening activity of toad-headed agamas, taken by I. L. Okshtein using Thermal imaging binocular. 1 – the warm burrow stands out vividly against the background of the cool sand surface; 2 – the fingerling of the toad-headed agama has a warmer body core; 3 – the toad-headed agama has a warmer lower part of its body, since it has clearly recently been sitting on a warmer area that it had excavated; 4 – a heated adult toad-headed agama is sitting on cool sand; 5 – the front part of the body of the toad-headed agama sticks out against the background of the cool sand surface, heated in the upper, still warm layers of sand; 6 – an adult toad-headed agama, which, apparently, had been active on the surface for some time, then cooled down in cooler air and at the moment turned out to be cooler than the sand surface
Показатели сезонного состояния биотопа и фаз сезонных физиологических циклов у рептилий
Если исследователь приезжает в какое-либо место, например весной на бархан Сарыкум в Дагестане, и обнаруживает, что там уже тепло, температура воздуха достигает 28–30°, а температура поверхности песка в середине дня 45–50°, то он может предположить, что здесь немного ранее прошел достаточно длительный период с теплой погодой, что рептилии уже давно вышли, у них в разгаре половая активность, самки начинают беременеть и т. п. Но на самом деле весной погода довольно изменчивая, и вполне вероятно, что предыдущее время было холодное, а теплая погода началась только с неделю назад, и рептилии еще совсем недавно вышли из зимовки. Как это распознать? Как точно определить фенологическую фазу на настоящий момент? Как понять, в каком физиологическом состоянии могут находиться в данный момент, например, ушастые круглоголовки на бархане?
Оказывается, возможность это оценить есть. Для этого имеет смысл изучить динамику температур в почве на глубине 80–100 см (рис. 7).
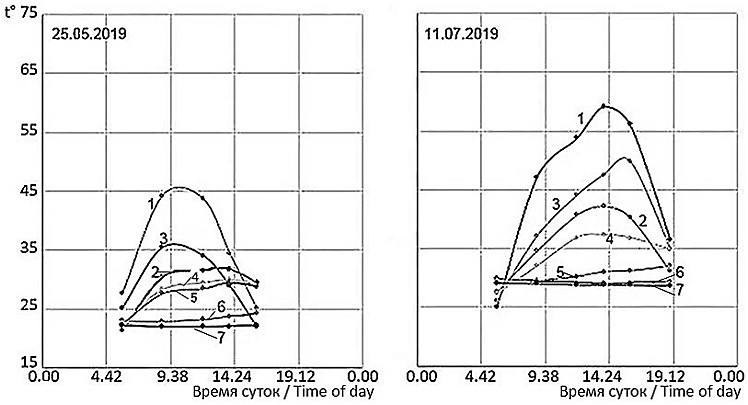
Рис. 7. Динамика климатических температур (пример типичных дней) в мае и июле на бархане Сарыкум. t° – температура; 1 – температура поверхности песка; 2 – температура воздуха на высоте 3 см; 3 – температура почвы на глубине 5 см; 4 – температура почвы на глубине 10 см; 5 – температура почвы на глубине 30 см; 6 – температура почвы на глубине 50 см; 7 – температура почвы на глубине 80 см
Fig. 7. The dynamics of climatic temperatures (example of typical days) in May and July on the Sarykum dune. t° – the temperature; 1 – the temperature of the sand surface; 2 – the air temperature at a height of 3 cm; 3 – the soil temperature at a depth of 5 cm; 4 – the soil temperature at a depth of 10 cm; 5 – the soil temperature at a depth of 30 cm; 6 – the soil temperature at a depth of 50 cm; 7 – the soil temperature at a depth of 80 cm
Из рис. 7 видно, что суточная динамика температур приземного слоя воздуха и почвы на небольших глубинах (примерно до 30 см) вполне отчетливая, она может быть изменчивой в связи с погодными колебаниями, ярко проявляющимися прежде всего в весенний и осенний периоды. А вот суточной динамики температуры в почве уже на глубинах примерно от 50 см и глубже вообще не прослеживается. Зато общий уровень этих температур на глубине 80 см с ранней весны (с конца марта – апреля) до середины лета и начала осени (до июля – начала августа) постепенно возрастает с 18–20° до 30–32°. И даже если на поверхности погода в это время изменчивая и неустойчивая, температуры на глубине 80–100 см будут показывать эти плавные, закономерно-сезонные изменения. Таким образом, температуры почвы на глубинах 80–100 см могут служить верным показателем фенологической фазы в сезоне. Что бы исследователь ни застал на поверхности, приехав на бархан, – холод или тепло, температура на этих больших глубинах четко покажет: сейчас фенологическая весна, лето или осень, чего можно ждать, например, от ящериц – бурной половой активности, встреч беременных самок, новорожденных, активного питания, вечерней активности и т. п. Так, например, застав весной на поверхности теплую погоду, но зная, что на глубине 80 см в это время температура 21°, исследователь должен понимать, что сейчас, несмотря на тепло, все равно еще ранняя весна со всеми соответствующими этому фенологическими и физиологическими процессами.
Другими словами, не бархане Сарыкум (как и в других пустынных регионах) температуры почвы на глубинах 80–100 см достаточно точно показывают (как индикатор), в какой фенологической фазе находится природа и в какой фазе сезонных физиологических циклов находятся живущие тут рептилии.
Обсуждение
Значение температуры приземного слоя воздуха
Температура приземного слоя воздуха (на высоте 2–3 см, т. е. в слое, в котором реально живут круглоголовки) также играет большую регулирующую роль. Она эффективно влияет на тепловой баланс тела, позволяя ящерицам вести полную активность при 36–39°, но не давая им быть активными, если она повышается до 41–42° и выше.
Значение температуры поверхности почвы
Как мы уже описывали выше, температура поверхности почвы (песка) эффективно участвует в формировании пространственно-временной структуры суточной активности ушастых круглоголовок. Она ограничивает и модифицирует активность ящериц, по крайней мере – сверху, но, скорее всего, и снизу тоже. Высокие температуры поверхности (выше 58–59°) не дают круглоголовкам находиться на открытых пространствах, «загоняют» их в тень, иногда даже заставляют забираться на пучки травы или на упавшие ветки, закапываться в верхние слои песка или уходить в норы.
Значение температуры на глубине 5–10 см
Одной из важных особенностей поведения ушастых круглоголовок является то, что при опасности и в ситуациях, когда им просто надо уходить с поверхности в жару, они боковыми движениями тела закапываются, как будто «тонут» в песке. Они оказываются на глубине около 5 см, редко – до 10 см. В этом положении и в этом слое песка они проводят довольно много времени. Поэтому знание температурных условий на этой глубине и их суточной динамики очень важно для изучения экологии ушастых круглоголовок.
Весной этот слой песка еще недостаточно прогревается. Границы суточных колебаний температуры в нем могут быть примерно от 10–12 до 30°, а суточная амплитуда колебаний около 15°. В это время круглоголовки большую часть времени проводят на открытых пространствах на поверхности песка, греясь и ведя активность на солнце. Важно то, что для правильного созревания половых продуктов ящерицам, как и всем рептилиям, требуется, чтобы хотя бы часть времени они проводили в достаточно холодных условиях (Angelini et al., 1979, 1982; Action…, 1981; Callard et al., 1972; Licht, 1973; Bradshaw et al., 1980; Cowles, Bogert, 1944; Lang, 1979; Matz, 1984; Regal, 1966; Saint-Girons, 1982; Witten, Heatwole, 1978). Именно такая ситуация складывается у них весной: уходя с поверхности в норы или закапываясь в верхний слой песка, они попадают в холодные условия. И именно в этот период у них начинается половая активность, у самцов и у самок созревают половые продукты.
Летом в этом верхнем слое песка температуры колеблются примерно от 23–25 до 39–43° с суточной амплитудой колебаний около 10–15°. Таким образом, общий уровень температуры в этом слое повышается до уровня, близкого к предпочитаемому (37–39°), низких температур в то время уже не наблюдается. Все это позволяет ящерицам заняться восстановлением сил после сезона половой активности, а беременным самкам – выносить яйца.
Осенью температуры в слое песка на глубине 5–10 см постепенно понижаются, приближаясь постепенно к весенним значениям.
Значение температуры на глубине 20–30 см
Как мы уже упоминали выше, 20–30 см – это глубина, на которой в основном залегают норы ушастых круглоголовок. Весной в течение дня температуры там колебались примерно от 17 до 28°, а суточная амплитуда колебаний составляла около 7°. В середине лета на глубине 20 см температура была ~25–36° с суточной амплитудой 7–8°. К осени почва прогревалась на достаточно большую глубину, вследствие чего суточная амплитуда колебаний температуры даже в верхнем слое (~ до 30 см) уменьшалась. В результате с середины августа и в сентябре температура на глубине 20 см находилась в диапазоне примерно 27.5–30.0°, а суточная амплитуда колебаний была около 2.0–2.5°. На глубине 30 см эти значения были соответственно 27.5–28.5° с амплитудой около 1°. Такая температурная динамика поддерживала сезонные физиологические циклы, о чем мы упоминали в предыдущем разделе.
Значение температуры на глубине 50–80 см
Нор ушастых круглоголовок на глубинах 50 см и глубже мы не обнаружили. Температуры на этих и бо́льших глубинах в биологии этих ящериц на бархане Сарыкум, видимо, существенного значения не имеют. Но исследователем они могут быть использованы как косвенные показатели фенологического состояния природной среды и фазы сезонного цикла жизнедеятельности ящериц, что было показано выше.
Заключение
Для того чтобы ушастые круглоголовки, как и любые другие виды рептилий, могли успешно поддерживать подходящие им параметры жизнедеятельности и реализовывать необходимые сезонные физиологические циклы, должна существовать гармоничная и оперативная связь между эндогенными свойствами животных (параметрами гомеостаза) и характеристиками внешней среды. Характеристики внешней среды напрямую влияют на тепловой баланс тела, т. е. на температуру тела рептилий. Температурные условия внешней среды достаточно разнообразны, их конкретные значения, закономерные суточная и сезонная динамики во всей зоне, задействованной круглоголовками для своей активности (от температуры приземного слоя воздуха на высоте 2–3 см до температур на поверхности песка и на разных глубинах, которые по-разному используются ящерицами для своей жизнедеятельности), предоставляют рептилиям богатый выбор, позволяющий регулировать температуру тела и организовывать их необходимые суточную и сезонную динамики, регулировать и способствовать протеканию у них сезонных физиологических циклов.
Библиография
Джамирзоев Г. С., Яровенко Ю. А., Букреев С. А. Результаты инвентаризации фауны наземных позвоночных животных участка «Сарыкумские барханы» ГПЗ «Дагестанский» // Труды Государственного природного заповедника «Дагестанский». 2011. № 4. С. 172–179.
Мазанаева Л. Ф. Ушастая круглоголовка, Phrynocephalus mystaceus (Pallas, 1776) // Красная книга Республики Дагестан. Махачкала: Тип. ИП Джамалудинов М. А., 2020. С. 521–523.
Окштейн И. Л., Черлин В. А. Необычная вечерняя активность ушастых круглоголовок (Phrynocephalus mystaceus) на бархане Сарыкум (Республика Дагестан, Российская Федерация) // Принципы экологии. 2021. Т. 11, № 4. С. 57–79.
Окштейн И. Л., Черлин В. А., Вето Д. В., Кожара В. В., Корнеева А. А., Ложкина А. В., Мироненко-Маренков А. Д., Рябова Е. Д., Тищенко М. А., Турова М. А., Фраерман А. Ю., Яцук А. А. Сумеречная активность ушастых круглоголовок (Phrynocephalus mystaceus) на песчаном массиве Большой Сарыкум в Дагестане // Вопросы герпетологии: Программа и тезисы докладов VIII съезда Герпетологического общества имени А. М. Никольского при РАН «Современные герпетологические исследования Евразии» 3–9 октября 2021 г. М.: КМК Scientific Press, 2021. С. 199–201.
Черлин В. А. К термобиологии серого геккона (Cyrtopodion russowi), полосатой ящурки (Eremias scripta) и степной агамы (Trapelus sanguinolentus) в Восточных Каракумах // Известия Академии наук Туркменской ССР. Серия биологических наук. 1988. № 5. С. 36–43.
Черлин В. А. Термобиология рептилий. Общие сведения и методы исследований (руководство) . СПб.: Русско-Балтийский информационный центр «БЛИЦ», 2010. 124 с.
Черлин В. А. Сложности и возможные ошибки при полевых исследованиях по термобиологии рептилий // Современная герпетология: проблемы и пути их решения: Статьи по материалам докладов Первой международной молодежной конференции герпетологов России и сопредельных стран (Санкт-Петербург, Россия, 25–27 ноября 2013 г.) / Зоологический институт РАН. СПб., 2013а. С. 32–39.
Черлин В. А. Сравнение термобиологии сцинкового (Teratoscincus scincus scincus) и гребнепалого (Crossobamon eversmanni) гекконов // Вестник Тамбовского университета. Сер. Естественные и технические науки. 2013б. Т. 18, вып. 6. С. 3110–3112.
Черлин В. А. Сравнение термобиологии трех видов змей Средней Азии (Echis multisquamatus, Spalerosophis diadema, Psammophis lineolatum) // Працi Украïнського герпетологiчного товариства. 2013в. № 4. С. 184–189.
Черлин В. А. Рептилии: температура и экология . Saarbrücken: Lambert Academic Publishing, 2014. 442 с.
Черлин В. А. Термобиологический механизм пространственно-временного разобщения активности средней, Eremias intermedia, и линейчатой, E. lineolata, ящурок (Reptilia, Lacertidae) в Кызылкумах // Известия высших учебных заведений. Поволжский регион. Естественные науки. 2019. № 1. С. 162–174.
Черлин В. А. Термобиология и экология четырех видов агамовых ящериц в Средней Азии и на Северном Кавказе // Горные экосистемы и их компоненты: Материалы VIII Всероссийской конференции с международным участием, посвященной Году науки и технологий в Российской Федерации. 10–25 сентября 2021. Нальчик, 2021. С. 124–125.
Черлин В. А. Приспособления разных эктотермных животных к глобальным изменениям климата // Принципы экологии. 2025. № 3. С. 4–39.
Черлин В. А., Лабазанов И. М., Значение ветра в экологии ушастых круглоголовок (Phrynocephalus mystaceus) на бархане Сарыкум (Республика Дагестан, Российская Федерация) // Принципы экологии. 2025 (в печати).
Черлин В. А., Музыченко И. В. Термобиология и экология сетчатой ящурки (Eremias grammica), ушастой (Phrynocephalus mystaceus) и песчаной (Ph. interscapularis) круглоголовок летом в Каракумах // Зоологический журнал. 1983а. Т. 62, № 6. С. 897–908.
Черлин В. А., Музыченко И. В. Использование нор в терморегуляции рептилий // Прикладная этология: Материалы III Всесоюзной конференции по поведению животных. Т. 3. М.: Наука, 1983б. С. 172–174.
Черлин В. А., Окштейн И. Л., Алигаджиев Э. Термобиологические характеристики ушастых круглоголовок (Phrynocephalus mystaceus) (Reptilia, Agamidae) на бархане Сарыкум (Дагестан, Россия), полученные с помощью логгеров температуры // Современная герпетология. 2023. Т. 23, вып. 3/4. С. 160–163.
Черлин В. А., Целлариус А. Ю. Зависимость поведения песчаной эфы, Echis multisquamatus Cherlin, 1981 от температурных условий в Южной Туркмении // Фауна и экология амфибий и рептилий палеарктической Азии. Л.: Наука, 1981. С. 96–108. (Труды Зоологического института АН СССР, т. 101.)
Хонякина З. П. Некоторые морфолого-меристические особенности ушастой круглоголовки Дагестана // Зоологический журнал. 1965. Т. XLIV, вып. 8. С. 1272–1274.
Action des facteurs externes sur les cycles de reproduction. Les strategies de reproduction. 2 part // Bull. Soc. Zool. France. Vol. 106, No 4. P. 398–502.
Angelini F., Botte V., D´Alterio E. Autumn photothermal regimes and reproductive functions in the female lizard Podarcis s. sicula Raf. // Monitore Zoologico Italiano, Firenze, (N.S.). 1982. Vol. 16. P. 133–148.
Angelini F., Brizzi R., Barone C. The annual spermatogenetic cycle of Podarcis sicula campestris De Betta (Reptilia, Lacertidae). I. The spermatogenetic cycle in nature // Monitore Zoologico Italiano, Firenze, (N.S.). 1979. Vol. 13. P. 279–301.
Callard J. P., Chan S. W. C., Potts M. The control of the reptilian gonsad // Amer. Zool. 1972. Vol. 12, No 2. P. 273–287.
Cowles R. B., Bogert C. M. Preliminary study of the thermal requirements of desert reptiles // Bulletin of the American Museum of Natural History. 1944. No 83. P. 261–296.
Bradshaw S. D., Gans C., Saint-Girons H. Behavioral thermoregulation in a Pygopodid lizard, Lialis burtonis // Copeia. 1980. No 4. P. 738–743.
Lang J. W. Thermophilic response of the American alligator and the American crocodile to feeding // Copeia. 1979. P. 48–59.
Licht P. Photoperiodic and thermal influences on reproductive cycles in reptiles // Endocrinology. Amsterdam; New-York, 1973. P. 185–190.
Matz G. La reproduction des reptiles et les facteurs de son induction // Acta zool. et pathol. Antverp. No 78. Mainten. and reprod. reptile captive. 1984. Vol. 1. P. 33–68.
Regal P. J. Thermophilic response following feeding in certain reptiles // Copeia. 1966. No 3. P. 588–590.
Petrovskii D., Zavjalov E. Development and validation of an implantable sensor for measuring activity and body temperature for long-term studies in small wild and laboratory animals // Biological Rhythm Research. 2023. Vol. 54, No 9. P. 523–534.
Saint-Girons H. Reproductive cycles of male snakes and their relationships with climate and female reproductive cycles // Herpetologica. 1982. Vol. 38, No 1. P. 5–16.
Witten G. J., Heatwole H. Preferred temperature of the agamid lizard Amphibolurus nobbinobbi // Copeia. 1978. No 2. P. 362–364.
Благодарности
Выражаем свою искреннюю благодарность заведующей кафедрой зоологии и физиологии Дагестанского государственного университета Людмиле Фейзулаевне Мазанаевой за организационную, информационную помощь и научную поддержку нашей работы, а также заместителю директора по науке Государственного природного заповедника «Дагестанский» Гаджибеку Сефибековичу Джамирзоеву за организационную помощь, поддержку работы в условиях бархана Сарыкум на территории заповедника и за предоставление тепловизионной техники для осуществления исследований.

 © 2011 - 2026
© 2011 - 2026