



Введение
В углеродном цикле лесных экосистем подземной фитомассе придается особое значение, т. к. углерод при отмирании и разложении корней переходит непосредственно в подстилку и нижележащие почвенные слои.
В настоящее время исследований по оценке вклада корней травяно-кустарничкового яруса (ТКЯ) и тонких корней деревьев, сосредоточенных непосредственно в лесной подстилке и верхнем слое почвы, немного. Многие авторы указывают на сложность в оценке подземной фитомассы в лесных экосистемах (Kurtz at al., 1996; Ruess et al., 1996; Cairns et al., 1997; Грозовская и др., 2015). Расчеты аллометрических моделей для биомассы корней не всегда представляются возможными в связи с нехваткой исходных данных (Усольцев и др., 2022). Изучение древесных корней также усложняется разным подходом исследователей к количеству выделяемых фракций по диаметру, и тонких корней в частности (Усольцев, 2018; Карпечко, Синькевич, 2023).
В лесах умеренной зоны корни составляют примерно 25 % от надземной фитомассы, по мере продвижения на север это соотношение меняется в связи с тем, что для северной растительности характерна более развитая, чем надземная фитомасса, корневая система. На европейском Севере определены запасы органического вещества в корнях ельников средней и северной тайги Архангельской (Руднева и др., 1966 и др.), Вологодской (Паршевников, 1962; Смирнов, 1962) областей, Республики Коми (Верхоланцева, Бобкова, 1972; Бобкова, 2001) и Карелии (Казимиров, Морозова, 1973). Подземная биомасса в лиственных лесах послерубочного происхождения в среднетаежной зоне на Европейском Севере изучена ранее (Смирнов, 1962; Паршевников, 1962; Казимиров и др., 1978; Вакуров, Полякова, 1982 и др.), в настоящее время она рассматривается реже (Пристова, 2010; Бобкова, Лиханова, 2012; Карпечко, Синькевич, 2023 и др.). В указанных исследованиях приведены данные либо в целом по подземной части фитомассы древесных растений и ТКЯ без разделения древесных корней по толщине, либо выделены тонкие (до 1 или 3 мм) и скелетные корни древесных растений.
Цель настоящей работы состояла в определении запасов органического вещества и углерода в древесных корнях диаметром до 20 мм и подземной фитомассе ТКЯ разновозрастных среднетаежных лиственных насаждений послерубочного происхождения.
Материалы
Исследования проводились на базе Ляльского лесоэкологического стационара Института биологии ФИЦ Коми НЦ УрО РАН, в Княжпогостском районе Республики Коми в подзоне средней тайги. Объекты расположены на территории ГУ РК «Железнодорожное лесничество» – березово-еловый молодняк разнотравно-зеленомошный (62°19' с.ш. 50°55' в.д.) (состав древостоя 5Б5ЕедСедОс, возраст 12 лет), расположен в Кылтовском участковом лесничестве ГУ (кв. 51), лиственно-хвойное (5Ос4Б1С+Е, 45 лет) (62º15' с.ш. 50º65' в.д.) и сосново-лиственное (5С2Б2Ос1Е, 45 лет) (62º15' с.ш. 50º64' в.д.) разнотравно-черничные насаждения – на территории Ляльского заказника. Исследуемые насаждения произрастают на месте еловых и сосновых вырубок. Древостой фитоценозов сложный по составу, представлен березой повислой (Betula pendula Roth.) и березой пушистой (B. pubescens Ehrh.), елью сибирской (Picea obovata Ledeb.), осиной обыкновенной (Populus tremula L.) и сосной обыкновенной (Pinus sylvestris L.), подрост – березой, елью и пихтой (Abies sibirica Ledeb.). Подлесок представлен ивой козьей (Salix caprea L.), ивой пятитычинковой (S. pentandra L.), ивой филиколистной (S. philicifolia L.), рябиной обыкновенной (Sorbus aucuparia L.), шиповником иглистым (Rosa acicularis Lindl.), жимолостью Палласа (Lonicera pallasii L.) и можжевельником обыкновенным (Juniperus communis L.). В ТКЯ исследуемых фитоценозов среди кустарничков наиболее распространены черника (Vaccinium myrtilus L.) и брусника (V. vitis-idaea L.), среди трав – щучка дернистая (Deschampsia cespitosa (L.) P. Beauv.), иван-чай (Chamerion angustifolium (L.) Scop.), золотая розга (Solidago virgaurea L.), в моховом покрове – зеленые и политриховые мхи. Почва 45-летних насаждений – иллювиально-железистый подзол, средняя толщина подстилки 5 см, 12-летнего березово-елового молодняка – торфянисто-подзолисто-глееватая, мощность подстилки 10 см. Более подробная характеристика исследуемых объектов представлена ранее (Пристова, 2010, 2019, 2022).
Методы
Подземная часть тонких древесных корней и растений ТКЯ определялась методом монолитов (Родин и др., 1968; Рожков и др., 2008). Отбор монолитов производился отдельно для подстилки (0–5 см, 0–10 см), отдельно для минеральных слоев почвы каждые 10 см на глубину до 50–55 см в 10-кратной повторности. В березово-еловом молодняке на глубине 40–50 см корни диаметром до 20 мм практически отсутствуют, поэтому данные по ним не приведены. Корни промывались через сито с размером ячеек 0.25 мм (Родин и др., 1968). Полученные образцы распределяли следующим образом: древесные корни по видам – на корни ели, березы, осины, ивы, подземные органы трав и кустарничков – без разделения на виды. Древесные корни разделяли на две основные фракции в зависимости от их диаметра: менее 1 мм и 1–20 мм. В образцах березово-елового молодняка дополнительно выделяли корни толщиной 1–2, 2–3, 3–5, 5–20 мм. Все полученные образцы высушивали при температуре 105 ˚С до абсолютно сухого веса (а.с.в.) и взвешивали (Орлов, 1967; Родин и др., 1968). Расчет содержания углерода в подземной фитомассе производился путем умножения веса корней каждого вида или группы видов на содержание в них углерода, данные выражались в кг/га. Для расчетов использованы ранее опубликованные данные по концентрации углерода в корнях растений лиственных фитоценозов средней тайги Республики Коми (Пристова, 2010, 2022). Средняя концентрация углерода в корнях ели, березы, осины и ивы составляет 47.5; 46.2; 46.4; 47.9 % соответственно, в подземных органах кустарничков – 48.4, травах – 43.1 % (Пристова, 2022). Статистическая обработка данных по подземной фитомассе и запасам углерода включала вычисление средней арифметической, среднего квадратического отклонения и коэффициента вариации (CV).
Результаты
Проведенное исследование затрагивает верхний и средний ярусы подземной части растений глубиной до 50–55 см. Как известно, подземные органы растений в лесных ценозах занимают 3 яруса: 1) верхний ярус (глубина 5–10 см) – обильно пронизан корнями, по массе преобладают физиологически активные корни деревьев и растений ТКЯ; 2) средний ярус (до 30–40 см), в котором преобладают корни кустарников и деревьев, при этом корней ТКЯ мало; 3) нижний ярус (до 1–2 м), в котором доминируют корни деревьев (скелетная часть) (Корчагин, 1976).
Запасы органического вещества в древесных корнях диаметром менее 20 мм и подземных органах травяно-кустарничкового яруса, сосредоточенных в 40–55-сантиметровом слое почвы, включая подстилку, в исследуемых фитоценозах составили от 3.9 до 9.8 т/га (табл. 1, 2, 3). При этом в древесных корнях толщиной до 20 мм накапливается от 1.4 до 8.5 т/га органического вещества.
Для березово-елового молодняка, в связи с небольшим возрастом, размерами деревьев, составом древостоя (Пристова, 2010, 2019) и менее развитой корневой системой, по сравнению со средневозрастными лиственными насаждениями, фракции мелких и тонких корней от 1 до 5 мм представлены более подробно (см. табл. 1). Основная масса корней ограничена глубиной 30–40 см, т. к. на глубине 40–50 см корни практически отсутствуют. В молодняке, в зависимости от технологических элементов прошедшей рубки, насыщенность почвы корнями различается, поэтому показатели фитомассы корней на месте пасечных участков и трелевочных волоков представлены отдельно (см. табл. 1). Общие запасы подземных органов растений в березово-еловом молодняке составляют 6.7 т/га на пасечном участке и 3.9 т/га на трелевочном, в т. ч. древесных корней 4.4 и 1.4 т/га соответственно (см. табл. 1). Масса корней березы и ели менее 3 мм в диаметре в 12-летнем березово-еловом молодняке составляет 1.6 т/га. Для 17-летних березово-еловых насаждений Вологодской области приводятся более высокие показатели для таких корней – 2.4–2.6 т/га (Паршевников, 1962). Корненасыщенность пасечных участков древесными корнями на 68 % выше, чем трелевочных, при этом для подземных органов ТКЯ она приблизительно одинакова. Вариабельность подземной фитомассы древесных растений с глубиной увеличивается. Так, коэффициент вариации (CV) показателя массы древесных корней на глубине до 30 см не превышает 16 %, на глубине более 30 см составляет около 30 %. На трелевочных участках масса корней, как правило, более вариабельна, чем на пасечных, особенно это выражено для корней ели. Запасы подземных органов растений травяно-кустарничкового яруса менее вариабельны, и с глубиной показатель CV увеличивается в среднем с 4 до 11 %.
Таблица 1. Распределение массы древесных корней (менее 20 мм) и подземных органов ТКЯ в березово-еловом молодняке, кг/га
| Жизненная форма, толщина древесных корней | Глубина, участок, см* | Всего | ||||||||
| 0–10 (n = 10) | 10–20 (n = 10) | 20–30 (n = 10) | 30–40 (n = 10) | |||||||
| пасечный | треле-вочный | пасеч- ный | треле-вочный | пасеч- ный | треле-вочный | пасеч- ный | треле-вочный | пасеч- ный | треле-вочный | |
| Древесные корни | ||||||||||
| Береза | 2050 | 300 | 780 | 110 | 40 | - | 20 | 10 | 2890 | 420 |
| >1 мм | 270 ± 18 | 40 ± 4 | 260 ± 11 | 40 ± 2 | 20 ± 3 | - | 10 ± 3 | 10 ± 3 | 560 | 90 |
| 1–2 мм | 300 ± 29 | 30 ± 3 | 250 ± 18 | 50 ± 7 | - | - | 10 ± 3 | - | 560 | 80 |
| 2–3 мм | 820 ± 73 | 90 ± 11 | 270 ± 21 | 20 ± 1 | 10 ± 1 | - | - | - | 1100 | 110 |
| 3–5 мм | 290 ± 34 | 140 ± 9 | - | - | 10 ± 1 | - | - | - | 300 | 140 |
| 5–20 мм | 370 ± 12 | - | - | - | - | - | - | - | 370 | - |
| Ива | 810 | 230 | 450 | 80 | 120 | 490 | - | 10 ± 3 | 1390 | 800 |
| >1 мм | 120 ± 9 | 20 ± 1 | 90 ± 7 | 30 ± 1 | 10 ± 1 | 80 ± 9 | - | - | 220 | 130 |
| 1–2 мм | 250 ± 23 | 60 ± 3 | 150 ± 17 | 40 ± 5 | 30 ± 3 | 140 ± 17 | - | 10 ± 3 | 440 | 240 |
| 2–3 мм | 330 ± 32 | - | 130 ± 7 | 10 ± 1 | - | 210 ± 19 | - | - | 460 | 220 |
| 3–5 мм | 110 ± 12 | - | 80 ± 11 | - | - | 60 ± 4 | - | - | 190 | 60 |
| 5–20 мм | - | 150 ± 12 | - | - | 80±6 | - | - | - | 80 | 150 |
| Ель | 190 | 30 | - | 80 | - | - | 10 ± 3 | - | 200 | 110 |
| >1 мм | 40 ±5 | 10 ± 2 | - | 40 ± 3 | - | - | - | - | 40 | 40 |
| 1–3 мм | 70 ± 11 | 10 ± 3 | - | 20 ± 1 | - | - | 10 ± 2 | - | 80 | 30 |
| 3–5 мм | - | - | - | - | - | - | - | - | - | - |
| 5–20 мм | 80 ± 12 | 10 ± 3 | - | 20 ± 2 | - | - | - | - | 80 | 30 |
| Подземные органы растений ТКЯ (всего) | 1180 | 570 | 350 | 1250 | 690 | 720 | - | - | 2220 | 2540 |
| травы | 1110 ± 46 | 540 ± 28 | 340 ± 15 | 1250 ± 97 | 690 ± 57 | 720 ± 65 | - | - | 2140 | 2510 |
| кустарнички | 70 ± 4 | 30 ± 4 | 10 ± 1 | - | - | - | - | - | 80 | 30 |
Примечание. - – данная фракция корней отсутствует, * – на глубине 40–50 см корни практически отсутствуют и в таблице не представлены. Береза представлена видами Betula pendula, B. pubescens, ива – Salix caprea, S. pentandra, S. philicifolia, ель – Picea obovata, кустарнички – Vaccinium myrtilus, V. vitis-idaea, травы – все травянистые виды растений, произрастающие в березово-еловом молодняке
Насыщенность почвы корнями (менее 20 мм) в средневозрастных смешанных лиственных насаждениях пространственно более однородна, чем в березово-еловом молодняке, и составляет 8.5 т/га в лиственно-хвойном (см. табл. 2) и 9.8 т/га в сосново-лиственном (см. табл. 3) насаждениях. Масса древесных корней в подстилке и минеральных слоях почвы сосново-лиственного насаждения на 1.2 т/га больше, чем в лиственно-хвойном. Это различие обусловлено тем, что доля участия сосны в составе древостоя сосново-лиственного насаждения выше, чем лиственно-хвойного. Вариабельность древесных корней с глубины 35 см увеличивается, в сосново-лиственном насаждении показатель CV для массы древесных корней возрастает с 6–12 до 43–50 %, в лиственно-хвойном – с 2–25 до 12–36 %. Для подземных органов растений ТКЯ наблюдается схожая тенденция – коэффициент вариации возрастает с 5-6 до 32–45 % и с 5–8 до 16–33 %.
Таблица 2. Распределение массы древесных корней (менее 20 мм) и подземных органов ТКЯ в лиственно-хвойном насаждении, кг/га
| Жизненная форма, толщина древесных корней | Глубина, см | Всего | |||||
|
0–5
(n = 10) |
5–15
(n = 10) |
15–25
(n = 10) |
25–35
(n = 10) |
35–45
(n = 10) |
45–55
(n = 10) |
||
| Древесные корни | |||||||
| Береза | 1355 | 1521 | 594 | 26 | 90 | 122 | 3708 |
| до 1 мм | 339 ± 41 | 155 ± 11 | 58 ± 7 | 26 ± 4 | 16 ± 2 | 11 ± 2 | 605 |
| 1–20 мм | 1016 ± 97 | 1366 ± 95 | 536 ± 31 | - | 74 ± 9 | 111 ± 19 | 3103 |
| Осина | 673 | 580 | 44 | 14 | 49 | 29 | 1389 |
| до 1 мм | 134 ± 11 | 52 ± 12 | 11 ± 1 | - | 13 ± 2 | - | 210 |
| 1–20 мм | 539 ± 46 | 528 ± 39 | 33 ± 4 | 14 ± 3 | 36 ± 6 | 29 ± 4 | 1179 |
| Сосна | 77 | 579 | 278 | - | - | - | 934 |
| до 1 мм | 41 ± 18 | 42 ± 4 | 56 ± 7 | - | - | - | 139 |
| 1–20 мм | 36 ± 5 | 537 ± 48 | 222 ± 17 | - | - | - | 795 |
| Ель | 882 | 333 | 24 | 3 | - | 3 ± 1 | 1245 |
| до 1 мм | 231 ± 18 | 48 ± 5 | 4 ± 1 | - | - | - | 283 |
| 1–20 мм | 651 ± 41 | 285 ± 25 | 20 ± 3 | 3 ± 1 | - | 3 ± 1 | 962 |
| Подземные органы ТКЯ (всего) | 580 | 440 | 80 | 28 | 28 | 73 | 1229 |
| травы | 250 ± 17 | 300 ± 19 | 20 ± 1 | 9 ± 1 | 9 ± 2 | 3 ± 1 | 591 |
| кустарнички | 330 ± 19 | 140 ± 8 | 60 ± 5 | 19 ± 2 | 19 ± 2 | 70 ± 9 | 638 |
Примечание. - – данная фракция корней отсутствует. Береза представлена Betula pendula, B. pubescens, ель – Picea obovata, осина – Populus tremula, сосна – Pinus sylvestris, кустарнички – Vaccinium myrtilus, V. vitis-idaea, травы – все травянистые виды растений, произрастающие в лиственно-хвойном фитоценозе
В средневозрастных насаждениях масса подземных органов ТКЯ приблизительно одинакова и составляет 1.2–1.3 т/га, что почти в 2 раза ниже, чем в березово-еловом молодняке. Такое отличие обусловлено разницей в видовом составе напочвенного покрова, происхождением и возрастом насаждений. В травяно-кустарничковом ярусе средневозрастных насаждений, по сравнению с березово-еловым молодняком, доля участия кустарничков выше, чем трав. Как показано ранее, березово-еловый молодняк находится на начальной стадии сукцессии и интенсивного роста, в связи с этим ТКЯ представлен травянистыми растениями, характерными для вырубок (Пристова, 2019, 2022).
В целом масса древесных корней (менее 20 мм) в средневозрастных насаждениях почти в 2 раза больше, чем в молодняке, и связано это в первую очередь с возрастом древостоя.
Таблица 3. Распределение массы древесных корней (менее 20 мм) и подземных органов ТКЯ в сосново-лиственном насаждении, кг/га
| Жизненная форма, вид, толщина корней | Глубина, см | Всего | |||||
| 0–5 (n = 10) | 5–15 (n = 10) | 15–25 (n = 10) | 25–35 (n = 10) | 35–45 (n = 10) | 45–55 (n = 10) | ||
| Береза | 1628 | 954 | 253 | 435 | 146 | 83 | 3499 |
| до 1 мм | 269 ± 31 | 150 ± 9 | 75 ± 4 | 56 ± 13 | 19 ± 3 | 7 ± 3 | 576 |
| 1–20 мм | 1359 ± 155 | 804 ± 65 | 178 ± 34 | 379 ± 39 | 127 ± 17 | 76 ± 17 | 2923 |
| Осина | 746 | 926 | 326 | - | 65 | 26 | 2089 |
| до 1 мм | 127 ± 14 | 105 ± 14 | 48 ± 3 | - | 11 ± 4 | 1 ± 0.2 | 292 |
| 1–20 мм | 619 ± 71 | 821 ± 132 | 278 ± 14 | - | 54 ± 7 | 25 ± 6 | 1797 |
| Сосна | 625 | 719 | 287 | - | - | - | 1631 |
| до 1 мм | 291 ± 33 | 244 ± 23 | 48 ± 3 | - | - | - | 583 |
| 1–20 мм | 334 ± 38 | 475 ± 122 | 239 ± 12 | - | - | - | 1048 |
| Ель | 629 | 294 | 20 | 101 | - | 4 | 1048 |
| до 1 мм | 121 ± 14 | 29 ± 4 | 3 ± 1 | 2 ± 1 | - | 1 ± 0.3 | 156 |
| 1–20 мм | 508 ± 58 | 265 ± 13 | 17 ± 1 | 99 ± 15 | - | 3 ± 1 | 892 |
| Ива | 46 | 101 | 53 | 26 | - | - | 226 |
| до 1 мм | 12 ± 1 | 35 ± 5 | 23 ± 3 | 3 ± 1 | - | - | 73 |
| 1–20 мм | 34 ± 2 | 66 ± 7 | 30 ± 2 | 23 ± 8 | - | - | 153 |
| Подземные органы ТКЯ | 657 | 309 | 192 | 20 | 27 | 62 | 1267 |
| травы | 243 ± 11 | 131 ± 7 | 33 ± 2 | 3 ± 0.1 | 11 ± 5 | 2 ± 1 | 423 |
| кустарнички | 414 ± 21 | 178 ± 9 | 159 ± 8 | 17 ± 3 | 16 ± 7 | 60 ± 21 | 844 |
Примечание. - – данная фракция корней отсутствует. Береза представлена Betula pendula, B. pubescens, ель – Picea obovata, осина – Populus tremula, сосна – Pinus sylvestris, ива – Salix caprea, S. pentandra, кустарнички – Vaccinium myrtilus, V. vitis-idaea, травы – все травянистые виды растений, произрастающие в лиственно-хвойном фитоценозе.
С глубиной в исследуемых насаждениях наблюдается уменьшение корненасыщенности почв (рис. 1, 2). В структуре подземной фитомассы древесных корней (менее 20 мм) исследуемых насаждений в верхнем 10–15 см слое почвы и органогенном горизонте сосредоточено наибольшее ее количество – от 68 до 82 %, при этом от 41 до 68 % от их массы накапливается в лесной подстилке. Особенностью березово-елового молодняка является более высокая корненасыщенность минерального слоя почвы на глубине 20–30 см на участке трелевочных волоков по сравнению с пасечными (см. рис. 1). На трелевочных участках в молодняке древесные корни на глубине 20–30 см представлены ивой, что обусловлено их зарастанием кустарниковыми видами ив (S. pentandra, S. philicifolia), реже березой и елью. Трелевочные участки отличаются переувлажнением и развитием сфагновых и зеленых мхов, что препятствует росту древесных пород и развитию их корневых систем (Пристова, 2019).
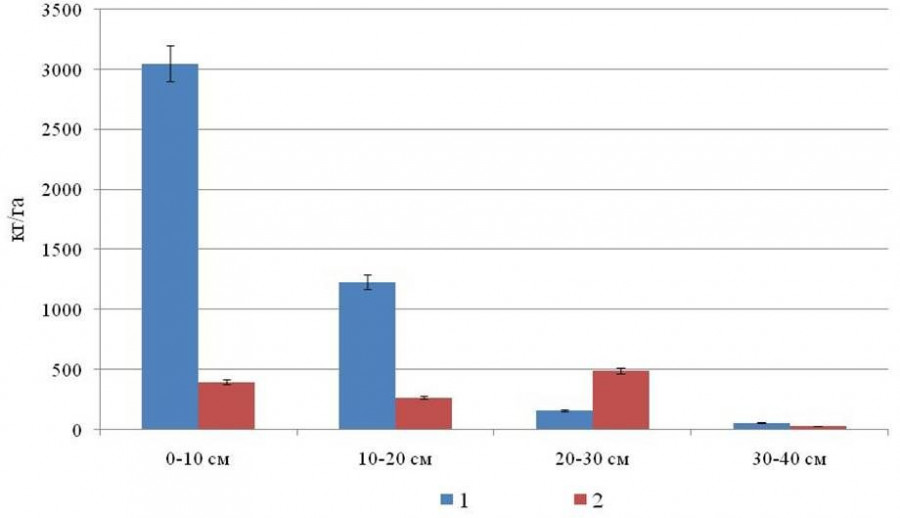
Рис. 1. Распределение массы древесных корней (менее 20 мм) на месте пасечных участков (1) и трелевочных волоков (2) в березово-еловом молодняке на разной глубине
Fig. 1. Distribution of the mass of tree roots (less than 20 mm) at the site of cutting sections (1) and skidding lines (2) in birch-spruce young growth at different depths
Структура распределения подземной фитомассы древесных корней сосново-лиственного и лиственно-хвойного насаждений схожа между собой (см. рис. 2). Отличием сосново-лиственного насаждения является более высокая корненасыщенность подстилки и минеральных горизонтов почвы на глубине 25–35 см.
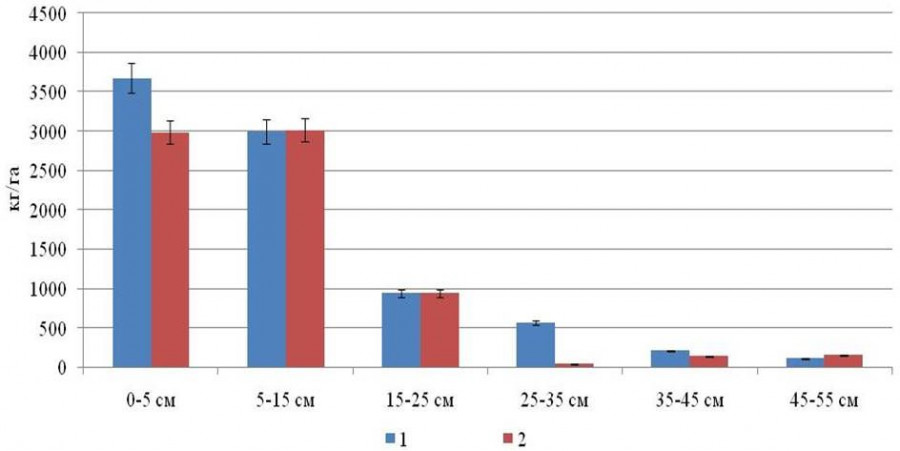
Рис. 2. Распределение массы древесных корней (менее 20 мм) в сосново-лиственном (1) и лиственно-хвойном (2) насаждениях на разной глубине, кг/га
Fig. 2. Distribution of the tree roots mass (less than 20 mm) in pine-deciduous (1) and deciduous-coniferous (2) stands at different depths, kg/ha
Масса физиологически активных корней толщиной до 1 мм, древесных пород (береза, ель, сосна, осина) в березово-еловом молодняке на пасечных участках составляет 0.6, на трелевочных – 0.1 т/га, в лиственно-хвойном – 1.2, в сосново-лиственном насаждении – 1.6 т/га. Более высокие результаты по этому показателю приводятся для 70-летних лиственно-еловых древостоев Карелии – 1.9 т/га (Карпечко, Синькевич, 2023).
Распределение фитомассы корней древесных растений в исследуемых насаждениях дифференцировано и представлено на рис. 3. Максимальная подземная фитомасса во всех трех фитоценозах характерна для корней березы (42–65 %), на втором месте в средневозрастных насаждениях – корни осины (19–25 %), в березово-еловом молодняке – ивы (31 %).
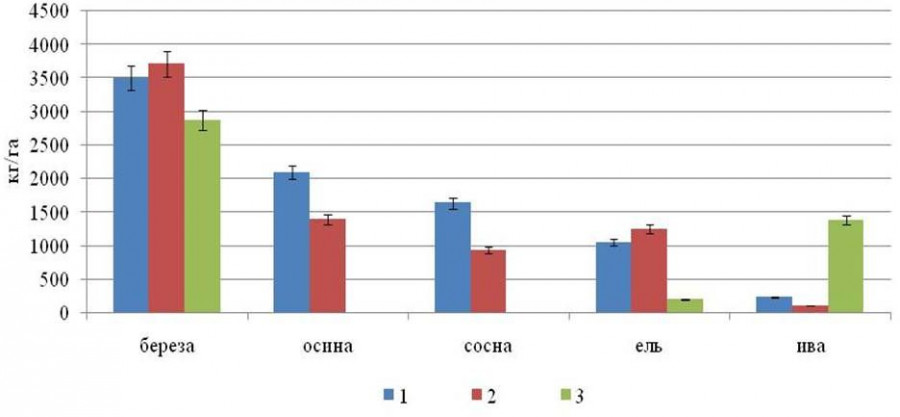
Рис. 3. Распределение массы древесных корней (менее 20 мм) в сосново-лиственном (1), лиственно-хвойном (2) насаждениях и березово-еловом молодняке (3), кг/га
Fig. 3. Distribution of tree roots mass (less than 20 mm) by species in pine-deciduous (1), deciduous-coniferous (2) stands and birch-spruce young forest (3), kg/ha
Запасы углерода в подземной фитомассе исследуемых фитоценозов составляют от 1.5 до 4.6 тС/га, при этом в древесных корнях накапливается от 0.4 до 4.0 тС/га (табл. 4). В средневозрастных насаждениях в подземной фитомассе депонируется в среднем почти в 2 раза больше углерода по сравнению с березово-еловым молодняком. Следует отметить, что на пасечных участках молодняка это различие составляет 1.3-1.5 раза, а на трелевочных – 2.7-3.1 раза.
Таблица 4. Запасы углерода в древесных корнях (менее 20 мм) и подземных органах трав и кустарничков лиственных фитоценозов
| Жизненная форма, вид | Лиственно-хвойное насаждение | Сосново-лиственное насаждение | Березово-еловый молодняк, участок | |||||
| пасечный | трелевочный | |||||||
| кгС/га | % | кгСга | % | кгС/га | % | кгС/га | % | |
| Древесные корни (всего) | 3410 | 86 | 3996 | 87 | 2077 | 68 | 393 | 26 |
| сосна | 461 | 12 | 804 | 18 | - | - | - | |
| береза | 1713 | 43 | 1617 | 35 | 1326 | 44 | - | - |
| ель | 591 | 15 | 498 | 11 | 90 | 3 | 10 | 1 |
| осина | 645 | 16 | 969 | 21 | - | - | ||
| ива | - | - | 108 | 2 | 661 | 22 | 383 | 26 |
| Подземные органы ТКЯ (всего) | 564 | 14 | 591 | 13 | 961 | 32 | 1097 | 74 |
| кустарнички | 309 | 8 | 409 | 9 | 39 | 1 | 15 | 1 |
| травы | 255 | 6 | 182 | 4 | 922 | 30 | 1082 | 73 |
| Итого | 3974 | 100 | 4587 | 100 | 3038 | 100 | 1490 | 100 |
Большая часть углерода (68–87 %) аккумулируется в корнях деревьев (менее 20 мм), исключение составляют трелевочные участки березово-елового молодняка, где накопление углерода в древесных корнях составляет 26 % (см. табл. 4). В лесной подстилке, в древесных корнях деревьев (менее 20 мм) накапливается 36–56 %, в подземных органах растений ТКЯ – 44–64 % от общего запаса углерода, сосредоточенного в определяемой подземной фитомассе.
Обсуждение
Одним из основных результатов проведенного исследования является получение данных по запасам органического вещества и углерода в подземной части фитомассы лиственных фитоценозов. Согласно полученным данным, древесные корни (менее 20 мм) исследуемых фитоценозов аккумулируют от 1.4 до 8.5 т/га органического вещества и от 0.4 до 3.4 т/га углерода. Эти показатели находятся в одном диапазоне с данными других исследователей для лиственных и хвойных лесов таежной зоны, в том случае если сравнивать с нашими результатами, полученными для древесных корней диаметром до 20 мм. Так, в таежных лесах Аляски фитомасса живых тонких древесных корней составляет 3–8 т/га (Ruess et al., 1996), в ельнике зеленомошном Архангельской области – 4 т/га (Руднева и др., 1966), в ельниках Республики Коми в 30-сантиметровом слое почвы от 4 до 16 т/га (Верхоланцева, Бобкова, 1972). Однако, если рассматривать только тонкие древесные корни толщиной до 1 мм, то различие с хвойными лесами будет выше, т. к. в исследуемых лиственных фитоценозах масса таких корней составляет 0.6–1.6 т/га, а в ельниках – 2–10 т/га (Верхоланцева, Бобкова, 1972). Масса корней березы и ели толщиной до 3 мм на пасечных участках в исследуемом березово-еловом насаждении составляет 2.3 т/га, что вполне сопоставимо с 17-летними березово-еловыми молодняками средней тайги Вологодской области, в которых этот показатель равняется 2.4–2.6 т/га (Паршевников, 1962), и лиственно-еловым насаждением Карелии (3.8 т/га) (Карпечко, Синькевич, 2023). Биомасса березовых и еловых корней толщиной до 2 мм в изучаемом молодняке составляет около 1.2 т/га, что в 2 раза меньше, чем в хвойных насаждениях Финляндии (Helmisaary et al., 2007). Таким образом, возникает определенная сложность при сравнении полученных результатов с данными других исследователей. Сравнение данных возможно только для корней определенной толщины. Одной из проблем при изучении древесных корней является то, что в настоящее время нет четкого подхода к разделению корней на фракции по диаметру, что отмечается многими исследователями (Усольцев, 2018; Карпечко, Синкевич, 2023 и др.). Так, корни деревьев, по мнению одних авторов, по толщине разделяют на следующие фракции: 1) 0–3 мм – тонкие сосущие корни; 2) 3–10 мм – ростовые корни; 3) 10–20 мм – ростовые корни; 4) менее 20 мм – корни, выполняющие опорную функцию (Базилевич, Титлянова, 1978). По мнению других исследователей, древесные корни разделяются так: 1) до 1 мм – сосущие корни; 2) 1–3 мм – полускелетные (проводящие); 3) более 3 мм – скелетные (проводящие) (Рахтеенко, 1952). Финские исследователи выделяют корни до 2 мм как тонкие, диаметром 2–5 мм как мелкие, а остальные – скелетные толстые корни (Helmisaary et al., 2007). Некоторые исследователи при изучении лиственных и лиственно-хвойных насаждений послерубочного происхождения относят к тонким корням фракции до 3 мм (Паршевников, 1962; Смирнов, 1962; Карпечко, Синькевич, 2023), дополнительно выделяя корни менее 1 мм (Карпечко, Синькевич, 2023). Таким образом, у исследователей подходы к количеству фракций при разделении древесных корней и выделении тонких корней разные. При проведении настоящих исследований применялись два варианта разделения древесных корней по толщине. В первом случае для всех трех насаждений были выделены корни толщиной до 1 мм и 1–20 мм. Во втором варианте для березово-елового молодняка из фракции диаметром 1–20 мм выделены корни толщиной 1–2, 2–3, 3–5, 5–20 мм. Такой подход к разделению корней в этом фитоценозе обусловлен рядом причин: особенностями вертикальной структуры подземной фитомассы, более ограниченной глубиной их проникновения по сравнению со средневозрастными насаждениями, небольшой массой корней толщиной 5–20 мм. Кроме того, выделение большего количества фракций корней толщиной до 5 мм позволило сопоставить полученные данные с результатами других исследователей.
Таким образом, количество выделенных фракций при распределении корней по толщине может быть связано как с особенностями изучаемого объекта, так и с поставленными задачами. При определении количества депонируемого углерода в подземной фитомассе древесных растений важное значение придается крупным скелетным корням, а при оценке пула МОВ в углеродном цикле – тонким древесным корням и подземным органам растений ТКЯ (Kurtz at al., 1996). Для исследований в области физиологии древесных растений изучение тонких физиологически активных корней диаметром до 1 мм более актуально, чем других фракций (Рахтеенко, 1952; Helmisaari et al, 2007; Карпечко, Синькевич, 2023).
В подземных органах растений ТКЯ исследуемых насаждений сосредоточено от 1.2 до 2.5 т/га органического вещества и от 0.6 до 1.1 т/га углерода. Подземные органы растений ТКЯ среднетаежных хвойных насаждений Республики Коми аккумулируют от 1.3 (Бобкова, 2001) до 2.4 т/га органического вещества (Запрудина, 2010), что довольно близко к нашим данным.
Полученные данные позволяют оценить не только запасы органического вещества и углерода в подземных органах растений, но и вертикальную структуру их распределения. В целом для всех исследуемых насаждений характерно преимущественное распределение корней деревьев (менее 20 мм) и подземных органов растений ТКЯ в верхнем 20–25-сантиметровом слое почвы – 87–95 % от общей массы корней. Похожие данные приводятся для хвойных лесов (Верхоланцева, Бобкова, 1972; Helmisaari, 2007). Средневозрастные лиственно-хвойные насаждения отличаются более глубоким распределением корней по сравнению с березово-еловым молодняком. Это в первую очередь обусловлено возрастом деревьев. Как известно, с возрастом у деревьев рост корней увеличивается (Рахтеенко, 1952). Также важным фактором является большая доля участия сосны в формировании древостоев средневозрастных насаждений. Как известно, в сосняках черничного типа корневая система сосны интенсивно использует весь освоенный корнями почвенный профиль (Тюкавина, Евдокимов, 2016). Показано, что вариабельность показателей подземной фитомассы увеличивается с глубиной, что обусловлено значительной разреженностью и снижением массы корней с 30–35 см глубины.
Результаты проведенных исследований также отражают распределение подземной фитомассы отдельных видов древесных растений. Согласно представленным данным, корни ели накапливают меньше органического вещества и углерода (5–17 %), чем корни лиственных пород (65–69 %). Участие ели в составе древостоя не превышает 10 %. Однако, как показано ранее, количество ели в подросте больше, чем в древостое: в березово-еловом молодняке – 12, в средневозрастных насаждениях – 1–2 тыс. экз./га (Пристова, 2010, 2019). По данным исследователей, изучавших корневые системы смешанных березово-еловых насаждений, корни ели могут угнетаться корнями березы, и лишь по мере естественного изреживания березы в древостое происходит разделение экологических ниш и снижение конкуренции между этими породами (Чибисов, 1971; Рахтеенко, 1981). Также многими авторами отмечается, что корни лиственных пород более активно осваивают почвенные горизонты и используют их ресурсы, а корни ели сосредоточены преимущественно в верхних слоях почвы (Рахтеенко, 1981; Mekontchou, 2020; Shanin, 2015). В то же время сосна в смешанных с березой насаждениях растет лучше и более устойчива, чем в чистых культурах (Рахтеенко, 1963).
Заключение
Проведена оценка запасов органического вещества и углерода в древесных корнях толщиной до 20 мм и подземных органах растений ТКЯ на глубине до 40–55 см в разновозрастных среднетаежных лиственно-хвойных фитоценозах послерубочного происхождения на территории Республики Коми. Установлено, что в подземной фитомассе аккумулируется от 3.9 до 9.8 т/га органического вещества и от 1.5 до 4.6 тС/га. Исследование структуры подземной фитомассы древесных корней (менее 20 мм) показало, что в верхнем 10–15-сантиметровом минеральном слое почвы и органогенном горизонте сосредоточено наибольшее количество корней – от 68 до 82 %. Согласно полученным данным, возраст и состав древостоя оказывают влияние на запасы органического вещества и углерода. В средневозрастных лиственно-хвойных насаждениях эти показатели в среднем в 2 раза выше, чем в молодняке. Показано, что более высокая доля участия сосны в древостое сосново-лиственного насаждения по сравнению с лиственно-хвойным увеличивает запасы подземной фитомассы на 1.2 т/га. Проведенные исследования по депонированию углерода в подземной фитомассе позволяют оценить ее участие в пуле МОВ и углеродном цикле в целом в лиственно-хвойных насаждениях послерубочного происхождения.
Библиография
Базилевич Н. И., Титлянова А. А., Смирнов В. В., Родин Л. Е., Нечаева Н. Т., Левин Ф. И. Методы изучения биологического круговорота в различных природных зонах . М.: Наука, 1978. 184 с.
Бобкова К. С. Еловые леса // Биопродукционный процесс в лесных экосистемах Севера / Отв. ред. К. С. Бобкова, Э. П. Галенко. СПб.: Наука, 2001. С. 52–67.
Бобкова К. С., Лиханова Н. В. Вынос углерода и элементов минерального питания при сплошнолесосечных рубках ельников средней тайги // Лесоведение. 2012. № 6. С. 44–54.
Вакуров А. Д., Полякова А. Ф. Круговорот азота и минеральных элементов в 35-летнем осиннике // Круговорот химических веществ в лесу. М.: Наука, 1982. С. 44–54.
Верхоланцева Л. А., Бобкова К. С. Влияние почвенных условий на корневые системы древесных пород в еловых насаждениях подзоны северной тайги . Сыктывкар: Коми филиал АН СССР, 1972. 56 с. (Серия препринтов «Научные доклады» / АН СССР. Коми филиал; Вып. 6).
Грозовская И. С., Ханина Л. Г., Смирнов В. Э., Бобровский М. В., Романов М. С., Глухова Е. М. Биомасса напочвенного покрова в еловых лесах Костромской области // Лесоведение. 2015. № 1. С. 63–76.
Запрудина М. В. Фитомасса травяно-кустарничкого и мохового ярусов темнохвойных высокотравных лесов Печоро-Илычского заповедника // Известия Самарского научного центра Российской академии наук. 2010. Т. 12, № 1 (3). С. 876–879.
Казимиров Н. И., Морозова Р. М. Биологический круговорот веществ в ельниках Карелии . Л.: Наука, 1973. 175 с.
Казимиров Н. И., Морозова Р. М., Куликова В. Н. Органическая масса и потоки веществ в березняках средней тайги . Л.: Наука, 1978. 216 с.
Карпечко А. Ю., Синькевич С. М. Влияние реконструктивной рубки в лиственно-еловом древостое на корневую конкуренцию пород // Лесной журнал. 2023. № 3. С. 99–112.
Корчагин А. А. Строение растительных сообществ // Полевая геоботаника. Л.: Наука, 1976. С. 5–320.
Орлов А. Я. Метод определения массы корней деревьев в лесу и возможность учета годичного прироста органической массы в толще лесной почвы // Лесоведение. 1967. № 1. С. 64–70.
Паршевников А. Л. Круговорот азота и зольных элементов в связи со сменой пород в лесах средней тайги // Типы леса и почвы северной части Вологодской области: Труды Института леса и древесины / Под ред. Г. П. Мотовилова. М.; Л.: Изд-во АН СССР, 1962. С. 196–209.
Пристова Т. А. Компоненты углеродного цикла в лиственно-хвойном насаждении средней тайги // Лесоведение. 2010. № 6. С. 12–19.
Пристова Т. А. Динамика древесной растительности в лиственных насаждениях послерубочного происхождения (подзона средней тайги Республики Коми) // Принципы экологии. 2019. Т. 8, № 3. С. 63–73.
Пристова Т. А. Содержание углерода в растениях среднетаежных лиственных фитоценозов Республики Коми // Принципы экологии. 2022. № 3. С. 43–49.
Рахтеенко И. Н. Корневое питание ели и березы при взаимодействии их корневых систем // Известия АН БССР. Сер.: биол. науки. 1981. № 3 (5–8). С. 123.
Рахтеенко И. Н. Корневые системы древесных и кустарниковых пород . М.; Л: Гослесбумиздат, 1952. 106 с.
Родин Л. Е., Ремезов Н. П., Базилевич Н. И. Методические указания к изучению динамики и биологического круговорота в фитоценозах . Л.: Наука, 1968. 145 с.
Рожков В. А., Кузнецова И. В., Рахматуллоев Х. Р. Методы изучения корневых систем в поле и лаборатории . М.: Изд-во МГУЛ, 2008. 51 с.
Руднева Е. Н., Тонконогов В. Д., Дорохова К. Я. Круговорот зольных элементов и азота в ельнике-зеленомошнике северной тайги бассейна р. Мезень // Почвоведение. 1966. № 1. С. 14–26.
Смирнов В. В. Изменение сырорастущей органической массы в еловых и елово-лиственных древостоях средней тайги в связи с их возрастом // Материалы к научному обоснованию некоторых лесохозяйственных мероприятий в северной части Вологодской области: Труды Института леса и древесины. М.; Л.: Изд-во АН СССР, 1962. Т. LIII. С. 103–125.
Тюкавина О. Н., Евдокимов В. Н. Корневая система сосны обыкновенной в условиях северотаежной зоны // ИВУЗ. «Лесной журнал». 2016. № 1. С. 55–65.
Усольцев В. А. В подвалах биосферы: что мы знаем о первичной продукции корней деревьев? // Эко-потенциал. 2018. № 4 (24). С. 24–77.
Усольцев В. А., Цепордей И. С., Норицын Д. В. Аллометрические модели биомассы деревьев лесообразующих пород Урала // Леса России и хозяйство в них. 2022. № 1 (80). С. 4–14.
Чибисов Г. А. Рост корневых систем в березово-еловых насаждениях и влияние на них рубок ухода // Сборник работ по лесному хозяйству и лесохимии. Архангельск: Сев.-зап. кн. изд-во, 1971. С. 121–130.
Cairns M. A., Brown S., Helmer E. H., Baumgardner G. A. Root biomass allocation in the world's upland forests // Oecologia. 1997. № 11. P. 1–11.
Helmisaari H.-S., Derome J., Nöjd P., Kukkola M. Fine Root Biomass in Relation to Site and Stand Characteristics in Norway Spruce and Scots Pine Stands // Tree Physiology. 2007. Vol. 27, No. 10. P. 1493–1504. DOI: 10.1093/treephys/27.10.1493.
Kurtz W. A., Beukema S. J., Apps M. J. Estimation of root biomass and dynamics for the carbon budget model of the Canadian forest sector // Can J. For. Res. 1996. Vol. 26. P. 1973–1979.
Mekontchou C. G., Houle D., Bergeron Y., Drobyshev I. Contrasting Root System Structure and Belowground Interactions Between Black Spruce (Picea mariana (Mill.) B.S.P) and Trembling Aspen (Populus tremuloides Michx) in Boreal Mixedwoods of Eastern Canada // Forests. 2020. Vol. 11, No. 2. P. 127. DOI: 10.3390/f11020127.
Ruess R. W., Van Cleve K., Yarie J., Viereck Z. V. Contributions of fine root production and turnover to the carbon and nitrogen cycling in taiga forests of the Alaskan interior // Can. J. For. Res. 1996. Vol. 28. P. 1326–1336.
Shanin V. N., Rocheva L. K., Shashkov M. P., Ivanova N. V., Moskalenko S. V., Burnasheva E. R. Spatial Distribution Features of the Root Biomass of Some Tree Species (Picea abies, Pinus sylvestris, Betula ) // Biology Bulletin. 2015. Vol. 42, No. 3. P. 260–268. DOI: 10.1134/S1062359015030115.
Благодарности
Работа выполнена при финансовой поддержке темы госзадания Института биологии Коми научного центра УрО РАН «Зональные закономерности динамики структуры и продуктивности первичных и антропогенно измененных фитоценозов лесных и болотных экосистем европейского северо-востока России» (№ 122040100031–8).

 © 2011 - 2026
© 2011 - 2026