



Введение
Фундаментальная задача ботанических садов на протяжении всей их истории состояла в распространении растений, полезных для человека, мобилизации генетических ресурсов растений и разработке основ их культивирования в новых условиях (Андреев и др., 2005; Камелин, 2006). Создавая ботанические коллекции и акклиматизируя растения на протяжении последних 6 веков как целенаправленно, так и спонтанно, ботанические сады стали новыми центрами распространения растений. Такие центры, обычно содержащие тысячи таксонов растений в открытом грунте, расположены в различных экологических условиях. Ботанические сады с их коллекциями живых растений являются неотъемлемой частью эффективного использования генетических ресурсов растений, изучения и сохранения биологического разнообразия, развития наукоемкой экономики, медицины, экобиотехнологии, сельского хозяйства и других сфер деятельности, определяющих поступательное социально-экономическое развитие, повышение уровня жизни людей, обеспечение продовольственной и биологической безопасности (Инвентаризация коллекций..., 2002). Осуществление ботаническими садами вышеуказанной научно-практической деятельности может иметь существенные эволюционные последствия, связанные: с распространением видов растений за пределы их естественного экологического оптимума; с адаптацией растений к нетипичным климатическим условиям; с созданием условий для межвидовой гибридизации близкородственных перекрестноопыляющихся видов и сортов растений; с селекцией и спонтанным размножением потенциально инвазионных гибридов. Требует критического анализа и участие ботанических садов в работах по сохранению биологического разнообразия, в связи с особенностью формирования коллекций растений.
Методы
Выполнение исследований базируется на ранее разработанной организационной и методологической базе. Инвентаризация генетических ресурсов сосудистых растений в коллекциях ботанических садов России и создание автоматизированных банков паспортных и оценочных данных осуществляются на основе локальных систем регистрации коллекций, поддерживающих международный стандарт обмена информацией о коллекциях растений (Wyse Jackson, 1997). Одной из таких систем является разработанная нами и распространенная в ботанических садах система «Калипсо» (Нестеренко и др., 1997; Андрюсенко, Прохоров, 2012).
Собранные данные обрабатываются с помощью информационно-аналитической системы «Ботанические коллекции России» (Прохоров, Нестеренко, 2000; Прохоров, Андрюсенко, 2012). По нашим данным, в ботанических садах России культивируется более 23000 видов и 24000 садовых форм сосудистых растений (Прохоров, 2002). Этот информационный инструмент может успешно применяться для оценки результатов мобилизации и сохранения генетических ресурсов сосудистых растений в ботанических садах. Информационно-аналитическая система «Ботанические коллекции России» (далее – ИАС) позволяет принимать участие в оценке результатов работы по сохранению биологического разнообразия в рамках задач GSPC (Глобальная стратегия сохранения растений, 2003). С позиций национальных интересов России ИАС необходима для оценки перспектив интродукции и создания необходимых коллекций генетических ресурсов растений для обеспечения ресурсной биологической безопасности и для развития биотехнологий в РФ (Инвентаризация коллекций..., 2002). С позиций ботанических садов ИАС позволяет планировать коллекционную политику для достижения максимальной уникальности коллекций и их оптимальной насыщенности в конкретных климатических условиях (Прохоров и др., 2012). В плане же ботанических исследований и применительно к рассматриваемому вопросу ИАС позволяет оценить степень нарушения естественной географии растений и эволюционный потенциал ботанических садов (Prokhorov, 2011).
Результаты
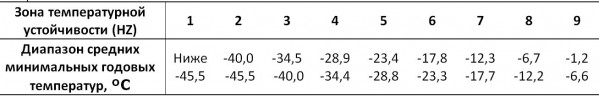
В ходе анализа Gymnospermae (Еглачева и др., 2011; Прохоров и др., 2012), был рассмотрен культурный ареал некоторых североамериканских видов, представленных в ботанических садах России. Abies concolor (Gordon) Lindl. ex Hildebr. и Abies fraserii (Pursh) Poir. (рис. 1, 2) наиболее ярко демонстрируют распространенное явление при интродукции растений – адаптацию к измененным климатическим условиям. В данном случае – к минимальным зимним температурам, наиболее значимом из климатических факторов для древесных растений (Rehder, 1949). На территории России отмечено 9 зон температурной устойчивости (табл. 1), границы которых совпадают с изотермами средних минимальных зимних температур за долгосрочный период с шагом 5° по шкале Фаренгейта.
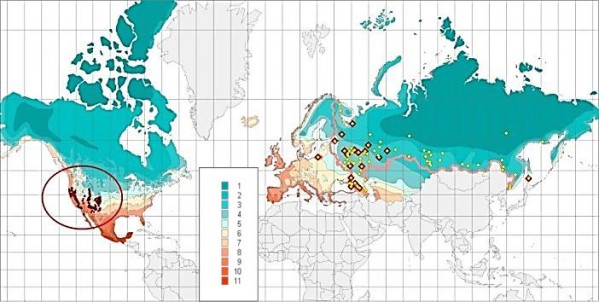
Рис. 1. Естественный ареал и встречаемость в ботанических садах России Abies concolor (Gordon) Lindl. ex Hildebr. в диапазоне зон температурной устойчивости растений (табл. 1)
Fig. 1. The natural habitat and occurrence of Abies concolor (Gordon) Lindl. ex Hildebr. in the botanical gardens of Russia in the range of hardiness zones of plants (table 1)
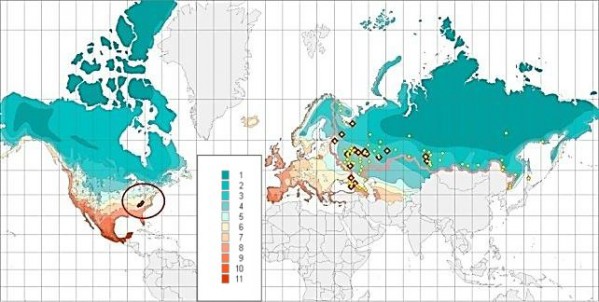
Рис. 2. Естественный ареал и встречаемость в ботанических садах России Abies fraserii (Pursh) Poir. в диапазоне зон температурной устойчивости растений (табл. 1)
Fig. 2. The natural habitat and occurrence of Abies fraserii (Pursh) Poir in the botanical gardens of Russia in the range of hardiness zones of plants (table 1)
Таблица 1. Характеристики зон температурной устойчивости растений
Table 1. The characteristics of the zones of plants hardiness
Сопоставление естественных ареалов видов растений с их распространением в коллекциях ботанических садов свидетельствует о выходе большинства рассмотренных видов за границы своих климатических предпочтений (Еглачева и др., 2011; Прохоров и др., 2012). Это происходит в результате соответствующих адаптаций (в онтогенезе или филогенезе), обеспечивающих отдельным особям или их потомкам приспособление к нетипичным условиям существования. Можно предположить, что на группу особей одного таксона в ботанической коллекции действуют те же факторы, не зависящие от плотности популяции, что и на периферийную малочисленную популяцию (Ивантер, 2012).
Процесс интродукции растений завершается образованием культигенных популяций, адаптированных к новым эколого-климатическим условиям (Карпун, 2003). Таким образом, формируется сеть своеобразных резерватов генетических ресурсов растений за пределами их естественных ареалов и, соответственно, экологических оптимумов.
Еще одной особенностью ботанических садов является наличие многих представителей одного рода в коллекции, родовых комплексов (Русанов, 1977). В результате на относительно небольшой территории ботанического сада в новых экологических условиях изменяются взаимоотношения между близкородственными видами, и создаются уникальные условия для спонтанной межвидовой гибридизации с неконтролируемыми и непредсказуемыми результатами.
Рассмотрим крупнейшие родовые комплексы Salix L. в ботанических садах России (табл. 2), для которых приведены значимые климатические параметры: HZ – зоны температурной устойчивости растений, в интервалах температур в ºC; FF – продолжительность безморозного периода, в днях; HT – число дней с температурой выше 30 ºC.
Таблица 2. Крупнейшие родовые комплексы Salix L. в БС России
Table 2. The largest generic complexes Salix L in botanical gardens in Russia
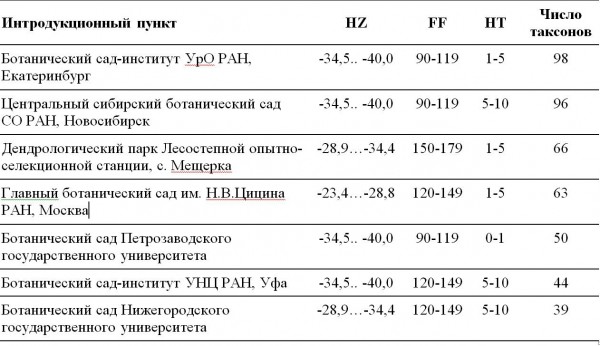
Представленные ботанические сады располагаются в различных условиях, характеризуемых комбинацией климатических параметров среды. В каждом из них собрана своя коллекция видов и сортов, которые в результате адаптации к местным условиям изменили сроки цветения и других фенологических фаз, что приводит к появлению новых вариантов родительских пар. Эти интродукционные пункты становятся источником новых гибридов ивы, появление которых в естественных условиях невозможно, поскольку в природе мы имеем как сформировавшиеся ареалы видов, так и сформировавшиеся зоны гибридизации на их границах. Только в случаях выявления хозяйственно ценных признаков селекция ведется целенаправленно и факты получения новых сортов фиксируются (Беляева и др., 2000).
Обсуждение
Адаптированные растения-интродуценты, новые местные сорта и спонтанные гибриды, появившиеся в условиях коллекции ботанического сада – это потенциальная угроза для соседствующих растительных сообществ (Виноградова и др., 2010), т. к. зачастую расселяются за пределы коллекций, входя в состав местных фитоценозов, а также переносятся в виде семян и вегетативного материала в коллекции других ботанических садов. Однако не следует забывать, что современное биологическое разнообразие в каждом регионе во многом является результатом естественных инвазий, осуществляющихся на протяжении миллионов лет эволюции.
Общеизвестно, что появление инвазионных видов в естественных растительных сообществах обусловлено как наличием потенциальных инвазионных видов, так и состоянием самих фитоценозов. Ботанические сады в основном располагаются в крупных населенных пунктах и контактируют с другими урбаноценозами и агроценозами, заселение которых адаптированными интродуцентами и местными гибридами значительно облегчено по сравнению с природными сообществами.
Нечто подобное происходило и ранее, с тех пор как человечество стало заниматься земледелием, и некоторые современные виды, принимаемые за вполне естественные, представляют собой давние стабилизировавшиеся гибриды (многие цитрусовые, пальмы, фикусы, отдельные виды роз и другие полезные растения). Ботанические сады, с их огромными коллекциями самых разнообразных растений, существенно ускорили это неоднозначное явление в эволюции растений.
Следует отметить, что ботанические сады, принявшие на себя функцию сохранения биологического разнообразия ex situ с 90-х годов прошлого века, невольно его еще и приумножают, порождая гибридные виды и даже роды растений. Одновременно невольно возникает вопрос: а возможно ли вообще сохранить для потомков сложившееся биологическое разнообразие, в данном случае растений? В частности, возможно ли это сделать, создавая коллекции живых растений?
Сохранение биологического разнообразия ex situ, тем более в коллекциях ботанического сада, предполагает выращивание ограниченного числа особей растений. Эти особи должны обходиться без привычных для вида исторически сложившихся ценотических связей и достаточно легко образовывать новые связи, не свойственные данному виду. Как следствие этого, мы имеем в культуре не сам вид и не типичных представителей данного вида, а особи, характеризующиеся большей устойчивостью в условиях культуры. Такие особи часто и внешне отличаются от типичных представителей вида, представляя собой своеобразные культивары, т. к. генетически однородный коллекционный образец формируется чаще всего путем вегетативного размножения.
Мы сохраняем под существующим видовым названием некий новый таксон, если работа по сохранению растений не идет на уровне популяций и за пределами естественного природного ареала и экотопа. Соответственно, реинтродуцируя такой культигенный таксон (понимая под термином «реинтродукция» возврат растения в естественную среду обитания в пределах исходного ареала вида), мы, по сути дела, интродуцируем нечто новое в природные условия. Такая «реинтродукция» контрпродуктивна в плане восстановления естественных фитоценозов, но одновременно способствует активизации эволюционных процессов у таких видов, благодаря несколько измененному геному «реинтродуцированных» особей.
Заключение
На карте (рис. 3), созданной международной организацией Botanic Gardens Conservation International (BGCI, 2012), показано более 3100 ботанических садов Земли, в коллекциях которых представлено около 1/3 существующих видов сосудистых растений. Эти растения в основном культивируются за пределами их экологического оптимума и ареала. В ходе интродукции растений осуществляется их адаптация к условиям новой среды обитания и селекция форм, устойчивых к новым экологическим условиям. В каждой коллекции представлены большие группы близкородственных видов, что создает оптимальные условия для межвидовой гибридизации. Таким образом, ботанические сады увеличивают биологическое разнообразие и расширяют ареалы видов. Эволюционные процессы, в естественных условиях занимающие миллионы лет, происходят в ботанических садах в течение короткого промежутка времени.
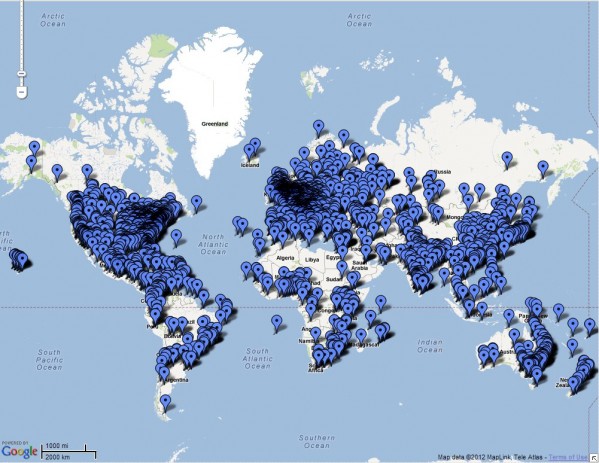
Рис. 3. Ботанические сады Земли (BGCI, 2012)
Fig. 3. The botanical gardens of the Earth (BGCI, 2012)
Многочисленные ботанические сады, являющиеся антропогенными центрами эволюции и распространения растений, оказывают влияние на формирование растительного мира Земли.
Библиография
Андреев Л. Н., Бер М. Н., Егоров А. А., Камелин Р. В., Лурье Е. А., Прохоров А. А., Стриханов М. Н., Селиховкин А. В. Ботанические сады и дендрологические парки высших учебных заведений. // Hortus botanicus. 2005. Т. 3. C. 5–27.
Андрюсенко В. В., Прохоров А. А. «Калипсо» – система управления базами данных ботанических коллекций : 2012615043: Свидетельство о государственной регистрации программы для ЭВМ. Российская Федерация, 6 июня 2012 г. Правообладатель: ФГБОУ ВПО «ПетрГУ».
Беляева И. В., Шабуров В. И., Дьяченко А. А. Гибридные ивы в декоративном садоводстве на Среднем Урале // Бюллетень Главного ботанического сада РАН. 2000. Т. 180. C. 102—109.
Виноградова Ю. К., Майоров С. Р., Хорун Л. В. Черная книга флоры Средней России. Чужеродные виды растений в экосистемах Средней России. М.: Геос, 2010. 512 с.
Глобальная стратегия сохранения растений. М.: Секретариат Конвенции о биологическом разнообразии, ОМСБСОР, 2003. 16 с.
Еглачева А. В., Андрюсенко В. В., Дементьев П. А., Савченко О. Н., Прохоров А. А. Интродукция аборигенных видов хвойных Северной Америки в ботанических садах России // Ботанические сады в современном мире: теоретические и прикладные исследования: Материалы Всероссийской научной конференции с международным участием, посвященной 80-летию со дня рождения академика Л. Н. Андреева (5–7 июля 2011 г., г. Москва). М.: Товарищество научных изданий КМК, 2011. C. 179–185.
Ивантер Э. В. Периферические популяции политипического вида и их роль в эволюционном процессе // Принципы экологии. 2012. № 2. C. 71–75. URL: http://ecopri.ru/journal/atricle.php?id=861.
Инвентаризация коллекций национальных ресурсов растений, животных, микроорганизмов и клеточных культур : Отчет по проекту № ВК-458/43-1349. М.: ГНИИ «Биоэффект», 2002.
Камелин Р. В. Ботанические сады в системе образования = Уникальные объекты высшей школы. Ботанические сады и дендрологические парки высших учебных заведений / Ред. А. А. Прохоров. Петрозаводск: Информационно-аналитический центр СБСР; «Копи-Диск», 2006. CD-ROM.
Карпун Ю. Н. Основы интродукции растений // Hortus botanicus. 2003. Т. 2. C. 17–32. URL: http://hb.karelia.ru/.
Нестеренко М. И., Прохоров А. А., Груздева Е. А., Холодкова Е. Ю. «Калипсо» – база данных коллекционных фондов для ботанических садов // Компьютерные базы данных в ботанических исследованиях. СПб.: ЗИН РАН, 1997. C. 70–71.
Прохоров А. А. Обеспечение открытого доступа к информации о коллекционных фондах ботанических садов // Ботанический журнал. 2002. Т. 87. № 11. C. 127–130.
Прохоров А. А., Андрюсенко В. В. Информационно-аналитическая система «Ботанические коллекции России» : 2012613396: Свидетельство о государственной регистрации программы для ЭВМ. Российская Федерация, 10 апрель 2012 г. Правообладатель: ФГБОУ ВПО «ПетрГУ».
Прохоров А. А., Еглачева А. В. и др. Влияние климатических изменений на состояние и формирование коллекционных фондов ботанических садов России – резерва генетических ресурсов растений : Отчет о НИР / Ботанический сад ПетрГУ; Петрозаводский государственный университет. Петрозаводск, 2012. 106 с. № ГР 01200901606. Инв. № 02201256108.
Прохоров А. А., Нестеренко М. И. Информационно-поисковая система «Ботанические коллекции России в Интернете» // Бюллетень Главного ботанического сада. 2000. Т. 180. C. 124–128.
Русанов Ф. Н. Метод родовых комплексов в интродукции растений // Бюллетень ГБС АН СССР. 1977. Т. 81. C. 15–20.
BGCI. Global Distribution of Botanic Gardens // Botanic Gardens Conservation International. : InGraphix, 2012. URL: http://www.bgci.org/map.php.
Prokhorov A. Botanic gardens and their role in plant evolution // XVIII International Botanical Congress. Melbourne, 2011. Pp. 225—226.
Rehder A. Manual of cultivated trees and shrubs hardy of in North America. New York: The Macmillan Company, 1949. 996 p.
Wyse Jackson D. International Transfer Format for Botanic Garden Plant Records (version 2.00 draft 3.2.). Richmond: Botanic Gardens Conservation International, 1997. 60 p. URL: http://www.tdwg.org/standards/102/download/102-525-1-RV.pdf
Благодарности
Исследования выполнены при поддержке Министерства образования и науки Российской Федерации (проект 4.1700.2011) и Российского фонда фундаментальных исследований (проект 12-04-90018).

 © 2011 - 2026
© 2011 - 2026