



Введение
Прикладные работы о вкладе видов, особенно редких, в поддержание и сохранение биоразнообразия достаточно распространены начиная с конца прошлого века, когда в связи с развитием природоохранных методов появился общественный запрос на соответствующие исследования. Попытки математически обосновать роль редких или обычных видов в поддержании биоразнообразия относятся к сравнительно недавнему времени. Веховой работой здесь является статья Джека Леннона с соавторами (Lennon et al., 2004), в которой было проанализировано влияние на распределение местного населения птиц трех территорий (Великобритании, отдельно Шотландии и Южной Африки) видов с разным обилием. В работе было показано, как меняется корреляция парциальной фауны отдельных пространственных ячеек с общим показателем видового разнообразия при добавлении вида за видом в градиенте от самого распространенного до самых редких с общим видовым списком. По итогам был сделан вывод о том, что большая часть пространственной вариации разнообразия обусловлена наличием сравнительно малого числа довольно частых видов.
На данный момент выполнено много работ на разных объектах, методологически одинаково или почти одинаково с цитируемой работой (Vázquez, Gaston, 2004; Mazaris et al., 2008; Sizling et al., 2009; Heegard et al., 2013), в т. ч. и на сообществах луговых растений (Lennon et al., 2011). Везде констатируется более высокая роль обычных видов в формировании видового богатства. Впрочем, заметно, что влияние это крайне неравномерное и отходит собственно от градиента «обычный – редкий». Даже соседние по рангу обычности виды могут оказывать совершенно разное влияние на корреляцию с итоговой величиной видового разнообразия.
В нашей работе мы попытались вычленить виды, присутствие или обилие которых оказывает наибольшее влияние на итоговое значение альфа- и бета-разнообразия сообществ, а также определить, какие признаки, кроме обычно рассматриваемой редкости – частоты, являются общими для таких видов.
Материалы
Работа основана на результатах анализа данных о структуре сообществ сухих и мезофитных лугов среднетаежной части Республики Карелия. Республика Карелия расположена на северо-западе Российской Федерации в таежной зоне, преобладающим биотопом здесь являются хвойные леса, занимающие 54.71 % территории, также обильно представлены болота и внутренние водоемы. Луга на 2022 г. занимали всего 0.39 % площади республики (Государственный доклад…, 2022). В среднетаежной части республики (граница между среднетаежной и северотаежной Карелией проходит примерно по 63 градусу северной широты) относительная площадь лугов несколько выше из-за исторически более развитого сельского хозяйства, но нигде не достигает значения выше 7 %. Фрагментация и изолированность отдельных луговых патчей в Карелии приводят к значительному островному эффекту. В то же самое время бореальные луга достаточно богаты видами: видовой фонд сосудистых растений отдельных лугов составляет от 30–40 до 120 видов травянистых растений. Все это делает северные луга достаточно удобным модельным объектом для исследований биоразнообразия.
Эколого-топологическая классификация сухих и мезофитных лугов среднетаежной Карелии, выполненная по описаниям со 102 луговых выделов, разделяет растительность на четыре ассоциации, следующие за почвенными факторами, прежде всего богатством почв азотом, от ассоциации сухих скальных лугов до нитрофильных группировок, формирующихся на заброшенных полях, пастбищах и сенокосах (Знаменский, 2015).
Полевые данные были получены в 2004–2016 гг. на 146 сайтах (здесь и далее этим термином обозначен конкретно локализованный выдел однородной растительности) сухих и мезофитных лугов среднетаежной части Республики Карелия (Российская Федерация). Сайты располагаются по всей территории юга республики, от Сортавалы на западе до Водлозерского национального парка на востоке (рис. 1).
Рис. 1. Карта-схема мест выполнения описаний
Fig. 1. Map-scheme of places where descriptions were performed
Методы
Растительность каждого сайта была описана при помощи 15 более или менее равномерно расположенных в границах однородного пятна луговой растительности пробных площадей размером 1 м2. Также для каждого выдела в пределах этого же пятна был составлен полный видовой список встреченных сосудистых растений. Виды в каждом списке были ранжированы по 4-балльной шкале: 4 – вид встречается по всей площади пятна, включая виды-доминанты; 3 – вид встречается спорадически по всей площади пятна, образуя локальные скопления; 2 – вид образует одно или два локальных скопления или распространен спорадически отдельными экземплярами; 1 – вид представлен единичными особями. Эти данные были сведены в рабочую таблицу.
Для каждого сайта по описаниям растительности были рассчитаны показатели альфа- и бета-разнообразия. В качестве модельных показателей мы использовали видовую насыщенность, т. е. среднее для 15 описаний число видов на 1 м2 (альфа) и среднее расстояние Сёренсена между 15 описаниями (бета). Для определения среднего расстояния Сёренсена в статистическом пакете Past 4.12 (Hammer et al., 2001) рассчитывалась матрица коэффициентов сходства Сёренсена dij (в меню Past – «коэффициент Брея – Кёртиса»), которая затем в MS Excel трансформировалась в матрицу расстояний (1 - dij), после чего рассчитывалась средняя величина для 105 расстояний между всеми парами среди 15 описаний. Все вместе 146 средних индексов для альфа- и бета-разнообразия составляли эмпирические векторы, которые фигурировали в дальнейшем анализе.
На первом этапе обработки использовали методику, похожую на подход, применяемый Ленноном и его последователями. Виды в сводной таблице также были проранжированы от самого распространенного до наиболее редких по суммарному обилию каждого вида по 4-балльной шкале на всех 146 сайтах, после чего, добавляя строку за строкой в таблице, мы отслеживали, как меняется связь вектора встречаемости отобранных видов с итоговыми показателями биоразнообразия. Основная разница в методике сводится к двум моментам.
1) В качестве индекса сходства мы использовали не коэффициент корреляции r, а коэффициент детерминации R2. Этот показатель позволяет оценить непосредственно вклад видов в дисперсию определенного индекса биоразнообразия.
2) В качестве показателей разнообразия мы брали не просто объем полного видового списка, а индексы, полученные при помощи геоботанических описаний. Таким образом, величина связи во время сканирования не просто изменялась от близкого к нулю значения до единицы, а достигала некоего максимума, после чего начинала падать. Особый интерес в этом и последующем экспериментах представляло именно максимальное значение коэффициента детерминации, ибо оно обозначало, что прибавление следующих видов только ослабляет связь.
На втором этапе мы отошли от градиента «обычный – редкий», проранжировав все виды, встреченные на луговых выделах именно по их вкладу в формирование итогового индекса биоразнообразия. Для этого был написан макрос Microsoft Excel, выполняющий следующие процедуры.
В начале работы ранжируемые виды в таблице перемешиваются случайным образом. Перебирая один вид за другим, мы подбирали в качестве опорного тот, у которого вектор, характеризующий наличие / отсутствие вида, показывал наиболее высокое значение R2 с вектором индекса биоразнообразия. Этот вид закреплялся вверху таблицы. На второй итерации программа перебирает один за другим все оставшиеся виды, при этом модельный вектор включает данные по наличию уже двух видов, и второй вид подбирается так, чтобы вместе с первым он давал модельный вектор, максимально схожий с вектором индекса разнообразия. Соответствующий вид закреплялся на второй строке таблицы, после чего мы переходили к третьей итерации и т. д. Итерации продолжаются до тех пор, пока вся таблица не будет упорядочена.
Экран расчета R2 показан на рис. 2. Расчеты были повторены в двух вариантах. В первый раз мы использовали исключительно данные по присутствию – отсутствию видов, во второй учитывалось их обилие по 4-балльной шкале.

Рис. 2. Экран расчета R2 в таблице Microsoft Excel. В данном примере показан расчет по таблице, включающей 10 видов, произрастающих на 7 сайтах. Сверху приведены два вектора, для которых определяется R2. Vector 1 – вектор индекса биоразнообразия (в данном примере – видовая насыщенность). Vector 2 – модельный вектор, рассчитываемый исходя из флористической таблицы. Формула в ячейках данного вектора подсчитывает число видов [=COUNNTIF(B6:Bx;”>0”)] или сумму [=SUM(B6:Bx)] обилий видов в х верхних строчках. В столбце ‘rank’ (столбец I) приведены ранги видов по сумме обилий. В ячейках столбца J содержится формула расчета R2 векторов 1 и 2. В столбце K записывается точное значение R2 на каждой итерации. Максимальное значение находится при сортировке таблицы по этому столбцу. Столбик L содержит случайное число [=RAND()] для смешивания видов в таблице в начале обработки. В показанный момент х = 3. Два верхних вида с максимальной R2 уже подобраны и зафиксированы вверху таблицы, ведется перебор оставшихся видов, причем будет подобран тот, который покажет наибольшее значение R2, после чего он будет зафиксирован на третьей строчке, и начнется подбор четвертого вида
Fig. 2. Screenshot of R2 calculation in Microsoft Excel worksheet. This example shows calculations in a table with 10 species growing in 7 sites. Two vectors for which R2 is determined are given at the top. Vector 1 is the vector of the biodiversity index (in this case, species density). Vector 2 is the model vector calculated from the flora worksheet. The formula in this vector’s cells calculates the number of species [=COUNNTIF(B6:Bx;”>0”)] or the sum [=SUM(B6:Bx)] of species abundances in the top х lines. The ‘rank’ column (column I) gives the ranks of species according to the sum of abundances. Cells in column J contain the formula for calculating R2 for vectors 1 and 2. The exact value of R2 at each iteration is recorded in column K. The maximum value is found by sorting the table according to this column. Column L contains the random number [=RAND()] for randomizing species in the table at the first step of processing. In the captured instant, х = 3. Two species with the highest R2 have already been selected and fixed at the top of the table; the remaining species are being scanned to select the one with the highest R2 to be fixed in the third line, after which scanning for the fourth species will begin
Для определения направления векторов биоразнообразия в рассмотренной растительности была применена ординация методом неметрического многомерного шкалирования (NMS) (Kruscal, Wish, 1978). Для ординации использовалась сводная таблица описаний растительности по 146 выделам, где учитывалась относительная встречаемость каждого вида в 15 описаниях на соответствующем выделе. NMS выполнена в пакете PC-ORD 7.08 (McCune, Mefford, 2018), при этом для расчета матрицы расстояний использовано также расстояние Сёренсена.
Результаты
В итоговой таблице фигурирует 292 вида травянистых растений и кустарников, встреченных нами на 146 луговых выделах. Ранг обычности видов варьирует от 1 (Achillea millefolium, найденный на 143 из 146 сайтов с суммарным обилием 469) до 255 (всего 38 видов, встреченных на одном выделе с обилием, равным единице). Если разделить этот набор по биотопическому признаку, то самую большую долю (37.67 %) в нем займут луговые виды, 26.37 % видов относятся к антропогенным биотопам, 15.07 % – к видам увлажненных местообитаний, 13.36 % – к видам лесов и 7.53 % – к видам скальных биотопов.
График изменения величины R2 при сканировании таблицы, упорядоченной по градиенту «обычный – редкий», показан на рис. 3.
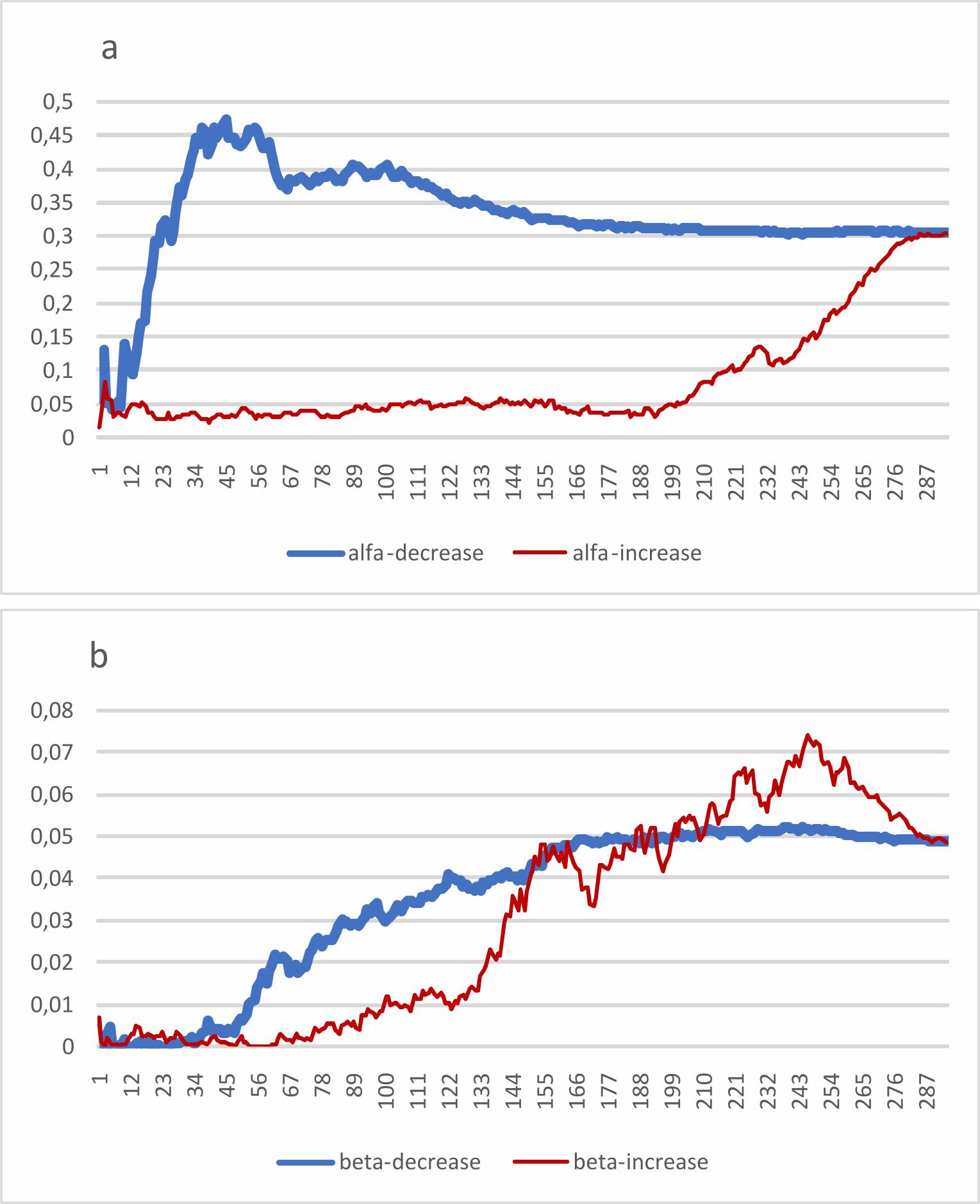
Рис. 3. Кривая изменения коэффициента детерминации по градиенту «частые – редкие»: a – для альфа-разнообразия (видовая насыщенность), b – для бета-разнообразия (дистанция Сёренсена). Толстой линией показана кривая для сканирования таблицы от наиболее частого вида к наиболее редким, тонкой – для сканирования от наиболее редких видов к наиболее частому
Fig. 3. The curve of change in the coefficient of determination along the gradient “frequent-to-rare” : a – for alpha diversity (species density), b – for beta diversity (mean Sørensen distance). The thick line shows the curve for scanning the table from the most common species to the most rare, the thin line shows the curve for scanning from the rarest species to the most frequent.
Кривая коэффициента детерминации альфа-разнообразия (рис. 3а) напоминает кривые, полученные в публикациях Леннона с соавторами (Lennon et al., 2004). Поскольку в нашем случае мы сравниваем не просто часть списка видов с полным списком, а берем независимые показатели биоразнообразия, при пополнении набора корреляция у нас не стремится к единице. Максимальное значение R2 составляет 0.4743, приходясь на уровень 45 наиболее обычных видов из 294. Дальнейшее добавление видов приводит только к снижению детерминации, и полному набору видов соответствует величина R2 = 0.3054. При этом самые верхние виды таблицы (т. е. самые частые) практически не изменяли величины детерминации, а сам прирост был крайне неравномерен: соседние по частоте виды могли давать и положительную, и отрицательную прибавку к кумулятивной величине коэффициента детерминации.
Кривая коэффициента детерминации бета-разнообразия (рис. 3b) выглядит беспорядочно и проходит через низкие значения при сканировании в обоих направлениях (максимальное значение R2 не превышает 0.08). Впрочем, некоторое повышение коэффициента детерминации при переходе от частых видов к редким наблюдается.
При сортировке видов в таблице при помощи макроса MS Excel кривая существенно сглаживается, а максимальное значение R2 заметно увеличивается. В итоге кривая имеет выраженную горбовидную форму. При этом кривые для альфа- и бета-разнообразия выглядят по-разному.
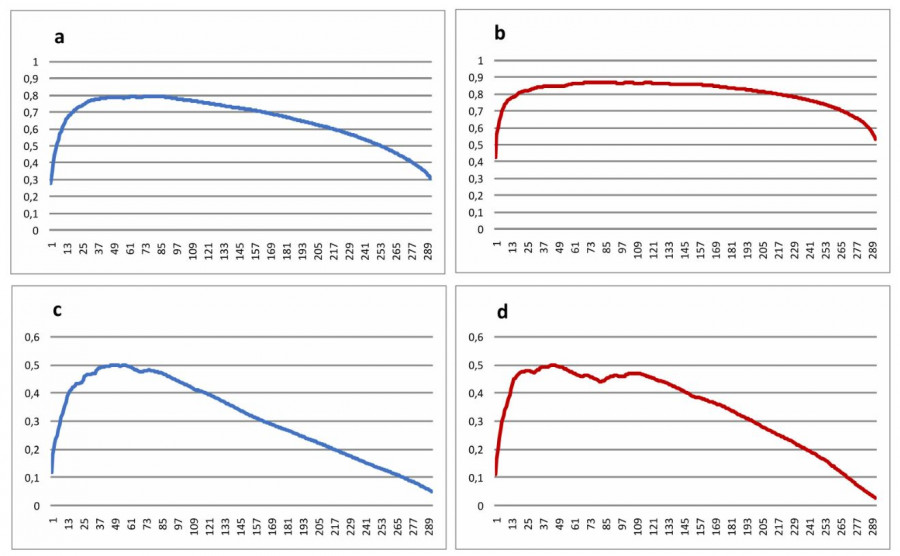
Рис. 4. Сглаженные кривые R2 для числа видов на 1 м2 (α-разнообразие) и среднего расстояния Сёренсена (β-разнообразие): a – альфа-разнообразие (видовая насыщенность) с учетом присутствия / отсутствия видов, b – альфа-разнообразие с учетом обилия видов, c – бета-разнообразие (средняя дистанция Сёренсена) с учетом присутствия / отсутствия видов, d – бета-разнообразие с учетом обилия видов
Fig. 4. Smoothed R2 curves for the number of species per 1 m2 (α-diversity) and mean Sørensen distance (β-diversity): a – alpha diversity (species density) taking into account the species presence-absence, b – alpha diversity accounting for species abundance, c – beta diversity (mean Sørensen distance) taking into account the species presence-absence, d – beta diversity taking into account the species abundance
На кривой для альфа-разнообразия (на рис. 4a, b показана кривая для видовой насыщенности) все виды можно разделить на три группы: «позитивные» виды, введение которых положительно влияет на итоговую величину R2; «нейтральные» виды, которые несущественно изменяют R2; «негативные» виды, которые уменьшают итоговое значение коэффициента детерминации. «Позитивные» виды достаточно четко отделяются от «нейтральных» по точке перегиба кривой, «негативные» виды не имеют четкой границы с «нейтральными», плавно переходя одни в другие.
Кривая детерминации для видов без учета обилия имеет максимальное значение на уровне 0.7945, что соответствует 80 видам (27.40 % от общего числа видов) в таблице со средним рангом 164.52 (медиана ранга 198). Точка перегиба соответствует уровню R2 = 0.7387; 24 (8.56 %) вида со средним рангом 119.41 (медиана 137). Эти 24 вида: Luzula multiflora, Potentilla erecta, Lathyrus pratensis, Cerastium fontanum, Rubus saxatilis, Viola epipsila, Pilosella caespitosa, Euphrasia officinalis, Ranunculus auricomus, Hieracium umbellatum, Trifolium spadiceum, Crepis biennis, Veronica scutellata, Calamagrostis arundinacea, Dactylorhyza maculata, Anthriscus sylvestris, Pilosella lactucella, Crepis paludosa, Stachys palustris, Convolvulus arvensis, Rubus idaeus, Trifolium sativum, Phleum pratense, Carex panicea. 10 видов из 24 являются индикаторными видами отдельных луговых ассоциаций, причем среди них представлены индикаторные виды всех четырех ассоциаций луговой растительности региона (Знаменский, 2015), еще два вида являются доминантными луговыми видами региона.
Использование данных с учетом обилия отдельных видов сохраняет общий вид кривой, но она сама претерпевает некоторые изменения. Максимальная величина R2 значительно возрастает и теперь составляет 0.8655, что соответствует 76 видам из 294 (26.03 %), со средним рангом 168.45 (медиана 191). Сама кривая значительно уплощается, т. е. больше видов переходит в разряд «нейтральных». Перегиб кривой происходит несколько раньше и соответствует 21 первым видам (7.19 % видов) с уровнем R2 = 0.8103 и средним рангом 120.19 (медиана 133): Anthoxanthum odoratum, Achillea millefolium, Pilosella caespitosa, Vicia sepium, Viola montana, Trifolium pratense, Rubus saxatilis, Viola epipsila, Hierocloë hirta, Cirsium vulgare, Campanula latifolia, Bistorta vivipara, Convallaria majalis, Thalictrum simplex, Millium effusum, Rumex thyrsiflorus, Rumex acetosa, Luzula pilosa, Campanula persicifolia, Allium schoenoprasum, Polemonium caeruleum. Тут увеличивают свое представительство наиболее обычные луговые виды, хотя увеличения среднего ранга при этом не происходит.
Кривая для бета-разнообразия (рис. 4c, d) также имеет горбовидную форму, но заметно отличается от кривой альфа-разнообразия. Так, на кривой, построенной только по данным о присутствии / отсутствии вида, практически отсутствует группа «нейтральных» видов: кривая, поднявшись вверх, тут же начинает опускаться. Точкой перегиба является максимум, соответствующий R2 = 0.4977, что приходится на 57 видов растений (19.52 % от общего числа) со средним рангом 189.1 (медиана 198). В списке данных видов преобладают виды других типов растительных сообществ: на 28 % это виды антропогенно нарушенных местообитаний (Sagina procumbens, Poa annua, Thlaspi arvense etc). 26 % списка составляют виды ветландов (Rorippa palustris, Lysimachia thyrsiflora, Carex juncella etc), 12 % – лесные виды (Lycopodium clavatum, Delphinium elatum, Viola epipsila etc.) и 10 % – виды скальных местообитаний (Polypodium vulgare, Scletanthus annuus, Sedum telephium etc.) Луговые виды составляют менее 22 %, в основном редкие в регионе (Avenula pubescens, Poa angustifolia, Carex muricata etc), однако имеются и обычные виды (Alchemilla vulgaris, ранг 5; Dactylis glomerata, ранг 4).
При учете в компьютерной обработке обилия отдельных видов результат несколько изменяется. На кривой теперь просматривается небольшая группа «нейтральных» видов, примерно с 21-го по 108-й вид. Максимальное R2 = 0.4965, и соответствует 45 видам (15.41 %) со средним рангом 196.79 (медиана также 198). 36 из них те же, что в предыдущем случае, 9 видов в предыдущий список не входят. Биотопическая структура также в значительной степени сохраняется: виды антропогенных местообитаний составляют 33 %, виды ветландов – 27 %, скальные виды – 13 % и лесные – 11 %. Луговые виды составляют 16 % списка «позитивных» видов, обычных луговых видов среди них теперь нет.
Ординация NMS вычленяет в рассматриваемой растительности два главных градиента, первая ординационная ось берет на себя 57.1 % общей изменчивости, вторая – 30.6 %. При этом показатель альфа-разнообразия значимо коррелирует с первой осью NMS (r = 0.749, t = 0.494), а показатель бета-разнообразия не коррелирует ни с первой осью, ни со второй (рис. 5).
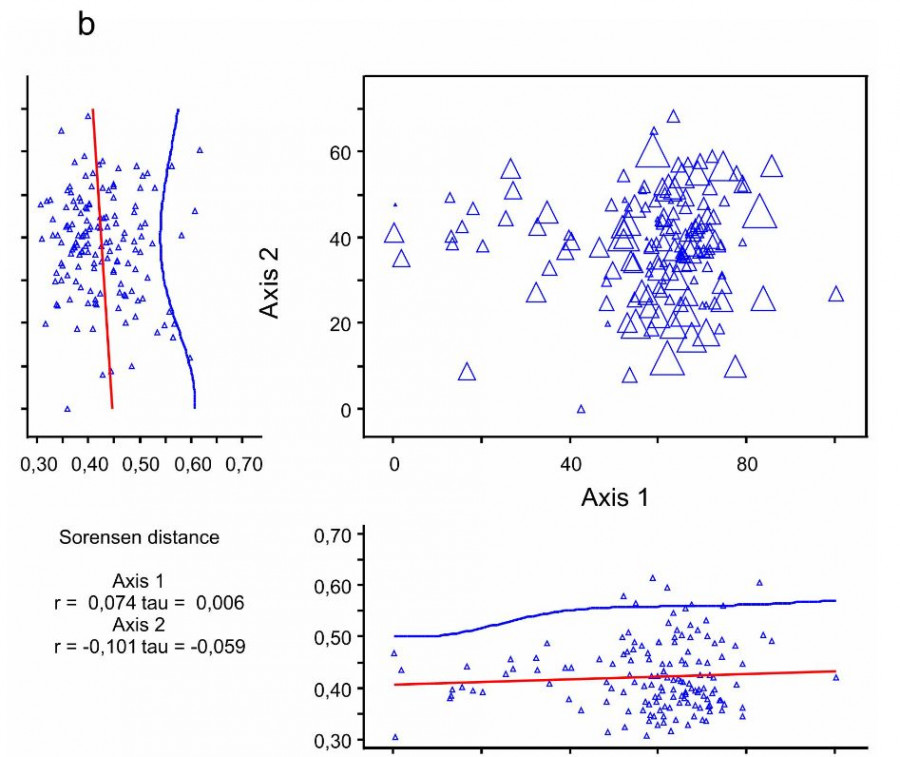
Рис. 5. Индексы альфа- и бета-разнообразия в ординационном пространстве NMS: a – видовая насыщенность, b – среднее расстояние Сёренсена. Размер значка на диаграмме соответствует величине индекса биоразнообразия
Fig. 5. Alpha- and beta-diversity indices in the NMS ordination space: a – species density, b – mean Sørensen distance. Symbol size in the diagram corresponds to the biodiversity index value
Обсуждение
Полученные результаты подтверждают вывод о том, что в величину биоразнообразия, в частности альфа-разнообразия, основной вклад вносит ограниченное число относительно частых видов. В дальнейшем, однако, оказывается, что ситуация несколько сложнее и выходит за рамки градиента «обычные виды – редкие».
Замена общего числа видов в наборе на другой показатель альфа-разнообразия мало что изменит в качественном плане, поскольку различные индексы альфа-разнообразия демонстрируют сильную связь между собой. Максимальные значения коэффициента R2 для альфа-разнообразия оказались неожиданно высокими. Они свидетельствуют, что итоговая величина альфа-разнообразия на 4/5 зависит от наличия 27 % встреченных на лугах региона видов и на 6/7 – от обилия немного меньшего числа видов. При этом на 3/4 она зависит от наличия менее 10 % видов и на 4/5 от обилия примерно такого же числа видов. Необъясненная часть дисперсии становится весьма малой, всего 15–20 %.
В приведенном нами случае альфа-разнообразие на лугах изменяется направленно, вдоль главного градиента растительности, соответствующего изменению уровня нутриентов, в частности почвенного азота. Обратная линейная зависимость альфа-разнообразия и почвенных нутриентов хорошо известна в науке о растительности (Tilman, 1984). В нашем случае вдоль этого же градиента выделены четыре различных ассоциации луговой растительности. Это объясняет, почему среди «позитивных» видов обильно представлены индикаторные виды соответствующих ассоциаций, изменяющие свое обилие вдоль главного градиента.
Привлечение во время обработки величин обилия отдельных видов должно было способствовать также появлению среди «позитивных» видов доминантов, которые присутствуют на всех или почти всех выделах, а на изменение уровня нутриентов реагируют своим обилием. Отчасти этот эффект наблюдался при учете обилия видов, среди «позитивных» появились Achillea millefolium (ранг 1), Anthoxanthum odoratum (9), Trifolium pratense (12) и Rumex acetosa (9). Однако при этом медианный ранг «позитивных» видов изменился несущественно (133 против 137), а средний ранг даже несущественно вырос.
В то же самое время среди «положительных» видов, увеличивающих связь между видовым набором и итоговым биоразнообразием, встречаются и редкие виды с весьма низким рангом, например Stachys palustris, Convolvulus arvensis (оба имеют ранг 232) или Carex panicea (ранг 255).
В формировании бета-разнообразия гораздо большую роль играют стохастические процессы. Во-первых, заметно меньше максимальное значение R2, которое на практике не достигает 0.5, во-вторых, средний ранг видов с позитивной реакцией на R2 значительно меньше, т. е. преобладают более редкие виды. Но и тут можно заметить некоторую закономерность. «Позитивные» для бета-разнообразия виды, как правило, появляются на чужеродных вкраплениях в луговых сообществах: местах с повышенным вытаптыванием и повышенным содержанием почвенного азота, например тропинках и свалках мусора (антропогенные виды), канавах и временных водоемах в микродепрессиях (виды ветландов), на кучах камней и скальных обнажениях (скальные виды). Не имеют своих типов инфрасообществ лесные виды, наличие которых в основном зависит от размеров лугового сайта и окружающих его биотопов. Надо отметить, что площадки, на которых описывалась растительность, размещались только на непосредственно травяной растительности, поэтому эти «позитивные» виды на растительность никак не влияли, но учитывались в парциальной флоре. Преобладание среди «позитивных» для бета-разнообразия антропогенных видов (однолетников, связанных с вытаптыванием, нитрофильных сорняков и адвентивных видов, сбежавших из культуры) наводит на мысль о том, что основным механизмом увеличения бета-разнообразия на описываемых лугах является антропогенный пресс. Немного менее значима мозаичность, создаваемая микрорельефом, в частности наличием депрессий, куч камней и скальных обнажений. При этом зачастую депрессии, например лужи на дорогах и кучи камней, также нередко имеют антропогенное происхождение.
Среди «положительных» видов, чье присутствие усиливает связь с итоговым бета-разнообразием, есть два обычных вида с высоким рангом: Alchemilla vulgaris L. s. l. (ранг 5) и Dactylis glomerata L. (ранг 4). И тот, и другой вид имеет свои выраженные экологические специфики и может формировать пятна доминирования на лугах, а значит, увеличивать мозаичность и через него – величину бета-разнообразия.
Привлечение данных по обилию видов практически не изменяет максимальную величину R2, поскольку среди «позитивных» видов преобладают виды редкие, имеющие стабильно низкое обилие, а значительное направленное изменение обилия можно заметить только у обычных видов.
Примененный метод сортировки видов по вкладу в корреляцию, очевидно, может использоваться не только для исследования биоразнообразия, но и для оценки связи наборов видов с любыми градиентами, например географическими или экологическими.
Стратификация видов по их вкладу в формирование разнообразия, очевидно, является необходимым условием при моделировании разнообразия сообществ.
Заключение
Проведено упорядочивание списков видов в сообществах по их вкладу в итоговые индексы биоразнообразия. Оно показало, что формирование индексов, на самом деле, весьма хорошо описывается простой линейной зависимостью, особенно в том случае, когда в сообществах имеется выраженный градиент среды, которому подчиняется индекс биоразнообразия. Простой видовой список объясняет до 80 % дисперсии альфа-разнообразия, а с учетом обилия – до 86 %. Бета-разнообразие, для которого отсутствует такой градиент, имеет существенно более низкую долю дисперсии, объясняемой наличием или обилием конкретных видов.
Сами виды могут быть условно разделены на три группы: 1) достаточно четко отделяемые по точке перегиба или по максимуму достигаемой детерминации сравнительно распространенные «положительные виды»; 2) составляющие заведомое меньшинство распространенные «нейтральные» виды, чье присутствие никак не влияет на итоговое разнообразие; 3) большую группу шумовых видов, встречающихся редко и оказывающих негативный эффект на показатель разнообразия.
Библиография
Государственный доклад о состоянии окружающей среды Республики Карелия в 2021 г. / Министерство природных ресурсов и экологии Республики Карелия; Ред. кол.: А. Н. Громцев (гл. ред.), О. Л. Кузнецов, А. Е. Курило, Е. Г. Полина. Петрозаводск, 2022. 263 с.
Знаменский С. Р. Растительность ксеромезофитных и мезофитных лугов среднетаежной Карелии: эколого-топологический подход // Труды Карельского научного центра Российской академии наук. 2015. № 2. С. 3–15. DOI: 10.17076/eco40
Hammer Ø., Harper D., Ryan P. PAST: Paleontological Statistics Software Package for Education and Data Analysis // Palaeontologia Electronica. Vol. 4, No 1. P. 1–9.
Heegaard E., Gjerde I., Saetersdal M. Contribution of rare and common species to richness patterns at local scales // Ecography. 2013. Vol. 36. P. 937–946. DOI: 10.1111/j.1600-0587.2013.00060.x
Kruskal J. B., Wish M. Multidimensional Scaling. Sage Publications, Beverly Hills, California, 1978. 93 p.
Lennon J. J., Koleff P., Greenwood J. J. D., Gaston K. J. Contribution of rarity and commonness to patterns of species richness // Ecology Letters. 2004. Vol. 7. P. 81–87. DOI: 10.1111/j.1600-0587.2010.06669.x
Lennon J. J., Beale C. M., Reid C., Kent M., Pakeman R. Are Richness Patterns of Common and Rare Species Equally Well Explained by Environmental Variables? // Ecography. 2011. Vol. 34. P. 529–539. DOI: 10.1111/j.1600-0587.2010.06669.x
Mazaris A. D., Tzanopoulos J., Kallimanis A., Matsinos Y., Sgardelis S., Pantis J. The contribution of common and rare species to plant species richness patterns: the effect of habitat type and size of sampling unit // Biodiversity and Conservation. 2008. Vol. 17. P. 3567–3577. DOI: 10.1007/s10531-008-9480-7
McCune B. and Mefford M. J. PC-ORD. Multivariate Analysis of Ecological Data. Version 7.08. Wild Blueberry Media. Corvallis, Oregon, U.S.A., 2018.
Sizling A., Sizlingová E., Storch D., Reif J., Gaston K. J. Rarity, commonness, and the contribution of individual species to species richness patterns // American Naturalist. 2009. Vol. 174. P. 82–93. DOI: 10.1086/599305
Tilman D. Plant dominance along an experimental nutrient gradient // Ecology. 2009. Vol. 65. P. 1445–1453.
Vázquez L., Gaston K. J. Rarity, Commonness, and Patterns of Species Richness: The Mammals of Mexico // Global Ecology and Biogeography. 2004. Vol. 13. P. 535–542.
Благодарности
Автор выражает благодарность за составление карты М. С. Потахину (Институт водных проблем Севера Карельского научного центра РАН), а также двум анонимным рецензентам, чьи замечания помогли существенно улучшить рукопись. Работа выполнена в рамках государственной темы НИР номер FMEN-2022-0008.

 © 2011 - 2026
© 2011 - 2026